
Cytokinin-Controlled Gradient Distribution of Auxin in Arabidopsis Root Tip



Institute of Cell Biology and MOE Key Laboratory of Cell Activities and Stress Adaptations, School of Life Sciences, Lanzhou University, Lanzhou 730000, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(8), 3874; https://doi.org/10.3390/ijms22083874
Submission received: 19 March 2021
/
Revised: 4 April 2021
/
Accepted: 6 April 2021
/
Published: 8 April 2021
(This article belongs to the Special Issue Signal Conversion and Auxin Transport in Plant Tropism)
{kind=link}
{kind=link}
Abstract
:The plant root is a dynamic system, which is able to respond promptly to external environmental stimuli by constantly adjusting its growth and development. A key component regulating this growth and development is the finely tuned cross-talk between the auxin and cytokinin phytohormones. The gradient distribution of auxin is not only important for the growth and development of roots, but also for root growth in various response. Recent studies have shed light on the molecular mechanisms of cytokinin-mediated regulation of local auxin biosynthesis/metabolism and redistribution in establishing active auxin gradients, resulting in cell division and differentiation in primary root tips. In this review, we focus our attention on the molecular mechanisms underlying the cytokinin-controlled auxin gradient in root tips.
1. Introduction
Roots are plant organs, usually located below the surface of the soil, where they grow and respond to various environmental stimuli. Plants receive essential water and nutrients from the soil through roots. Thus, overall plant survival depends on root growth and development. Auxin is the most important phytohormone that regulates the growth and development of plant roots [1,2,3,4,5]. Although auxin plays a central role in the control of root growth and development, much evidences have now been collected showing that the correct growth and development of roots also depends on auxin cross-talk with other phytohormones, such as cytokinin [6,7]. Since the discovery of cytokinin as an inducer of cell division in cultured plants, the function of cytokinin has been linked to that of auxin [8,9]. Earlier studies have suggested an antagonistic relationship between these two phytohormones; however, the truth is more complicated, with both antagonistic and supportive interactions that are usually cell and/or tissue-specific [10,11,12].
It is usually considered that the key to the function of auxin in plant growth and development is its gradient distribution [13,14]. Of all the mechanisms that regulate auxin distribution, cytokinin plays a prominent role, not only by regulating local auxin metabolism [15,16,17,18,19,20,21,22], but also by modulating PAT (polar auxin transport) [11,23,24,25,26,27]. In the last decade, multiple points of cross-talk between auxin and cytokinin, including biosynthesis/metabolism, transport, and signaling, have been revealed [12,23,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48]. In this review, we focus on cytokinin-controlled gradient distribution of auxin by regulating its biosynthesis and transport, and its role in regulating root growth and development.
2. Cytokinin Signal Pathway
The cytokinin signaling pathway in plants is similar to the bacterial multi-step two-component signal transduction system [12,34,36,49]. In Arabidopsis, cytokinin binding leads to autophosphorylation of membrane-bound cytokinin receptors AHK2 (Arabidopsis histidine kinase 2), AHK3 and AHK4/CRE1 (cytokinin response 1), followed by a phosphorylation cascade [50,51,52,53].The phosphoryl group is transfered from receptors to AHPs (Arabidopsis histidine phosphotransferase proteins) [54,55,56,57], which enters the nucleus and phosphorylates the ARRs (Arabidopsis response regulators). ARRs can be divided into two types according to their structure. Phosphorylated type-B ARRs work as TFs (transcription factors), activating cytokinin-responsive genes [58,59,60,61]. Unlike the type-B ARRs, the type-A ARRs lack a DNA-binding domain, and their expression is rapidly induced by cytokinin, which forms a feedback loop by negatively regulating type-B ARRs [62,63,64,65]. Furthermore, some CRFs (cytokinin responsive factors), identified as AP2 TFs [66,67], also play a role in cytokinin-regulated gene expression [67].
3. Cytokinin-Regulated IAA Biosynthesis
Based on biochemical and genetic evidences, the major natural auxin in plants, IAA (indole-3-acetic acid), is synthesized via two major pathways: Trp (Tryptophan)-independent (TI) and Trp-dependent (TD) pathways [5,22,68]. So far, the molecular components of the TI pathway have been poorly understood [69].
At present, it appears that the best understood IPA (indole pyruvic acid) pathway is the main TD pathway of auxin biosynthesis in Arabidopsis thaliana [48,70,71], in which TAA (tryptophan aminotransferase of Arabidopsis) family proteins catalyze the conversion of Trp to IPA [16,72,73,74], and YUC (YUCCA) flavin monooxygenase-like proteins catalyze the conversion of IPA to IAA [70,71,75]. Overexpression of YUCs, but not TAA family genes, leads to auxin overproduction, implying that the YUCs, rather than TAA family proteins, catalyze the rate-limiting step of the IPA pathway [70,76,77,78].
Besides TAA1/WEI8/SAV3/TIR2/CKRC1 (weak ethylene insensitive 8/shade avoidance 3/transport inhibitor response 2/cytokinin induced root curling 1) [16,72,73,74], the TAA family also includes two other homologous proteins: TAR1 (tryptophan aminotransferase related 1) and TAR2, which have overlapping functions [72]. YUCs belongs to a large gene family with 11 members in the Arabidopsis genome, which are functionally redundant to each other [75,76,77,78,79,80,81]. The importance of the IPA pathway in plant growth and development has made it the focus of research in recent years, and it is regulated by various developmental and environmental signals, including phytohormones [22,48,82].
Early definite evidence of the effect of cytokinin on auxin biosynthesis was reported by Jones et al. (2010), showing that cytokinin can induce auxin biosynthesis and that some signaling components of auxin and cytokinin are involved in this process; however, the molecular mechanism remains unclear [15]. Later, the authors found that cytokinin can induce the expression of both TAA1 and YUC8 genes to enhance auxin production [16,17]. In the adventitious root apex of Arabidopsis, cytokinin-mediated up-regulation of YUC6 was found to be involved in the formation of the QC (Quiescent Center) [83]. Cytokinin-induced expression of YUC1 and YUC4 in the gynoecia primordium has been reported to ensure correct domain patterning [39].
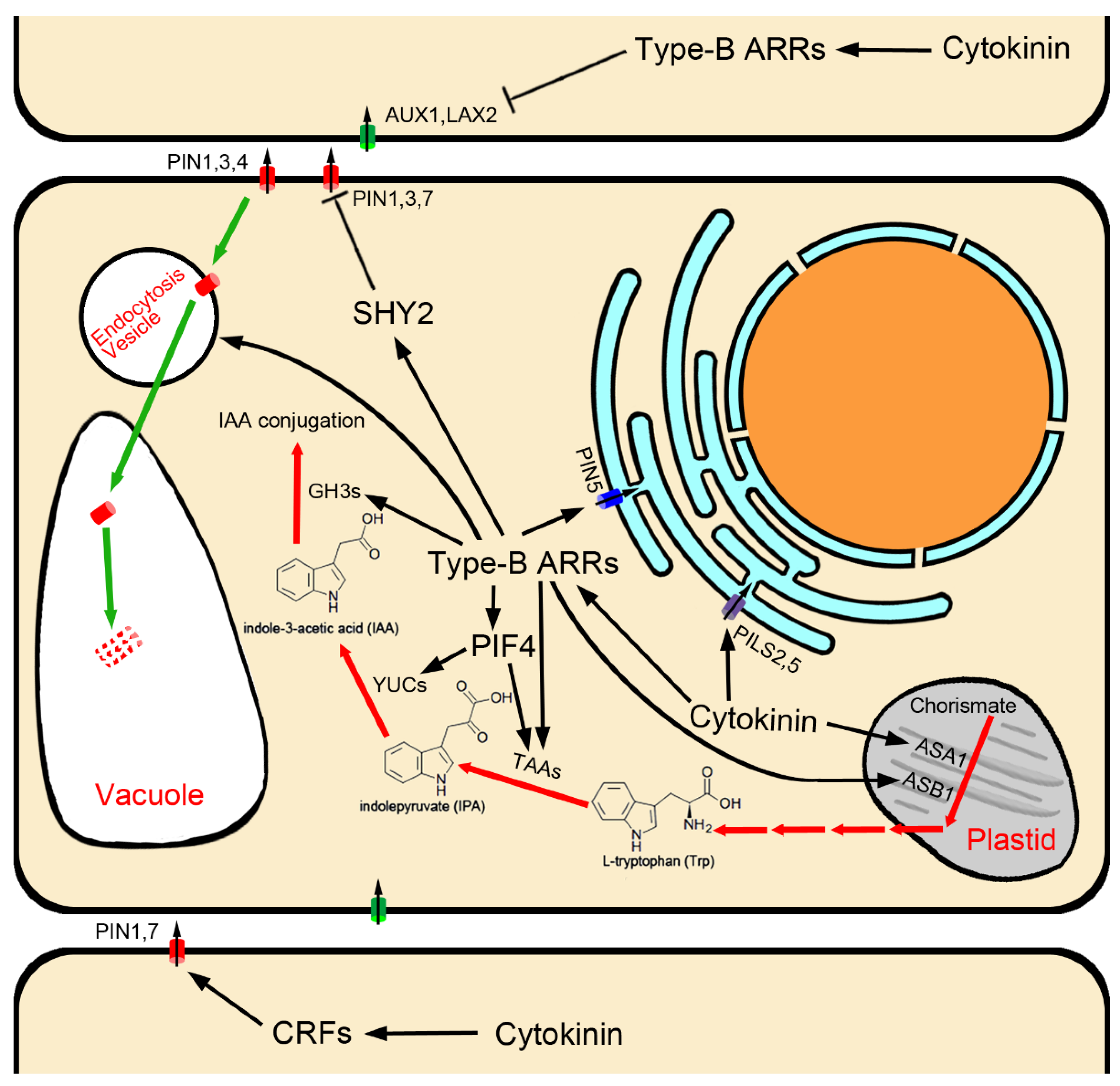
Transcriptional activation of TAA and YUC family genes by cytokinin is dependent on cytokinin signaling transduction, as has been shown for TAA1/CKRC1 [16] and YUC8/CKRC2 [17] in Arabidopsis (Figure 1). Yan et al. (2017) found that type-B ARRs can directly bind two cis elements in the promoter and the second intron of TAA1 to activate its transcription [18]. The AHKs-ARR1/12-mediated cytokinin signaling pathway is also reported to be necessary for cytokinin-induced up-regulation of TAA1 and YUC8, and the PIF4 (phytochrome-interacting factors 4) is required for this upregulation. Transcription of PIF4 itself is induced by cytokinin via the AHKs-ARR1/12 signaling pathway, indicating that PIF4 plays an essential role in mediating the regulatory effect of cytokinin on the transcriptions of TAA1 and YUC8 genes in the IPA pathway of auxin biosynthesis [17].
