
Terpenoid-Mediated Targeting of STAT3 Signaling in Cancer: An Overview of Preclinical Studies




Abstract
1. Introduction
2. Terpenoid-Mediated Cancer Chemoprevention
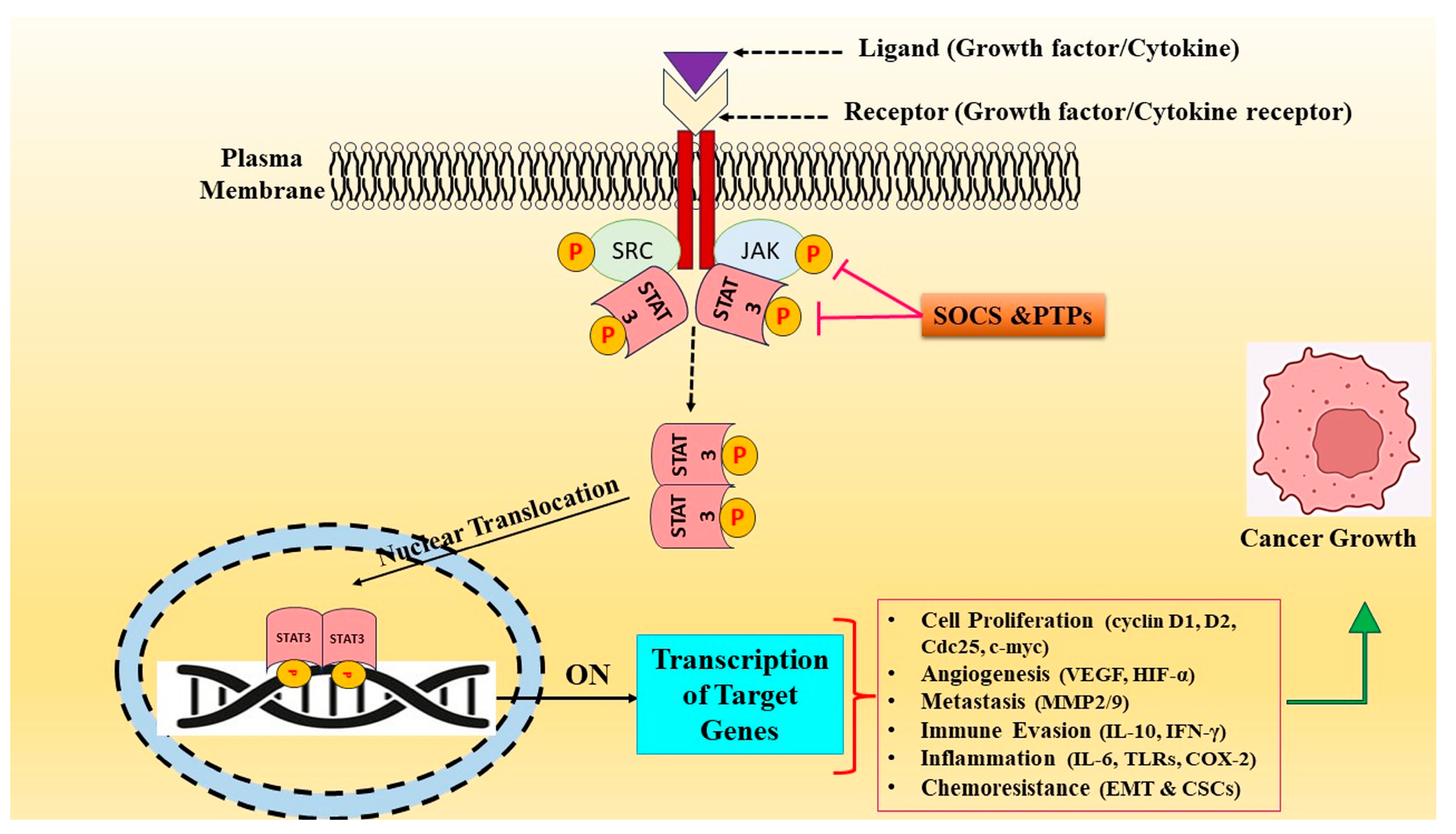
3. STAT3 Signaling Cascade and Tumorigenesis
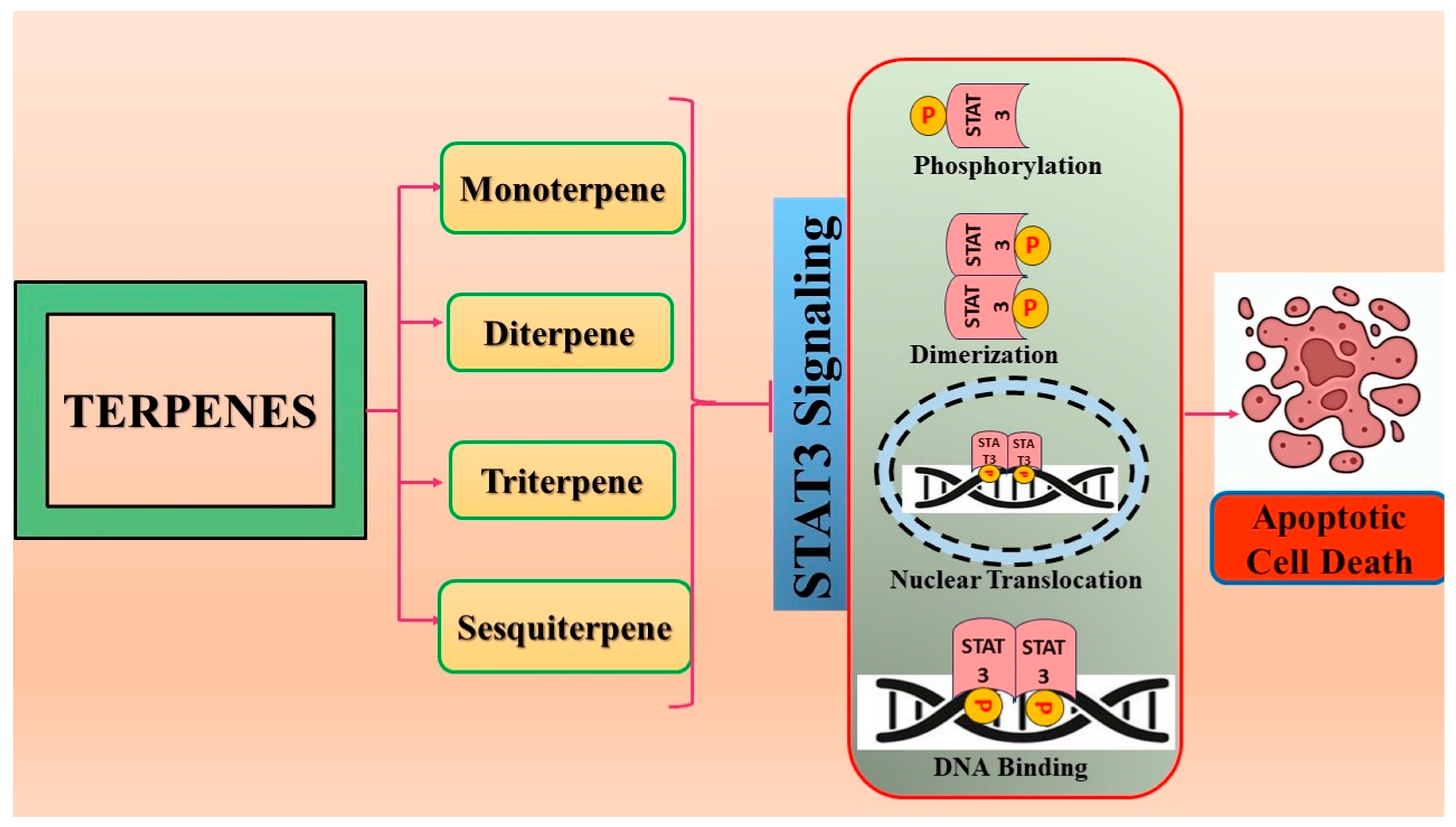
4. Terpenoids: Modulators of STAT3 Signaling
4.1. Monoterpene
Thymoquinone
4.2. Diterpene
4.2.1. Andrographolide
4.2.2. Cryptotanshinone
4.2.3. Oridonin
4.3. Triterpene
4.3.1. Brusatol
4.3.2. Betulinic Acid
4.3.3. Celastrol
4.3.4. Cucurbitacin B
4.3.5. Cucurbitacin E
4.3.6. Cucurbitacin I
4.3.7. Ursolic Acid
4.4. Sesquiterpene
4.4.1. Alantolactone
4.4.2. β-Caryophyllene Oxide
4.4.3. Dihydroartemisinin
4.4.4. Parthenolide
4.4.5. γ-Tocotrienol
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Li, Z.; Zhang, M.; Bao, M.; He, B.; Zhou, X. LncRNA FAS-AS1 upregulated by its genetic variation rs6586163 promotes cell apoptosis in nasopharyngeal carcinoma through regulating mitochondria function and Fas splicing. Sci. Rep. 2023, 13, 8218. [Google Scholar] [CrossRef] [PubMed]
- Taga, T.; Hibi, M.; Hirata, Y.; Yamasaki, K.; Yasukawa, K.; Matsuda, T.; Hirano, T.; Kishimoto, T. Interleukin-6 triggers the association of its receptor with a possible signal transducer, gp130. Cell 1989, 58, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target. Ther. 2021, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jove, R. The STATs of cancer--new molecular targets come of age. Nat. Rev. Cancer 2004, 4, 97–105. [Google Scholar] [CrossRef]
- Molenda, S.; Sikorska, A.; Florczak, A.; Lorenc, P.; Dams-Kozlowska, H. Oligonucleotide-Based Therapeutics for STAT3 Targeting in Cancer-Drug Carriers Matter. Cancers 2023, 15, 5647. [Google Scholar] [CrossRef]
- Herrmann, A.; Kortylewski, M.; Kujawski, M.; Zhang, C.; Reckamp, K.; Armstrong, B.; Wang, L.; Kowolik, C.; Deng, J.; Figlin, R.; et al. Targeting Stat3 in the myeloid compartment drastically improves the in vivo antitumor functions of adoptively transferred T cells. Cancer Res. 2010, 70, 7455–7464. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, Y.; Wang, S.; Shen, Q.; Zhou, X. The role of STAT3 in leading the crosstalk between human cancers and the immune system. Cancer Lett. 2018, 415, 117–128. [Google Scholar] [CrossRef]
- Chen, R.Y.; Yen, C.J.; Liu, Y.W.; Guo, C.G.; Weng, C.Y.; Lai, C.H.; Wang, J.M.; Lin, Y.J.; Hung, L.Y. CPAP promotes angiogenesis and metastasis by enhancing STAT3 activity. Cell Death Differ. 2020, 27, 1259–1273. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, X.; Liu, Y.; Singhal, M.; Gürkaşlar, C.; Valls, A.F.; Lei, Y.; Hu, W.; Schermann, G.; Adler, H.; et al. STAT3-YAP/TAZ signaling in endothelial cells promotes tumor angiogenesis. Sci. Signal. 2021, 14, eabj8393. [Google Scholar] [CrossRef]
