
Endometriosis Is Associated with Functional Polymorphism in the Promoter of Heme Oxygenase 1 (HMOX1) Gene

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Control Subjects
2.2. Typing for −413A>T and (GT)n Polymorphisms
2.3. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, S.W.; Wang, Y. Sources of heterogeneities in estimating the prevalence of endometriosis in infertile and previously fertile women. Fertil. Steril. 2006, 86, 1584–1595. [Google Scholar] [CrossRef]
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. N. Engl. J. Med. 2020, 382, 1244–1256. [Google Scholar] [CrossRef]
- Zondervan, K.T.; Cardon, L.R.; Kennedy, S.H. The genetic basis of endometriosis. Curr. Opin. Obstet. Gynecol. 2001, 13, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Koninckx, P.R.; Ussia, A.; Adamyan, L.; Wattiez, A.; Gomel, V.; Martin, D.C. Pathogenesis of endometriosis: The genetic/epigenetic theory. Fertil. Steril. 2019, 111, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Sampson, J.A. Peritoneal endometriosis due to menstrual dissemination of endometrial tissue into the peritoneal cavity. Am. J. Obst. Gynecol. 1927, 14, 442–469. [Google Scholar] [CrossRef]
- Sciezynska, A.; Komorowski, M.; Soszynska, M.; Malejczyk, J. NK Cells as Potential Targets for Immunotherapy in Endometriosis. J. Clin. Med. 2019, 8, 1468. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Velasco, J.A.; Arici, A. Apoptosis and the pathogenesis of endometriosis. Semin. Reprod. Med. 2003, 21, 165–172. [Google Scholar] [CrossRef]
- Becker, C.M.; D’Amato, R.J. Angiogenesis and antiangiogenic therapy in endometriosis. Microvasc. Res. 2007, 74, 121–130. [Google Scholar] [CrossRef]
- Harada, T.; Kaponis, A.; Iwabe, T.; Taniguchi, F.; Makrydimas, G.; Sofikitis, N.; Paschopoulos, M.; Paraskevaidis, E.; Terakawa, N. Apoptosis in human endometrium and endometriosis. Hum. Reprod. Update 2004, 10, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazvani, R.; Templeton, A. Peritoneal environment, cytokines and angiogenesis in the pathophysiology of endometriosis. Reproduction 2002, 123, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Van Langendonckt, A.; Casanas-Roux, F.; Donnez, J. Oxidative stress and peritoneal endometriosis. Fertil. Steril. 2002, 77, 861–870. [Google Scholar] [CrossRef]