Upstream of the IPA pathway, the two subunits of rate-limiting anthranilate synthase, ASA1 (anthranilate synthase alpha-subunit 1)/WEI2/CKRC6 and ASB1 (anthranilate synthase beta-subunit 1)/WEI7 in the Trp biosynthesis pathway [72,84], are specifically expressed in the root tip [72,73,79,85]. It has been shown that the transcriptional levels of ASA1/WEI2/CKRC6 and ASB1/WEI7 can be induced by cytokinin [15,85,86], and ARR1 promotes auxin biosynthesis in the stem cell niche via the up-regulation of ASB1/WEI7 in the root meristem (Figure 1) [85].
In another proposed but still somewhat obscure TD pathway named the IAOx (indole-3-acetaldoxime) pathway, the key enzymes of cytochrome P450 proteins CYP79B2 and CYP79B3 convert Trp to IAOx. Overexpression lines of these two genes showed auxin overproduction phenotypes and a high level of auxin; on the contrary, plants showed auxin-deficient phenotype and decreased endogenous auxin content when the enzyme function was lost [87,88]. Expressed in the root meristem [87], CYP79B2 and CYP79B3 are involved in root elongation [87,88]. Microarray and qRT-PCR results show that their transcription is activated when the plant is treated with cytokinin [15,89], and the result of ChIP-seq (chromatin immunoprecipitation sequencing) shows that ARR1 can bind to a partial sequence of a CYP79B3 gene after 3 h cytokinin treatment [90]. It is likely that the IAOx pathway could be another cytokinin-regulated auxin biosynthesis pathway.
Most recently, it was found that exogenous cytokinin can stimulate the expression of CKRW2/HUB1 (cytokinin induced root waving 2/histone monoubiquitinate 1), which encodes an E3 ligase required for histone H2B mono-ubiquitination (H2Bub1) to promote the transcription of auxin biosynthetic genes TRP2/TSB1 (tryptophan biosynthesis 2/tryptophan synthase beta-subunit 1), ASB1/WEI7, YUC7 and AMI1 (amidase 1) [91]. This discovery reveals an epigenetic mechanism of cytokinin-regulated IAA biosynthesis at the chromatin level.
4. Cytokinin-Regulated IAA Conjugation and Degradation
In Arabidopsis, the GH3 (Gretchen Hagen 3) family belongs to a large gene family with three groups, of which group II (GH3.1-6, GH3.9 and GH3.17) has been shown to convert IAA to IAA amino acid conjugates [95,96,97]. The conjugation of amino acids with IAA is usually classified into two categories: one that can be converted to free IAA through hydrolysis and is considered to be related to the storage of auxin, such as IAA-Ala and IAA-Leu [98,99]; the other is believed to be related to the degradation of auxin, which can be irretrievably oxidized after formation and then degraded, such as IAA-Asp and IAA-Glu [100,101,102]. The process of IAA amino acid conjugation is generally considered to be associated with auxin homeostasis, which may play a role in cases where plant cells have to rapidly alter the relative amount of IAA in response to developmental and environmental changes [19,22,95,102,103,104,105,106].
The LRC (lateral root cap) is the outermost tissue of the root meristem [107]. If the LRC is lost, the size of the meristem will be greatly reduced [108,109]. This is because bPAT (basipetal PAT, from the root tip to the elongation zone) starts at the lateral root cap. If the LRC is lost, auxin transported from the root tip to the elongation zone will be disturbed. The defect of bPAT makes it impossible to establish a normal gradient distribution of auxin, so that the root meristem becomes smaller [21,110]. However, it is interesting to note that several members of group II GH3 (GH3.5, GH3.6 and GH3.17) are specifically expressed in the LRC [19,20,21], in which GH3.17 catalyzes IAA to IAA-Glu to participate in IAA degradation [19,21]. It has been reported that these three conjugation enzymes play a key role in controlling auxin flow in bPAT, as they determine the amount of auxin transport from the root tip to the elongation zone [19,20,21].
Surprisingly, GH3.5, GH3.6 and GH3.17 are downstream of type-B ARR1 in cytokinin signal transduction, and are targets of cytokinin–auxin antagonism [20,21]. Cytokinin suppresses bPAT by activating transcription of GH3.5, GH3.6 and GH3.17, which convert free IAA to IAA amino acid conjugates, thus regulating the size of the root meristem (Figure 1) [20,21].
5. Cytokinin-Regulated Intercellular Auxin Transport
The carriers that mediate auxin transport between cells contain three protein families: (1) AUX1/LAX (AUX1/LIKE AUX1) family proteins, responsible for the transport of auxin from the apoplast into the cell [111,112,113,114,115]; (2) PIN (PIN-formed) family proteins that mediate auxin output cells [116,117,118,119,120]; (3) ABCB/PGP/MDR (ATP-binding cassette protein subfamily B/P-Glyco protein/multidrug resistance) family proteins, involved in the ATP-driven influx or efflux of auxin [121,122].
Of these three families, only AUX1/LAX influx and PIN efflux carriers are involved in PAT machinery, directing the flow of auxin from the shoot acropetally through the stele toward the root tip (aPAT, acropetal PAT). From here it is basipetally redistributed via the epidermis to the elongation zone (bPAT) [115,116,120,123,124,125,126,127]. The pattern of expression of the various AUX1/LAX and PIN genes and the localization of them on specific cell faces play a key role in PAT machinery to determine the distribution of auxin in plant tissues [115,116,120,123,124,125,126,127]. Unlike AUX1/LAX influx and PIN efflux carriers, the ABCB/PGP/MDR family proteins have also been shown to act as auxin transporters to mediate auxin in and out of cells; however, because they are uniformly localized in the cell, they are considered to be unrelated to PAT [128,129].
In the last 10 years, studies on cytokinin-regulated plant development have revealed that a number of processes are involved in cytokinin interaction with PAT (e.g., root and shoot apical meristem activity maintenance, lateral root organogenesis, vasculature differentiation, or phyllotaxis [11,26,47,130,131]). In primary roots, previous studies suggested that cytokinin inhibition of cell expansion depended on cytokinin-induced ethylene biosynthesis [132]. The inhibition of root cell elongation requires ethylene regulated transport-dependent auxin distribution [27,133]. Although the role of ethylene in the cytokinin response has been demonstrated, the direct regulation of PAT by cytokinin is more important for root growth and development.
5.1. PINs Efflux Carriers
In Arabidopsis thaliana, according to the length of the hydrophilic loop in the middle of the polypeptide chain, the PINs family is divided into two subfamilies: as auxin efflux carriers, PIN1, PIN2, PIN3, PIN4 and PIN7 contain a long hydrophilic loop and are located in the PM (plasma membrane) [117,119,120], while PIN5, PIN6 and PIN8 with a short hydrophilic loop are mainly located in the ER (endoplasmic reticulum), which are involved in intracellular auxin transport [117,119,120,134]. All of the PIN efflux carriers are expressed and active in the root tip and perform their respective functions [116,117,119,120,129,135].
Cytokinin has been shown to influence cell-to-cell auxin transport by regulating the expression of several PIN genes, thereby modulating auxin distribution, which is essential for root development [11,26,131,136,137]. In Arabidopsis roots, through the cytokinin receptor AHK3 and the downstream signaling components ARR1 and ARR12, cytokinin has been shown to activate SHY2 (short hypocotyl 2), which is a member of the AUX/IAA (Auxin/Indole-3-Acetic Acid) protein family that heterodimerizes with ARFs (auxin response factors), preventing the activation of auxin responses. Therefore, as a downstream gene of AFRs, the expression of PIN1, PIN3 and PIN7 was inhibited when SHY2 was activated (Figure 1) [11,138].
Some CRFs also directly fine-tune PIN expression, providing a direct regulatory link between cytokinin signaling and the auxin transport machinery. Plants lacking CRF activity show developmental pattern aberrations consistent with abnormal auxin distribution. Removal of specific cis-regulatory elements (PCRE (PIN cytokinin response element) domain 5′-AGCAGAC-3′-like motif) effectively uncouples PIN1 and PIN7 transcription from the CRF-dependent regulation, and attenuates plant cytokinin sensitivity (Figure 1) [137]. Furthermore, the bHLH TF SPATULA enables cytokinin signaling, and activates the expression of PIN3 [139].
Besides transcriptional regulation, cytokinin also negatively regulates PINs at the post-transcriptional level [140]. Cytokinin can affect endomembrane trafficking of PIN1, PIN3 and PIN7 to redirect them for lytic degradation in vacuoles to reduce their abundance on the plasma membrane [141], and this function relies on canonical cytokinin signaling components, including the cytokinin receptor AHK4/CRE1 and some type-B ARRs [47]. PIN1 phosphorylation status is also involved (Figure 1) [142].
The complexity of cytokinin effects on PINs to regulate PAT in various cells/tissues/organs and developmental stages by various mechanisms has led to some confusing or even seemingly contradictory results. For example, in studying the effect of exogenous cytokinin on the transcription of PIN1, cytokinin was found to inhibit PIN1 transcription by using a 2-mm root tip with meristem/transition/elongation zones as the material [140], but was reported to have no such effect on a 0.5-mm root tip mainly with meristem zones [11,26,140]. In fact, cytokinin still causes PIN1 inactivation in the meristem, which depends on cytokinin-induced post-transcriptional regulation [140]. As another example, cytokinin down-regulates PIN1 and PIN3 proteins in primary roots to inhibit aPAT [11,26,136], but promotes the accumulation of PIN3, PIN4, and PIN7 in shoots, thereby coordinating bud outgrowth and branching [47].
