
Blood Eosinophils Are Associated with Efficacy of Targeted Therapy in Patients with Advanced Melanoma

 , , , , and
, , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Cohort and Healthy Donors
2.2. Isolation and Purification of Eosinophils
2.3. Purity and Phenotyping of Eosinophils
2.4. Cell Lines
2.5. Cytotoxicity Assays
2.6. Lysis of Eosinophils and Inactivation of Contents
2.7. Cytospins and HE staining
2.8. Proliferation and Cell Cycle Assays
2.9. Colony Formation Assay
2.10. ELISA
2.11. Multiplex-Analysis
2.12. Statistical Analysis
3. Results
3.1. Patients
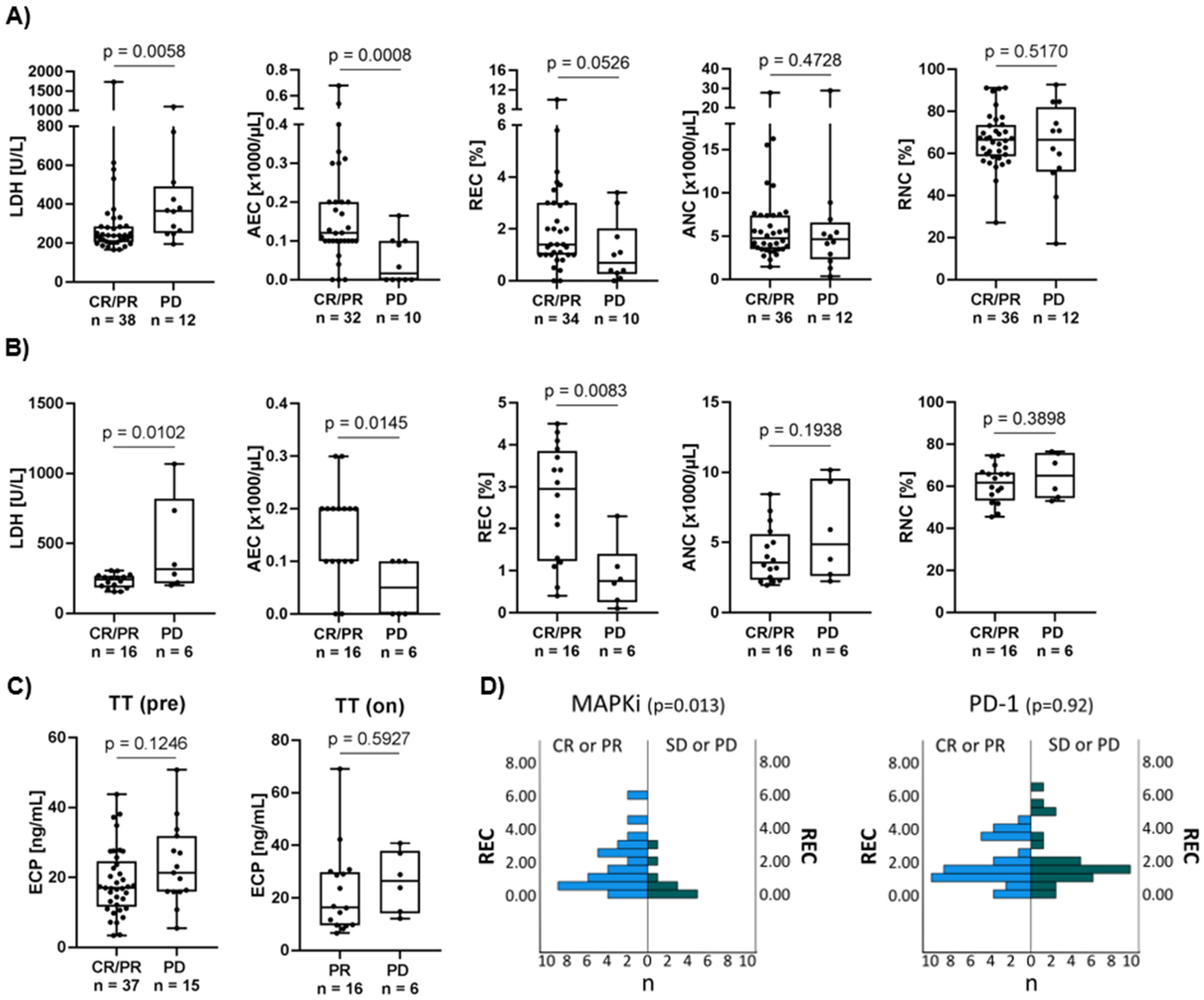
3.2. High Eosinophil Count Is Associated with Better Response to Targeted Therapy
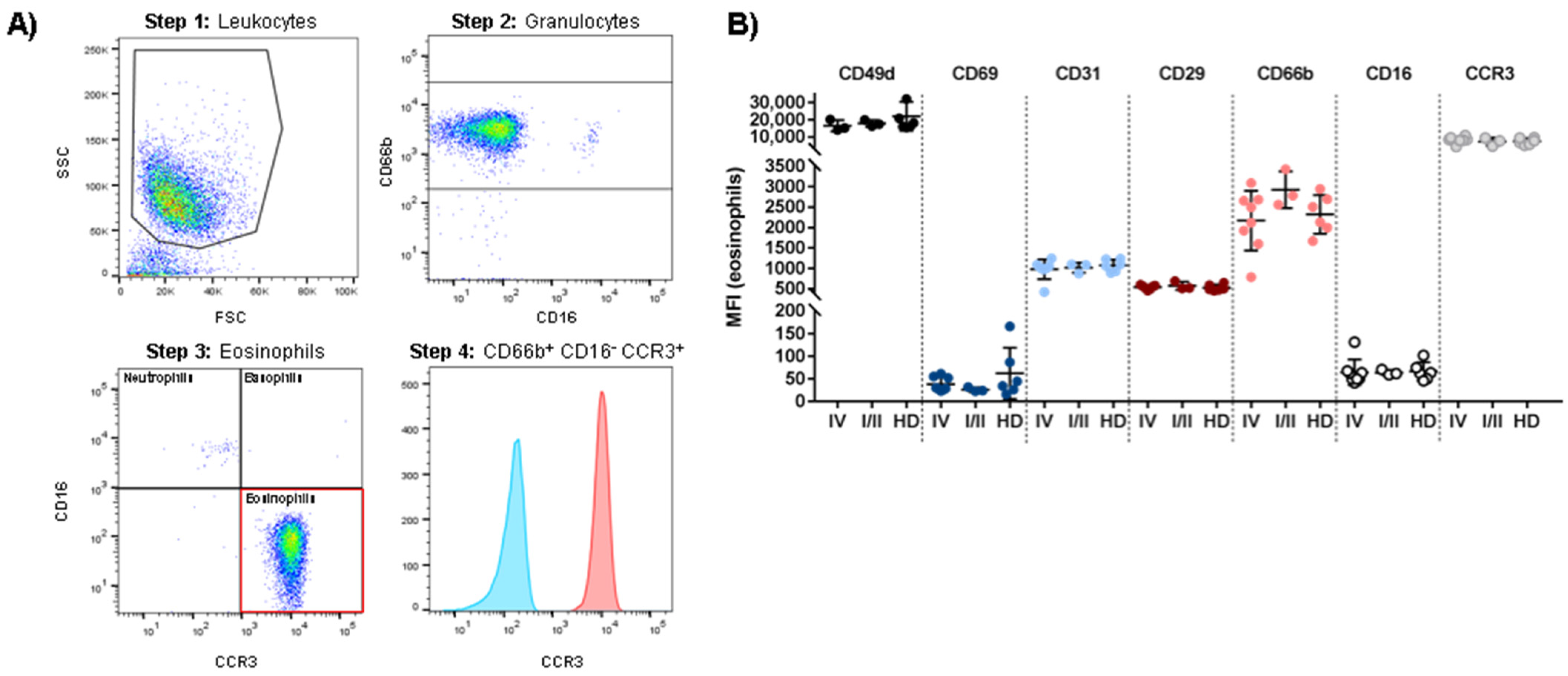
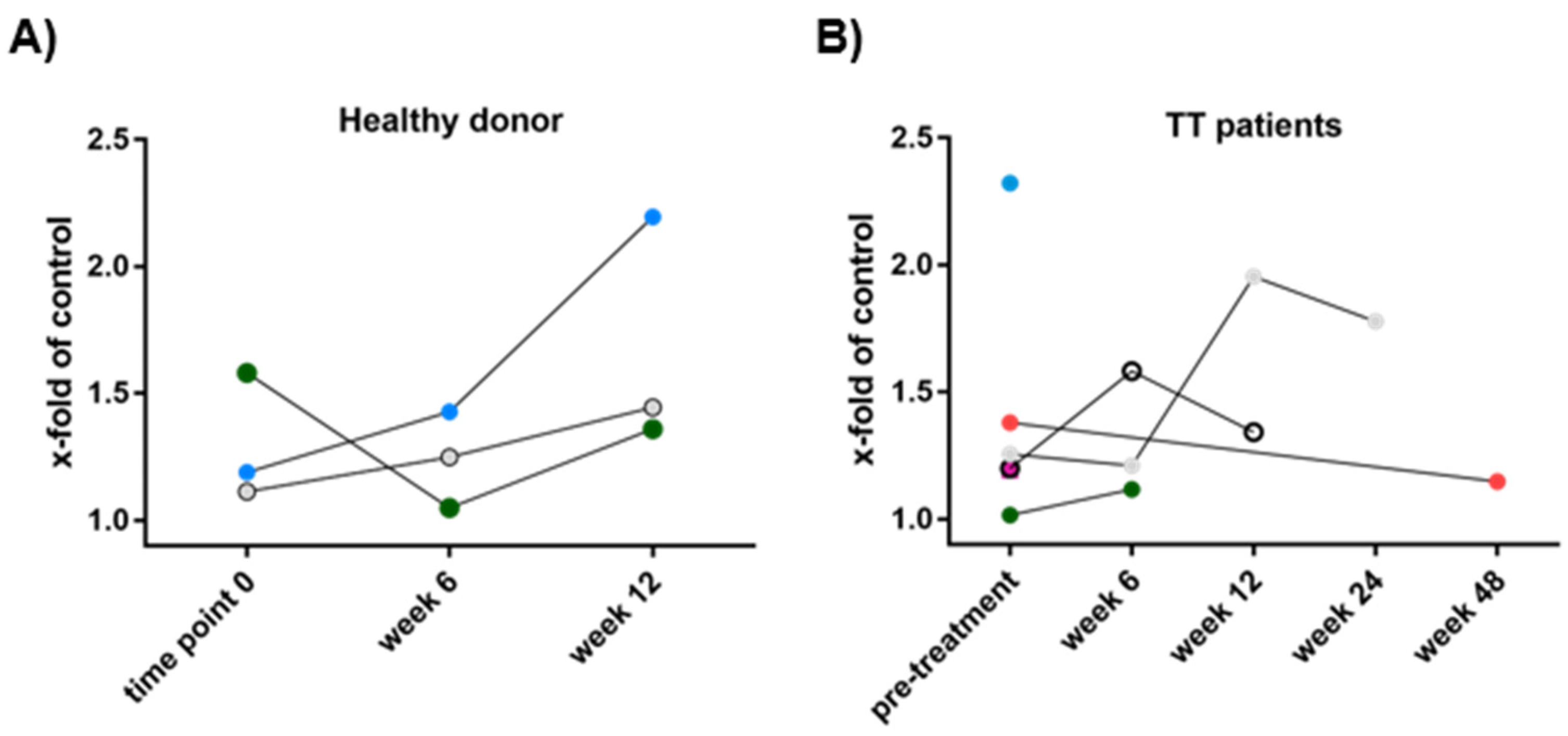
3.3. Patient-Derived Eosinophils Show a Comparable Phenotype to Eosinophils from Healthy Donors
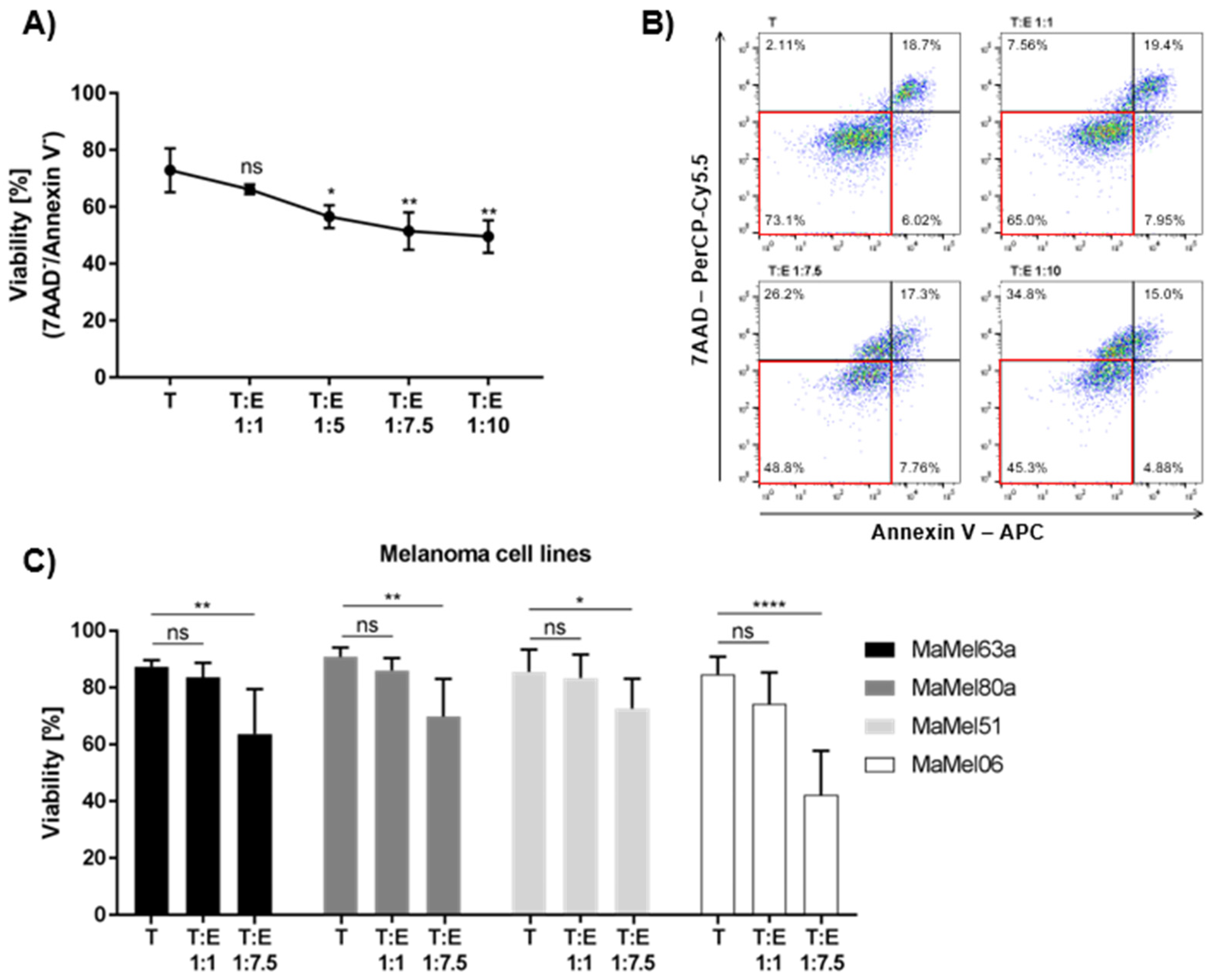
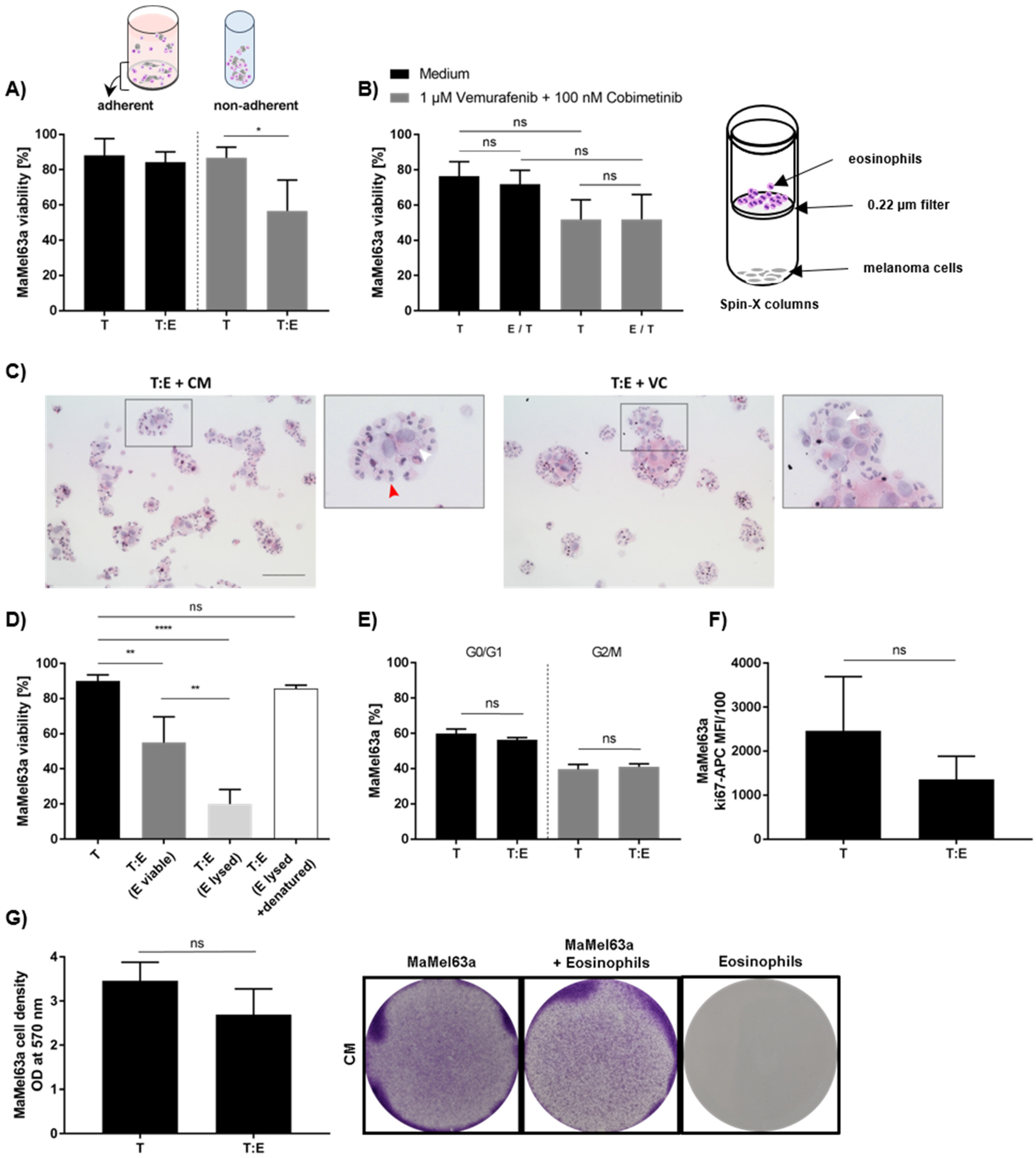
3.4. Susceptibility of Melanoma Cells to Eosinophil-Mediated Killing Is Volatile
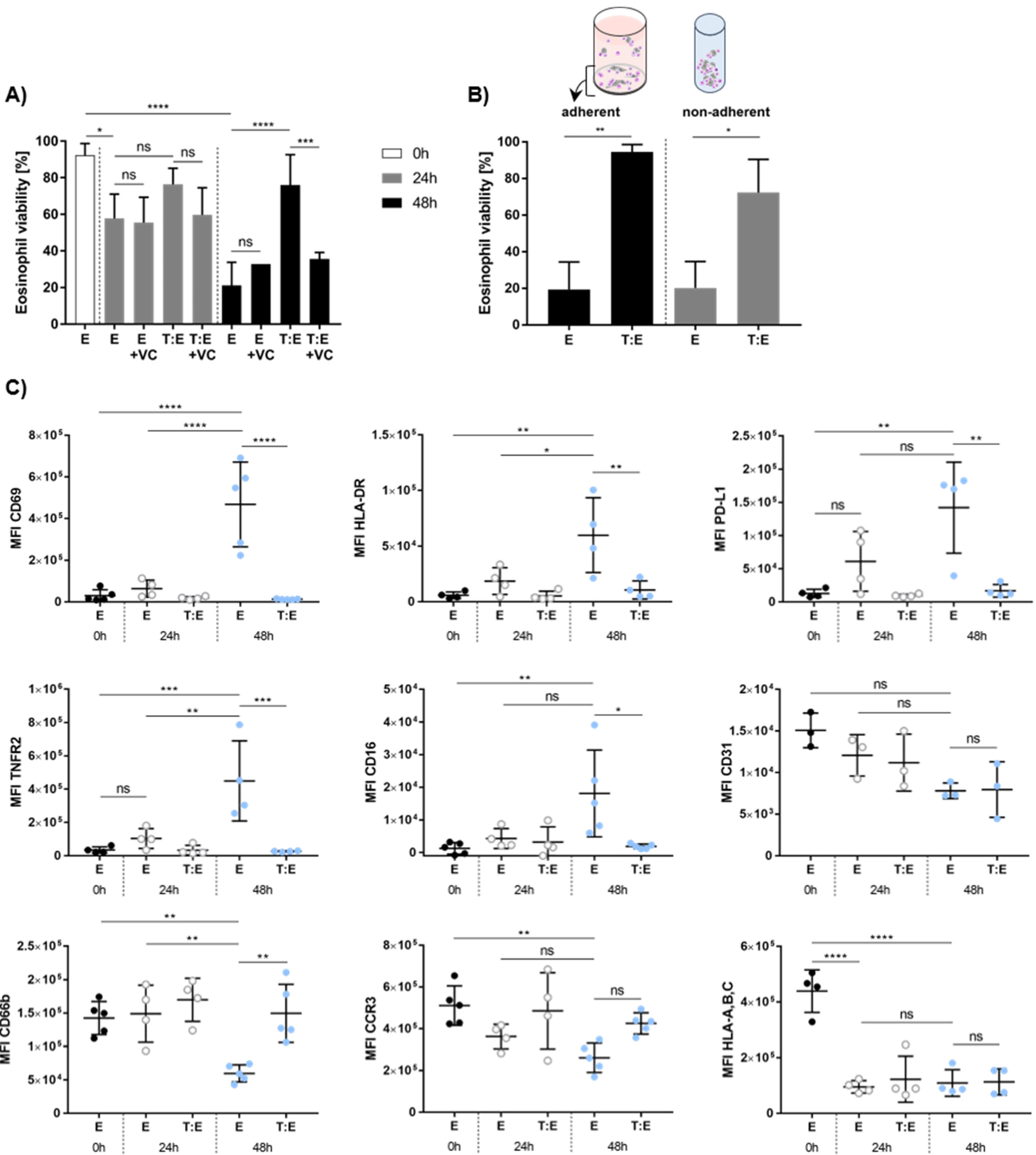
3.5. Melanoma Cells Affect Eosinophil Viability and Antigen Expression In Vitro
3.6. Eosinophils from Peripheral Blood Donor-Dependently Reduce Melanoma Cell Viability
3.7. Tumoricidal Effect of Eosinophils Depends on Culture Conditions and It Is an Active Process Requiring Cell–Cell Contact
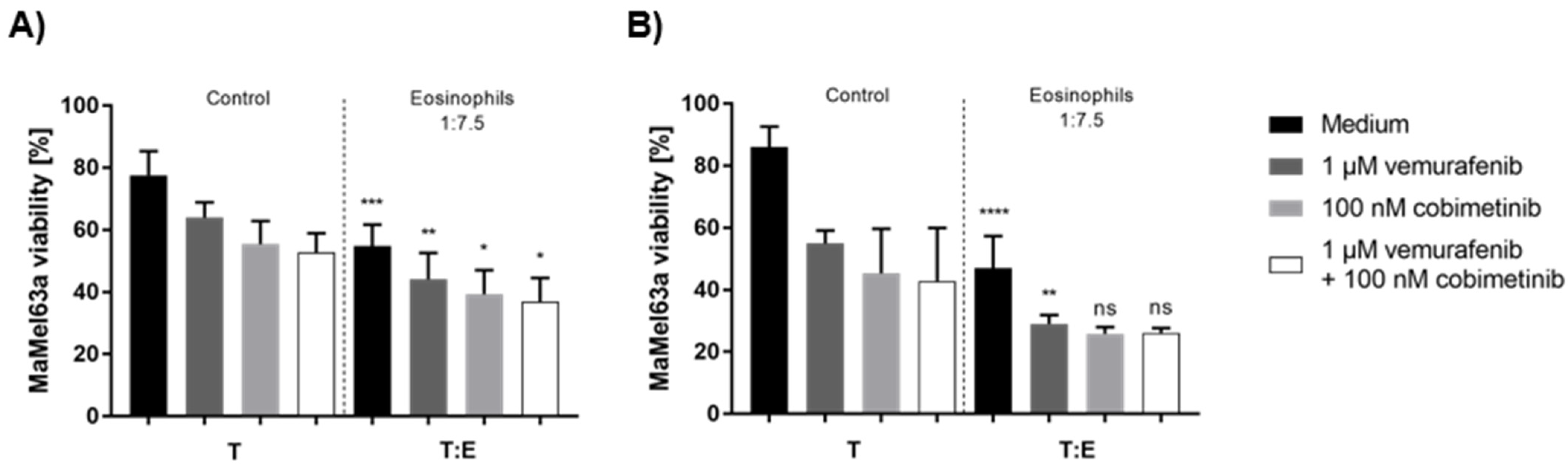
3.8. Tumoricidal Effects of Eosinophils and Systemic Therapy Are Additively Cytotoxic to Melanoma Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavinskiene, S.; Malakauskas, K.; Jeroch, J.; Hoppenot, D.; Sakalauskas, R. Functional activity of peripheral blood eosinophils in allergen-induced late-phase airway inflammation in asthma patients. J. Inflamm. 2015, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Domachowske, J.B. Eosinophils, eosinophil ribonucleases, and their role in host defense against respiratory virus pathogens. J. Leukoc. Biol. 2001, 70, 691–698. [Google Scholar] [PubMed]
- Na, H.J.; Hamilton, R.G.; Klion, A.D.; Bochner, B.S. Biomarkers of eosinophil involvement in allergic and eosinophilic diseases: Review of phenotypic and serum markers including a novel assay to quantify levels of soluble Siglec-8. J. Immunol. Methods 2012, 383, 39–46. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maizels, R.M.; Allen, J.E. Immunology. Eosinophils forestall obesity. Science 2011, 332, 186–187. [Google Scholar] [CrossRef]
- Carretero, R.; Sektioglu, I.M.; Garbi, N.; Salgado, O.C.; Beckhove, P.; Hammerling, G.J. Eosinophils orchestrate cancer rejection by normalizing tumor vessels and enhancing infiltration of CD8+ T cells. Nat. Immunol. 2015, 16, 609–617. [Google Scholar] [CrossRef]
- Huang, L.; Appleton, J.A. Eosinophils in Helminth Infection: Defenders and Dupes. Trends Parasitol. 2016, 32, 798–807. [Google Scholar] [CrossRef]
- Dricu, A.; Sergiu-Bogdan, C.; Brismar, K.; Biberfeld, P.; Andersson, L.C. A synthetic peptide derived from the human eosinophil-derived neurotoxin induces apoptosis in Kaposi’s sarcoma cells. Anticancer Res. 2004, 24, 1427–1432. [Google Scholar]