- Lee, H.; Zhang, P.; Herrmann, A.; Yang, C.; Xin, H.; Wang, Z.; Hoon, D.S.; Forman, S.J.; Jove, R.; Riggs, A.D.; et al. Acetylated STAT3 is crucial for methylation of tumor-suppressor gene promoters and inhibition by resveratrol results in demethylation. Proc. Natl. Acad. Sci. USA 2012, 109, 7765–7769. [Google Scholar] [CrossRef] [PubMed]
- Tolomeo, M.; Cascio, A. The Multifaced Role of STAT3 in Cancer and Its Implication for Anticancer Therapy. Int. J. Mol. Sci. 2021, 22, 603. [Google Scholar] [CrossRef]
- Hossain, D.M.; Dos Santos, C.; Zhang, Q.; Kozlowska, A.; Liu, H.; Gao, C.; Moreira, D.; Swiderski, P.; Jozwiak, A.; Kline, J.; et al. Leukemia cell-targeted STAT3 silencing and TLR9 triggering generate systemic antitumor immunity. Blood 2014, 123, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Eyking, A.; Ey, B.; Rünzi, M.; Roig, A.I.; Reis, H.; Schmid, K.W.; Gerken, G.; Podolsky, D.K.; Cario, E. Toll-like receptor 4 variant D299G induces features of neoplastic progression in Caco-2 intestinal cells and is associated with advanced human colon cancer. Gastroenterology 2011, 141, 2154–2165. [Google Scholar] [CrossRef] [PubMed]
- Tye, H.; Kennedy, C.L.; Najdovska, M.; McLeod, L.; McCormack, W.; Hughes, N.; Dev, A.; Sievert, W.; Ooi, C.H.; Ishikawa, T.O.; et al. STAT3-driven upregulation of TLR2 promotes gastric tumorigenesis independent of tumor inflammation. Cancer Cell 2012, 22, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Fehri, E.; Ennaifer, E.; Bel Haj Rhouma, R.; Ardhaoui, M.; Boubaker, S. TLR9 and Glioma: Friends or Foes? Cells 2022, 12, 152. [Google Scholar] [CrossRef]
- Li, L.; Wang, S.; Zhou, W. Balance Cell Apoptosis and Pyroptosis of Caspase-3-Activating Chemotherapy for Better Antitumor Therapy. Cancers 2022, 15, 26. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Albonici, L.; Focaccetti, C.; Ciuffa, S.; Fazi, S.; Cifaldi, L.; Miele, M.T.; De Maio, F.; Tresoldi, I.; Manzari, V.; et al. Polyphenol-Mediated Autophagy in Cancer: Evidence of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2020, 21, 6635. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F. A mechanistic review of the anticancer potential of hesperidin, a natural flavonoid from citrus fruits. Nutr. Res. 2021, 92, 21–31. [Google Scholar] [CrossRef]