5.2. AUX1/LAX Influx Carriers
In Arabidopsis thaliana, AUX1/LAX influx carriers are encoded by a small multigene family comprised of four members: AUX1, LAX1, LAX2, and LAX3 [111,112,113,114,115]. They display reasonably distinct expression patterns and are suggested to participate in different developmental processes [111]. Of all the members, except for LAX1, which is not involved in root development [111], the AUX1 gene is mainly expressed in the LRC, epidermal and phloem tissues near the root tip [113,143], and has been shown to play a role in gravitropism [143]; both AUX1 and LAX3 are shown to regulate lateral root development [112], and LAX2 is strongly expressed in the QC and the LRC [111], where it plays a key role in maintaining the stem cell fate surrounding the QC [144]. It was found that disruption of the LAX2 gene results in a phenotype similar to that observed in type-A ARR mutants, such as increased division of cells in the QC [144]. This is because auxin influx carriers, LAX2 genes, act downstream of cytokinin in the root tip, whose transcription is suppressed by cytokinin [27,144]. The decrease in AUX1 and LAX2 expression in response to cytokinin requires cytokinin response transcriptional effector type-B ARRs, which mediate the primary transcriptional response to cytokinin (Figure 1) [27,144,145]. CHIP assays showed that the AUX1 gene was enriched for extended type-B ARR12 binding motifs in intron 8 [27,145], and type-B ARR1 was found to bind directly to intron 2, intron 4 and 1.2 kb upstream motifs of the LAX2 gene [144]. These studies indicate that cytokinin response transcriptional effector type-B ARRs directly down-regulate the expression of AUX1/LAX influx carriers.
6. Cytokinin-Regulated Intracellular Auxin Transport
In addition to the above-mentioned PINs for intercellular PAT, the auxin carrier proteins for intracellular auxin transport include ER-localized PIN5, PIN6, PIN8, and other PILSs (PIN-like proteins), which are likely older than PINs by phylogenetic analysis [120,146,147,148]. There are seven known members of the PILS family. Although the PILS proteins share only 10–18% of their sequence with PIN proteins, they are topologically similar [147,148,149]. Members of the PILS family are identified by the presence of an auxin carrier domain that spans almost the entire length of the PILS proteins; therefore, PILS proteins still have the ability to transport auxin across the membrane [120,146].
Compared with the auxin efflux PINs located on the plasma membrane, which are involved in the intercellular transport of auxin, ER-localized PINs and PILSs mediate the intracellular transport of auxin [120,134,150,151,152,153,154]. ER-localized PINs are speculated to mediate auxin flow into (PIN5) or out (PIN8) of the ER lumen [120,152,154], or hypothetically from the ER lumen into the nucleus (PIN6 and PIN8) to open the auxin downstream genes’ transcription [150,152]. Like PIN5, the expression of PILS2 and PILS5 transporters causes cytosolic auxin to be transported into the ER lumen, leading to reduced transcriptional regulation of downstream genes by auxin in the nucleus, thus reducing auxin signals and cell sensitivity to auxin [120,146,147,148,155,156].
PIN5 is expressed in the vasculature of the mature root zone [157] and epidermis of the meristem zone [21]; PILS2 and PILS5 showed a particular overlapping expression in the root transition zone [146]. In root growth and development, PIN5, PILS2 and PILS5 play a negative role in primary root elongation [21,146,148]. The roots of PIN5, PILS2, or PILS5 gain-of-function mutants become shorter; on the contrary, the roots of loss-of-function mutants become longer [21,146,148]. This is because ER-localization auxin transport carriers, PIN5 and PILSs negatively regulate PAT and auxin signaling [21,134,146,147,154,155,156]. Interestingly, unlike PINs involved in intercellular transport, transcription of PIN5 and PILS5 is induced by cytokinin (Figure 1) [15,21,158,159], suggesting that PIN5 and PILS5 are other targets of cytokinin–auxin antagonism besides auxin conjugation enzymes GH3.5, GH3.6 and GH3.17. In other words, the process of cytokinin activating PIN5 and PILS5 expression reduces the amount of auxin transport to root elongation zones through intracellular auxin accumulation in bPAT transport cells. A recent paper reported that ARR1 binds directly to the PIN5 promoter to mediate cytokinin induction of PIN5 expression (Figure 1) [21]. Furthermore, it has been reported that PIN5-mediated intracellular auxin accumulation and GH3.17-mediated auxin conjugation with Glu are inextricably linked, which together regulate auxin homeostasis and signal transduction [21,160]; however, the details require further study.
7. Concluding Remarks
Once the seed germinates, the root meristem, which is derived from the proximal stem cells, proliferates and expands rapidly. After approximately 5 days of growth and development, the meristem of the root tip reaches a stable size, by which root growth is sustained. Cytokinin and auxin interactions play key roles in controlling the balance between the rate of cell division and differentiation, which is crucial for the maintenance of root meristems [11,19,26,161]. Here, we reviewed the cytokinin-mediated regulation on the components involved in auxin biosynthesis/metabolism, polar auxin transport, and intracellular transport of auxin in root tips (Figure 1). These processes are all related to establish the gradient distribution of auxin in root tips. They work together to control the size of the meristem, which is significantly reduced in taa1/ckrc1 [16], yuc8/ckrc2 [17], and the triple mutant pin1 pin3 pin7 [11], but incteased in pin5-3 [21], gh3.5, gh3.6 and gh3.17 [19,20,21].
