Polyamine Action under Metal/Metalloid Stress: Regulation of Biosynthesis, Metabolism, and Molecular Interactions

 ,
, 
 , ,
, ,  ,
,
Abstract
:1. Introduction
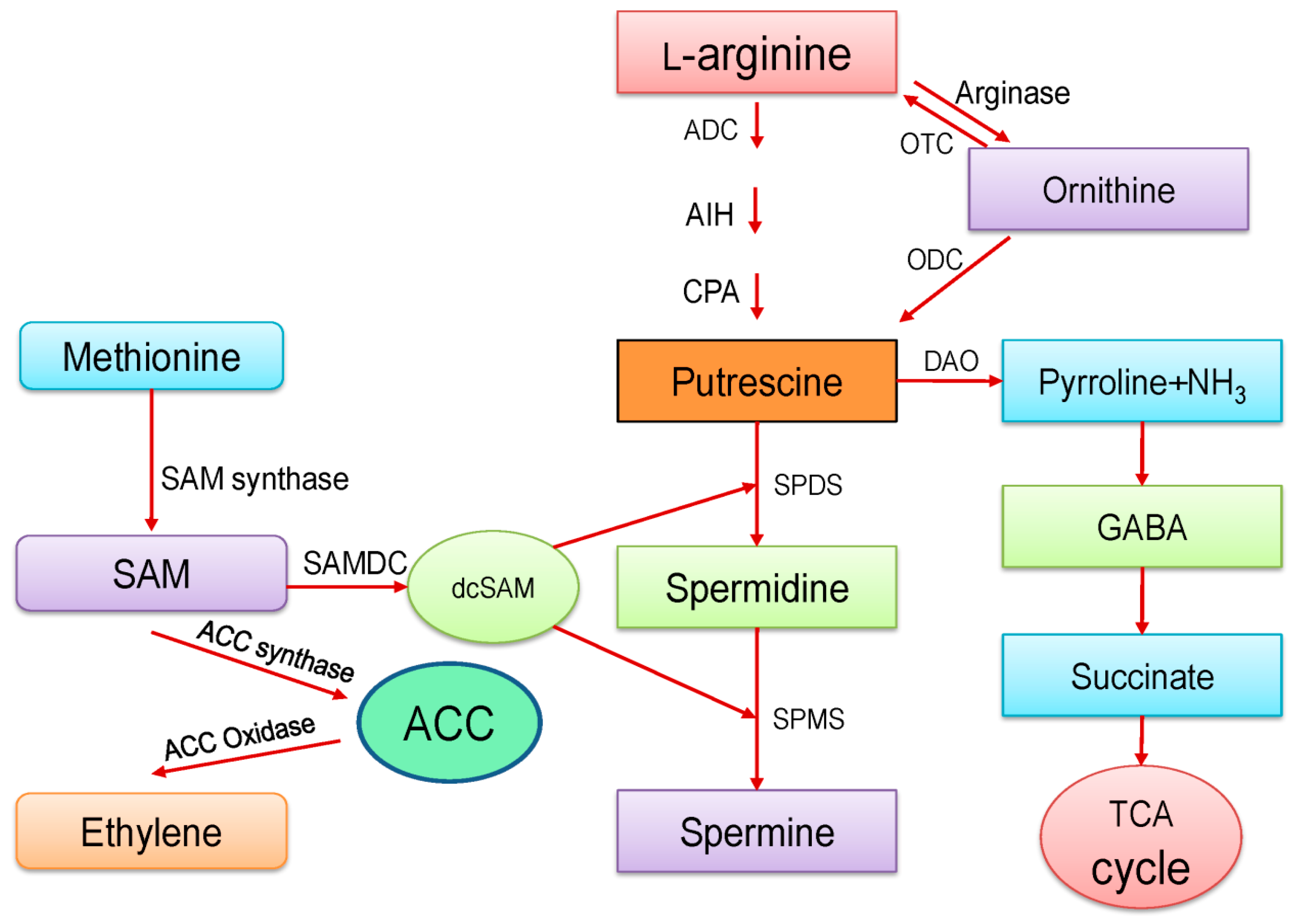
2. Polyamines Biosynthesis and Metabolism
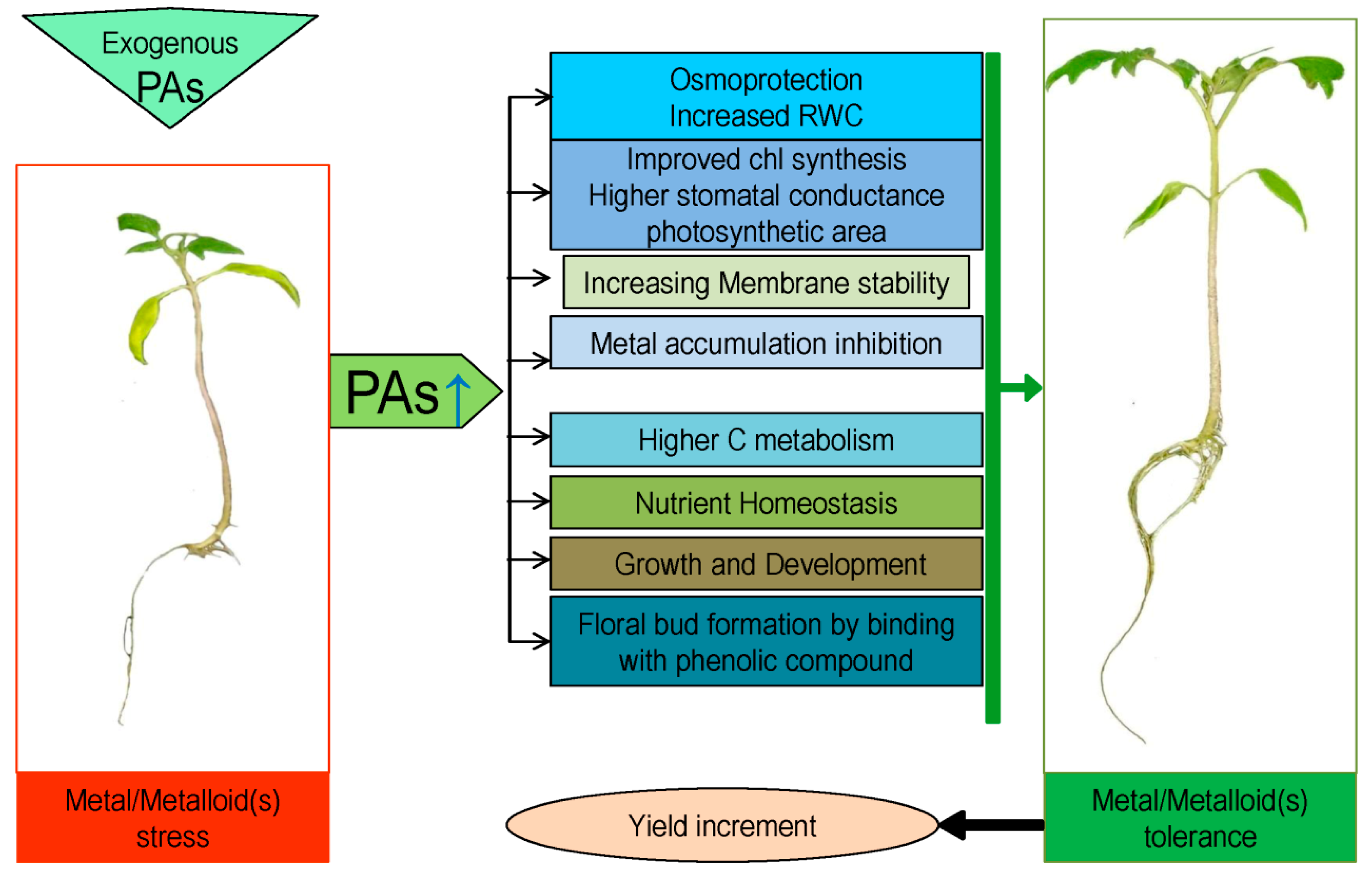
3. Polyamine-Induced Metal/Metalloids Tolerance in Plants
4. Polyamine-Induced Antioxidant Defense in Plants under Metal/Metalloid(S) Toxicity
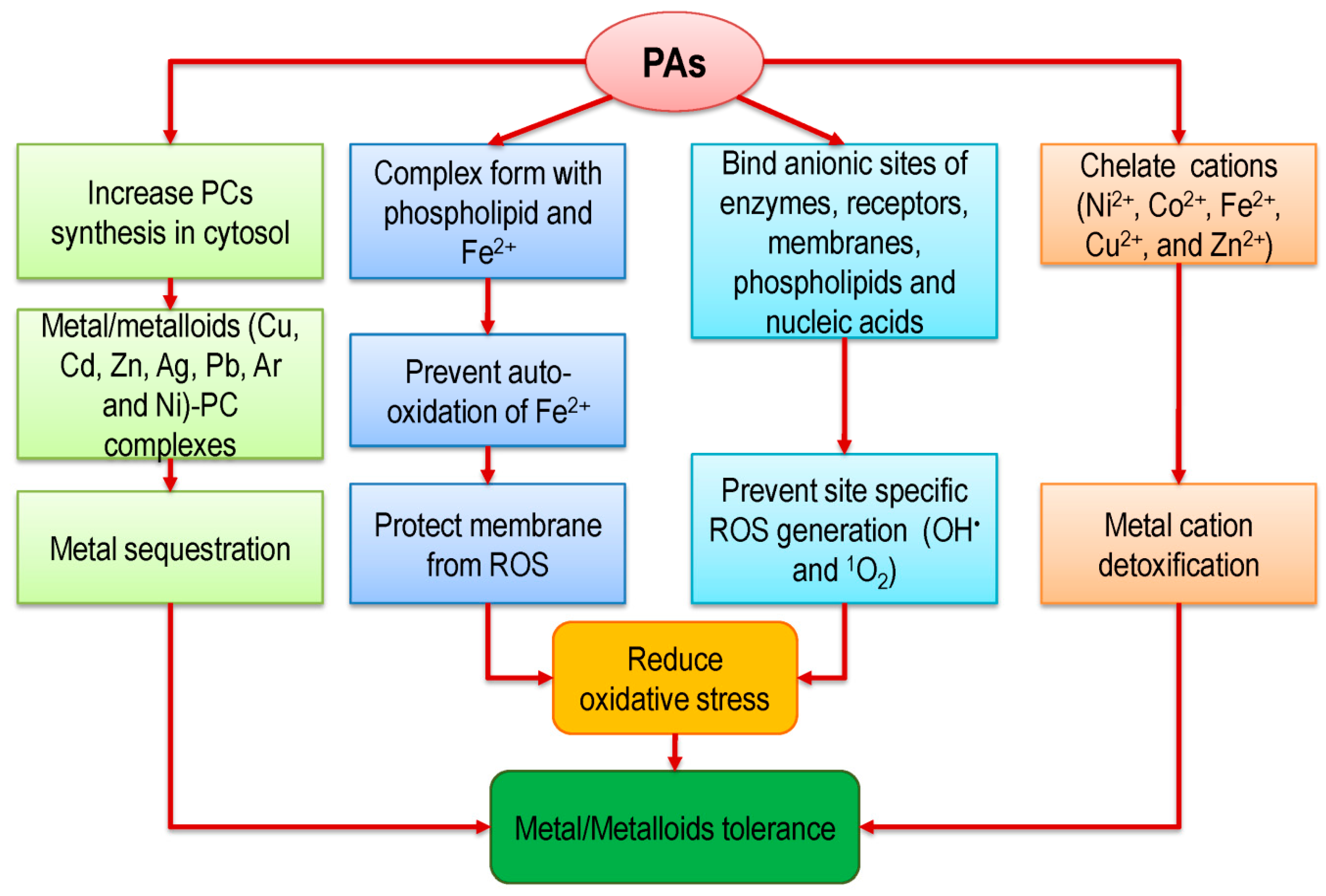
5. Polyamine-Induced Metal/Metalloid(s) Chelation and Phytoremediation in Plants
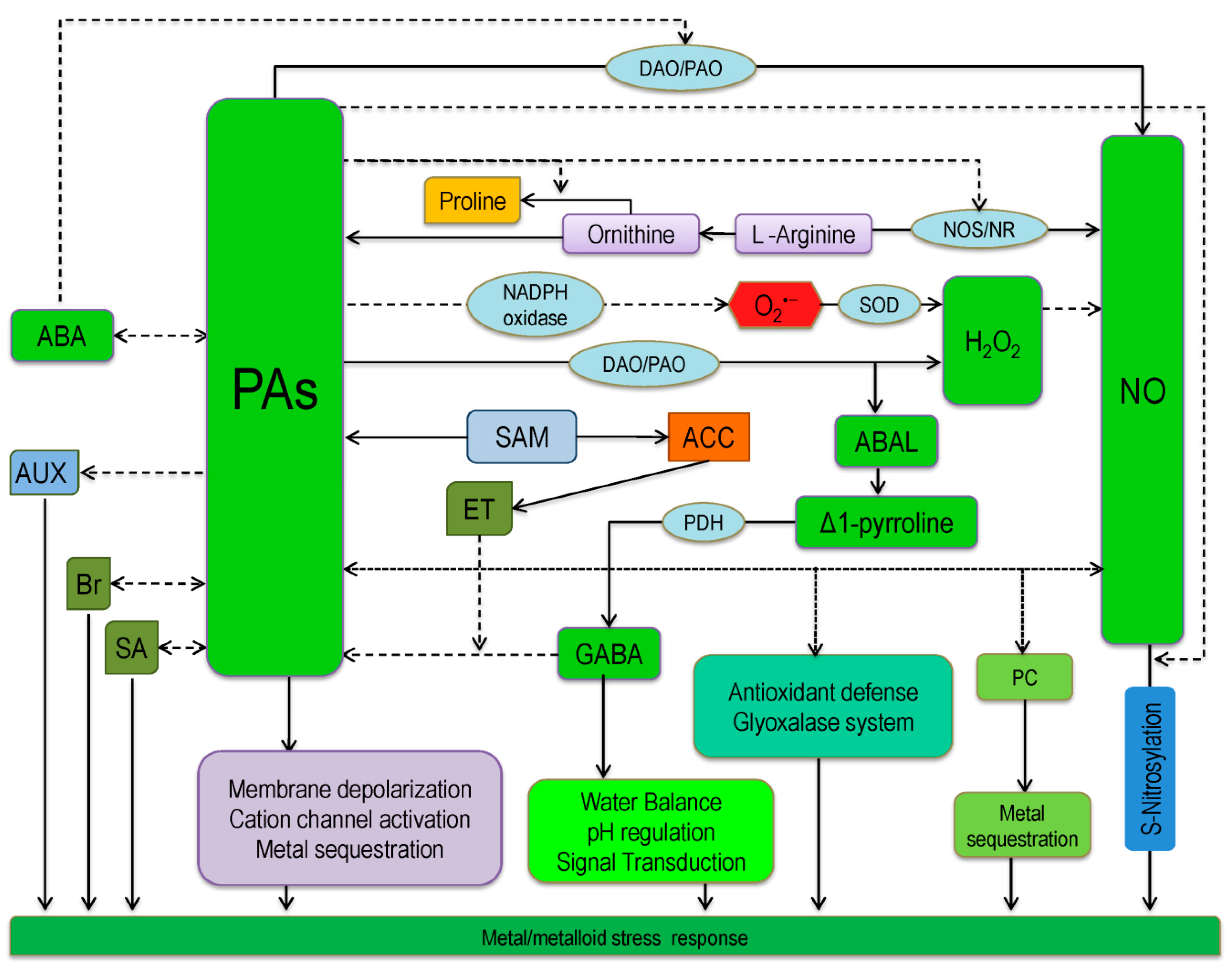
6. Interaction of Polyamines with other Molecules in Conferring Metal/Metalloids Tolerance in Plants
6.1. Nitric Oxide and Polyamines Cross-talk and the Reversal of Metal Phytotoxicity
6.2. Reactive Oxygen Species and Polyamines Interaction and the Reversal of Metal Phytotoxicity
6.3. Interaction of Polyamines with GABA and Pro Conferring Metal/Metalloid(s) Stress
6.4. Interaction of Plant Hormones and Other Hormone-Like Protective Molecules with Polyamines
6.5. Polyamine Interacts with Ion Channels Modulate Metal/Metalloid(S) Stress Tolerance
7. Omics Approaches to Improve Polyamines Actions towards Metals/Metalloid(s) Action
7.1. Transcriptomics
7.2. Proteomics
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAO | Ascorbic acid oxidase |
| ACC synthase | 1-aminocyclopropane-1-carboxylic-acid synthase |
| ACS | 1-aminocyclopropane-1-carboxylic-acid synthase |
| ACC | 1-aminocyclopropane-1-carboxylic-acid |
| ABAL | 4-aminobutanal |
| ABA | Abscisic acid |
| ATP | Adenosine triphosphate |
| AIH | Agmatin iminohydrolase |
| ADC | Arginine decarboxylase |
| APX | Ascorbate peroxidase |
| AsA | Ascorbate |
| AUX | Auxins |
| BAC | Biological accumulation coefficient |
| BCF | Biological concentration factor |
| Br | Brassinosteroids |
| CAT | Catalase |
| CW-PAO | Cell wall-PAO |
| CK | Cytokinins |
| dcSAM | Decarboxylated S-adenosylmethionine |
| DHAR | Dehydroascorbate reductase |
| DAO | Diamine oxidase |
| EL | Electrolyte leakage |
| EBL | Epibrassinolide |
| ET | Ethylene |
| GA | Gibberellins |
| GABA | γ-aminobutyric acid |
| GPX | Glutathione peroxidase |
| GR | Glutathione reductase |
| GST | Glutathione S-transferase |
| GSH | Glutathione |
| GB | Glycine betaine |
| Gly I | Glyoxalase I |
| Gly II | Glyoxalase II |
| IAA | Indole acetic acid |
| JA | Jasmonic acids |
| LOX | Lipoxygenase |
| MDA | Malondialdehyde |
| MSI | Membrane stability index |
| MTs | Metallothioneins |
| MG | Methylglyoxal |
| MAPKs | Mitogen-activated protein kinases |
| MDHAR | Monodehydroascorbate reductase |
| CPA | N-carbamoylputrescine amidohydrolase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NR | Nitrate reductase |
| NOS | NO synthase |
| OTC | Ornithine transcarbamoylase |
| δ-OAT | Ornithine δ-aminotransferase |
| ODC | Ortinine decarboxylase |
| PC | Phytochelatins |
| PAs | Polyamines |
| PPO | Polyphenol oxidase |
| Pro | Proline |
| Put | Putrescine |
| PDH | Pyrroline dehydrogenase |
| ROS | Reactive oxygen species |
| RWC | Relative water content |
| SAMDC | S-adenosylmethionine decarboxylase |
| SAM | S-adenosylmethionine |
| SA | Salicylic acid |
| SNP | Sodium nitroprusside |
| SPDS | Spermidine synthase |
| Spd | Spermidine |
| SPMS | Spermine synthase |
| Spm | Spermine |
| SOD | Superoxide dismutase |
| TBARS | Thiobarbituric acid |
| TF | Translocation factor |
References
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Basu, S.; Kumar, G. Polyamines metabolism: A way ahead for abiotic stress tolerance in crop plants. In Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Wani, S.H., Ed.; Academic Press: London, UK, 2018; pp. 39–55. [Google Scholar]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef] [PubMed]
- Minois, N. Molecular basis of the ‘anti-aging’ effect of spermidine and other natural polyamines—A mini-review. Gerontology 2014, 60, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Benavides, M.P.; Groppa, M.D.; Recalde, L.; Verstraeten, S.V. Effects of polyamines on cadmium-and copper-mediated alterations in wheat (Triticum aestivum L.) and sunflower (Helianthus annuus L.) seedling membrane fluidity. Arch. Biochem. Biophys. 2018, 654, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Groppa, M.D.; Benavides, M.P.; Tomaro, M.L. Polyamine metabolism in sunflower and wheat leaf discs under cadmium or copper stress. Plant Sci. 2003, 164, 293–299. [Google Scholar] [CrossRef]
- Fahad, S.; Rehman, A.; Shahzad, B.; Tanveer, M.; Saud, S.; Kamran, M.; Ihtisham, M.; Khan, S.U.; Turan, V.V.; ur Rahman, M.H. Rice responses and tolerance to metal/metalloid toxicity. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Eds.; Woodhead Publishing: London, UK, 2019; pp. 299–312. [Google Scholar]