- Muhammad, N.; Usmani, D.; Tarique, M.; Naz, H.; Ashraf, M.; Raliya, R.; Tabrez, S.; Zughaibi, T.A.; Alsaieedi, A.; Hakeem, I.J.; et al. The Role of Natural Products and Their Multitargeted Approach to Treat Solid Cancer. Cells 2022, 11, 2209. [Google Scholar] [CrossRef]
- Rahman, M.M.; Sarker, M.T.; Alam Tumpa, M.A.; Yamin, M.; Islam, T.; Park, M.N.; Islam, M.R.; Rauf, A.; Sharma, R.; Cavalu, S.; et al. Exploring the recent trends in perturbing the cellular signaling pathways in cancer by natural products. Front. Pharmacol. 2022, 13, 950109. [Google Scholar] [CrossRef]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Ge, J.; Liu, Z.; Zhong, Z.; Wang, L.; Zhuo, X.; Li, J.; Jiang, X.; Ye, X.Y.; Xie, T.; Bai, R. Natural terpenoids with anti-inflammatory activities: Potential leads for anti-inflammatory drug discovery. Bioorg. Chem. 2022, 124, 105817. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S.; Daglia, M.; Rengasamy, K.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacol. Res. 2020, 157, 104830. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Gonzalez, P.; Garza-Treviño, E.N.; de la Garza Kalife, D.A.; Quiroz Reyes, A.; Hernández-Tobías, E.A. Bioactive Compounds of Dietary Origin and Their Influence on Colorectal Cancer as Chemoprevention. Life 2023, 13, 1977. [Google Scholar] [CrossRef] [PubMed]
- Sporn, M.B.; Suh, N. Chemoprevention: An essential approach to controlling cancer. Nat. Rev. Cancer 2002, 2, 537–543. [Google Scholar] [CrossRef]
- Steward, W.P.; Brown, K. Cancer chemoprevention: A rapidly evolving field. Br. J. Cancer 2013, 109, 1–7. [Google Scholar] [CrossRef]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Withers, S.T.; Keasling, J.D. Biosynthesis and engineering of isoprenoid small molecules. Appl. Microbiol. Biotechnol. 2007, 73, 980–990. [Google Scholar] [CrossRef]
- Rabi, T.; Bishayee, A. Terpenoids and breast cancer chemoprevention. Breast Cancer Res. Treat. 2009, 115, 223–239. [Google Scholar] [CrossRef]
- Sheikh, I.; Sharma, V.; Tuli, H.S.; Aggarwal, D.; Sankhyan, A.; Vyas, P.; Sharma, A.K.; Bishayee, A. Cancer chemoprevention by flavonoids, dietary polyphenols and terpenoids. Biointerface Res. Appl. Chem. 2020, 11, 8502–8537. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, Z.; Chen, J.; Zheng, Y.; Limsila, B.; Lu, M.; Gao, T.; Yang, Q.; Fu, C.; Liao, W. Terpenoids from Curcumae Rhizoma: Their anticancer effects and clinical uses on combination and versus drug therapies. Biomed. Pharmacother. 2021, 138, 111350. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, Y.; Shen, F.; Long, D.; Yu, T.; Lin, X. Introduction of exogenous wild-type p53 mediates the regulation of oncoprotein 18/stathmin signaling via nuclear factor-κB in non-small cell lung cancer NCI-H1299 cells. Oncol. Rep. 2019, 41, 2051–2059. [Google Scholar] [CrossRef]