- Ngo, C.; Chereau, C.; Nicco, C.; Weill, B.; Chapron, C.; Batteux, F. Reactive oxygen species controls endometriosis progression. Am. J. Pathol. 2009, 175, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Kajihara, H.; Yamada, Y.; Kanayama, S.; Furukawa, N.; Noguchi, T.; Haruta, S.; Yoshida, S.; Sado, T.; Oi, H.; Kobayashi, H. New insights into the pathophysiology of endometriosis: From chronic inflammation to danger signal. Gynecol. Endocrinol. 2011, 27, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Van Langendonckt, A.; Casanas-Roux, F.; Donnez, J. Iron overload in the peritoneal cavity of women with pelvic endometriosis. Fertil. Steril. 2002, 78, 712–718. [Google Scholar] [CrossRef]
- Van Langendonckt, A.; Casanas-Roux, F.; Dolmans, M.M.; Donnez, J. Potential involvement of hemoglobin and heme in the pathogenesis of peritoneal endometriosis. Fertil. Steril. 2002, 77, 561–570. [Google Scholar] [CrossRef]
- Kobayashi, H.; Yamada, Y.; Kanayama, S.; Furukawa, N.; Noguchi, T.; Haruta, S.; Yoshida, S.; Sakata, M.; Sado, T.; Oi, H. The role of iron in the pathogenesis of endometriosis. Gynecol. Endocrinol. 2009, 25, 39–52. [Google Scholar] [CrossRef]
- Woo, J.H.; Choi, Y.S.; Choi, J.H. Iron-Storage Protein Ferritin Is Upregulated in Endometriosis and Iron Overload Contributes to a Migratory Phenotype. Biomedicines 2020, 8, 454. [Google Scholar] [CrossRef]
- Defrere, S.; Gonzalez-Ramos, R.; Lousse, J.C.; Colette, S.; Donnez, O.; Donnez, J.; Van Langendonckt, A. Insights into iron and nuclear factor-kappa B (NF-kappaB) involvement in chronic inflammatory processes in peritoneal endometriosis. Histol. Histopathol. 2011, 26, 1083–1092. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, X.; Lu, D.; Yin, M.; Shan, M.; Gao, Y. Erastin induces ferroptosis via ferroportin-mediated iron accumulation in endometriosis. Hum. Reprod. 2020. [Google Scholar] [CrossRef]
- Agarwal, A.; Nick, H.S. Renal response to tissue injury: Lessons from heme oxygenase-1 GeneAblation and expression. J. Am. Soc. Nephrol. 2000, 11, 965–973. [Google Scholar]
- Kumar, S.; Bandyopadhyay, U. Free heme toxicity and its detoxification systems in human. Toxicol. Lett. 2005, 157, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Dulak, J.; Deshane, J.; Jozkowicz, A.; Agarwal, A. Heme oxygenase-1 and carbon monoxide in vascular pathobiology: Focus on angiogenesis. Circulation 2008, 117, 231–241. [Google Scholar] [CrossRef]
- Keyse, S.M.; Tyrrell, R.M. Heme oxygenase is the major 32-kDa stress protein induced in human skin fibroblasts by UVA radiation, hydrogen peroxide, and sodium arsenite. Proc. Natl. Acad. Sci. USA 1989, 86, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, D.; Choi, A.M. Heme oxygenase-1: The “emerging molecule” has arrived. Am. J. Respir. Cell Mol. Biol. 2002, 27, 8–16. [Google Scholar] [CrossRef]