- Hamann, K.J.; Barker, R.L.; Ten, R.M.; Gleich, G.J. The molecular biology of eosinophil granule proteins. Int. Arch. Allergy Appl. Immunol. 1991, 94, 202–209. [Google Scholar] [CrossRef]
- Pereira, M.C.; Oliveira, D.T.; Kowalski, L.P. The role of eosinophils and eosinophil cationic protein in oral cancer: A review. Arch. Oral Biol. 2011, 56, 353–358. [Google Scholar] [CrossRef]
- Boix, E.; Carreras, E.; Nikolovski, Z.; Cuchillo, C.M.; Nogues, M.V. Identification and characterization of human eosinophil cationic protein by an epitope-specific antibody. J. Leukoc. Biol. 2001, 69, 1027–1035. [Google Scholar]
- Rosenberg, H.F.; Ackerman, S.J.; Tenen, D.G. Human eosinophil cationic protein. Molecular cloning of a cytotoxin and helminthotoxin with ribonuclease activity. J. Exp. Med. 1989, 170, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Young, J.D.; Peterson, C.G.; Venge, P.; Cohn, Z.A. Mechanism of membrane damage mediated by human eosinophil cationic protein. Nature 1986, 321, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Caruso, R.A.; Parisi, A.; Quattrocchi, E.; Scardigno, M.; Branca, G.; Parisi, C.; Luciano, R.; Paparo, D.; Fedele, F. Ultrastructural descriptions of heterotypic aggregation between eosinophils and tumor cells in human gastric carcinomas. Ultrastruct. Pathol. 2011, 35, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Simson, L.; Ellyard, J.I.; Dent, L.A.; Matthaei, K.I.; Rothenberg, M.E.; Foster, P.S.; Smyth, M.J.; Parish, C.R. Regulation of carcinogenesis by IL-5 and CCL11: A potential role for eosinophils in tumor immune surveillance. J. Immunol. 2007, 178, 4222–4229. [Google Scholar] [CrossRef]
- Kozar, I.; Margue, C.; Rothengatter, S.; Haan, C.; Kreis, S. Many ways to resistance: How melanoma cells evade targeted therapies. Biochim. Biophys. Acta (BBA) Rev. Cancer 2019, 1871, 313–322. [Google Scholar] [CrossRef]
- Kruckel, A.; Moreira, A.; Frohlich, W.; Schuler, G.; Heinzerling, L. Eosinophil-cationic protein—A novel liquid prognostic biomarker in melanoma. BMC Cancer 2019, 19, 207. [Google Scholar] [CrossRef]
- Kanda, A.; Fleury, S.; Kobayashi, Y.; Tomoda, K.; Julia, V.; Dombrowicz, D. Th2-activated eosinophils release Th1 cytokines that modulate allergic inflammation. Allergol. Int. Off. J. Jpn. Soc. Allergol. 2015, 64, S71–S73. [Google Scholar] [CrossRef]
- Legrand, F.; Driss, V.; Delbeke, M.; Loiseau, S.; Hermann, E.; Dombrowicz, D.; Capron, M. Human eosinophils exert TNF-alpha and granzyme A-mediated tumoricidal activity toward colon carcinoma cells. J Immunol. 2010, 185, 7443–7451. [Google Scholar] [CrossRef]
- Spencer, L.A.; Szela, C.T.; Perez, S.A.; Kirchhoffer, C.L.; Neves, J.S.; Radke, A.L.; Weller, P.F. Human eosinophils constitutively express multiple Th1, Th2, and immunoregulatory cytokines that are secreted rapidly and differentially. J. Leukoc. Biol. 2009, 85, 117–123. [Google Scholar] [CrossRef]
