
Rooting in the Desert: A Developmental Overview on Desert Plants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
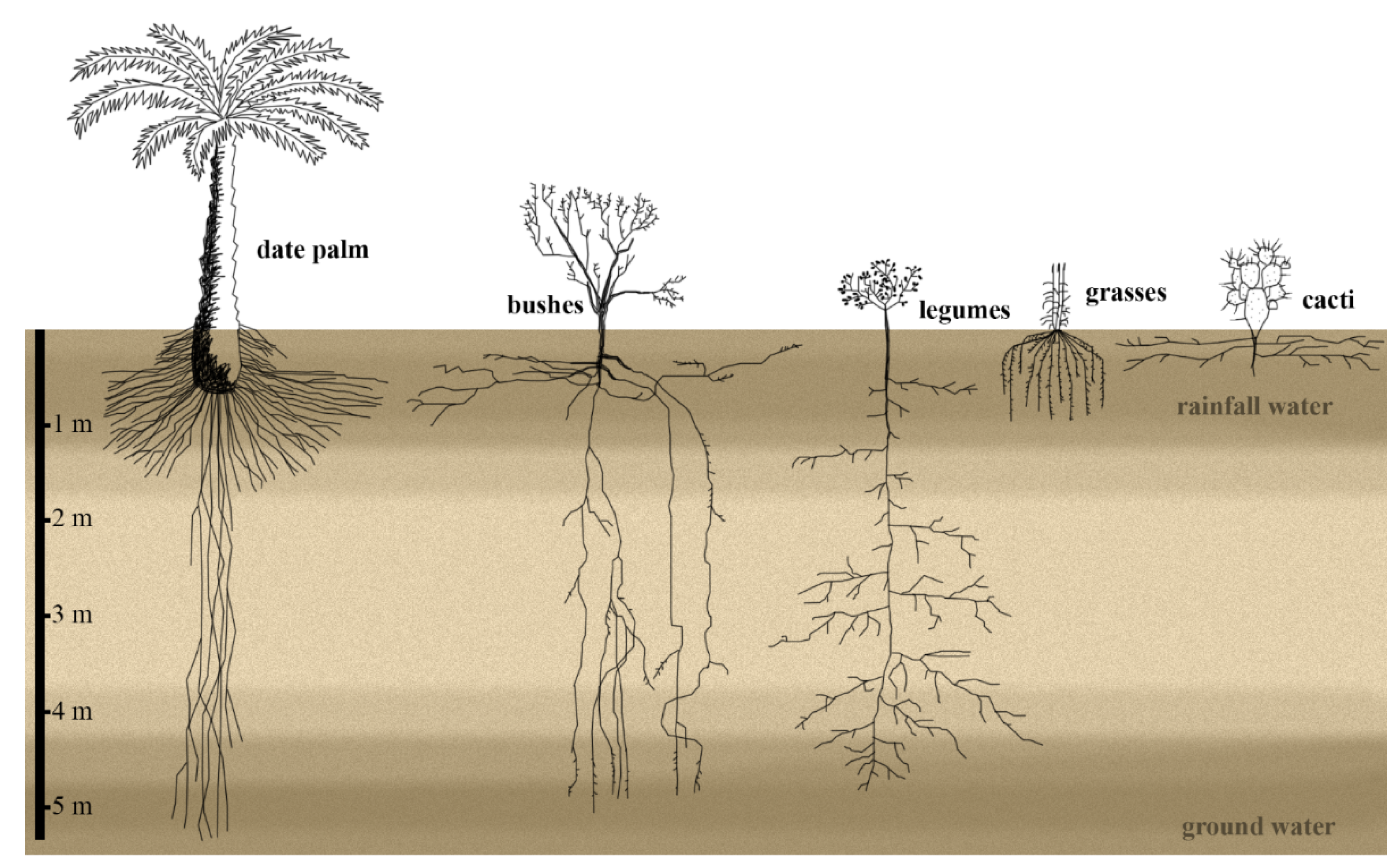
2. Below-Ground Organs: Rooting in the Desert
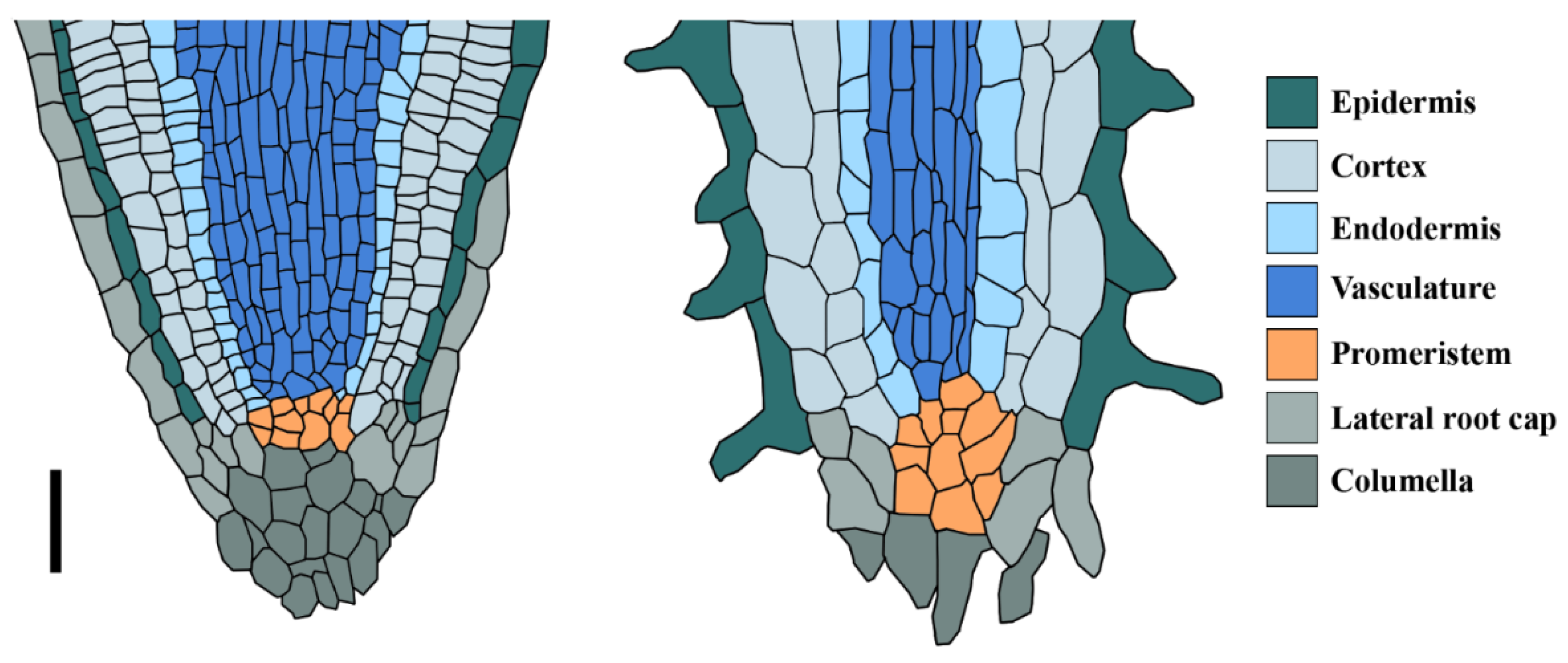
3. The Cactus Root System Is Optimized to Exploit Top-Soil Water
4. Hydraulic Conductivity in Cactus Root Tissue Is Adapted to Drought Conditions
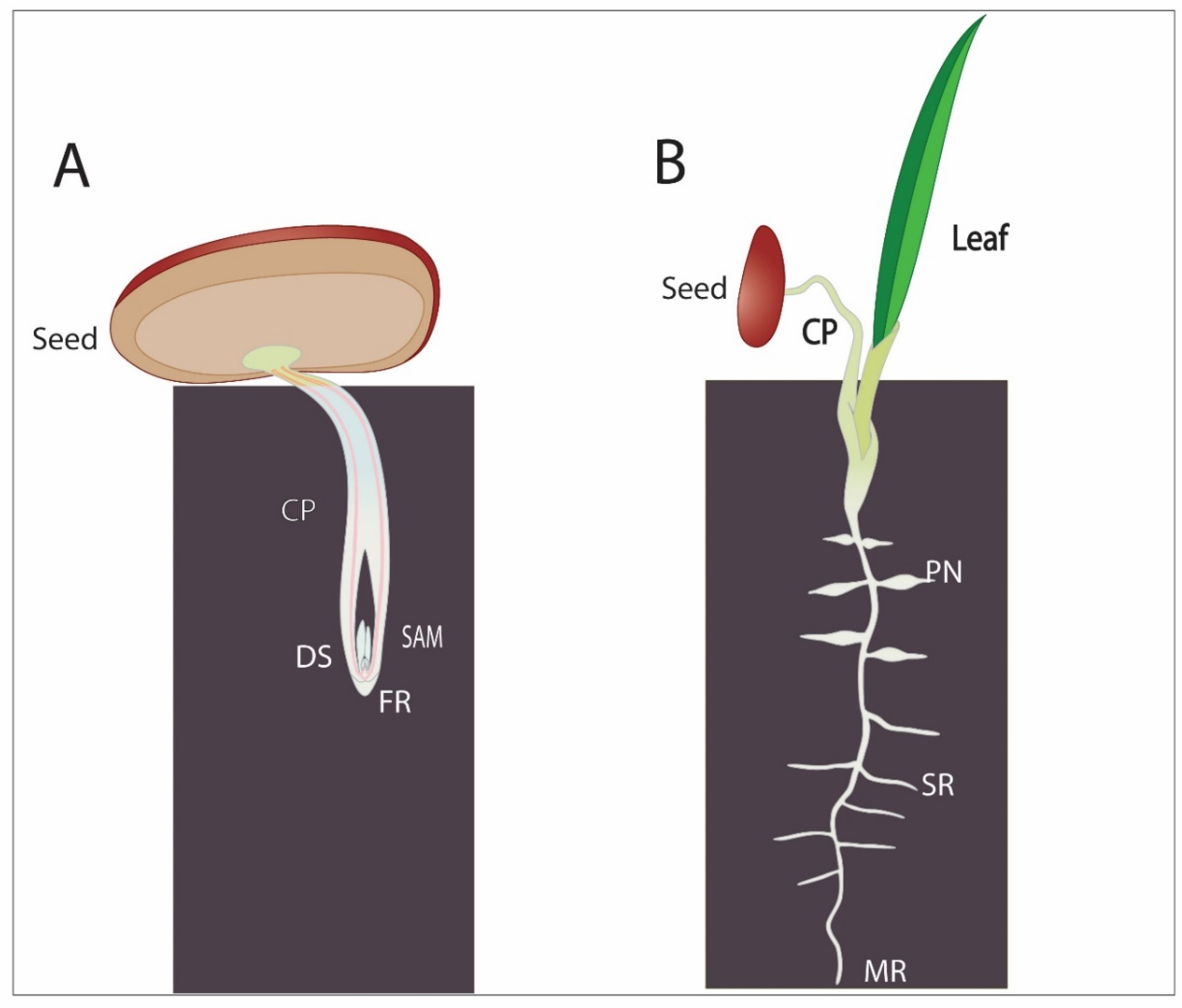
5. Date Palms Adaptation Strategies to Desert Soil
6. Desert Microbiome: The Invisible Root Ally
7. Conclusions: Lessons from Desert Plants
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nicholson, S.E. Desert hydrology. In Encyclopedia of Hydrology and Lakes; Springer: Dordrecht, The Netherlands, 1998; pp. 176–183. ISBN 978-1-4020-4497-7. [Google Scholar]
- Ehleringer, J.R.; Phillips, S.L.; Schuster, W.S.F.; Sandquist, D.R. Differential utilization of summer rains by desert plants. Oecologia 1991, 88, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.C. Photosynthetic organs of desert plants: Structural designs of nonsucculent desert plants cast doubt on the popular view that saving water is the key strategy. Bioscience 1998, 48, 911–920. [Google Scholar] [CrossRef]
- Gibbens, R.P.; Lenz, J.M. Root systems of some Chihuahuan Desert plants. J. Arid Environ. 2001, 49, 221–263. [Google Scholar] [CrossRef]
- Müller, H.M.; Schäfer, N.; Bauer, H.; Geiger, D.; Lautner, S.; Fromm, J.; Riederer, M.; Bueno, A.; Nussbaumer, T.; Mayer, K.; et al. The desert plant Phoenix dactylifera closes stomata via nitrate-regulated SLAC1 anion channel. New Phytol. 2017, 150–162. [Google Scholar] [CrossRef]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.T.; Krueger, R.R. The date palm (Phoenix dactylifera L.): Overview of biology, uses, and cultivation. HortScience 2007, 42, 1077–1082. [Google Scholar] [CrossRef]
- Dubrovsky, J.G.; North, G.B. Root structure and function. In CactiBiology and Uses; University of California Press: Oakland, CA, USA, 2002; pp. 41–56. [Google Scholar]
- Xiao, T.T.; Raygoza, A.A.; Pérez, J.C.; Kirschner, G.; Deng, Y.; Atkinson, B.; Sturrock, C.; Lube, V.; Wang, J.Y.; Lubineau, G.; et al. Emergent protective organogenesis in date palms: A morpho-devo-dynamic adaptive strategy during early development. Plant Cell 2019, 31, 1751–1766. [Google Scholar] [CrossRef]