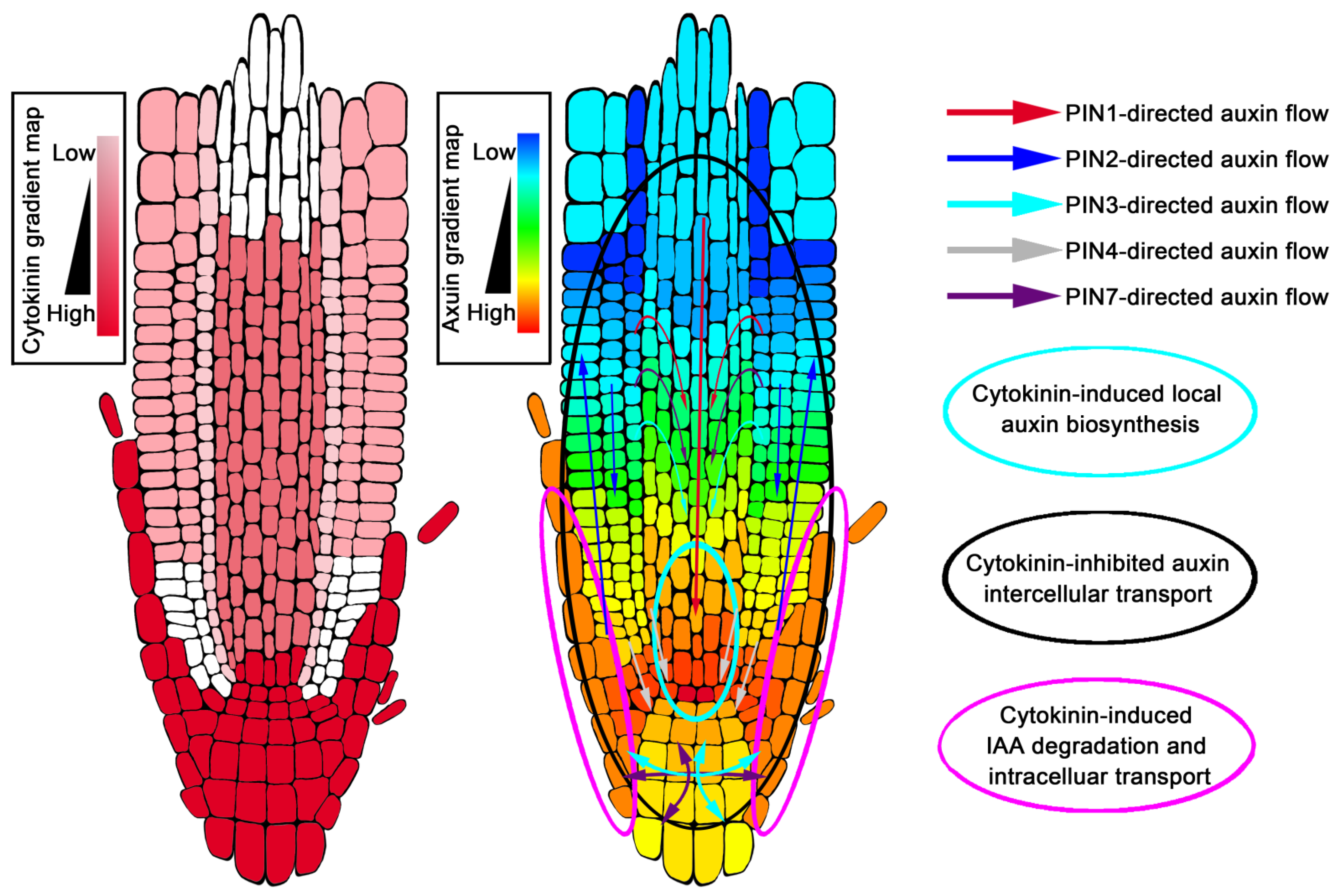
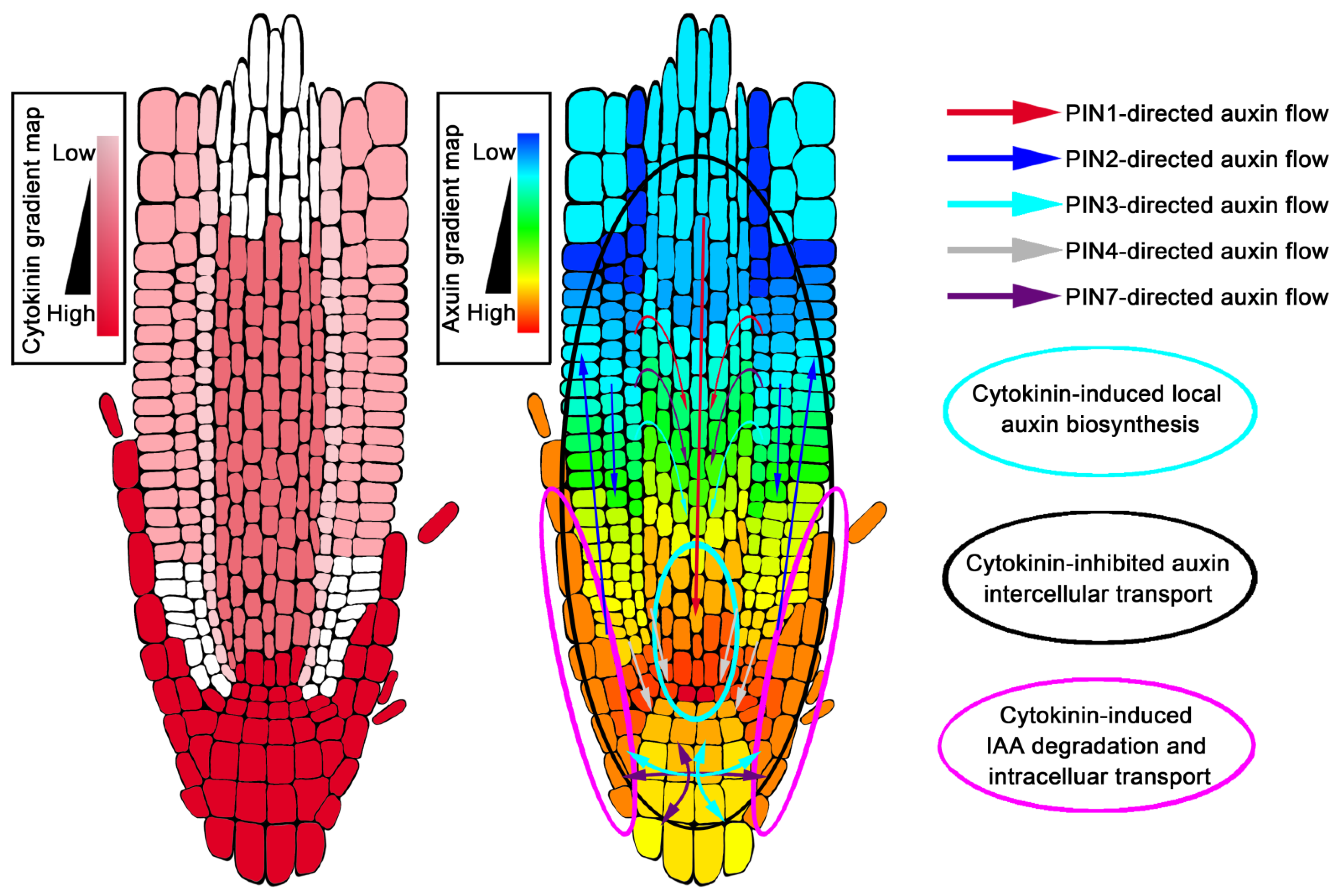
According to the current auxin gradient distribution model, auxin concentration is maximal at the QC of the root tip, but minimal at the boundary of the transition zone (Figure 2) [19,162]. The auxin maximum is very important for the maintenance of the QC and stem cell fate surrounding the QC [162], while the auxin minimum at the boundary of the transition zone acts as a signal to control the developmental switch from cell division to cell differentiation [19]. Earlier studies believed that the establishment of the maximum value of auxin in the root tip depended on the auxin transport mediated by PINs. This auxin maximum involves three transport processes, which are aPAT, bPAT and reflux of auxin. The auxin reflux redistributes auxin to form a characteristic gradient, and auxin is maximally located at the QC [162]. IAA3/SHY2, a negative regulator of auxin signal transduction, is a direct transcriptional target downstream of cytokinin signal AHK3-ARR1,12, which is one of the sites where the two hormones interact [11,138]. Cytokinin signaling suppresses auxin efflux carriers PIN1, PIN3 and PIN7 expressions through IAA3/SHY2 upregulation [11,26], and at the same time, promotes PIN1, PIN3 and PIN4 protein degradation to decrease PIN abundance at the post-transcriptional level [140]. Thus, cytokinin controls the flow of auxin to the root tip through the precise regulation of PINs. This process not only determines auxin levels that generate a minimum in the vascular tissue of the transition zone, but also determines the maximum value of auxin in the QC of the root tip (Figure 2) [11,19,26,140,163].
In addition, the auxin reflux mechanism gradually decreases auxin when it is transported along the external tissues of roots from the top to the base [162]; however, the formation of the external tissue auxin gradient is also regulated by cytokinin-mediated auxin degradation [19]. Several years ago, fluorescence-activated cell sorting of green fluorescent protein (GFP)-marked cell types, combined with solid-phase microextraction and an ultra-high-sensitivity mass spectrometry (MS) assay, was applied to analyze the levels of cytokinin in each tissue of the root tip (Figure 2) [164]. Cytokinin was found to be mainly concentrated in the lateral root cap, columella, columella initials, QC cells, and in the epidermis of the root tip, forming a gradient down to a maxima at the root cap [164]. Several components involved in cytokinin regulating auxin gradient distribution are specifically expressed in high cytokinin level regions of the root. When the auxin in the root tip was transported from the LRC to the elongation zone through bPAT, cytokinin could up-regulate the expression of genes GH3.5, GH3.6, GH3.17 and PIN5 in the LRC and the epidermis near the root tip to gradually reduce the amount of auxin transported by bPAT (Figure 2) [19,21,160].
Although calculated models based on the PINs and other auxin transporters indicate that the transport and redistribution of auxin from the above-ground parts is sufficient to produce the auxin gradient and auxin maximum in the root [165], the effect of local auxin biosynthesis on growth and development cannot be ignored [81,82,94,163]. Disturbing local auxin biosynthesis in the root tip will cause developmental defects, such as the small meristem and gravitropic defect [16,17,86]. There are evidences that auxin overproduction in shoots cannot completely rescue root auxin deficiency phenotypes, highlighting the importance of local auxin biosynthesis for root development [79]. In Section 3, we reviewed several processes by which local auxin biosynthesis is activated by cytokinin. At present, little is known about the physiological functions of these processes. We speculate that the local auxin biosynthesis induced by cytokinin may be a compensation mechanism of auxin. The establishment and maintenance of the gradient distribution of auxin in the root tip depends on auxin transport and reflux, and auxin is constantly attenuated by the downstream signal of cytokinin in the flow (Section 4 and Section 6). This compensation mechanism can offset the attenuated auxin, thereby maintaining the stability of the meristem.
In summary, recent studies highlight the important role of the cytokinin-controlled gradient distribution of auxin in root growth and development. However, some questions remain. For example, there are many mechanisms by which cytokinins regulate the gradient distribution of auxin; how do plants balance the local auxin biosynthesis, degradation, intercellular transport and intracellular transport in this process? Are these processes connected or dissected during plant growth and development?