- Paul, S.; Banerjee, A.; Roychoudhury, A. Role of polyamines in mediating antioxidant defense and epigenetic regulation in plants exposed to heavy metal toxicity. In Plants Under Metal and Metalloid Stress; Hasanuzzaman, M., Nahar, K., Fujita, M., Eds.; Eds.; Springer: Singapore, 2018; pp. 229–247. [Google Scholar]
- Tajti, J.; Janda, T.; Majláth, E.; Szalai, G.; Pál, M. Comparative study on the effects of putrescine and spermidine pre-treatment on cadmium stress in wheat. Ecotoxicol. Environ. Saf. 2018, 148, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Liu, Y.; Huang, D.; Zeng, G.; Liu, S.; Tang, H.; Zhou, L.; Hu, X.; Zhou, Y.; Tan, X. Effects of exogenous calcium and spermidine on cadmium stress moderation and metal accumulation in Boehmeria nivea (L.) Gaudich. Environ. Sci. Pollut. Res. 2016, 23, 8699–8708. [Google Scholar] [CrossRef]
- Rady, M.M.; Hemida, K.A. Modulation of cadmium toxicity and enhancing cadmium-tolerance in wheat seedlings by exogenous application of polyamines. Ecotoxicol. Environ. Saf. 2015, 119, 178–185. [Google Scholar] [CrossRef]
- Taie, H.A.; El-Yazal, M.A.S.; Ahmed, S.M.; Rady, M.M. Polyamines modulate growth, antioxidant activity, and genomic DNA in heavy metal–stressed wheat plant. Environ. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef]
- Alcázar, R.; Cuevas, J.C.; Patrón, M.; Altabella, T.; Tiburcio, A.F. Abscisic acid modulates polyamine metabolism under water stress in Arabidopsis thaliana. Physiol. Plant 2006, 128, 448–455. [Google Scholar] [CrossRef]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Arabidopsis polyamine biosynthesis: Absence of ornithine decarboxylase and the mechanism of arginine decarboxylase activity. Plant J. 2001, 27, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; García-Martínez, J.L.; Cuevas, J.C.; Tiburcio, A.F.; Altabella, T. Overexpression of ADC2 in Arabidopsis induces dwarfism and late-flowering through GA deficiency. Plant J. 2005, 43, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Hanzawa, Y.; Takahashi, T.; Michael, A.J.; Burtin, D.; Long, D.; Pineiro, M.; Coupland, G.; Komeda, Y. ACAULIS5, an Arabidopsis gene required for stem elongation, encodes a spermine synthase. EMBO J. 2000, 19, 4248–4256. [Google Scholar] [CrossRef] [PubMed]
- Panicot, M.; Minguet, E.G.; Ferrando, A.; Alca´zar, R.; Bla´zquez, M.A.; Carbonell, J.; Altabella, T.; Koncz, C.; Tiburcio, A.F. A polyamine metabolon involving aminopropyl transferases complexes in Arabidopsis. Plant Cell 2002, 14, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Ito, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Arabidopsis stress-inducible gene for arginine decarboxylase AtADC2 is required for accumulation of putrescine in salt tolerance. Biochem. Biophys. Res. Comm. 2004, 313, 369–375. [Google Scholar] [CrossRef]
- Kakkar, R.K.; Sawhney, V.K. Polyamine research in plants. A changing perspective. Physiol. Plant 2002, 116, 281–292. [Google Scholar] [CrossRef]
- Majumdar, R.; Shao, L.; Minocha, R.; Long, S.; Minocha, S.C. Ornithine: The overlooked molecule in regulation of polyamine metabolism. Plant Cell Physiol. 2013, 54, 990–1004. [Google Scholar] [CrossRef]
- Kuthanová, A.; Gemperlová, L.; Zelenková, S.; Eder, J.; Machácková, I.; Opatrny, Z.; Cvikrová, M. Cytological changes and alterations in polyamine contents induced by cadmium in tobacco BY-2 cells. Plant Physiol. Biochem. 2004, 42, 149–156. [Google Scholar] [CrossRef]
- Choudhary, A.; Singh, R.P. Cadmium-induced changes in diamine oxidase activity and polyamine levels in Vigna radiata Wilczek seedlings. J. Plant Physiol. 2000, 156, 704–710. [Google Scholar] [CrossRef]
- Chen, L.; Wang, L.; Chen, F.; Korpelainen, H.; Li, C. The effects of exogenous putrescine on sex-specific responses of Populus cathayana to copper stress. Ecotoxicol. Environ. Saf. 2013, 97, 94–102. [Google Scholar] [CrossRef]
- Podlešáková, K.; Ugena, L.; Spíchal, L.; Doležal, K.; De Diego, N. Phytohormones and polyamines regulate plant stress responses by altering GABA pathway. New Biotechnol. 2019, 48, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Regulatory role of polyamines in growth, development and abiotic stress tolerance in plants. In Plant Adaptation to Environmental Change: Significance of Amino Acids and their Derivatives; Anjum, N.A., Gill, S.S., Gill, R., Eds.; CABI: Wallingford, UK, 2014; pp. 157–193. [Google Scholar]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Balal, R.M.; Khan, N.; Rossi, L.; Rathinasabapathi, B.; Liu, G.; Khan, J.; Cámara-Zapata, J.M.; Martínez-Nicolas, J.J.; Garcia-Sanchez, F. Polyamines provide new insights into the biochemical basis of Cr-tolerance in Kinnow mandarin grafted on diploid and double-diploid rootstocks. Environ. Exp. Bot. 2018, 156, 248–260. [Google Scholar] [CrossRef]
- Rady, M.M.; Ahmed, S.M.; El-Yazal, M.A.S.; Taie, H.A. Alleviation of cadmium stress in wheat by polyamines. In Cadmium Tolerance in Plants: Agronomic, Molecular, Signaling, and Omic Approaches; Hasanuzzaman, M., Prasad, M., Nahar, K., Eds.; Elsevier: London, UK, 2019; pp. 463–496. [Google Scholar]