- Xue, C.; Yao, Q.; Gu, X.; Shi, Q.; Yuan, X.; Chu, Q.; Bao, Z.; Lu, J.; Li, L. Evolving cognition of the JAK-STAT signaling pathway: Autoimmune disorders and cancer. Signal. Transduct. Target. Ther. 2023, 8, 204. [Google Scholar] [CrossRef]
- Kwon, S. Molecular dissection of Janus kinases as drug targets for inflammatory diseases. Front. Immunol. 2022, 13, 1075192. [Google Scholar] [CrossRef]
- Turkson, J. STAT proteins as novel targets for cancer drug discovery. Expert. Opin. Ther. Targets 2004, 8, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sak, K.; Iqubal, A.; Garg, V.K.; Varol, M.; Sharma, U.; Chauhan, A.; Yerer, M.B.; Dhama, K.; Jain, M.; et al. STAT signaling as a target for intervention: From cancer inflammation and angiogenesis to non-coding RNAs modulation. Mol. Biol. Rep. 2022, 49, 8987–8999. [Google Scholar] [CrossRef]
- Rusek, M.; Smith, J.; El-Khatib, K.; Aikins, K.; Czuczwar, S.J.; Pluta, R. The Role of the JAK/STAT Signaling Pathway in the Pathogenesis of Alzheimer’s Disease: New Potential Treatment Target. Int. J. Mol. Sci. 2023, 24, 864. [Google Scholar] [CrossRef] [PubMed]
- Villarino, A.V.; Kanno, Y.; O’Shea, J.J. Mechanisms and consequences of Jak-STAT signaling in the immune system. Nat. Immunol. 2017, 18, 374–384. [Google Scholar] [CrossRef]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT Signaling as a Target for Inflammatory and Autoimmune Diseases: Current and Future Prospects. Drugs 2017, 77, 521–546, Erratum in Drugs 2017, 77, 939. [Google Scholar] [CrossRef]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef]
- Gao, Q.; Liang, X.; Shaikh, A.S.; Zang, J.; Xu, W.; Zhang, Y. JAK/STAT Signal Transduction: Promising Attractive Targets for Immune, Inflammatory and Hematopoietic Diseases. Curr. Drug Targets 2018, 19, 487–500. [Google Scholar] [CrossRef]
- El-Tanani, M.; Al Khatib, A.O.; Aladwan, S.M.; Abuelhana, A.; McCarron, P.A.; Tambuwala, M.M. Importance of STAT3 signalling in cancer, metastasis and therapeutic interventions. Cell. Signal. 2022, 92, 110275. [Google Scholar] [CrossRef]
- Takeda, J.; Yoshida, K.; Nakagawa, M.M.; Nannya, Y.; Yoda, A.; Morishita, D.; Saiki, R.; Chiba, K.; Tanaka, H.; Shiraishi, Y.; et al. EPOR/JAK/STAT signaling pathway as therapeutic target of acute erythroid leukemia. Blood 2021, 138, 610. [Google Scholar] [CrossRef]