- Woerly, G.; Roger, N.; Loiseau, S.; Dombrowicz, D.; Capron, A.; Capron, M. Expression of CD28 and CD86 by human eosinophils and role in the secretion of type 1 cytokines (interleukin 2 and interferon gamma): Inhibition by immunoglobulin a complexes. J. Exp. Med. 1999, 190, 487–495. [Google Scholar] [CrossRef]
- Legrand, F.; Driss, V.; Woerly, G.; Loiseau, S.; Hermann, E.; Fournie, J.J.; Heliot, L.; Mattot, V.; Soncin, F.; Gougeon, M.L.; et al. A functional gammadeltaTCR/CD3 complex distinct from gammadeltaT cells is expressed by human eosinophils. PLoS ONE 2009, 4, e5926. [Google Scholar] [CrossRef] [PubMed]
- Reinbach, G. Über das Verhalten der Leukocyten bei malignen Tumoren. Arch. Für Klin. Chir. 1893, 46, 486–562. [Google Scholar]
- Caruso, R.A.; Ieni, A.; Fedele, F.; Zuccala, V.; Riccardo, M.; Parisi, E.; Parisi, A. Degranulation patterns of eosinophils in advanced gastric carcinoma: An electron microscopic study. Ultrastruct. Pathol. 2005, 29, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Ieni, A.; Barresi, V.; Branca, G.; Alberto Caruso, R.; Tuccari, G. Mast Cell Interaction with Neutrophils in Human Gastric Carcinomas: Ultrastructural Observations. Anal. Cell. Pathol. 2016, 2016, 6891971. [Google Scholar] [CrossRef]
- Gatault, S.; Delbeke, M.; Driss, V.; Sarazin, A.; Dendooven, A.; Kahn, J.E.; Lefevre, G.; Capron, M. IL-18 Is Involved in Eosinophil-Mediated Tumoricidal Activity against a Colon Carcinoma Cell Line by Upregulating LFA-1 and ICAM-1. J. Immunol. 2015, 195, 2483–2492. [Google Scholar] [CrossRef]
- Reichman, H.; Itan, M.; Rozenberg, P.; Yarmolovski, T.; Brazowski, E.; Varol, C.; Gluck, N.; Shapira, S.; Arber, N.; Qimron, U.; et al. Activated Eosinophils Exert Antitumorigenic Activities in Colorectal Cancer. Cancer Immunol. Res. 2019, 7, 388–400. [Google Scholar] [CrossRef]
- Andreone, S.; Spadaro, F.; Buccione, C.; Mancini, J.; Tinari, A.; Sestili, P.; Gambardella, A.R.; Lucarini, V.; Ziccheddu, G.; Parolini, I.; et al. IL-33 Promotes CD11b/CD18-Mediated Adhesion of Eosinophils to Cancer Cells and Synapse-Polarized Degranulation Leading to Tumor Cell Killing. Cancers 2019, 11, 1664. [Google Scholar] [CrossRef]
- Feldbausch, F. The occurrence of eosinophilic leukocytes in tumours. Arch. Pathol. Anat. Physiol. Klin. Med. 1900, 161, 1–18. [Google Scholar] [CrossRef]
- von Wasielewski, R.; Seth, S.; Franklin, J.; Fischer, R.; Hubner, K.; Hansmann, M.L.; Diehl, V.; Georgii, A. Tissue eosinophilia correlates strongly with poor prognosis in nodular sclerosing Hodgkin’s disease, allowing for known prognostic factors. Blood 2000, 95, 1207–1213. [Google Scholar] [CrossRef]
- Horiuchi, K.; Mishima, K.; Ohsawa, M.; Sugimura, M.; Aozasa, K. Prognostic factors for well-differentiated squamous cell carcinoma in the oral cavity with emphasis on immunohistochemical evaluation. J. Surg. Oncol. 1993, 53, 92–96. [Google Scholar] [CrossRef]
- Sakkal, S.; Miller, S.; Apostolopoulos, V.; Nurgali, K. Eosinophils in Cancer: Favourable or Unfavourable? Curr. Med. Chem. 2016, 23, 650–666. [Google Scholar] [CrossRef] [PubMed]
- van Driel, W.J.; Hogendoorn, P.C.; Jansen, F.W.; Zwinderman, A.H.; Trimbos, J.B.; Fleuren, G.J. Tumor-associated eosinophilic infiltrate of cervical cancer is indicative for a less effective immune response. Hum. Pathol. 1996, 27, 904–911. [Google Scholar] [CrossRef]