- Yu, Y.; Zhou, W.; Zhou, K.; Liu, W.; Liang, X.; Chen, Y.; Dasheng, S.; Xianyong, L. Polyamines modulate aluminum-induced oxidative stress differently by inducing or reducing H2O2 production in wheat. Chemosphere 2018, 212, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Howladar, S.M.; Al-Robai, S.A.; Al-Zahrani, F.S.; Howladar, M.M.; Aldhebiani, A.Y. Silicon and its application method effects on modulation of cadmium stress responses in Triticum aestivum (L.) through improving the antioxidative defense system and polyamine gene expression. Ecotoxicol. Environ. Saf. 2018, 159, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.M.; El-Yazal, M.A.S.; Taie, H.A.A.; Ahmed, S.M. Response of wheat growth and productivity to exogenous polyamines under lead stress. J. Crop Sci. Biotechnol. 2016, 19, 363–371. [Google Scholar] [CrossRef]
- Mir, B.A.; Khan, T.A.; Fariduddin, Q. 24-epibrassinolide and spermidine modulate photosynthesis and antioxidant systems in Vigna radiata under salt and zinc stress. Int. J. Adv. Res. 2015, 3, 592–608. [Google Scholar]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Zambryzycka, E.; Godlewska-Zylkiewicz, B. Phytohormones as regulators of heavy metal biosorption and toxicity in green alga Chlorella vulgaris (Chlorophyceae). Plant Physiol. Biochem. 2012, 52, 52–65. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Kanwar, M.; Bhardwaj, R.; Yu, J.Q.; Tran, L.S.P. Chromium stress mitigation by polyamine-brassinosteroid application involves phytohormonal and physiological strategies in Raphanus sativus L. PLoS ONE 2012, 7, e33210. [Google Scholar] [CrossRef]
- Tang, C.; Zhang, R.; Hu, X.; Song, J.; Li, B.; Ou, D.; Hu, X.; Zhao, Y. Exogenous spermidine elevating cadmium tolerance in Salix matsudana involves cadmium detoxification and antioxidant defense. Int. J. Phytorem. 2019, 21, 305–315. [Google Scholar] [CrossRef]
- Yang, H.; Shi, G.; Wang, H.; Xu, Q. Involvement of polyamines in adaptation of Potamogeton crispus L. to cadmium stress. Aquat. Toxicol. 2010, 100, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Groppa, M.D.; Tomaro, M.L.; Benavides, M.P. Polyamines as protectors against cadmium or copper-induced oxidative damage in sunflower leaf discs. Plant Sci. 2001, 161, 481–488. [Google Scholar] [CrossRef]
- Hsu, Y.T.; Kao, C.H. Cadmium-induced oxidative damage in rice leaves is reduced by polyamines. Plant Soil 2007, 291, 27–37. [Google Scholar] [CrossRef]
- Groppa, M.D.; Tomaro, M.L.; Benavides, M.P. Polyamines and heavy metal stress: The antioxidant behavior of spermine in cadmium and copper-treated wheat leaves. Biometals 2007, 20, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Hasanuzzaman, M.; Hossain, M.A.; Thangavel, P.; Roychoudhury, A.; Gill, S.S.; Rodrigo, M.A.M.; Adam, V.; Fujita, M.; Kizek, R.; et al. Jacks of metal/metalloid chelation trade in plants—an overview. Front. Plant Sci. 2015, 6, 192. [Google Scholar] [CrossRef] [PubMed]
- Amist, N.; Singh, N.B. Responses of enzymes involved in proline biosynthesis and degradation in wheat seedlings under stress. Allelopathy J. 2017, 42, 195–205. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, L.Y.; Guo, S.R.; Sun, J.; Liu, C.J. Effects of exogenous spermidine on photosynthesis, xanthophyll cycle and endogenous polyamines in cucumber seedlings exposed to salinity. Afr. J. Biotechnol. 2012, 11, 6064–6074. [Google Scholar]
- Balal, R.M.; Shahid, M.A.; Javaid, M.M.; Iqbal, Z.; Liu, G.D.; Zotarelli, L.; Khan, N. Chitosan alleviates phytotoxicity caused by boron through augmented polyamine metabolism and antioxidant activities and reduced boron concentration in Cucumis sativus L. Acta Physiol. Plant 2016, 39, 31–36. [Google Scholar] [CrossRef]
- Kumar, M.; Kuzhiumparambil, U.; Ralph, P.; Contreras-Porcia, L. Polyamines: Stress metabolite in marine macrophytes. In Algal Green Chemistry, Recent Progress in Biotechnology; Rastogi, R.P., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 243–255. [Google Scholar]
- Ghabriche, R.; Ghnaya, T.; Zaier, H.; Baioui, R.; Vromman, D.; Abdelly, C.; Lutts, S. Polyamine and tyramine involvement in NaCl-induced improvement of Cd resistance in the halophyte Inula chrithmoides L. J. Plant Physiol. 2017, 216, 136–144. [Google Scholar] [CrossRef]
- Baniasadi, F.; Saffari, V.R.; Moud, A.A.M. Physiological and growth responses of Calendula officinalis L. plants to the interaction effects of polyamines and salt stress. Sci. Hortic. 2018, 234, 312–317. [Google Scholar] [CrossRef]
- Hassan, F.A.S.; Ali, E.F.; Alamer, K.H. Exogenous application of polyamines alleviates water stress-induced oxidative stress of Rosa damascena Miller var. Trigintipetala Dieck. South Afr. J. Bot. 2018, 116, 96–102. [Google Scholar] [CrossRef]