- Farombi, E.O.; Surh, Y.J. Heme oxygenase-1 as a potential therapeutic target for hepatoprotection. J. Biochem. Mol. Biol. 2006, 39, 479–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Goto, Y.; Takagi, S.; Baba, S.; Tago, N.; Nonogi, H.; Iwai, N. A promoter variant of the heme oxygenase-1 gene may reduce the incidence of ischemic heart disease in Japanese. Atherosclerosis 2004, 173, 315–319. [Google Scholar] [CrossRef]
- Ono, K.; Mannami, T.; Iwai, N. Association of a promoter variant of the haeme oxygenase-1 gene with hypertension in women. J. Hypertens. 2003, 21, 1497–1503. [Google Scholar] [CrossRef]
- Exner, M.; Minar, E.; Wagner, O.; Schillinger, M. The role of heme oxygenase-1 promoter polymorphisms in human disease. Free Radic. Biol. Med. 2004, 37, 1097–1104. [Google Scholar] [CrossRef]
- Andrews, W.C.; Buttram, V.C., Jr.; Weed, J.C.; Hammond, C.B.; Thomas, H.H. Revised American Fertility Society classification of endometriosis: 1985. Fertil. Steril. 1985, 43, 351–352. [Google Scholar] [CrossRef]
- Skorka, A.; Bednarczuk, T.; Bar-Andziak, E.; Nauman, J.; Ploski, R. Lymphoid tyrosine phosphatase (PTPN22/LYP) variant and Graves’ disease in a Polish population: Association and gene dose-dependent correlation with age of onset. Clin. Endocrinol. 2005, 62, 679–682. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Kimpara, T.; Takeda, A.; Watanabe, K.; Itoyama, Y.; Ikawa, S.; Watanabe, M.; Arai, H.; Sasaki, H.; Higuchi, S.; Okita, N.; et al. Microsatellite polymorphism in the human heme oxygenase-1 gene promoter and its application in association studies with Alzheimer and Parkinson disease. Hum. Genet. 1997, 100, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2007, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.M.; Ehm, M.G.; Weir, B.S. Detecting marker-disease association by testing for Hardy-Weinberg disequilibrium at a marker locus. Am. J. Hum. Genet. 1998, 63, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Balding, D.J. A tutorial on statistical methods for population association studies. Nat. Rev. Genet. 2006, 7, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Denschlag, D.; Marculescu, R.; Unfried, G.; Hefler, L.A.; Exner, M.; Hashemi, A.; Riener, E.K.; Keck, C.; Tempfer, C.B.; Wagner, O. The size of a microsatellite polymorphism of the haem oxygenase 1 gene is associated with idiopathic recurrent miscarriage. Mol. Hum. Reprod. 2004, 10, 211–214. [Google Scholar] [CrossRef]
- Hirai, H.; Kubo, H.; Yamaya, M.; Nakayama, K.; Numasaki, M.; Kobayashi, S.; Suzuki, S.; Shibahara, S.; Sasaki, H. Microsatellite polymorphism in heme oxygenase-1 gene promoter is associated with susceptibility to oxidant-induced apoptosis in lymphoblastoid cell lines. Blood 2003, 102, 1619–1621. [Google Scholar] [CrossRef] [PubMed]