- Luna-More, S.; Florez, P.; Ayala, A.; Diaz, F.; Santos, A. Neutral and acid mucins and eosinophil and argyrophil crystalloids in carcinoma and atypical adenomatous hyperplasia of the prostate. Pathol. Res. Pract. 1997, 193, 291–298. [Google Scholar] [CrossRef]
- McNeel, D.G.; Gardner, T.A.; Higano, C.S.; Kantoff, P.W.; Small, E.J.; Wener, M.H.; Sims, R.B.; DeVries, T.; Sheikh, N.A.; Dreicer, R. A transient increase in eosinophils is associated with prolonged survival in men with metastatic castration-resistant prostate cancer who receive sipuleucel-T. Cancer Immunol. Res. 2014, 2, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Prizment, A.E.; Anderson, K.E.; Visvanathan, K.; Folsom, A.R. Inverse association of eosinophil count with colorectal cancer incidence: Atherosclerosis risk in communities study. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1861–1864. [Google Scholar] [CrossRef]
- Prizment, A.E.; Vierkant, R.A.; Smyrk, T.C.; Tillmans, L.S.; Lee, J.J.; Sriramarao, P.; Nelson, H.H.; Lynch, C.F.; Thibodeau, S.N.; Church, T.R.; et al. Tumor eosinophil infiltration and improved survival of colorectal cancer patients: Iowa Women’s Health Study. Mod. Pathol. 2016, 29, 516–527. [Google Scholar] [CrossRef]
- Taghizadeh, N.; Vonk, J.M.; Boezen, H.M. Peripheral blood eosinophil counts and risk of colorectal cancer mortality in a large general population-based cohort study. J. Clin. Oncol. 2011, 29, 1583. [Google Scholar] [CrossRef]
- Ferrucci, P.F.; Gandini, S.; Cocorocchio, E.; Pala, L.; Baldini, F.; Mosconi, M.; Antonini Cappellini, G.C.; Albertazzi, E.; Martinoli, C. Baseline relative eosinophil count as a predictive biomarker for ipilimumab treatment in advanced melanoma. Oncotarget 2017, 8, 79809–79815. [Google Scholar] [CrossRef]
- Moreira, A.; Leisgang, W.; Schuler, G.; Heinzerling, L. Eosinophilic count as a biomarker for prognosis of melanoma patients and its importance in the response to immunotherapy. Immunotherapy 2017, 9, 115–121. [Google Scholar] [CrossRef]
- Bernard-Tessier, A.; Jeanville, P.; Champiat, S.; Lazarovici, J.; Voisin, A.L.; Mateus, C.; Lambotte, O.; Annereau, M.; Michot, J.M. Immune-related eosinophilia induced by anti-programmed death 1 or death-ligand 1 antibodies. Eur. J. Cancer 2017, 81, 135–137. [Google Scholar] [CrossRef]
- Delyon, J.; Mateus, C.; Lefeuvre, D.; Lanoy, E.; Zitvogel, L.; Chaput, N.; Roy, S.; Eggermont, A.M.; Routier, E.; Robert, C. Experience in daily practice with ipilimumab for the treatment of patients with metastatic melanoma: An early increase in lymphocyte and eosinophil counts is associated with improved survival. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2013, 24, 1697–1703. [Google Scholar] [CrossRef]
- Martens, A.; Wistuba-Hamprecht, K.; Geukes Foppen, M.; Yuan, J.; Postow, M.A.; Wong, P.; Romano, E.; Khammari, A.; Dreno, B.; Capone, M.; et al. Baseline Peripheral Blood Biomarkers Associated with Clinical Outcome of Advanced Melanoma Patients Treated with Ipilimumab. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 2908–2918. [Google Scholar] [CrossRef] [PubMed]