- Al-Mssallem, I.S.; Hu, S.; Zhang, X.; Lin, Q.; Liu, W.; Tan, J.; Yu, X.; Liu, J.; Pan, L.; Zhang, T.; et al. Genome sequence of the date palm Phoenix dactylifera L. Nat. Commun. 2013, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Silos-Espino, H.; Valdez-Ortiz, A.; Rascón-Cruz, Q.; Rodríguez-Salazar, E.; Paredes-López, O. Genetic transformation of prickly-pear cactus (Opuntia ficus-indica) by Agrobacterium tumefaciens. Plant Cell Tissue Organ Cult. 2006, 86, 397–403. [Google Scholar] [CrossRef]
- Hochholdinger, F.; Park, W.J.; Sauer, M.; Woll, K. From weeds to crops: Genetic analysis of root development in cereals. Trends Plant Sci. 2004, 9, 42–48. [Google Scholar] [CrossRef]
- Orman-Ligeza, B.; Parizot, B.; Gantet, P.P.; Beeckman, T.; Bennett, M.J.; Draye, X. Post-embryonic root organogenesis in cereals: Branching out from model plants. Trends Plant Sci. 2013, 18, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.F.; Kemp, P.R.; Tenhunen, J.D. Effects of long-term rainfall variability on evapotranspiration and soil water distribution in the Chihuahuan Desert: A modeling analysis. Plant Ecol. 2000, 150, 145–159. [Google Scholar] [CrossRef]
- Xu, S.; Ji, X.; Jin, B.; Zhang, J. Root distribution of three dominant desert shrubs and their water uptake dynamics. J. Plant Ecol. 2016, 10, rtw079. [Google Scholar] [CrossRef]
- Guevara, A.; Giordano, C.V. Hydrotropism in lateral but not in pivotal roots of desert plant species under simulated natural conditions. Plant Soil 2015, 389, 257–272. [Google Scholar] [CrossRef]
- Liu, B.; Zeng, F.J.; Arndt, S.K.; He, J.X.; Luo, W.C.; Song, C. Patterns of root architecture adaptation of a phreatophytic perennial desert plant in a hyperarid desert. S. Afr. J. Bot. 2013, 86, 56–62. [Google Scholar] [CrossRef]
- Snyman, H.A. Root distribution with changes in distance and depth of two-year-old cactus pears Opuntia ficus-indica and O. robusta plants. S. Afr. J. Bot. 2006, 72, 434–441. [Google Scholar] [CrossRef]
- Saïd Ahmed, H.; Coquet, Y. Water uptake by date palm on Haplic Luvisols in the Djibouti coastal plain. Geoderma Reg. 2018, 15, e00189. [Google Scholar] [CrossRef]
- Snyman, H.A. A greenhouse study on root dynamics of cactus pears, Opuntia ficus-indica and Orobusta. J. Arid Environ. 2006, 65, 529–542. [Google Scholar] [CrossRef]
- Dubrovsky, J.G. Determinate primary-root growth in seedlings of Sonoran Desert Cactaceae; its organization, cellular basis, and ecological significance. Planta 1997, 203, 85–92. [Google Scholar] [CrossRef]