- Taha, H.; Skrzypek, K.; Guevara, I.; Nigisch, A.; Mustafa, S.; Grochot-Przeczek, A.; Ferdek, P.; Was, H.; Kotlinowski, J.; Kozakowska, M.; et al. Role of heme oxygenase-1 in human endothelial cells: Lesson from the promoter allelic variants. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1634–1641. [Google Scholar] [CrossRef] [Green Version]
- Yamada, N.; Yamaya, M.; Okinaga, S.; Nakayama, K.; Sekizawa, K.; Shibahara, S.; Sasaki, H. Microsatellite polymorphism in the heme oxygenase-1 gene promoter is associated with susceptibility to emphysema. Am. J. Hum. Genet. 2000, 66, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Lin, S.J.; Lin, M.W.; Tsai, H.L.; Kuo, S.S.; Chen, J.W.; Charng, M.J.; Wu, T.C.; Chen, L.C.; Ding, Y.A.; et al. Microsatellite polymorphism in promoter of heme oxygenase-1 gene is associated with susceptibility to coronary artery disease in type 2 diabetic patients. Hum. Genet. 2002, 111, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Brouard, S.; Berberat, P.O.; Tobiasch, E.; Seldon, M.P.; Bach, F.H.; Soares, M.P. Heme oxygenase-1-derived carbon monoxide requires the activation of transcription factor NF-kappa B to protect endothelial cells from tumor necrosis factor-alpha-mediated apoptosis. J. Biol. Chem. 2002, 277, 17950–17961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, G.; Cunha, A.; Gregoire, I.P.; Seldon, M.P.; Soares, M.P. The antiapoptotic effect of heme oxygenase-1 in endothelial cells involves the degradation of p38 alpha MAPK isoform. J. Immunol. 2006, 177, 1894–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Was, H.; Cichon, T.; Smolarczyk, R.; Rudnicka, D.; Stopa, M.; Chevalier, C.; Leger, J.J.; Lackowska, B.; Grochot, A.; Bojkowska, K.; et al. Overexpression of heme oxygenase-1 in murine melanoma: Increased proliferation and viability of tumor cells, decreased survival of mice. Am. J. Pathol. 2006, 169, 2181–2198. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.S.; Chau, L.Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat. Med. 2002, 8, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.S.; Tsai, H.L.; Chau, L.Y. Induction of heme oxygenase-1 expression in murine macrophages is essential for the anti-inflammatory effect of low dose 15-deoxy-Delta 12,14-prostaglandin J2. J. Biol. Chem. 2003, 278, 19325–19330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, A.; Alves-Filho, J.C.; Secco, D.D.; Neto, A.F.; Ferreira, S.H.; Barja-Fidalgo, C.; Cunha, F.Q. Heme oxygenase/carbon monoxide-biliverdin pathway down regulates neutrophil rolling, adhesion and migration in acute inflammation. Br. J. Pharmacol. 2006, 149, 345–354. [Google Scholar] [CrossRef]