Author Contributions
L.W. and G.-Q.G. drafted, wrote, and edited this review. J.-L.W. and X.-F.L. participated in the discussion of this review. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (31970713), Fundamental Research Funds for the Central Universities (lzujbky-2020-33) and Natural Science Foundation of Gansu Province (20JR5RA270).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Enders, T.A.; Strader, L.C. Auxin activity: Past, present, and future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavy, M.; Estelle, M. Mechanisms of auxin signaling. Development 2016, 143, 3226–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatini, S.; Beis, D.; Wolkenfelt, H.; Murfett, J.; Guilfoyle, T.; Malamy, J.; Benfey, P.; Leyser, O.; Bechtold, N.; Weisbeek, P.; et al. An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell 1999, 99, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Weijers, D.; Nemhauser, J.; Yang, Z. Auxin: Small molecule, big impact. J. Exp. Bot. 2018, 69, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Pacifici, E.; Polverari, L.; Sabatini, S. Plant hormone cross-talk: The pivot of root growth. J. Exp. Bot. 2015, 66, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis root development. Annu Rev. Plant Biol. 2012, 63, 563–590. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.O.; Skoog, F.; Von Saltza, M.H.; Strong, F. Kinetin, a cell division factor from deoxyribonucleic acid. J. Am. Chem. Soc. 1955, 77, 1392. [Google Scholar] [CrossRef]
- Miller, C.O.; Skoog, F.; Okomura, F.S.; von Saltza, M.H.; Strong, F.M. Isolation, structure and synthesis of kinetin, a substance promoting cell division. J. Am. Chem. Soc. 1956, 78, 1345–1350. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Dello Ioio, R.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A genetic framework for the control of cell division and differentiation in the root meristem. Science 2008, 322, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar] [CrossRef] [PubMed]
- Vieten, A.; Sauer, M.; Brewer, P.B.; Friml, J. Molecular and cellular aspects of auxin-transport-mediated development. Trends Plant Sci. 2007, 12, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Gunneras, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandberg, G.; Ljung, K. Cytokinin regulation of auxin synthesis in Arabidopsis involves a homeostatic feedback loop regulated via auxin and cytokinin signal transduction. Plant Cell 2010, 22, 2956–2969. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.Y.; Zhang, C.G.; Wu, L.; Zhang, C.G.; Chai, J.; Wang, M.; Jha, A.; Jia, P.F.; Cui, S.J.; Yang, M.; et al. Functional characterization of the CKRC1/TAA1 gene and dissection of hormonal actions in the Arabidopsis root. Plant J. 2011, 66, 516–527. [Google Scholar] [CrossRef]
- Di, D.W.; Wu, L.; Zhang, L.; An, C.W.; Zhang, T.Z.; Luo, P.; Gao, H.H.; Kriechbaumer, V.; Guo, G.Q. Functional roles of Arabidopsis CKRC2/YUCCA8 gene and the involvement of PIF4 in the regulation of auxin biosynthesis by cytokinin. Sci. Rep. 2016, 6, 36866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Liu, X.; Ljung, K.; Li, S.; Zhao, W.; Yang, F.; Wang, M.; Tao, Y. Type B Response Regulators Act As Central Integrators in Transcriptional Control of the Auxin Biosynthesis Enzyme TAA1. Plant Physiol. 2017, 175, 1438–1454. [Google Scholar] [CrossRef] [PubMed]
- Di Mambro, R.; De Ruvo, M.; Pacifici, E.; Salvi, E.; Sozzani, R.; Benfey, P.N.; Busch, W.; Novak, O.; Ljung, K.; Di Paola, L.; et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2017, 114, E7641–E7649. [Google Scholar] [CrossRef] [Green Version]
- Pierdonati, E.; Unterholzner, S.J.; Salvi, E.; Svolacchia, N.; Bertolotti, G.; Dello Ioio, R.; Sabatini, S.; Di Mambro, R. Cytokinin-Dependent Control of GH3 Group II Family Genes in the Arabidopsis Root. Plants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Di Mambro, R.; Svolacchia, N.; Dello Ioio, R.; Pierdonati, E.; Salvi, E.; Pedrazzini, E.; Vitale, A.; Perilli, S.; Sozzani, R.; Benfey, P.N.; et al. The Lateral Root Cap Acts as an Auxin Sink that Controls Meristem Size. Curr. Biol. 2019, 29, 1199–1205.e4. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Saez, R.; Mateo-Bonmati, E.; Ljung, K. Auxin Metabolism in Plants. Cold Spring Harb. Perspect. Biol. 2021, 13, a039867. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Moore, S.; Chen, C.; Lindsey, K. Crosstalk Complexities between Auxin, Cytokinin, and Ethylene in Arabidopsis Root Development: From Experiments to Systems Modeling, and Back Again. Mol. Plant 2017, 10, 1480–1496. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.; Zhang, X.; Mudge, A.; Rowe, J.H.; Topping, J.F.; Liu, J.; Lindsey, K. Spatiotemporal modelling of hormonal crosstalk explains the level and patterning of hormones and gene expression in Arabidopsis thaliana wild-type and mutant roots. New Phytol. 2015, 207, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraro, D.; Larrieu, A.; Lucas, M.; Chopard, J.; Byrne, H.; Godin, C.; King, J. A multi-scale model of the interplay between cell signalling and hormone transport in specifying the root meristem of Arabidopsis thaliana. J. Theor. Biol. 2016, 404, 182–205. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, K.; Simaskova, M.; Duclercq, J.; Petrasek, J.; Zazimalova, E.; Simon, S.; Friml, J.; Van Montagu, M.C.; Benkova, E. Cytokinin regulates root meristem activity via modulation of the polar auxin transport. Proc. Natl. Acad. Sci. USA 2009, 106, 4284–4289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Street, I.H.; Mathews, D.E.; Yamburkenko, M.V.; Sorooshzadeh, A.; John, R.T.; Swarup, R.; Bennett, M.J.; Kieber, J.J.; Schaller, G.E. Cytokinin acts through the auxin influx carrier AUX1 to regulate cell elongation in the root. Development 2016, 143, 3982–3993. [Google Scholar] [CrossRef] [Green Version]
- Argueso, C.T.; Ferreira, F.J.; Kieber, J.J. Environmental perception avenues: The interaction of cytokinin and environmental response pathways. Plant Cell Environ. 2009, 32, 1147–1160. [Google Scholar] [CrossRef]
- Bishopp, A.; Benkova, E.; Helariutta, Y. Sending mixed messages: Auxin-cytokinin crosstalk in roots. Curr. Opin. Plant Biol. 2011, 14, 10–16. [Google Scholar] [CrossRef]
- Chandler, J.W.; Werr, W. Cytokinin-auxin crosstalk in cell type specification. Trends Plant Sci. 2015, 20, 291–300. [Google Scholar] [CrossRef]
- Del Bianco, M.; Giustini, L.; Sabatini, S. Spatiotemporal changes in the role of cytokinin during root development. New Phytol. 2013, 199, 324–338. [Google Scholar] [CrossRef]
- Domagalska, M.A.; Leyser, O. Signal integration in the control of shoot branching. Nat. Rev. Mol. Cell Biol. 2011, 12, 211–221. [Google Scholar] [CrossRef] [PubMed]
- El-Showk, S.; Ruonala, R.; Helariutta, Y. Crossing paths: Cytokinin signalling and crosstalk. Development 2013, 140, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Sheen, J.; Muller, B. Cytokinin signaling networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, H.; Strader, L.C. Interplay of Auxin and Cytokinin in Lateral Root Development. Int. J. Mol. Sci. 2019, 20, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arab. Book 2014, 12, e0168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurepa, J.; Shull, T.E.; Smalle, J.A. Antagonistic activity of auxin and cytokinin in shoot and root organs. Plant Direct 2019, 3, e00121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin-auxin crosstalk. Trends Plant Sci. 2009, 14, 557–562. [Google Scholar] [CrossRef]
- Muller, C.J.; Larsson, E.; Spichal, L.; Sundberg, E. Cytokinin-Auxin Crosstalk in the Gynoecial Primordium Ensures Correct Domain Patterning. Plant Physiol. 2017, 175, 1144–1157. [Google Scholar] [CrossRef]
- Naseem, M.; Dandekar, T. The role of auxin-cytokinin antagonism in plant-pathogen interactions. PLoS Pathog. 2012, 8, e1003026. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.A.; Benkova, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef] [Green Version]
- Pernisova, M.; Kuderova, A.; Hejatko, J. Cytokinin and auxin interactions in plant development: Metabolism, signalling, transport and gene expression. Curr. Protein Pept. Sci. 2011, 12, 137–147. [Google Scholar] [CrossRef]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The yin-yang of hormones: Cytokinin and auxin interactions in plant development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-cytokinin interaction regulates meristem development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Tognetti, V.B.; Bielach, A.; Hrtyan, M. Redox regulation at the site of primary growth: Auxin, cytokinin and ROS crosstalk. Plant Cell Environ. 2017, 40, 2586–2605. [Google Scholar] [CrossRef] [PubMed]
- Vanstraelen, M.; Benkova, E. Hormonal interactions in the regulation of plant development. Annu. Rev. Cell Dev. Biol. 2012, 28, 463–487. [Google Scholar] [CrossRef] [PubMed]
- Waldie, T.; Leyser, O. Cytokinin Targets Auxin Transport to Promote Shoot Branching. Plant Physiol. 2018, 177, 803–818. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y. Auxin biosynthesis. Arab. Book 2014, 12, e0173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, J.P.; Kieber, J.J. Cytokinin signaling: Two-components and more. Trends Plant Sci. 2008, 13, 85–92. [Google Scholar] [CrossRef]