- Arshad, S.; Naveed, M.; Ullia, M.; Javed, K.; Butt, A.; Khawar, M.; Amjad, F. Targeting STAT-3 signaling pathway in cancer for development of novel drugs: Advancements and challenges. Genet. Mol. Biol. 2020, 43, e20180160. [Google Scholar] [CrossRef] [PubMed]
- Aigner, P.; Just, V.; Stoiber, D. STAT3 isoforms: Alternative fates in cancer? Cytokine 2019, 118, 27–34. [Google Scholar] [CrossRef]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef]
- Liu, W.H.; Chen, M.T.; Wang, M.L.; Lee, Y.Y.; Chiou, G.Y.; Chien, C.S.; Huang, P.I.; Chen, Y.W.; Huang, M.C.; Chiou, S.H.; et al. Cisplatin-selected resistance is associated with increased motility and stem-like properties via activation of STAT3/Snail axis in atypical teratoid/rhabdoid tumor cells. Oncotarget 2015, 6, 1750–1768. [Google Scholar] [CrossRef]
- Lin, J.C.; Tsai, J.T.; Chao, T.Y.; Ma, H.I.; Liu, W.H. The STAT3/Slug Axis Enhances Radiation-Induced Tumor Invasion and Cancer Stem-like Properties in Radioresistant Glioblastoma. Cancers 2018, 10, 512. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.C.; Mei, K.C. Role of STAT3 signaling transduction pathways in cancer stem cell-associated chemoresistance. Drug Discov. Today 2021, 26, 1450–1458. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: A novel therapeutic strategy. Semin. Cancer Biol. 2022, 80, 1–17. [Google Scholar] [CrossRef]
- Huang, A.; Zhou, W. Mn-based cGAS-STING activation for tumor therapy. Chin. J. Cancer Res. 2023, 35, 19–43. [Google Scholar] [CrossRef]
- Yang, J.; Ren, B.; Yang, G.; Wang, H.; Chen, G.; You, L.; Zhang, T.; Zhao, Y. The enhancement of glycolysis regulates pancreatic cancer metastasis. Cell. Mol. Life Sci. 2020, 77, 305–321. [Google Scholar] [CrossRef]
- Park, J.H.; Pyun, W.Y.; Park, H.W. Cancer Metabolism: Phenotype, Signaling and Therapeutic Targets. Cells 2020, 9, 2308. [Google Scholar] [CrossRef]
- Wang, Z.; Hui, C.; Xie, Y. Natural STAT3 inhibitors: A mini perspective. Bioorg. Chem. 2021, 115, 105169. [Google Scholar] [CrossRef]
- Mohan, C.D.; Rangappa, S.; Preetham, H.D.; Chandra Nayaka, S.; Gupta, V.K.; Basappa, S.; Sethi, G.; Rangappa, K.S. Targeting STAT3 signaling pathway in cancer by agents derived from Mother Nature. Semin. Cancer Biol. 2022, 80, 157–182. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Sikka, S.; Surana, R.; Dai, X.; Zhang, J.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Targeting the STAT3 signaling pathway in cancer: Role of synthetic and natural inhibitors. Biochim. Biophys. Acta 2014, 1845, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.R.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 9, 1451. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, L.; Guan, X.; Qin, J.J. Inhibiting STAT3 signaling pathway by natural products for cancer prevention and therapy: In vitro and in vivo activity and mechanisms of action. Pharmacol. Res. 2022, 182, 106357. [Google Scholar] [CrossRef] [PubMed]