- Shishkova, S.; Las Peñas, M.L.; Napsucialy-Mendivil, S.; Matvienko, M.; Kozik, A.; Montiel, J.; Patiño, A.; Dubrovsky, J.G. Determinate primary root growth as an adaptation to aridity in Cactaceae: Towards an understanding of the evolution and genetic control of the trait. Ann. Bot. 2013, 112, 239–252. [Google Scholar] [CrossRef]
- Stahl, Y.; Simon, R. Plant primary meristems: Shared functions and regulatory mechanisms. Curr. Opin. Plant Biol. 2010, 13, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Heidstra, R.; Sabatini, S. Plant and animal stem cells: Similar yet different. Nat. Rev. Mol. Cell Biol. 2014, 15, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.; Scheres, B. Root development-two meristems for the price of one? Curr. Top. Dev. Biol. 2010, 91, 67–102. [Google Scholar] [CrossRef] [PubMed]
- Dello Ioio, R.; Linhares, F.S.; Scacchi, E.; Casamitjana-Martinez, E.; Heidstra, R.; Costantino, P.; Sabatini, S. Cytokinins determine Arabidopsis root-meristem size by controlling cell differentiation. Curr. Biol. 2007, 17, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, G.K.; Stahl, Y.; Von Korff, M.; Simon, R. Unique and conserved features of the barley root meristem. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Gundu, S.; Tabassum, N.; Blilou, I. Moving with purpose and direction: Transcription factor movement and cell fate determination revisited. Curr. Opin. Plant Biol. 2020, 57, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Clowes, F.A.L. Duration of the mitotic cycle in a meristem. J. Exp. Bot. 1961, 12, 283–293. [Google Scholar] [CrossRef]
- Feldman, L.J.; Torrey, J.G. The isolation and culture in vitro of the quiescent center of Zea mays. Am. J. Bot. 1976, 63, 345. [Google Scholar] [CrossRef]
- van den Berg, C.; Willemsen, V.; Hendriks, G.; Weisbeek, P.; Scheres, B. Short-range control of cell differentiation in the Arabidopsis root meristem. Nature 1997, 390, 287–289. [Google Scholar] [CrossRef]
- Heyman, J.; Kumpf, R.P.; De Veylder, L. A quiescent path to plant longevity. Trends Cell Biol. 2014, 24, 443–448. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, J.F.; Shishkova, S.; Napsucialy-Mendivil, S.; Dubrovsky, J.G. Apical meristem organization and lack of establishment of the quiescent center in Cactaceae roots with determinate growth. Planta 2003, 217, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Dubrovsky, J.G.; Gómez-Lomelí, L.F. Water deficit accelerates determinate developmental program of the primary root and does not affect lateral root initiation in a Sonoran Desert cactus (Pachycereus pringlei, Cactaceae). Am. J. Bot. 2003, 90, 823–831. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rodriguez-Alonso, G.; Matvienko, M.; López-Valle, M.L.; Lázaro-Mixteco, P.E.; Napsucialy-Mendivil, S.; Dubrovsky, J.G.; Shishkova, S. Transcriptomics insights into the genetic regulation of root apical meristem exhaustion and determinate primary root growth in Pachycereus pringlei (Cactaceae). Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Welch, D.; Hassan, H.; Blilou, I.; Immink, R.; Heidstra, R.; Scheres, B. Arabidopsis JACKDAW and MAGPIE zinc finger proteins delimit asymmetric cell division and stabilize tissue boundaries by restricting SHORT-ROOT action. Genes Dev. 2007, 21, 2196–2204. [Google Scholar] [CrossRef]
- Maud, A.W.H.; Rost, T.L. The control of lateral root development in cultured pea seedlings. I. The role of seedling organs and plant growth regulators. Bot. Gaz. 1986, 147, 137–147. [Google Scholar] [CrossRef]
- Wightman, F.; Thimann, K.V. Hormonal factors controlling the initiation and development of lateral roots: II. Effects of exogenous growth factors on lateral root formation in pea roots. Physiol. Plant. 1980, 49, 304–314. [Google Scholar] [CrossRef]
- Kirschner, G.K.; Stahl, Y.; Imani, J.; von Korff, M.; Simon, R. Fluorescent reporter lines for auxin and cytokinin signalling in barley (Hordeum vulgare). PLoS ONE 2018, 13, e0196086. [Google Scholar] [CrossRef] [PubMed]
- Di Mambro, R.; Svolacchia, N.; Dello Ioio, R.; Pierdonati, E.; Salvi, E.; Pedrazzini, E.; Vitale, A.; Perilli, S.; Sozzani, R.; Benfey, P.N.; et al. The lateral root cap acts as an auxin sink that controls meristem size. Curr. Biol. 2019, 29, 1199–1205. [Google Scholar] [CrossRef]
- Di Mambro, R.; De Ruvo, M.; Pacifici, E.; Salvi, E.; Sozzani, R.; Benfey, P.N.; Busch, W.; Novak, O.; Ljung, K.; Di Paola, L.; et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2017, 114, E7641–E7649. [Google Scholar] [CrossRef]
- Shishkova, S.; Rost, T.L.; Dubrovsky, J.G. Determinate root growth and meristem maintenance in angiosperms. Ann. Bot. 2008, 101, 319–340. [Google Scholar] [CrossRef]