- Groppa, M.D.; Benavides, M.P. Polyamines and abiotic stress: Recent advances. Amino Acids 2008, 34, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Løvaas, E. Antioxidative and metal-chelating effects of polyamines. Adv. Pharmacol. 1996, 38, 119–149. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of endogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Romero, L.; Ruiz, J.M. Accumulation of free polyamines enhances the antioxidant response in fruits of grafted tomato plants under water stress. J. Plant Physiol. 2016, 190, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2001, 212, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.; Aarts, M.G.M. Opportunities and feasibilities for biotechnological improvement of Zn, Cd or Ni tolerance and accumulation in plants. Environ. Exp. Bot. 2011, 72, 53–63. [Google Scholar] [CrossRef]
- Mahmud, J.A.; Hasanuzzaman, M.; Nahar, K.; Bhuyan, M.B.; Fujita, M. Insights into citric acid-induced cadmium tolerance and phytoremediation in Brassica juncea L.: Coordinated functions of metal chelation, antioxidant defense and glyoxalase systems. Ecotoxicol. Environ. Saf. 2018, 147, 990–1001. [Google Scholar] [CrossRef]
- Lv, Y.; Deng, X.; Quan, L.; Xia, Y.; Shen, Z. Metallothioneins BcMT1 and BcMT2 from Brassica campestris enhance tolerance to cadmium and copper and decrease production of reactive oxygen species in Arabidopsis thaliana. Plant Soil 2013, 367, 507–519. [Google Scholar] [CrossRef]
- Wen, X.P.; Ban, Y.; Inoue, H.; Matsuda, N.; Moriguchi, T. Spermidine levels are implicated in heavy metal tolerance in a spermidine synthase overexpressing transgenic European pear by exerting antioxidant activities. Transgenic Res. 2010, 19, 91–103. [Google Scholar] [CrossRef]
- Flink, L.; Pettijohn, D.E. Polyamines stabilize DNA folds. Nature 1975, 253, 62–63. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Csávás, G.; Szalai, G.; Oláh, T.; Khalil, R.; Yordanova, R.; Gell, G.; Birinyi, Z.; Németh, E.; Janda, T. Polyamines may influence phytochelatin synthesis during Cd stress in rice. J. Hazard. Mater. 2017, 340, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Abhilash, P.C.; Powell, J.R.; Singh, H.B.; Singh, B.K. Plant–microbe interactions: Novel applications for exploitation in multipurpose remediation technologies. Trends Biotechnol. 2012, 30, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Keymirov, M.A. Water Purification of Ions of Heavy Metals by Montmorillonite Modified with Polyamine. J. Water Chem. Technol. 2018, 40, 320–326. [Google Scholar] [CrossRef]
- Nahar, K.; Rahman, M.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Physiological and biochemical mechanisms of spermine-induced cadmium stress tolerance in mung bean (Vigna radiata L.) seedlings. Environ Sci. Pollut. Res. 2016, 23, 21206–21218. [Google Scholar] [CrossRef] [PubMed]
- Shevyakova, N.I.; Il’ina, E.N.; Kuznetsov, V.V. Polyamines increase plant potential for phytoremediation of soils polluted with heavy metals. Doklady Biol. Sci. 2008, 423, 457–460. [Google Scholar] [CrossRef]
- Soudek, P.; Ursu, M.; Petrová, S.; Vaněk, T. Improving crop tolerance to heavy metalstress by polyamine application. Food Chem. 2016, 213, 223–229. [Google Scholar] [CrossRef]
- Guo, Z.; Tan, J.; Zhuo, C.; Wang, C.; Xiang, B.; Wang, Z. Abscisic acid H2O2 and nitric oxide interactions mediated cold-induced S-adenosyl methionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. 2014, 12, 601–612. [Google Scholar] [CrossRef]
- Gao, H.J.; Yang, H.Q.; Wang, J.X. Arginine metabolism in roots and leaves of apple (Malus domestica Borkh.): The tissue-specific formation of both nitric oxide and polyamines. Sci. Hortic. 2009, 119, 147–152. [Google Scholar] [CrossRef]
- Shi, H.; Chan, Z. Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J. Integr. Plant Biol. 2014, 56, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.S.; Scherer, G.F. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Wimalasekera, R.; Tebartz, F.; Scherer, G.F. Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011, 181, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Saha, J.; Brauer, E.K.; Sengupta, A.; Popescu, S.C.; Gupta, K.; Gupta, B. Polyamines as redox homeostasis regulators during salt stress in plants. Front. Environ. Sci. 2015, 3, 21. [Google Scholar] [CrossRef]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Fuglsang, A.T.; Shabala, S. Polyamines cause plasma membrane depolarization, activate Ca2+-, and modulate H+-ATPase pump activity in pea roots. J. Exp. Bot. 2014, 65, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Moschou, P.N.; Paschalidis, K.A.; Delis, I.D.; Andriopoulou, A.H.; Lagiotis, G.D.; Yakoumakis, D.I.; Roubelakis-Angelakis, K.A. Spermidine exodus and oxidation in the apoplast induced by abiotic stress is responsible for H2O2 signatures that direct tolerance responses in tobacco. Plant Cell 2008, 20, 1708–1724. [Google Scholar] [CrossRef] [PubMed]