- Piantadosi, C.A.; Withers, C.M.; Bartz, R.R.; MacGarvey, N.C.; Fu, P.; Sweeney, T.E.; Welty-Wolf, K.E.; Suliman, H.B. Heme oxygenase-1 couples activation of mitochondrial biogenesis to anti-inflammatory cytokine expression. J. Biol. Chem. 2011, 286, 16374–16385. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, P.; Lu, J.; Xiong, W.; Oger, J.; Tetzlaff, W.; Cynader, M. Bilirubin possesses powerful immunomodulatory activity and suppresses experimental autoimmune encephalomyelitis. J. Immunol. 2008, 181, 1887–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegiel, B.; Baty, C.J.; Gallo, D.; Csizmadia, E.; Scott, J.R.; Akhavan, A.; Chin, B.Y.; Kaczmarek, E.; Alam, J.; Bach, F.H.; et al. Cell surface biliverdin reductase mediates biliverdin-induced anti-inflammatory effects via phosphatidylinositol 3-kinase and Akt. J. Biol. Chem. 2009, 284, 21369–21378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Wang, J.; Jiang, H.; Ma, Y.; Pan, S.; Reddy, S.; Sun, X. Bilirubin protects grafts against nonspecific inflammation-induced injury in syngeneic intraportal islet transplantation. Exp. Mol. Med. 2010, 42, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Grochot-Przeczek, A.; Dulak, J.; Jozkowicz, A. Heme oxygenase-1 in neovascularisation: A diabetic perspective. Thromb. Haemost. 2010, 104, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.P.; Seldon, M.P.; Gregoire, I.P.; Vassilevskaia, T.; Berberat, P.O.; Yu, J.; Tsui, T.Y.; Bach, F.H. Heme oxygenase-1 modulates the expression of adhesion molecules associated with endothelial cell activation. J. Immunol. 2004, 172, 3553–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Allele/Genotype | Endometriosis (n = 228) | Control (n = 415) | p-Value * | OR (95% CI) | |
|---|---|---|---|---|---|
| -413A>T | A T | 269 (59.0%) 187 (41.0%) | 459 (55.3%) 371 (44.7%) | 0.2017 | 1.16 (0.92–1.47) |
| AA AT TT | 90 (39.5%) 89 (39.0%) 49 (21.5%) | 127 (30.6%) 205 (49.4%) 83 (20.0%) | 0.0228 0.0132 0.6547 | 1.48 (1.06–2.07) 0.66 (0.48–0.92) 1.09 (0.74–1.63) | |
| HWE P | 0.004 | 0.987 | |||
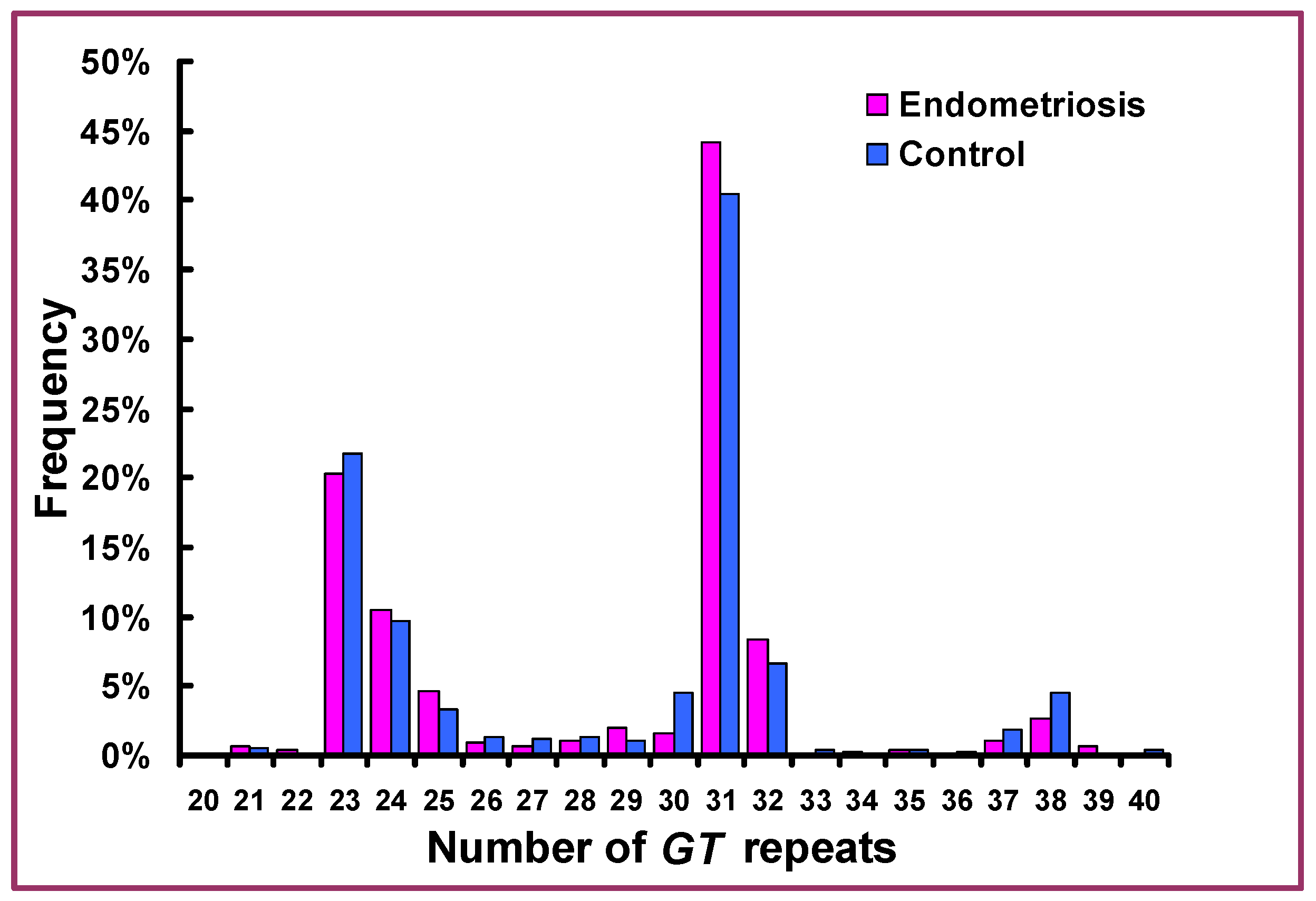
| (GT)n | (GT)23,24 (GT)31,32 | 141 (30.8%) 240 (52.4%) | 262 (31.6%) 392 (47.2%) | 0.777 0.0754 | 0.96 (0.75–1.23) 1.23 (0.98–1.55) |
| (GT)23,24(GT)23,24 (GT)23,24(GT)31,32 (GT)31,32(GT)31,32 | 23 (10.1%) 62 (27.2%) 72 (31.6%) | 41 (9.1%) 119 (28.7%) 101 (24.3%) | 0.9203 0.6891 0.0480 | 1.02 (0.60–1.75) 0.93 (0.65–1.33) 1.43 (1.01–2.05) | |
| HWE P | 0.09 ** | 0.39 ** |
| (GT)n Genotype | 416A>T SNP Genotype | ||||||
|---|---|---|---|---|---|---|---|
| AA | AT | TT | |||||
| Endometriosis n = 90 | Control n = 127 | Endometriosis n = 89 | Control n = 205 | Endometriosis n = 49 | Control n = 83 | ||