- Weide, B.; Martens, A.; Hassel, J.C.; Berking, C.; Postow, M.A.; Bisschop, K.; Simeone, E.; Mangana, J.; Schilling, B.; Di Giacomo, A.M.; et al. Baseline Biomarkers for Outcome of Melanoma Patients Treated with Pembrolizumab. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 5487–5496. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.O.; Downey, S.G.; Klapper, J.A.; Yang, J.C.; Sherry, R.M.; Royal, R.E.; Kammula, U.S.; Hughes, M.S.; Restifo, N.P.; Levy, C.L.; et al. Treatment of metastatic melanoma using interleukin-2 alone or in conjunction with vaccines. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 5610–5618. [Google Scholar] [CrossRef] [PubMed]
- Schindler, K.; Harmankaya, K.; Postow, M.; Frantal, S.; Bello, D.; Ariyan, C.; Michielin, O.; Hoeller, C.; Pehamberger, H.; Wolchok, J. Pretreatment levels of absolute and relative eosinophil count to improve overall survival (OS) in patients with metastatic melanoma under treatment with ipilimumab, an anti CTLA-4 antibody. J. Clin. Oncol. 2013, 31, 9024. [Google Scholar] [CrossRef]
- Swami, U.; Monga, V.; Knutson, T.; Bossler, A.D.; Zakharia, Y.; Milhem, M.M. Prognostic markers for progression free survival (PFS) to anti PD-1 therapies in metastatic melanoma. J. Clin. Oncol. 2018, 36, e21527. [Google Scholar] [CrossRef]
- Gaba, L.; Victoria, I.; Pineda, E.; Fernandez, A.; Aya, F.; Prat, A.; Arance, A.M. Changes in blood eosinophilia during anti-PD1 therapy as a predictor of long term disease control in metastatic melanoma. J. Clin. Oncol. 2015, 33, 9069. [Google Scholar] [CrossRef]
- Simon, S.C.S.; Hu, X.; Panten, J.; Grees, M.; Renders, S.; Thomas, D.; Weber, R.; Schulze, T.J.; Utikal, J.; Umansky, V. Eosinophil accumulation predicts response to melanoma treatment with immune checkpoint inhibitors. Oncoimmunology 2020, 9, 1727116. [Google Scholar] [CrossRef]
- Ugurel, S.; Thirumaran, R.K.; Bloethner, S.; Gast, A.; Sucker, A.; Mueller-Berghaus, J.; Rittgen, W.; Hemminki, K.; Becker, J.C.; Kumar, R.; et al. B-RAF and N-RAS mutations are preserved during short time in vitro propagation and differentially impact prognosis. PLoS ONE 2007, 2, e236. [Google Scholar] [CrossRef]
- Mattes, J.; Hulett, M.; Xie, W.; Hogan, S.; Rothenberg, M.E.; Foster, P.; Parish, C. Immunotherapy of cytotoxic T cell-resistant tumors by T helper 2 cells: An eotaxin and STAT6-dependent process. J. Exp. Med. 2003, 197, 387–393. [Google Scholar] [CrossRef]
- Gershenwald, J.E.; Scolyer, R.A.; Hess, K.R.; Sondak, V.K.; Long, G.V.; Ross, M.I.; Lazar, A.J.; Faries, M.B.; Kirkwood, J.M.; McArthur, G.A.; et al. Melanoma staging: Evidence-based changes in the American Joint Committee on Cancer eighth edition cancer staging manual. CA Cancer J. Clin. 2017, 67, 472–492. [Google Scholar] [CrossRef]