- Andronis, E.A.; Moschou, P.N.; Toumi, I.; Roubelakis-Angelakis, K.A. Peroxisomal polyamine oxidase and NADPH-oxidase cross-talk for ROS homeostasis which affects respiration rate in Arabidopsis thaliana. Front. Plant Sci. 2014, 5, 132. [Google Scholar] [CrossRef] [PubMed]
- Kabała, K.; Zboińska, M.; Głowiak, D.; Reda, M.; Jakubowska, D.; Janicka, M. Interaction between the signaling molecules hydrogen sulfide and hydrogen peroxide and their role in vacuolar H+-ATPase regulation in cadmium-stressed cucumber roots. Physiol. Plant 2018. [Google Scholar] [CrossRef]
- Yang, R.; Guo, Q.; Gu, Z. GABA shunt and polyamine degradation pathway on γ-aminobutyric acid accumulation in germinating fava bean (Vicia faba L.) under hypoxia. Food Chem. 2013, 136, 152–159. [Google Scholar] [CrossRef]
- Hatmi, S.; Gruau, C.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Baillieul, F.; Eullaffroy, P.; Clément, C.; Ferchichi, A.; Aziz, A. Drought stress tolerance in grapevine involves activation of polyamine oxidation contributing to improved immune response and low susceptibility to Botrytis cinerea. J. Exp. Bot. 2015, 66, 775–787. [Google Scholar] [CrossRef]
- Al-Quraan, N.A. GABA shunt deficiencies and accumulation of reactive oxygen species under UV treatments: Insight from Arabidopsis thaliana calmodulin mutants. Acta Physiol. Plant 2015, 37, 1–11. [Google Scholar] [CrossRef]
- Mahmud, J.A.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Hossain, M.S.; Fujita, M. γ-aminobutyric acid (GABA) confers chromium stress tolerance in Brassica juncea L. by modulating the antioxidant defense and glyoxalase systems. Ecotoxicology 2017, 26, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Xu, X.; Wang, H.; Wang, H.; Tao, Y. Exogenous γ-aminobutyric acid alleviates oxidative damage caused by aluminium and proton stresses on barley seedlings. J. Sci. Food Agric. 2010, 90, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.Q.; Shi, Z.; Jiang, Z.P.; Qi, L.W.; Sun, X.M.; Li, C.X.; Liu, J.F.; Xiao, W.F.; Zhang, S.G. Effects of exogenous GABA on gene expression of Caragana intermedia roots under NaCl stress: Regulatory roles for H2O2 and ethylene production. Plant Cell Environ 2010, 33, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fan, L.; Gao, H.; Wu, X.; Li, J.; Lv, G.; Gong, B. Polyamine biosynthesis and degradation are modulated by exogenous gamma-aminobutyric acid in root-zone hypoxia-stressed melon roots. Plant Physiol. Biochem. 2014, 82, 17–26. [Google Scholar] [CrossRef]
- Hu, X.; Xu, Z.; Xu, W.; Li, J.; Zhao, N.; Zhou, Y. Application of γ-amino butyric acid demonstrates a protective role of polyamine and GABA metabolism in muskmelon seedlings under Ca (NO3)2 stress. Plant Physiol. Biochem. 2015, 92, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Becker, D.F. Connecting proline metabolism and signaling pathways in plant senescence. Front. Plant Sci. 2015, 6, 552. [Google Scholar] [CrossRef] [Green Version]
- Seifikalhor, M.; Aliniaeifard, S.; Hassani, B.; Niknam, V.; Lastochkina, O. Diverse role of γ-aminobutyric acid in dynamic plant cell responses. Plant Cell Rep. 2019. [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Suzuki, T.; Fujita, M. Polyamines-induced aluminum tolerance in mung bean: A study on antioxidant defense and methylglyoxal detoxification systems. Ecotoxicology 2017, 26, 58–73. [Google Scholar] [CrossRef]
- Cvikrová, M.; Gemperlová, L.; Martincová, O.; Vanková, R. Effect of drought and combined drought and heat stress on polyamine metabolism in proline-over-producing tobacco plants. Plant Physiol. Biochem. 2013, 73, 7–15. [Google Scholar] [CrossRef]
- Agami, R.A. Pre-soaking in indole-3-acetic acid or spermidine enhances copper tolerance in wheat seedlings. South. Afr. J. Bot. 2016, 104, 167–174. [Google Scholar] [CrossRef]
- Cui, X.; Ge, C.; Wang, R.; Wang, H.; Chen, W.; Fu, Z.; Jiang, X.; Li, J.; Wang, Y. The BUD2 mutation affects plant architecture through altering cytokinin and auxin responses in Arabidopsis. Cell Res. 2010, 20, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, S.; Liu, P.; Wang, W.; Cao, D.; Deng, X.; Zhang, S. Silicon-mediated changes in polyamine and 1-aminocyclopropane-1-carboxylic acid are involved in silicon-induced drought resistance in Sorghum bicolor L. Plant Physiol. Biochem. 2014, 80, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Cong, R.; Sagor, G.H.M.; Niitsu, M.; Berberich, T.; Kusano, T. Characterization of five polyamine oxidase isoforms in Arabidopsis thaliana. Plant Cell Rep. 2010, 29, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjunatha, G.; Gupta, K.J.; Lokesh, V.; Mur, L.A.; Neelwarne, B. Nitric oxide counters ethylene effects on ripening fruits. Plant Signal. Behav. 2012, 7, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Verma, S.; Vyas, D.; Per, T.S.; Masood, A.; Khan, N.A. Ethylene and polyamines in counteracting heavy metal phytotoxicity: A crosstalk perspective. J. Plant Growth Regul. 2018, 37, 1050–1065. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Lefèvre, I.; Lutts, S.; Deckert, J. Short term signaling responses in roots of young soybean seedlings exposed to cadmium stress. J. Plant Physiol. 2013, 170, 1585–1594. [Google Scholar] [CrossRef]