| 21 | 23 | 0 | 0 | 1 (0.4%) | 0 | 0 | 1 (0.2%) |
| 21 | 24 | 0 | 0 | 0 | 1 (0.2%) | 1 (0.4%) | 0 |
| 21 | 26 | 0 | 0 | 0 | 0 | 1 (0.4%) | 0 |
| 21 | 31 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 21 | 32 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 22 | 27 | 0 | 0 | 0 | 0 | 1 (0.4%) | 0 |
| 22 | 31 | 0 | 0 | 1 (0.4%) | 1 (0.2%) | 0 | 0 |
| 23 | 23 | 0 | 0 | 0 | 1 (0.2%) | 11 (4.8%) | 16 (3.9%) |
| 23 | 24 | 0 | 0 | 1 (0.4%) | 0 | 6 (2.6%) | 21 (5.1%) |
| 23 | 25 | 0 | 0 | 3 (1.3%) | 2 (0.5%) | 2 (0.9%) | 6 (1.4%) |
| 23 | 26 | 0 | 0 | 0 | 1 (0.2%) | 1 (0.4%) | 2 (0.5%) |
| 23 | 27 | 0 | 0 | 0 | 1 (0.2%) | 0 | 1 (0.2%) |
| 23 | 28 | 0 | 0 | 0 | 2 (0.5%) | 0 | 1 (0.2%) |
| 23 | 29 | 0 | 0 | 0 | 0 | 2 (0 (1.9%) | 1 (0.2%) |
| 23 | 30 | 0 | 0 | 1 (0.4%) | 10 (2.4%) | 0 | 0 |
| 23 | 31 | 2 (0.9%) | 1 (0.2%) | 34 (14.9%) | 74 (17.8%) | 1 (0.4%) | 0 |
| 23 | 32 | 0 | 0 | 8 (3.5%) | 9 (2.1%) | 0 | 0 |
| 23 | 33 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 23 | 35 | 0 | 0 | 0 | 0 | 1 (0.4%) | 2 (0.5%) |
| 23 | 37 | 0 | 0 | 0 | 0 | 2 (0.9%) | 3 (0.7%) |
| 23 | 38 | 0 | 0 | 0 | 0 | 4 (1.7%) | 8 (1.9%) |
| 23 | 39 | 0 | 0 | 0 | 0 | 1 (0.4%) | 0 |
| 24 | 24 | 0 | 0 | 0 | 0 | 5 (2.2%) | 3 (0.7%) |
| 24 | 25 | 0 | 0 | 0 | 1 (0.2%) | 5 (2.2%) | 1 (0.2%) |
| 24 | 26 | 0 | 0 | 0 | 1 (0.2%) | 0 | 2 (0.5%) |
| 24 | 28 | 0 | 0 | 1 (0.4%) | 1 (0.2%) | 0 | 0 |
| 24 | 29 | 0 | 0 | 1 (0.4%) | 1 (0.2%) | 0 | 0 |
| 24 | 30 | 0 | 0 | 1 (0.4%) | 4 (1%) | 1 (0.4%) | 0 |
| 24 | 31 | 0 | 0 | 15 (6.6%) | 30 (7.2%) | 0 | 0 |
| 24 | 32 | 0 | 0 | 2 (0.9%) | 5 (1.2%) | 0 | 0 |
| 24 | 38 | 0 | 0 | 0 | 0 | 3 (1.3%) | 6 (1.4%) |
| 24 | 39 | 0 | 0 | 0 | 0 | 1 (0.4%) | 0 |
| 24 | 40 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| 25 | 27 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| 25 | 28 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| 25 | 30 | 0 | 0 | 1 (0.4%) | 2 (0.5%) | 0 | 0 |
| 25 | 31 | 3 (1.3%) | 2 (0.5%) | 3 (1.3%) | 5 (1.2%) | 0 | 0 |
| 25 | 32 | 2 (0.9%) | 0 | 1 (0.4%) | 1 (0.2%) | 0 | 0 |
| 25 | 34 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 25 | 37 | 0 | 0 | 0 | 1 (0.2%) | 0 | 2 (0.5%) |
| 25 | 39 | 0 | 0 | 1 (0.4%) | 0 | 0 | 1 (0.2%) |
| 26 | 29 | 1 (0.4%) | 0 | 0 | 0 | 0 | 0 |
| 26 | 31 | 1 (0.4%) | 1 (0.2%) | 0 | 3 (0.7%) | 0 | 0 |
| 26 | 32 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 27 | 31 | 1 (0.4%) | 2 (0.5%) | 1 (0.4%) | 4 (1%) | 0 | 0 |
| 27 | 32 | 0 | 1 (0.2%) | 0 | 0 | 0 | 0 |
| 28 | 31 | 2 (0.9%) | 4 (1%) | 2 | 1 (0.2%) | 0 | 0 |
| 28 | 32 | 0 | 1 (0.2%) | 0 | 0 | 0 | 0 |