- Bochner, B.S. Siglec-8 on human eosinophils and mast cells, and Siglec-F on murine eosinophils, are functionally related inhibitory receptors. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2009, 39, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Lucarini, V.; Marone, G.; Mattei, F.; Marone, G.; Schiavoni, G. Eosinophils: The unsung heroes in cancer? Oncoimmunology 2018, 7, e1393134. [Google Scholar] [CrossRef] [PubMed]
- Lucey, D.R.; Nicholson-Weller, A.; Weller, P.F. Mature human eosinophils have the capacity to express HLA-DR. Proc. Natl. Acad. Sci. USA 1989, 86, 1348–1351. [Google Scholar] [CrossRef] [PubMed]
- Amschler, K.; Kossmann, E.; Erpenbeck, L.; Kruss, S.; Schill, T.; Schon, M.; Mockel, S.M.C.; Spatz, J.P.; Schon, M.P. Nanoscale Tuning of VCAM-1 Determines VLA-4-Dependent Melanoma Cell Plasticity on RGD Motifs. Mol. Cancer Res. MCR 2018, 16, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Giommoni, E.; Giorgione, R.; Paderi, A.; Pellegrini, E.; Gambale, E.; Marini, A.; Antonuzzo, A.; Marconcini, R.; Roviello, G.; Matucci-Cerinic, M.; et al. Eosinophil Count as Predictive Biomarker of Immune-Related Adverse Events (irAEs) in Immune Checkpoint Inhibitors (ICIs) Therapies in Oncological Patients. Immuno 2021, 1, 253–263. [Google Scholar] [CrossRef]
- Scanvion, Q.; Bene, J.; Gautier, S.; Grandvuillemin, A.; Le Beller, C.; Chenaf, C.; Etienne, N.; Brousseau, S.; Cortot, A.B.; Mortier, L.; et al. Moderate-to-severe eosinophilia induced by treatment with immune checkpoint inhibitors: 37 cases from a national reference center for hypereosinophilic syndromes and the French pharmacovigilance database. Oncoimmunology 2020, 9, 1722022. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, R.; Worschech, A.; Cardone, M.; Jones, Y.; Gyulai, Z.; Dai, R.M.; Wang, E.; Ma, W.; Haines, D.; O’HUigin, C.; et al. MyD88-mediated signaling prevents development of adenocarcinomas of the colon: Role of interleukin 18. J. Exp. Med. 2010, 207, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Tripathy, D.; Frenkel, E.P.; Shete, S.; Naftalis, E.Z.; Huth, J.F.; Beitsch, P.D.; Leitch, M.; Hoover, S.; Euhus, D.; et al. Circulating tumor cells in patients with breast cancer dormancy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 8152–8162. [Google Scholar] [CrossRef]
- Muller, V.; Stahmann, N.; Riethdorf, S.; Rau, T.; Zabel, T.; Goetz, A.; Janicke, F.; Pantel, K. Circulating tumor cells in breast cancer: Correlation to bone marrow micrometastases, heterogeneous response to systemic therapy and low proliferative activity. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 3678–3685. [Google Scholar] [CrossRef]
- Seyfried, T.N.; Huysentruyt, L.C. On the origin of cancer metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Ikutani, M.; Yanagibashi, T.; Ogasawara, M.; Tsuneyama, K.; Yamamoto, S.; Hattori, Y.; Kouro, T.; Itakura, A.; Nagai, Y.; Takaki, S.; et al. Identification of innate IL-5-producing cells and their role in lung eosinophil regulation and antitumor immunity. J. Immunol. 2012, 188, 703–713. [Google Scholar] [CrossRef]
- Lucarini, V.; Ziccheddu, G.; Macchia, I.; La Sorsa, V.; Peschiaroli, F.; Buccione, C.; Sistigu, A.; Sanchez, M.; Andreone, S.; D’Urso, M.T.; et al. IL-33 restricts tumor growth and inhibits pulmonary metastasis in melanoma-bearing mice through eosinophils. Oncoimmunology 2017, 6, e1317420. [Google Scholar] [CrossRef] [PubMed]