- Zhou, W.; Lozano-Torres, J.L.; Blilou, I.; Zhang, X.; Zhai, Q.; Smant, G.; Li, C.; Scheres, B. A Jasmonate signaling network activates root stem cells and promotes regeneration. Cell 2019, 177, 942–956.e14. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, N.; Marcotte, L.; Giraudat, J. Drought rhizogenesis in Arabidopsis thaliana. Differential responses of hormonal mutants. Plant Physiol. 1994, 104, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Nawrath, C.; Schreiber, L.; Franke, R.B.; Geldner, N.; Reina-Pinto, J.J.; Kunst, L. Apoplastic diffusion barriers in Arabidopsis. Arab. Book Am. Soc. Plant Biol. 2013, e0167. [Google Scholar] [CrossRef] [PubMed]
- Heimsch, C.; Seago, J.L. Organization of the root apical meristem in angiosperms. Am. J. Bot. 2008, 95, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hunt, E.R.; Nobel, P.S. Allometric root/shoot relationships and predicted water uptake for desert succulents. Ann. Bot. 1987, 59, 571–577. [Google Scholar] [CrossRef]
- North, G.B.; Huang, B.; Nobel, P.S. Changes in structure and hydraulic conductivity for root junctions of desert succulents as soil water status varies. Bot. Acta 1993, 106, 126–135. [Google Scholar] [CrossRef]
- North, G.B.; Nobel, P.S. Changes in hydraulic conductivity and anatomy caused by drying and rewetting roots of agave deserti (Agavaceae). Am. J. Bot. 1991, 78, 906. [Google Scholar] [CrossRef]
- North, G.B.; Nobel, P.S. Drought-induced changes in hydraulic conductivity and structure in roots of Ferocactus acanthodes and Opuntia ficus-indica. New Phytol. 1992, 120, 9–19. [Google Scholar] [CrossRef]
- Dubrovsky, J.G.; North, G.B.; Nobel, P.S. Root growth, developmental changes in the apex, and hydraulic conductivity for Opuntia ficus-indica during drought. New Phytol. 1998, 138, 75–82. [Google Scholar] [CrossRef]
- Cuneo, I.F.; Knipfer, T.; Brodersen, C.R.; McElrone, A.J. Mechanical failure of fine root cortical cells initiates plant hydraulic decline during drought. Plant Physiol. 2016, 172, 1669–1678. [Google Scholar] [CrossRef]
- Nobel, P.S.; Cui, M. Shrinkage of attached roots of opuntia ficus-indica in response to lowered water potentials—Predicted consequences for water uptake or loss to soil. Ann. Bot. 1992, 70, 485–491. [Google Scholar] [CrossRef]
- Nobel, P.S. Recent ecophysiological findings for Opuntia ficus-indica. J. Prof. Assoc. Cactus Dev. 1997, 2, 89–96. [Google Scholar]