| 29 | 29 | 1 (0.4%) | 0 | 0 | 0 | 0 | 0 |
| 29 | 30 | 0 | 1 (0.2%) | 0 | 0 | 0 | 0 |
| 29 | 31 | 2 (0.9%) | 5 (1.2%) | 1 (0.4%) | 0 | 0 | 0 |
| 29 | 38 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| 30 | 30 | 0 | 5 (1.2%) | 0 | 0 | 0 | 0 |
| 30 | 31 | 1 (0.4%) | 3 (0.7%) | 1 (0.4%) | 1 (0.2%) | 0 | 0 |
| 30 | 32 | 1 (0.4%) | 2 (0.5%) | 0 | 0 | 0 | 0 |
| 30 | 35 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 30 | 38 | 0 | 0 | 0 | 4 (1%) | 0 | 0 |
| 31 | 31 | 50 (21.9%) | 72 (17.3%) | 0 | 3 (0.7%) | 0 | 0 |
| 31 | 32 | 22 (9.6%) | 19 (4.6%) | 0 | 1 (0.2%) | 0 | 0 |
| 31 | 33 | 0 | 2 (0.5%) | 0 | 0 | 0 | 0 |
| 31 | 34 | 1 (0.4%) | 0 | 0 | 0 | 0 | 0 |
| 31 | 35 | 0 | 0 | 1 (0.4%) | 0 | 0 | 0 |
| 31 | 36 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 31 | 37 | 0 | 0 | 2 (0.9%) | 9 (2.2%) | 0 | 0 |
| 31 | 38 | 0 | 0 | 4 | 15 (3.6%) | 0 | 0 |
| 31 | 40 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 32 | 32 | 0 | 6 (1.4%) | 0 | 0 | 0 | 0 |
| 32 | 37 | 0 | 0 | 1 (0.4%) | 1 (0.2%) | 0 | 0 |
| 32 | 38 | 0 | 0 | 1 (0.4%) | 0 | 0 | 0 |
| 32 | 40 | 0 | 0 | 0 | 1 (0.2%) | 0 | 0 |
| 36 | 38 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| 38 | 38 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| Haplotype/Genotype | Endometriosis (n = 228) | Control (n = 415) | p-Value * | OR (95%CI) |
|---|---|---|---|---|
| −413A(GT)31,32 −413T(GT)23,24 | 238 (52.1%) 138 (30.2%) | 388 (46.8%) 260 (31.3%) | 0.0381 0.6954 | 1.27 (1.01–1.60) 0.95 (0.74–1.15) |
| −413A(GT)31,32/413A(GT)31,32 −413A(GT)other/−413A(GT)other | 72 (31.6%) 18 (7.9%) | 97 (23.4%) 30 (7.2%) | 0.0238 0.7641 | 1.51 (1.06–2.17) 1.10 (0.60–2.02) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milewski, Ł.; Ścieżyńska, A.; Ponińska, J.; Soszyńska, M.; Barcz, E.; Roszkowski, P.I.; Kamiński, P.; Włodarski, P.; Płoski, R.; Malejczyk, J. Endometriosis Is Associated with Functional Polymorphism in the Promoter of Heme Oxygenase 1 (HMOX1) Gene. Cells 2021, 10, 695. https://doi.org/10.3390/cells10030695
Milewski Ł, Ścieżyńska A, Ponińska J, Soszyńska M, Barcz E, Roszkowski PI, Kamiński P, Włodarski P, Płoski R, Malejczyk J. Endometriosis Is Associated with Functional Polymorphism in the Promoter of Heme Oxygenase 1 (HMOX1) Gene. Cells. 2021; 10(3):695. https://doi.org/10.3390/cells10030695
Chicago/Turabian StyleMilewski, Łukasz, Aneta Ścieżyńska, Joanna Ponińska, Marta Soszyńska, Ewa Barcz, Piotr I. Roszkowski, Paweł Kamiński, Paweł Włodarski, Rafał Płoski, and Jacek Malejczyk. 2021. "Endometriosis Is Associated with Functional Polymorphism in the Promoter of Heme Oxygenase 1 (HMOX1) Gene" Cells 10, no. 3: 695. https://doi.org/10.3390/cells10030695