- Reichman, H.; Karo-Atar, D.; Munitz, A. Emerging Roles for Eosinophils in the Tumor Microenvironment. Trends Cancer 2016, 2, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Marsavela, G.; Aya-Bonilla, C.A.; Warkiani, M.E.; Gray, E.S.; Ziman, M. Melanoma circulating tumor cells: Benefits and challenges required for clinical application. Cancer Lett. 2018, 424, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Terada, A.; Kita, H. CD66b regulates adhesion and activation of human eosinophils. J. Immunol. 2007, 179, 8454–8462. [Google Scholar] [CrossRef]
- Hansel, T.T.; De Vries, I.J.; Carballido, J.M.; Braun, R.K.; Carballido-Perrig, N.; Rihs, S.; Blaser, K.; Walker, C. Induction and function of eosinophil intercellular adhesion molecule-1 and HLA-DR. J. Immunol. 1992, 149, 2130–2136. [Google Scholar]
- Johansson, M.W. Activation states of blood eosinophils in asthma. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2014, 44, 482–498. [Google Scholar] [CrossRef]
- Weller, P.F.; Rand, T.H.; Barrett, T.; Elovic, A.; Wong, D.T.; Finberg, R.W. Accessory cell function of human eosinophils. HLA-DR-dependent, MHC-restricted antigen-presentation and IL-1 alpha expression. J. Immunol. 1993, 150, 2554–2562. [Google Scholar]
- Cormier, S.A.; Taranova, A.G.; Bedient, C.; Nguyen, T.; Protheroe, C.; Pero, R.; Dimina, D.; Ochkur, S.I.; O’Neill, K.; Colbert, D.; et al. Pivotal Advance: Eosinophil infiltration of solid tumors is an early and persistent inflammatory host response. J. Leukoc. Biol. 2006, 79, 1131–1139. [Google Scholar] [CrossRef]
- Spencer, L.A.; Bonjour, K.; Melo, R.C.; Weller, P.F. Eosinophil secretion of granule-derived cytokines. Front. Immunol. 2014, 5, 496. [Google Scholar] [CrossRef] [PubMed]
- Radonjic-Hoesli, S.; Wang, X.; de Graauw, E.; Stoeckle, C.; Styp-Rekowska, B.; Hlushchuk, R.; Simon, D.; Spaeth, P.J.; Yousefi, S.; Simon, H.U. Adhesion-induced eosinophil cytolysis requires the receptor-interacting protein kinase 3 (RIPK3)-mixed lineage kinase-like (MLKL) signaling pathway, which is counterregulated by autophagy. J. Allergy Clin. Immunol. 2017, 140, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.D.; Cannistra, S.A.; Sullivan, R.; Demetri, G.D.; Ernst, T.J.; Kanakura, Y. The biology of GM-CSF: Regulation of production and interaction with its receptor. Int. J. Cell Cloning 1990, 8 (Suppl. 1), 35–44; discussion 44–45. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. GM-CSF in inflammation. J. Exp. Med. 2020, 217, e20190945. [Google Scholar] [CrossRef]
- Dillman, R.O. An update on GM-CSF and its potential role in melanoma management. Melanoma Manag. 2020, 7, MMT49. [Google Scholar] [CrossRef]
- Zhang, S.; Mohammed, Q.; Burbidge, A.; Morland, C.M.; Roche, W.R. Cell cultures from bronchial subepithelial myofibroblasts enhance eosinophil survival in vitro. Eur. Respir. J. 1996, 9, 1839–1846. [Google Scholar] [CrossRef]