- Stevenson, J.F.; Mauseth, J.D. Effects of environment on vessel characters in cactus wood. Int. J. Plant Sci. 2004, 165, 347–357. [Google Scholar] [CrossRef][Green Version]
- Graça, J.; Santos, S. Suberin: A biopolyester of plants’ skin. Macromol. Biosci. 2007, 7, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Iossi, E.; Vittimoro, F.; Sader, R. Seed anatomy and germination of Phoenix roebelenii O’Brien (Arecaceae). Rev. Bras. Sementes 2006, 28, 121–128. [Google Scholar] [CrossRef]
- Seubert, E. Root anatomy of palms: I. Coryphoideae. Flora 1997, 192, 81–103. [Google Scholar] [CrossRef]
- Abd Elhalim, M.E.; Abo-Alatta, O.K.; Habib, S.A.; Abd Elbar, O.H. The anatomical features of the desert halophytes Zygophyllum album L.F. and Nitraria retusa (Forssk.) Asch. Ann. Agric. Sci. 2016, 61, 97–104. [Google Scholar] [CrossRef]
- Bokor, B.; Soukup, M.; Vaculík, M.; Vd’ačný, P.; Weidinger, M.; Lichtscheidl, I.; Vávrová, S.; Šoltys, K.; Sonah, H.; Deshmukh, R.; et al. Silicon uptake and localisation in date palm (Phoenix dactylifera)—A unique association with sclerenchyma. Front. Plant Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.L.; Ewers, F.W.; Pratt, R.B.; Paddock, W.A.; Davis, S.D. Do xylem fibers affect vessel cavitation resistance? Plant Physiol. 2005, 139, 546–556. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The soil-borne supremacy. Trends Plant Sci. 2016, 21, 171–173. [Google Scholar] [CrossRef]
- York, L.M.; Carminati, A.; Mooney, S.J.; Ritz, K.; Bennett, M.J. The holistic rhizosphere: Integrating zones, processes, and semantics in the soil influenced by roots. J. Exp. Bot. 2016, 67, 3629–3643. [Google Scholar] [CrossRef]
- Eida, A.A.; Ziegler, M.; Lafi, F.F.; Michell, C.T.; Voolstra, C.R.; Hirt, H.; Saad, M.M. Desert plant bacteria reveal host influence and beneficial plant growth properties. PLoS ONE 2018, 13, e0208223. [Google Scholar] [CrossRef] [PubMed]
- Eida, A.A.; Bougouffa, S.; L’Haridon, F.; Alam, I.; Weisskopf, L.; Bajic, V.B.; Saad, M.M.; Hirt, H. Genome insights of the plant-growth promoting bacterium Cronobacter muytjensii JZ38 with volatile-mediated antagonistic activity against phytophthora infestans. Front. Microbiol. 2020, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Cherif, H.; Marasco, R.; Rolli, E.; Ferjani, R.; Fusi, M.; Soussi, A.; Mapelli, F.; Blilou, I.; Borin, S.; Boudabous, A.; et al. Oasis desert farming selects environment-specific date palm root endophytic communities and cultivable bacteria that promote resistance to drought. Environ. Microbiol. Rep. 2015, 7, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, W.; Li, Q.; Cui, R.; Wang, Z.; Wang, Y.; Zhang, Y.