- Alet, A.I.; Sanchez, D.H.; Cuevas, J.C.; del Valle, S.; Altabella, T.; Tiburcio, A.F.; Marco, F.; Ferrando, A.; Espasandín, F.D.; González, M.E.; et al. Putrescine accumulation in Arabidopsis thaliana transgenic lines enhances tolerance to dehydration and freezing stress. Plant Signal. Behav. 2011, 6, 278–286. [Google Scholar] [CrossRef]
- Espasandin, F.D.; Maiale, S.J.; Calzadilla, P.; Ruiz, O.A.; Sansberro, P.A. Transcriptional regulation of 9-cis-epoxycarotenoid dioxygenase (NCED) gene by putrescine accumulation positively modulates ABA synthesis and drought tolerance in Lotus tenuis plants. Plant Physiol. Biochem. 2014, 76, 29–35. [Google Scholar] [CrossRef]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine is involved in Arabidopsis freezing tolerance and cold acclimation by regulating abscisic acid levels in response to low temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Keunen, E.; Schellingen, K.; Vangronsveld, J.; Cuypers, A. Ethylene and metal stress: Small molecule, big impact. Front. Plant Sci. 2016, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Szepesi, Á.; Gémes, K.; Orosz, G.; Peto, A.; Takács, Z.; Vorák, M.; Tari, I. Interaction between salicylic acid and polyamines and their possible roles in tomato hardening processes. Acta Biol. Szegediensis 2011, 55, 165–166. [Google Scholar]
- Baronas, V.A.; Kurata, H.T. Inward rectifiers and their regulation by endogenous polyamines. Front. Physiol. 2014, 5, 325. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Kim, D.W.; Niitsu, M.; Berberich, T.; Kusano, T. Oryza sativa polyamine oxidase 1 back-converts tetraamines, spermine and thermospermine, to spermidine. Plant Cell Rep. 2014, 33, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.P.; Pang, X.M.; Matsuda, N.; Kita, M.; Inoue, H.; Hao, Y.J.; Honda, C.; Moriguchi, T. Over-expression of the apple spermidine synthase gene in pear confers multiple abiotic stress tolerance by altering polyamine titers. Transgenic Res. 2008, 17, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.P.; Ban, Y.; Pang, X.M.; Moriguchi, T. Identification of differentially-expressed genes potentially related to stress tolerance in a transgenic line of European pear over-expressing an apple spermidine synthase gene (MdSPDS1). J. Hort. Sci. Biotechnol. 2011, 86, 146–152. [Google Scholar] [CrossRef]
- Prabhavathi, V.R.; Rajam, M.V. Polyamine accumulation in transgenic eggplant enhances tolerance to multiple abiotic stresses and fungal resistance. Plant Biotechnol. 2007, 24, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.P.; Ban, Y.; Inoue, H.; Matsuda, N.; Moriguchi, T. Aluminum tolerance in a spermidine synthase-overexpressing transgenic European pear is correlated with the enhanced level of spermidine via alleviating oxidative status. Environ. Exp. Bot. 2009, 66, 471–478. [Google Scholar] [CrossRef]
- Sunkar, R.; Bartels, D.; Kirch, H.H. Overexpression of a stress-inducible aldehyde dehydrogenase gene from Arabidopsis thaliana in transgenic plants improves stress tolerance. Plant J. 2003, 35, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Kitashiba, H.; Wang, J.; Ban, Y.; Moriguchi, T. Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol. 2007, 24, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, P.P.; Chen, C.L.; Wang, Y.; Fu, X.Z.; Liu, J.H. An arginine decarboxylase gene PtADC from Poncirus trifoliata confers abiotic stress tolerance and promotes primary root growth in Arabidopsis. J. Exp. Bot. 2011, 62, 2899–2914. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Ban, Y.; Wen, X.P.; Nakajima, I.; Moriguchi, T. Molecular cloning and expression analysis of an arginine decarboxylase gene from peach (Prunus persica). Gene 2009, 429, 10–17. [Google Scholar] [CrossRef]
- Das, K.C.; Mishra, H.P. Hydroxyl radical scavenging and singlet oxygen quenching properties of polyamines. Mol. Cell. Biochem. 2004, 262, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.V.; Sharma, M.L.; Rajam, M.V. Polyamine biosynthetic pathway as a novel target for potential applications in plant biotechnology. Physiol. Mol. Biol. Plants 2006, 12, 13–28. [Google Scholar]
- You, J.; Hu, H.; Xiong, L. An ornithine δ-aminotransferase gene OsOAT confers drought and oxidative stress tolerance in rice. Plant Sci. 2012, 197, 59–69. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Trivedi, P.K.; Misra, P.; Tiwari, M.; Shri, M.; Shukla, D.; Kumar, S.; Rai, A.; Pandey, A.; Nigam, D.; et al. Comparative transcriptome analysis of arsenate and arsenite stresses in rice seedlings. Chemosphere 2009, 74, 688–702. [Google Scholar] [CrossRef]
- Tripathi, R.D.; Tripathi, P.; Dwivedi, S.; Dubey, S.; Chakrabarty, D. Arsenomics: Omics of arsenic metabolism in plants. Front. Physiol. 2012, 3, 275. [Google Scholar] [CrossRef]