- Inoue, T.; Higuchi, M.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Kato, T.; Tabata, S.; Shinozaki, K.; Kakimoto, T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 2001, 409, 1060–1063. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miwa, K.; Ishikawa, K.; Yamada, H.; Aiba, H.; Mizuno, T. The Arabidopsis sensor His-kinase, AHk4, can respond to cytokinins. Plant Cell Physiol. 2001, 42, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Ueguchi, C.; Sato, S.; Kato, T.; Tabata, S. The AHK4 gene involved in the cytokinin-signaling pathway as a direct receptor molecule in Arabidopsis thaliana. Plant Cell Physiol. 2001, 42, 751–755. [Google Scholar] [CrossRef]
- Yamada, H.; Suzuki, T.; Terada, K.; Takei, K.; Ishikawa, K.; Miwa, K.; Yamashino, T.; Mizuno, T. The Arabidopsis AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin signals across the membrane. Plant Cell Physiol. 2001, 42, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, C.E.; Li, J.; Argueso, C.; Gonzalez, M.; Lee, E.; Lewis, M.W.; Maxwell, B.B.; Perdue, T.D.; Schaller, G.E.; Alonso, J.M.; et al. The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling. Plant Cell 2006, 18, 3073–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, S.; Urao, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of genes for two-component phosphorelay mediators with a single HPt domain in Arabidopsis thaliana. FEBS Lett. 1998, 437, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Imamura, A.; Ueguchi, C.; Mizuno, T. Histidine-containing phosphotransfer (HPt) signal transducers implicated in His-to-Asp phosphorelay in Arabidopsis. Plant Cell Physiol. 1998, 39, 1258–1268. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Suzuki, T.; Yamashino, T.; Mizuno, T. Comparative studies of the AHP histidine-containing phosphotransmitters implicated in His-to-Asp phosphorelay in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2004, 68, 462–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argyros, R.D.; Mathews, D.E.; Chiang, Y.H.; Palmer, C.M.; Thibault, D.M.; Etheridge, N.; Argyros, D.A.; Mason, M.G.; Kieber, J.J.; Schaller, G.E. Type B response regulators of Arabidopsis play key roles in cytokinin signaling and plant development. Plant Cell 2008, 20, 2102–2116. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Yamashino, T.; Yokoyama, A.; Mizuno, T. Three type-B response regulators, ARR1, ARR10 and ARR12, play essential but redundant roles in cytokinin signal transduction throughout the life cycle of Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Mason, M.G.; Mathews, D.E.; Argyros, D.A.; Maxwell, B.B.; Kieber, J.J.; Alonso, J.M.; Ecker, J.R.; Schaller, G.E. Multiple type-B response regulators mediate cytokinin signal transduction in Arabidopsis. Plant Cell 2005, 17, 3007–3018. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, A.; Yamashino, T.; Amano, Y.; Tajima, Y.; Imamura, A.; Sakakibara, H.; Mizuno, T. Type-B ARR transcription factors, ARR10 and ARR12, are implicated in cytokinin-mediated regulation of protoxylem differentiation in roots of Arabidopsis thaliana. Plant Cell Physiol. 2007, 48, 84–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiba, T.; Yamada, H.; Sato, S.; Kato, T.; Tabata, S.; Yamashino, T.; Mizuno, T. The type-A response regulator, ARR15, acts as a negative regulator in the cytokinin-mediated signal transduction in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 868–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.J.; Park, J.Y.; Ku, S.J.; Ha, Y.M.; Kim, S.; Kim, M.D.; Oh, M.H.; Kim, J. Genome-wide expression profiling of ARABIDOPSIS RESPONSE REGULATOR 7(ARR7) overexpression in cytokinin response. Mol. Genet. Genom. 2007, 277, 115–137. [Google Scholar] [CrossRef] [PubMed]
- To, J.P.; Deruere, J.; Maxwell, B.B.; Morris, V.F.; Hutchison, C.E.; Ferreira, F.J.; Schaller, G.E.; Kieber, J.J. Cytokinin regulates type-A Arabidopsis Response Regulator activity and protein stability via two-component phosphorelay. Plant Cell 2007, 19, 3901–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, J.P.; Haberer, G.; Ferreira, F.J.; Deruere, J.; Mason, M.G.; Schaller, G.E.; Alonso, J.M.; Ecker, J.R.; Kieber, J.J. Type-A Arabidopsis response regulators are partially redundant negative regulators of cytokinin signaling. Plant Cell 2004, 16, 658–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashotte, A.M.; Carson, S.D.; To, J.P.; Kieber, J.J. Expression profiling of cytokinin action in Arabidopsis. Plant Physiol. 2003, 132, 1998–2011. [Google Scholar] [CrossRef] [Green Version]
- Rashotte, A.M.; Mason, M.G.; Hutchison, C.E.; Ferreira, F.J.; Schaller, G.E.; Kieber, J.J. A subset of Arabidopsis AP2 transcription factors mediates cytokinin responses in concert with a two-component pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 11081–11085. [Google Scholar] [CrossRef] [Green Version]
- Normanly, J. Approaching cellular and molecular resolution of auxin biosynthesis and metabolism. Cold Spring Harb. Perspect. Biol. 2010, 2, a001594. [Google Scholar] [CrossRef]
- Nonhebel, H.M. Tryptophan-Independent Indole-3-Acetic Acid Synthesis: Critical Evaluation of the Evidence. Plant Physiol. 2015, 169, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [Green Version]
- Won, C.; Shen, X.; Mashiguchi, K.; Zheng, Z.; Dai, X.; Cheng, Y.; Kasahara, H.; Kamiya, Y.; Chory, J.; Zhao, Y. Conversion of tryptophan to indole-3-acetic acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18518–18523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanova, A.N.; Robertson-Hoyt, J.; Yun, J.; Benavente, L.M.; Xie, D.Y.; Dolezal, K.; Schlereth, A.; Jurgens, G.; Alonso, J.M. TAA1-mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell 2008, 133, 177–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Ferrer, J.L.; Ljung, K.; Pojer, F.; Hong, F.; Long, J.A.; Li, L.; Moreno, J.E.; Bowman, M.E.; Ivans, L.J.; et al. Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants. Cell 2008, 133, 164–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Greenham, K.; Prigge, M.J.; Jensen, P.J.; Estelle, M. The TRANSPORT INHIBITOR RESPONSE2 gene is required for auxin synthesis and diverse aspects of plant development. Plant Physiol. 2009, 151, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanova, A.N.; Yun, J.; Robles, L.M.; Novak, O.; He, W.; Guo, H.; Ljung, K.; Alonso, J.M. The Arabidopsis YUCCA1 flavin monooxygenase functions in the indole-3-pyruvic acid branch of auxin biosynthesis. Plant Cell 2011, 23, 3961–3973. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Sharkhuu, A.; Jin, J.B.; Li, P.; Jeong, J.C.; Baek, D.; Lee, S.Y.; Blakeslee, J.J.; Murphy, A.S.; Bohnert, H.J.; et al. yucca6, a dominant mutation in Arabidopsis, affects auxin accumulation and auxin-related phenotypes. Plant Physiol. 2007, 145, 722–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef]
- Chen, Q.; Dai, X.; De-Paoli, H.; Cheng, Y.; Takebayashi, Y.; Kasahara, H.; Kamiya, Y.; Zhao, Y. Auxin overproduction in shoots cannot rescue auxin deficiencies in Arabidopsis roots. Plant Cell Physiol. 2014, 55, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin synthesized by the YUCCA flavin monooxygenases is essential for embryogenesis and leaf formation in Arabidopsis. Plant Cell 2007, 19, 2430–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, M.; Liang, N.; Zheng, Y.; Yu, Q.; Wu, S. Symplastic communication spatially directs local auxin biosynthesis to maintain root stem cell niche in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 4005–4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumos, J.; Robles, L.M.; Yun, J.; Vu, T.C.; Jackson, S.; Alonso, J.M.; Stepanova, A.N. Local Auxin Biosynthesis Is a Key Regulator of Plant Development. Dev. Cell 2018, 47, 306–318.e5. [Google Scholar] [CrossRef] [Green Version]
- Della Rovere, F.; Fattorini, L.; D’Angeli, S.; Veloccia, A.; Falasca, G.; Altamura, M.M. Auxin and cytokinin control formation of the quiescent centre in the adventitious root apex of Arabidopsis. Ann. Bot. 2013, 112, 1395–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanova, A.N.; Hoyt, J.M.; Hamilton, A.A.; Alonso, J.M. A Link between ethylene and auxin uncovered by the characterization of two root-specific ethylene-insensitive mutants in Arabidopsis. Plant Cell 2005, 17, 2230–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moubayidin, L.; Di Mambro, R.; Sozzani, R.; Pacifici, E.; Salvi, E.; Terpstra, I.; Bao, D.; van Dijken, A.; Dello Ioio, R.; Perilli, S.; et al. Spatial coordination between stem cell activity and cell differentiation in the root meristem. Dev. Cell 2013, 26, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, D.W.; Wu, L.; Luo, P.; Zhang, L.; Zhang, T.Z.; Sun, X.; Wei, S.D.; An, C.W.; Guo, G.Q. Analysis the role of Arabidopsis CKRC6/ASA1 in auxin and cytokinin biosynthesis. J. Plant Biol. 2016, 59, 162–171. [Google Scholar] [CrossRef]
- Ljung, K.; Hull, A.K.; Celenza, J.; Yamada, M.; Estelle, M.; Normanly, J.; Sandberg, G. Sites and regulation of auxin biosynthesis in Arabidopsis roots. Plant Cell 2005, 17, 1090–1104. [Google Scholar] [CrossRef]