- El-Baba, C.; Baassiri, A.; Kiriako, G.; Dia, B.; Fadlallah, S.; Moodad, S.; Darwiche, N. Terpenoids’ anti-cancer effects: Focus on autophagy. Apoptosis 2021, 26, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.Q.; Wang, J.; Guo, X.F.; Liu, Z.; Dong, W.G. Thymoquinone inhibits proliferation in gastric cancer via the STAT3 pathway in vivo and in vitro. World J. Gastroenterol. 2016, 22, 4149–4159. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.; Choi, B.Y.; Jeong, C.H.; Kundu, J.K.; Chun, K.S. Thymoquinone induces apoptosis in human colon cancer HCT116 cells through inactivation of STAT3 by blocking JAK2- and Src mediated phosphorylation of EGF receptor tyrosine kinase. Oncol. Rep. 2014, 32, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Al-Rawashde, F.A.; Wan Taib, W.R.; Ismail, I.; Johan, M.F.; Al-Wajeeh, A.S.; Al-Jamal, H.A.N. Thymoquinone Induces Downregulation of BCR-ABL/JAK/STAT Pathway and Apoptosis in K562 Leukemia Cells. Asian Pac. J. Cancer Prev. 2021, 22, 3959–3965. [Google Scholar] [CrossRef]
- Almajali, B.; Johan, M.F.; Al-Wajeeh, A.S.; Wan Taib, W.R.; Ismail, I.; Alhawamdeh, M.; Al-Tawarah, N.M.; Ibrahim, W.N.; Al-Rawashde, F.A.; Al-Jamal, H.A.N. Gene Expression Profiling and Protein Analysis Reveal Suppression of the C-Myc Oncogene and Inhibition JAK/STAT and PI3K/AKT/mTOR Signaling by Thymoquinone in Acute Myeloid Leukemia Cells. Pharmaceuticals 2022, 15, 307. [Google Scholar] [CrossRef] [PubMed]
- Atteia, H.H.; Arafa, M.H.; Mohammad, N.S.; Amin, D.M.; Sakr, A.T. Thymoquinone upregulates miR-125a-5p, attenuates STAT3 activation, and potentiates doxorubicin antitumor activity in murine solid Ehrlich carcinoma. J. Biochem. Mol. Toxicol. 2021, 35, e22924. [Google Scholar] [CrossRef] [PubMed]
- Raut, P.K.; Lee, H.S.; Joo, S.H.; Chun, K.S. Thymoquinone induces oxidative stress-mediated apoptosis through downregulation of Jak2/STAT3 signaling pathway in human melanoma cells. Food Chem. Toxicol. 2021, 157, 112604. [Google Scholar] [CrossRef]
- Chae, I.G.; Song, N.Y.; Kim, D.H.; Lee, M.Y.; Park, J.M.; Chun, K.S. Thymoquinone induces apoptosis of human renal carcinoma Caki-1 cells by inhibiting JAK2/STAT3 through pro-oxidant effect. Food Chem. Toxicol. 2020, 139, 111253. [Google Scholar] [CrossRef]