- Rose, J.C.; Bashir, S.; Giovannoni, J.J.; Jahn, M.M.; Saravanan, R.S. Tackling the plant proteome: Practical approaches, hurdles and experimental tools. Plant J. 2004, 39, 715–733. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.A.; Ahmad, J.; Bagheri, R.; Ali, A.A.; Al-Huqail, A.A.; Ibrahim, M.M.; Qureshi, M.I. Proteomic and ecophysiological responses of soybean (Glycine max L.) root nodules to Pb and hg stress. BMC Plant Biol. 2018, 18, 283. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, B.C.; Xu, Y.; Zhu, Y.X. Systematic studies of 12S seed storage protein accumulation and degradation patterns during Arabidopsis seed maturation and early seedling germination stages. J. Biochem. Mol. Biol. 2007, 40, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ge, Q.; Zhang, J.; Zhou, J.; Xu, J. Proteomic analysis of Cd-responsive proteins in Solanum torvum. Plant Mol. Biol. Rep. 2013, 31, 485–491. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.G.; Alam, I.; Kim, P.J.; Lee, J.J.; Ahn, Y.O.; Kwak, S.S.; Lee, I.J.; Bahk, J.D.; Kang, K.Y.; et al. Comparative proteomic study of arsenic-induced differentially expressed proteins in rice roots reveals glutathione plays a central role during As stress. Proteomics 2008, 8, 3561–3576. [Google Scholar] [CrossRef] [PubMed]
- Aloui, A.; Recorbet, G.; Gollotte, A.; Robert, F.; Valot, B.; Gianinazzi-Pearson, V.; Aschi-Smiti, S.; Dumas-Gaudot, E. On the mechanisms of cadmium stress alleviation in Medicago truncatula by arbuscular mycorrhizal symbiosis: A root proteomic study. Proteomics 2009, 9, 420–433. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, X.; Yang, S.; Zhou, Y.; Dong, C.; Ren, J.; Sun, X.; Yang, Y. Comparative physiological and proteomic analysis reveals the leaf response to cadmium-induced stress in poplar (Populus yunnanensis). PLoS ONE 2015, 10, e0137396. [Google Scholar] [CrossRef]
- Naka, Y.; Watanabe, K.; Sagor, G.H.M.; Niitsu, M.; Pillai, M.A.; Kusano, T.; Takahashi, Y. Quantitative analysis of plant polyamines including thermospermine during growth and salinity stress. Plant Physiol. Biochem. 2010, 48, 527–533. [Google Scholar] [CrossRef]
- Brumbarova, T.; Matros, A.; Mock, H.P.; Bauer, P. A proteomic study showing differential regulation of stress, redox regulation and peroxidase proteins by iron supply and the transcription factor FER. Plant J. 2008, 54, 321–334. [Google Scholar] [CrossRef]
- Kim, Y.K.; Lee, M.Y. Proteomic analysis of differentially expressed proteins of rice in response to cadmium. J. Korean Soc. App.l Biol. Chem. 2009, 52, 428–436. [Google Scholar] [CrossRef]


{kind=link}
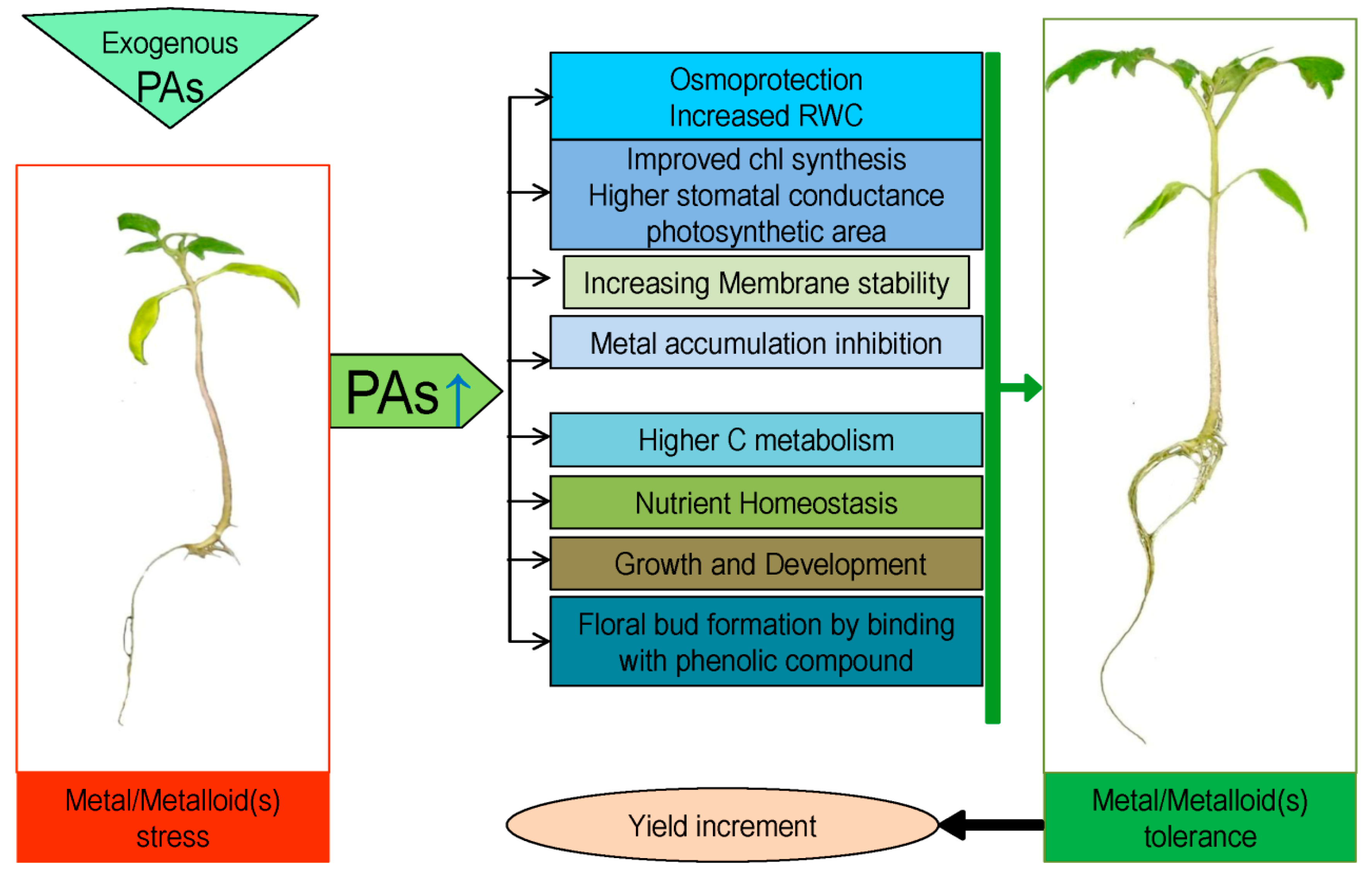{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Metal(s) Exposed | Changes in Endogenous PAs Level | Exogenous PAs Used | Tolerance | References |
|---|---|---|---|---|---|
| Triticum aestivum L. | 1 mM CdCl2 for 5–15 d | - | 0.1 mM Put, Spd or Spm pretreatment for 5 and 10 d |
| Benavides et al. [5] |
| T. aestivum | 2 mM Cd as CdCl2 | - | 0.25 mM Spm, 0.50 mM Spd, and 1 mM Put as seed priming or as a foliar spray at 20, 40, and 60 DAS |
| Taie et al. [12] |
| T. aestivum | 2 mM as PbCl2 for 45 d | - | 0.25 mM Spm, 0.50 mM Spd, and 1 mM Put as seed priming or as a foliar spray at 20, 40, and 60 DAS |
| Taie et al. [12] |
| T. aestivum | 2 mM CdCl2 for 58 d | - | 0.25 mM Spm, 0.50 mM Spd, or 1.0 mM put as seed priming and later foliar spraying |
| Rady et al. [28] |
| T. aestivum | 1 mM CuCl2 for 5–15 d | - | 0.1 mM Put, Spd, or Spm pretreatment for 5 and 10 d |
| Benavides et al. [5] |
| T. aestivum | 1 mM Cd as CdCl2 | Increased PAs content | 2 mM Spd or 2 mM Spm as a seed treatment for 6 h |
| Rady and Hemida [11] |
| T. aestivum | 30 µM AlCl3 | Increased Spd | Put, 2 mM |
| Yu et al. [29] |
| T. aestivum | 2 mM Cd | Increased Spm and Spd content | - |
| Howladar et al. [30] |
| T. aestivum | 2.0 mM Pb2+ | - | 0.25 mM Spm, 0.50 mM Spd or 1.0 mM Put as seed treatment |
| Rady et al. [31] |
| Helianthus annuus L. | 1 mM CdCl2 for 5–15 d | Increased Put, Spd, and Spm content | 0.1 mM Put, Spd, or Spm pretreatment for 5 and 10 d |
| Benavides et al. [5] |
| H. annuus | 1 mM CuCl2 for 5–15 d | Increased Put, Spd, and Spm content | 0.1 mM Put, Spd, or Spm pretreatment for 5 and 10 d |
| Benavides et al. [5] |
| Vigna radiata L. | 1.5 mM CdCl2 | Increased Spd and Spm content with decrease the Put/PAs ratio | Put, 0.2 mM |
| Nahar et al. [1] |
| V. radiata | Zn, 200 mg kg−1 soil as ZnSO4·7H2O | - | 1.0 mM Spd, foliar application |
| Mir et al. [32] |
| Chlorella vulgaris Beijerinck | 100 µM of Cu as Cu(NO3)2·3H2O | - | 100 μM Spd |
| Piotrowska-Niczyporuk et al. [33] |
| C. vulgaris | 100 µM Cd, as Cd(NO3)2·4H2O | - | 100 μM Spd |
| Piotrowska-Niczyporuk et al. [33] |
| C. vulgaris | 100 µM of Pb Pb(NO3)2 | - | 100 μM Spd |
| Piotrowska-Niczyporuk et al. [33] |