- Zhao, Y.; Hull, A.K.; Gupta, N.R.; Goss, K.A.; Alonso, J.; Ecker, J.R.; Normanly, J.; Chory, J.; Celenza, J.L. Trp-dependent auxin biosynthesis in Arabidopsis: Involvement of cytochrome P450s CYP79B2 and CYP79B3. Genes Dev. 2002, 16, 3100–3112. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Ljung, K. Auxin and cytokinin regulate each other’s levels via a metabolic feedback loop. Plant Signal. Behav. 2011, 6, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Chen, H.; Huang, L.; O’Neil, R.C.; Shokhirev, M.N.; Ecker, J.R. A B-ARR-mediated cytokinin transcriptional network directs hormone cross-regulation and shoot development. Nat. Commun. 2018, 9, 1604. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Luo, P.; Bai, J.; Wu, L.; Di, D.W.; Liu, H.Q.; Li, J.J.; Liu, Y.L.; Khaskheli, A.J.; Zhao, C.M.; et al. Function of histone H2B monoubiquitination in transcriptional regulation of auxin biosynthesis in Arabidopsis. Commun Biol. 2021, 4, 206. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olatunji, D.; Geelen, D.; Verstraeten, I. Control of Endogenous Auxin Levels in Plant Root Development. Int. J. Mol. Sci. 2017, 18, 2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, B.; Yan, Z.; Tian, H.; Zhang, X.; Ding, Z. Local Auxin Biosynthesis Mediates Plant Growth and Development. Trends Plant Sci. 2019, 24, 6–9. [Google Scholar] [CrossRef]
- Mellor, N.; Band, L.R.; Pencik, A.; Novak, O.; Rashed, A.; Holman, T.; Wilson, M.H.; Voss, U.; Bishopp, A.; King, J.R.; et al. Dynamic regulation of auxin oxidase and conjugating enzymes AtDAO1 and GH3 modulates auxin homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 11022–11027. [Google Scholar] [CrossRef] [Green Version]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Westfall, C.S.; Hicks, L.M.; Wang, S.; Jez, J.M. Kinetic basis for the conjugation of auxin by a GH3 family indole-acetic acid-amido synthetase. J. Biol. Chem. 2010, 285, 29780–29786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeClere, S.; Tellez, R.; Rampey, R.A.; Matsuda, S.P.; Bartel, B. Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J. Biol. Chem. 2002, 277, 20446–20452. [Google Scholar] [CrossRef] [Green Version]
- Rampey, R.A.; LeClere, S.; Kowalczyk, M.; Ljung, K.; Sandberg, G.; Bartel, B. A family of auxin-conjugate hydrolases that contributes to free indole-3-acetic acid levels during Arabidopsis germination. Plant Physiol. 2004, 135, 978–988. [Google Scholar] [CrossRef] [Green Version]
- Kai, K.; Horita, J.; Wakasa, K.; Miyagawa, H. Three oxidative metabolites of indole-3-acetic acid from Arabidopsis thaliana. Phytochemistry 2007, 68, 1651–1663. [Google Scholar] [CrossRef]
- Ostin, A.; Kowalyczk, M.; Bhalerao, R.P.; Sandberg, G. Metabolism of indole-3-acetic acid in Arabidopsis. Plant Physiol. 1998, 118, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Pencik, A.; Simonovik, B.; Petersson, S.V.; Henykova, E.; Simon, S.; Greenham, K.; Zhang, Y.; Kowalczyk, M.; Estelle, M.; Zazimalova, E.; et al. Regulation of auxin homeostasis and gradients in Arabidopsis roots through the formation of the indole-3-acetic acid catabolite 2-oxindole-3-acetic acid. Plant Cell 2013, 25, 3858–3870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljung, K. Auxin metabolism and homeostasis during plant development. Development 2013, 140, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.E.; Park, J.Y.; Kim, Y.S.; Staswick, P.E.; Jeon, J.; Yun, J.; Kim, S.Y.; Kim, J.; Lee, Y.H.; Park, C.M. GH3-mediated auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. J. Biol. Chem. 2007, 282, 10036–10046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westfall, C.S.; Sherp, A.M.; Zubieta, C.; Alvarez, S.; Schraft, E.; Marcellin, R.; Ramirez, L.; Jez, J.M. Arabidopsis thaliana GH3.5 acyl acid amido synthetase mediates metabolic crosstalk in auxin and salicylic acid homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 13917–13922. [Google Scholar] [CrossRef] [Green Version]
- Skalicky, V.; Kubes, M.; Napier, R.; Novak, O. Auxins and Cytokinins-The Role of Subcellular Organization on Homeostasis. Int. J. Mol. Sci. 2018, 19, 3115. [Google Scholar] [CrossRef] [Green Version]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organisation of the Arabidopsis thaliana root. Development 1993, 119, 71–84. [Google Scholar]
- Blancaflor, E.B.; Fasano, J.M.; Gilroy, S. Laser ablation of root cap cells: Implications for models of graviperception. Adv. Space Res. 1999, 24, 731–738. [Google Scholar] [CrossRef]
- Tsugeki, R.; Fedoroff, N.V. Genetic ablation of root cap cells in Arabidopsis. Proc. Natl. Acad. Sci. USA 1999, 96, 12941–12946. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Kramer, E.M.; Perry, P.; Knox, K.; Leyser, H.M.; Haseloff, J.; Beemster, G.T.; Bhalerao, R.; Bennett, M.J. Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nat. Cell Biol. 2005, 7, 1057–1065. [Google Scholar] [CrossRef]
- Peret, B.; Swarup, K.; Ferguson, A.; Seth, M.; Yang, Y.; Dhondt, S.; James, N.; Casimiro, I.; Perry, P.; Syed, A.; et al. AUX/LAX genes encode a family of auxin influx transporters that perform distinct functions during Arabidopsis development. Plant Cell 2012, 24, 2874–2885. [Google Scholar] [CrossRef] [Green Version]
- Swarup, K.; Benkova, E.; Swarup, R.; Casimiro, I.; Peret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S.; et al. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef]
- Swarup, R.; Friml, J.; Marchant, A.; Ljung, K.; Sandberg, G.; Palme, K.; Bennett, M. Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex. Genes Dev. 2001, 15, 2648–2653. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Peret, B. AUX/LAX family of auxin influx carriers-an overview. Front. Plant Sci. 2012, 3, 225. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Bhosale, R. Developmental Roles of AUX1/LAX Auxin Influx Carriers in Plants. Front. Plant Sci. 2019, 10, 1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krecek, P.; Skupa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zazimalova, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zazimalova, E.; Murphy, A.S.; Yang, H.; Hoyerova, K.; Hosek, P. Auxin transporters—Why so many? Cold Spring Harb. Perspect. Biol. 2010, 2, a001552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwiewka, M.; Bilanovicova, V.; Seifu, Y.W.; Nodzynski, T. The Nuts and Bolts of PIN Auxin Efflux Carriers. Front. Plant Sci. 2019, 10, 985. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Kleine-Vehn, J. PIN-FORMED and PIN-LIKES auxin transport facilitators. Development 2019, 146, dev168088. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Cho, H.T. The function of ABCB transporters in auxin transport. Plant Signal. Behav. 2013, 8, e22990. [Google Scholar] [CrossRef] [Green Version]
- Verrier, P.J.; Bird, D.; Burla, B.; Dassa, E.; Forestier, C.; Geisler, M.; Klein, M.; Kolukisaoglu, U.; Lee, Y.; Martinoia, E.; et al. Plant ABC proteins—a unified nomenclature and updated inventory. Trends Plant Sci. 2008, 13, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Benkova, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertova, D.; Jurgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Blakeslee, J.J.; Peer, W.A.; Murphy, A.S. Auxin transport. Curr. Opin Plant Biol. 2005, 8, 494–500. [Google Scholar] [CrossRef]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jurgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Grebe, M.; Friml, J.; Swarup, R.; Ljung, K.; Sandberg, G.; Terlou, M.; Palme, K.; Bennett, M.J.; Scheres, B. Cell polarity signaling in Arabidopsis involves a BFA-sensitive auxin influx pathway. Curr. Biol. 2002, 12, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Retzer, K.; Vosolsobe, S.; Napier, R. Advances in Understanding the Mechanism of Action of the Auxin Permease AUX1. Int. J. Mol. Sci. 2018, 19, 3391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubes, M.; Yang, H.; Richter, G.L.; Cheng, Y.; Mlodzinska, E.; Wang, X.; Blakeslee, J.J.; Carraro, N.; Petrasek, J.; Zazimalova, E.; et al. The Arabidopsis concentration-dependent influx/efflux transporter ABCB4 regulates cellular auxin levels in the root epidermis. Plant J. 2012, 69, 640–654. [Google Scholar] [CrossRef]
- Yang, H.; Murphy, A.S. Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 2009, 59, 179–191. [Google Scholar] [CrossRef]
- Bishopp, A.; Lehesranta, S.; Vaten, A.; Help, H.; El-Showk, S.; Scheres, B.; Helariutta, K.; Mahonen, A.P.; Sakakibara, H.; Helariutta, Y. Phloem-transported cytokinin regulates polar auxin transport and maintains vascular pattern in the root meristem. Curr. Biol. 2011, 21, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Pernisova, M.; Prat, T.; Grones, P.; Harustiakova, D.; Matonohova, M.; Spichal, L.; Nodzynski, T.; Friml, J.; Hejatko, J. Cytokinins influence root gravitropism via differential regulation of auxin transporter expression and localization in Arabidopsis. New Phytol. 2016, 212, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.; Chae, H.S.; Kieber, J.J. Regulation of ACS protein stability by cytokinin and brassinosteroid. Plant J. 2009, 57, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, K.; Ljung, K.; Vanneste, S.; Podhorska, R.; Beeckman, T.; Friml, J.; Benkova, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [Green Version]