-Z.; Ding, W.; Shen, X. Deciphering the root endosphere microbiome of the desert plant alhagi sparsifolia for drought resistance-promoting bacteria. Appl. Environ. Microbiol. 2020, 86, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zamioudis, C.; Mastranesti, P.; Dhonukshe, P.; Blilou, I.; Pieterse, C.M.J. Unraveling root developmental programs initiated by beneficial Pseudomonas spp. Bacteria. Plant Physiol. 2013, 162, 304–318. [Google Scholar] [CrossRef] [PubMed]
- de Zélicourt, A.; Synek, L.; Saad, M.M.; Alzubaidy, H.; Jalal, R.; Xie, Y.; Andrés-Barrao, C.; Rolli, E.; Guerard, F.; Mariappan, K.G.; et al. Ethylene induced plant stress tolerance by Enterobacter sp. SA187 is mediated by 2-keto-4-methylthiobutyric acid production. PLoS Genet. 2018, 14, 1–28. [Google Scholar] [CrossRef]
- Lynch, J.P.; Wojciechowski, T. Opportunities and challenges in the subsoil: Pathways to deeper rooted crops. J. Exp. Bot. 2015, 66, 2199–2210. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Partitioning of assimilates to deeper roots is associated with cooler canopies and increased yield under drought in wheat. Funct. Plant Biol. 2010, 37, 147. [Google Scholar] [CrossRef]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Liang, W.; Sturrock, C.J.; Pandey, B.K.; Giri, J.; Mairhofer, S.; Wang, D.; Muller, L.; Tan, H.; York, L.M.; et al. Rice actin binding protein RMD controls crown root angle in response to external phosphate. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirschner, G.K.; Xiao, T.T.; Blilou, I. Rooting in the Desert: A Developmental Overview on Desert Plants. Genes 2021, 12, 709. https://doi.org/10.3390/genes12050709
Kirschner GK, Xiao TT, Blilou I. Rooting in the Desert: A Developmental Overview on Desert Plants. Genes. 2021; 12(5):709. https://doi.org/10.3390/genes12050709
Chicago/Turabian StyleKirschner, Gwendolyn K., Ting Ting Xiao, and Ikram Blilou. 2021. "Rooting in the Desert: A Developmental Overview on Desert Plants" Genes 12, no. 5: 709. https://doi.org/10.3390/genes12050709
APA StyleKirschner, G. K., Xiao, T. T., & Blilou, I. (2021). Rooting in the Desert: A Developmental Overview on Desert Plants. Genes, 12(5), 709. https://doi.org/10.3390/genes12050709