| Raphanus sativus L. | 1.2 mM Cr as (K2CrO4) | Increased Put and Spd content | 1 mM Spd as cotreatment |
| Choudhary et al. [34] |
| Poncirus trifoliata L. | 0, 0.25, 0.50, 0.75, 1.0, 1.25 mM Cr as K2Cr2O7 | Increased PAs content | - |
| Shahid et al. [27] |
| Citrus reshni L. | 0, 0.25, 0.50, 0.75, 1.0, 1.25 mM Cr as K2Cr2O7 | Increased PAs content | - |
| Shahid et al. [27] |
| C. limonia Osbeck (CL) | 0, 0.25, 0.50, 0.75, 1.0, 1.25 mM Cr as K2Cr2O7 | Increased PAs content | - |
| Shahid et al. [27] |
| Salix matsudana Koidz. | 0.05 and 0.10 mM Cd | Increased endogenous Spd and Put contents | 0.25 mM Spd as cotreatment for 3 d |
| Tang et al. [35] |
| Potamogeton crispus L. | 30, 50, 70 µM Cd as CdCl2 | Increased Put, PS Put, and PIS Put content | - |
| Yang et al. [36] |
| Crop Species | Metal Exposure | Changes in Endogenous PAs Level | Exogenous PAs Applications | Antioxidant Defense System | References |
|---|---|---|---|---|---|
| Triticum aestivum L. | 2 mM Cd as CdCl2 and 2 mM as PbCl2 for 45 d | - | 0.25 mM Spm, 0.50 mM Spd, and 1 mM Put as seed priming or as a foliar spray at 20, 40, and 60 DAS |
| Taie et al. [12] |
| T. aestivum | 2 mM as PbCl2 for 45 d | - | 0.25 mM Spm, 0.50 mM Spd, and 1 mM Put as seed priming or as a foliar spray at 20, 40 and 60 DAS |
| Taie et al. [12] |
| T. aestivum | 2 mM CdCl2 for 58 d | - | 0.25 mM Spm, 0.50 mM Spd, 1.0 mM Put as seed priming and later foliar spraying |
| Rady et al. [28] |
| T. aestivum | 30 µM AlCl3 | Increased Spd | Put, 2 mM as cotreatment |
| Yu et al. [29] |
| T. aestivum | 2 mM Cd | Increased endogenous Spm and Spd upon Cd exposure | - |
| Howladar et al. [30] |
| T. aestivum | 1 mM Cd as CdCl2 | Increased PAs content by pretreated seedlings under stress affection | 2 mM Spd or 2 mM Spm as a seed treatment for 6 h |
| Rady and Hemida [11] |
| T. aestivum | 0.5 mM Cd as CdCl2 | Increased PAs content such as Put, Spd, and Spm | 0.5 and 1.0 mM Spm |
| Groppa et al. [37] |
| T. aestivum | 0.5 mM Cu as CuCl2 | - | 0.5 and 1.0 mM Spm |
| Groppa et al. [37] |
| Oryza sativa L. | 5 mM CdCl2 | - | 5 mM Put, 5 mM Spd, and 5 mM Spm |
| Hsu and Kao [38] |
| Vigna radiata | 1.5 mMCdCl2 | Increased Spd and Spd content with decrease the Put/PAs ratio | Put, 0.2 mM, as pretreatment for 24 h |
| Nahar et al. [1] |
| V. radiata | 200 mg kg−1 soil as ZnSO4·7H2O | - | 1.0 mM Spd, foliar application |
| Mir et al. [32] |
| Chlorella vulgaris Beijerinck | 100 µM of Cd as Cd(NO3)2·4H2O | - | 100 μM Spd |
| Piotrowska-Niczyporuk et al. [33] |
| C. vulgaris | 100 µM Pb as, Pb(NO3)2 | - | 100 μM Spd |
| Piotrowska-Niczyporuk et al. [33] |
| C. vulgaris | 100 µM Cu as Cu(NO3)2·3H2O | - | 100 μM Spd |
| Piotrowska-Niczyporuk et al. [33] |
| Raphanus sativus | 1.2 mM Cr as (K2CrO4) | Increased Put and Spd content | 1 mM Spd as cotreatment |
| Choudhary et al. [34] |
| Salix matsudana Koidz. | 0.05 and 0.10 mM Cd | Increased endogenous Spd and Put contents | 0.25 mM Spd as cotreatment for 3 d |
| Tang et al. [35] |
| Helianthus annuus | 0.5 mM Cd as CdCl2 | Increased endogenous Put and Spd levels | 1.0 mM Spd and Spm |
| Groppa et al. [39] |
| H. annuus | 0.5 mM Cu as CuCl2 | Increased endogenous Put and Spd levels | 1.0 mM Spd and Spm |
| Groppa et al. [39] |
| PA gene | Host plant/organism | Transgenic plant | Targeted metals (tolerance) | Reference |
|---|---|---|---|---|
| SPDS 1 | Apple | European Pear | Cd, Zn, and Pb | Wen et al. [56] |
| SPDS 1 | Apple | European Pear | Cu stress | Wen et al. [104] |
| SPDS 1 | Apple | European Pear | Cd stress | Wen et al. [105] |
| ADC gene | Agrobacterium | Eggplant | Cd stress | Prabhavathi and Rajam [106] |
| SPDS 1 | Apple | European Pear | Al stress | Wen et al. [107] |
| ALD | Arabidopsis | Cd and Cu stress | Sunkar et al. [108] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasanuzzaman, M.; Alhaithloul, H.A.S.; Parvin, K.; Bhuyan, M.H.M.B.; Tanveer, M.; Mohsin, S.M.; Nahar, K.; Soliman, M.H.; Mahmud, J.A.; Fujita, M. Polyamine Action under Metal/Metalloid Stress: Regulation of Biosynthesis, Metabolism, and Molecular Interactions. Int. J. Mol. Sci. 2019, 20, 3215. https://doi.org/10.3390/ijms20133215
Hasanuzzaman M, Alhaithloul HAS, Parvin K, Bhuyan MHMB, Tanveer M, Mohsin SM, Nahar K, Soliman MH, Mahmud JA, Fujita M. Polyamine Action under Metal/Metalloid Stress: Regulation of Biosynthesis, Metabolism, and Molecular Interactions. International Journal of Molecular Sciences. 2019; 20(13):3215. https://doi.org/10.3390/ijms20133215
Chicago/Turabian StyleHasanuzzaman, Mirza, Haifa Abdulaziz S. Alhaithloul, Khursheda Parvin, M.H.M. Borhannuddin Bhuyan, Mohsin Tanveer, Sayed Mohammad Mohsin, Kamrun Nahar, Mona H. Soliman, Jubayer Al Mahmud, and Masayuki Fujita. 2019. "Polyamine Action under Metal/Metalloid Stress: Regulation of Biosynthesis, Metabolism, and Molecular Interactions" International Journal of Molecular Sciences 20, no. 13: 3215. https://doi.org/10.3390/ijms20133215