- Abdollahi Sisi, N.; Ruzicka, K. ER-Localized PIN Carriers: Regulators of Intracellular Auxin Homeostasis. Plants 2020, 9, 1527. [Google Scholar] [CrossRef]
- Petrasek, J.; Mravec, J.; Bouchard, R.; Blakeslee, J.J.; Abas, M.; Seifertova, D.; Wisniewska, J.; Tadele, Z.; Kubes, M.; Covanova, M.; et al. PIN proteins perform a rate-limiting function in cellular auxin efflux. Science 2006, 312, 914–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernisova, M.; Klima, P.; Horak, J.; Valkova, M.; Malbeck, J.; Soucek, P.; Reichman, P.; Hoyerova, K.; Dubova, J.; Friml, J.; et al. Cytokinins modulate auxin-induced organogenesis in plants via regulation of the auxin efflux. Proc. Natl. Acad. Sci. USA 2009, 106, 3609–3614. [Google Scholar] [CrossRef] [Green Version]
- Simaskova, M.; O’Brien, J.A.; Khan, M.; Van Noorden, G.; Otvos, K.; Vieten, A.; De Clercq, I.; Van Haperen, J.M.A.; Cuesta, C.; Hoyerova, K.; et al. Cytokinin response factors regulate PIN-FORMED auxin transporters. Nat. Commun. 2015, 6, 8717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, M.; Sasaki, N.; Tsuge, T.; Aoyama, T.; Oka, A. ARR1 directly activates cytokinin response genes that encode proteins with diverse regulatory functions. Plant Cell Physiol. 2007, 48, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Olalde, J.I.; Zuniga-Mayo, V.M.; Serwatowska, J.; Chavez Montes, R.A.; Lozano-Sotomayor, P.; Herrera-Ubaldo, H.; Gonzalez-Aguilera, K.L.; Ballester, P.; Ripoll, J.J.; Ezquer, I.; et al. The bHLH transcription factor SPATULA enables cytokinin signaling, and both activate auxin biosynthesis and transport genes at the medial domain of the gynoecium. PLoS Genet. 2017, 13, e1006726. [Google Scholar] [CrossRef]
- Zhang, W.; To, J.P.; Cheng, C.Y.; Schaller, G.E.; Kieber, J.J. Type-A response regulators are required for proper root apical meristem function through post-transcriptional regulation of PIN auxin efflux carriers. Plant J. 2011, 68, 1–10. [Google Scholar] [CrossRef]
- Marhavy, P.; Bielach, A.; Abas, L.; Abuzeineh, A.; Duclercq, J.; Tanaka, H.; Parezova, M.; Petrasek, J.; Friml, J.; Kleine-Vehn, J.; et al. Cytokinin modulates endocytic trafficking of PIN1 auxin efflux carrier to control plant organogenesis. Dev. Cell 2011, 21, 796–804. [Google Scholar] [CrossRef]
- Marhavy, P.; Duclercq, J.; Weller, B.; Feraru, E.; Bielach, A.; Offringa, R.; Friml, J.; Schwechheimer, C.; Murphy, A.; Benkova, E. Cytokinin controls polarity of PIN1-dependent auxin transport during lateral root organogenesis. Curr. Biol. 2014, 24, 1031–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchant, A.; Kargul, J.; May, S.T.; Muller, P.; Delbarre, A.; Perrot-Rechenmann, C.; Bennett, M.J. AUX1 regulates root gravitropism in Arabidopsis by facilitating auxin uptake within root apical tissues. EMBO J. 1999, 18, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Swarup, R.; Bennett, M.; Schaller, G.E.; Kieber, J.J. Cytokinin induces cell division in the quiescent center of the Arabidopsis root apical meristem. Curr. Biol. 2013, 23, 1979–1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Zorrilla, J.M.; Lopez-Vidriero, I.; Carrasco, J.L.; Godoy, M.; Vera, P.; Solano, R. DNA-binding specificities of plant transcription factors and their potential to define target genes. Proc. Natl. Acad. Sci. USA 2014, 111, 2367–2372. [Google Scholar] [CrossRef] [Green Version]
- Barbez, E.; Kubes, M.; Rolcik, J.; Beziat, C.; Pencik, A.; Wang, B.; Rosquete, M.R.; Zhu, J.; Dobrev, P.I.; Lee, Y.; et al. A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nature 2012, 485, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Feraru, E.; Vosolsobe, S.; Feraru, M.I.; Petrasek, J.; Kleine-Vehn, J. Evolution and Structural Diversification of PILS Putative Auxin Carriers in Plants. Front. Plant Sci. 2012, 3, 227. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Feraru, E.; Feraru, M.I.; Waidmann, S.; Wang, W.; Passaia, G.; Wang, Z.Y.; Wabnik, K.; Kleine-Vehn, J. PIN-LIKES Coordinate Brassinosteroid Signaling with Nuclear Auxin Input in Arabidopsis thaliana. Curr. Biol. 2020, 30, 1579–1588.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paponov, I.A.; Teale, W.D.; Trebar, M.; Blilou, I.; Palme, K. The PIN auxin efflux facilitators: Evolutionary and functional perspectives. Trends Plant Sci. 2005, 10, 170–177. [Google Scholar] [CrossRef]
- Simon, S.; Skupa, P.; Viaene, T.; Zwiewka, M.; Tejos, R.; Klima, P.; Carna, M.; Rolcik, J.; De Rycke, R.; Moreno, I.; et al. PIN6 auxin transporter at endoplasmic reticulum and plasma membrane mediates auxin homeostasis and organogenesis in Arabidopsis. New Phytol. 2016, 211, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Wang, B.; Moreno, I.; Duplakova, N.; Simon, S.; Carraro, N.; Reemmer, J.; Pencik, A.; Chen, X.; Tejos, R.; et al. ER-localized auxin transporter PIN8 regulates auxin homeostasis and male gametophyte development in Arabidopsis. Nat. Commun. 2012, 3, 941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Bosco, C.; Dovzhenko, A.; Palme, K. Intracellular auxin transport in pollen: PIN8, PIN5 and PILS5. Plant Signal. Behav. 2012, 7, 1504–1505. [Google Scholar] [CrossRef] [Green Version]
- Sawchuk, M.G.; Edgar, A.; Scarpella, E. Patterning of leaf vein networks by convergent auxin transport pathways. PLoS Genet. 2013, 9, e1003294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mravec, J.; Skupa, P.; Bailly, A.; Hoyerova, K.; Krecek, P.; Bielach, A.; Petrasek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.D.; et al. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef]
- Beziat, C.; Barbez, E.; Feraru, M.I.; Lucyshyn, D.; Kleine-Vehn, J. Light triggers PILS-dependent reduction in nuclear auxin signalling for growth transition. Nat. Plants 2017, 3, 17105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feraru, E.; Feraru, M.I.; Barbez, E.; Waidmann, S.; Sun, L.; Gaidora, A.; Kleine-Vehn, J. PILS6 is a temperature-sensitive regulator of nuclear auxin input and organ growth in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2019, 116, 3893–3898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, A.; Park, M.; Kesawat, M.S.; Cho, H.T. Functional Analysis of the Hydrophilic Loop in Intracellular Trafficking of Arabidopsis PIN-FORMED Proteins. Plant Cell 2014, 26, 1570–1585. [Google Scholar] [CrossRef] [Green Version]
- Brenner, W.G.; Ramireddy, E.; Heyl, A.; Schmulling, T. Gene regulation by cytokinin in Arabidopsis. Front. Plant Sci. 2012, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Brenner, W.G.; Schmulling, T. Transcript profiling of cytokinin action in Arabidopsis roots and shoots discovers largely similar but also organ-specific responses. BMC Plant Biol. 2012, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, K.; Hejatko, J. Auxin transport and conjugation caught together. J. Exp. Bot. 2017, 68, 4409–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dello Ioio, R.; Linhares, F.S.; Scacchi, E.; Casamitjana-Martinez, E.; Heidstra, R.; Costantino, P.; Sabatini, S. Cytokinins determine Arabidopsis root-meristem size by controlling cell differentiation. Curr. Biol. 2007, 17, 678–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieneisen, V.A.; Xu, J.; Maree, A.F.; Hogeweg, P.; Scheres, B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007, 449, 1008–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersson, S.V.; Johansson, A.I.; Kowalczyk, M.; Makoveychuk, A.; Wang, J.Y.; Moritz, T.; Grebe, M.; Benfey, P.N.; Sandberg, G.; Ljung, K. An auxin gradient and maximum in the Arabidopsis root apex shown by high-resolution cell-specific analysis of IAA distribution and synthesis. Plant Cell 2009, 21, 1659–1668. [Google Scholar] [CrossRef]
- Antoniadi, I.; Plackova, L.; Simonovik, B.; Dolezal, K.; Turnbull, C.; Ljung, K.; Novak, O. Cell-Type-Specific Cytokinin Distribution within the Arabidopsis Primary Root Apex. Plant Cell 2015, 27, 1955–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armengot, L.; Marques-Bueno, M.M.; Jaillais, Y. Regulation of polar auxin transport by protein and lipid kinases. J. Exp. Bot. 2016, 67, 4015–4037. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Auxin biosynthesis/metabolism and transport regulated by cytokinin.
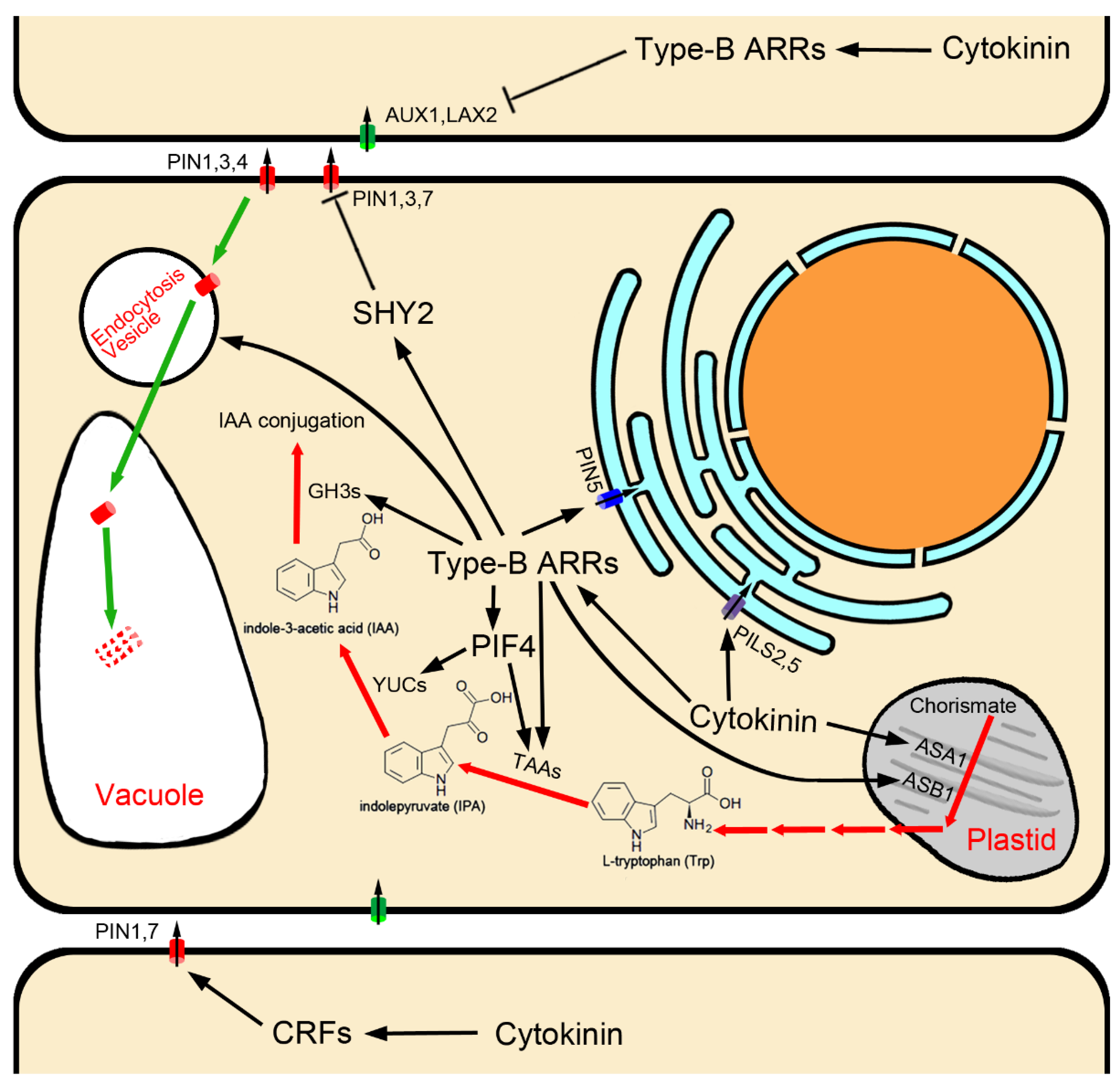
Figure 2.
Cytokinin-controlled gradient distribution of auxin in Arabidopsis root tip.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, L.; Wang, J.-L.; Li, X.-F.; Guo, G.-Q. Cytokinin-Controlled Gradient Distribution of Auxin in Arabidopsis Root Tip. Int. J. Mol. Sci. 2021, 22, 3874. https://doi.org/10.3390/ijms22083874
AMA Style
Wu L, Wang J-L, Li X-F, Guo G-Q. Cytokinin-Controlled Gradient Distribution of Auxin in Arabidopsis Root Tip. International Journal of Molecular Sciences. 2021; 22(8):3874. https://doi.org/10.3390/ijms22083874
Chicago/Turabian StyleWu, Lei, Jun-Li Wang, Xiao-Feng Li, and Guang-Qin Guo. 2021. "Cytokinin-Controlled Gradient Distribution of Auxin in Arabidopsis Root Tip" International Journal of Molecular Sciences 22, no. 8: 3874. https://doi.org/10.3390/ijms22083874
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.