- Curran, C.S.; Evans, M.D.; Bertics, P.J. GM-CSF production by glioblastoma cells has a functional role in eosinophil survival, activation, and growth factor production for enhanced tumor cell proliferation. J. Immunol. 2011, 187, 1254–1263. [Google Scholar] [CrossRef]
- Solomon, A.; Shmilowich, R.; Shasha, D.; Frucht-Pery, J.; Pe’er, J.; Bonini, S.; Levi-Schaffer, F. Conjunctival fibroblasts enhance the survival and functional activity of peripheral blood eosinophils in vitro. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1038–1044. [Google Scholar]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- De Lima, P.O.; Dos Santos, F.V.; Oliveira, D.T.; Carvalho de Figueiredo, R.; Pereira, M.C. Effect of eosinophil cationic protein on human oral squamous carcinoma cell viability. Mol. Clin. Oncol. 2015, 3, 353–356. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Patients | ||
|---|---|---|---|
| Age | median (range) | 71 years (27–93) | |
| % | |||
| Individual Patients | 206 | 100 | |
| Sex | male | 97 | 47.1 |
| female | 79 | 38.3 | |
| unknown 1 | 30 | 14.6 | |
| Stage | III | 15 | 7.3 |
| IV | 191 | 92.7 | |
| M-Category | M1a | 23 | 11.2 |
| M1b | 51 | 24.8 | |
| M1c | 75 | 36.4 | |
| M1d | 42 | 20.4 | |
| First-Line Therapy | yes | 180 | 87.4 |
| no | 26 | 12.6 | |
| Therapy after Study Inclusion | anti-PD-1 | 86 | 41.8 |
| anit-PD-1 + anti-CTLA-4 | 26 | 12.6 | |
| BRAFi + MEKi 2 | 94 | 45.6 | |
| LDH | >1× ULN 3 | 68 | 33.0 |
| <1× ULN 3 | 134 | 65.1 | |
| missing | 4 | 1.9 | |
| Variables | Donor | |||
|---|---|---|---|---|
| % | ||||
| Individual Donors | 31 | 100 | ||
| Patient | Stage | I/II | 6 | 19.4 |
| IV 2 | 13 | 41.9 | ||
| Healthy Donor | 12 | 38.7 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wendlinger, S.; Wohlfarth, J.; Kreft, S.; Siedel, C.; Kilian, T.; Dischinger, U.; Heppt, M.V.; Wistuba-Hamprecht, K.; Meier, F.; Goebeler, M.; et al. Blood Eosinophils Are Associated with Efficacy of Targeted Therapy in Patients with Advanced Melanoma. Cancers 2022, 14, 2294. https://doi.org/10.3390/cancers14092294
Wendlinger S, Wohlfarth J, Kreft S, Siedel C, Kilian T, Dischinger U, Heppt MV, Wistuba-Hamprecht K, Meier F, Goebeler M, et al. Blood Eosinophils Are Associated with Efficacy of Targeted Therapy in Patients with Advanced Melanoma. Cancers. 2022; 14(9):2294. https://doi.org/10.3390/cancers14092294
Chicago/Turabian StyleWendlinger, Simone, Jonas Wohlfarth, Sophia Kreft, Claudia Siedel, Teresa Kilian, Ulrich Dischinger, Markus V. Heppt, Kilian Wistuba-Hamprecht, Friedegund Meier, Matthias Goebeler, and et al. 2022. "Blood Eosinophils Are Associated with Efficacy of Targeted Therapy in Patients with Advanced Melanoma" Cancers 14, no. 9: 2294. https://doi.org/10.3390/cancers14092294
APA StyleWendlinger, S., Wohlfarth, J., Kreft, S., Siedel, C., Kilian, T., Dischinger, U., Heppt, M. V., Wistuba-Hamprecht, K., Meier, F., Goebeler, M., Schadendorf, D., Gesierich, A., Kosnopfel, C., & Schilling, B. (2022). Blood Eosinophils Are Associated with Efficacy of Targeted Therapy in Patients with Advanced Melanoma. Cancers, 14(9), 2294. https://doi.org/10.3390/cancers14092294