- Park, J.E.; Kim, D.H.; Ha, E.; Choi, S.M.; Choi, J.S.; Chun, K.S.; Joo, S.H. Thymoquinone induces apoptosis of human epidermoid carcinoma A431 cells through ROS-mediated suppression of STAT3. Chem. Biol. Interact. 2019, 312, 108799. [Google Scholar] [CrossRef]
- Li, F.; Rajendran, P.; Sethi, G. Thymoquinone inhibits proliferation, induces apoptosis and chemosensitizes human multiple myeloma cells through suppression of signal transducer and activator of transcription 3 activation pathway. Br. J. Pharmacol. 2010, 161, 541–554. [Google Scholar] [CrossRef]
- Wang, X.R.; Jiang, Z.B.; Xu, C.; Meng, W.Y.; Liu, P.; Zhang, Y.Z.; Xie, C.; Xu, J.Y.; Xie, Y.J.; Liang, T.L.; et al. Andrographolide suppresses non-small-cell lung cancer progression through induction of autophagy and antitumor immune response. Pharmacol. Res. 2022, 179, 106198. [Google Scholar] [CrossRef]
- Zhou, J.; Ong, C.N.; Hur, G.M.; Shen, H.M. Inhibition of the JAK-STAT3 pathway by andrographolide enhances chemosensitivity of cancer cells to doxorubicin. Biochem. Pharmacol. 2010, 79, 1242–1250. [Google Scholar] [CrossRef]
- Shin, D.S.; Kim, H.N.; Shin, K.D.; Yoon, Y.J.; Kim, S.J.; Han, D.C.; Kwon, B.M. Cryptotanshinone inhibits constitutive signal transducer and activator of transcription 3 function through blocking the dimerization in DU145 prostate cancer cells. Cancer Res. 2009, 69, 193–202. [Google Scholar] [CrossRef]
- Yang, Y.; Cao, Y.; Chen, L.; Liu, F.; Qi, Z.; Cheng, X.; Wang, Z. Cryptotanshinone suppresses cell proliferation and glucose metabolism via STAT3/SIRT3 signaling pathway in ovarian cancer cells. Cancer Med. 2018, 7, 4610–4618. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Yang, B.; Chen, Z.; Cheng, R. Cryptotanshinone suppresses the proliferation and induces the apoptosis of pancreatic cancer cells via the STAT3 signaling pathway. Mol. Med. Rep. 2015, 12, 7782–7788. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Liu, Y.; Xue, N.; Du, T.; Wang, L.; Huang, R.; Li, L.; Yan, C.; Chen, X. Cryptotanshinone inhibits esophageal squamous-cell carcinoma in vitro and in vivo through the suppression of STAT3 activation. OncoTargets Ther. 2019, 12, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhu, R.; Zheng, J.; Chen, C.; Huang, C.; Ma, J.; Xu, C.; Zhai, W.; Zheng, J. Cryptotanshinone inhibits proliferation yet induces apoptosis by suppressing STAT3 signals in renal cell carcinoma. Oncotarget 2017, 8, 50023–50033. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, C.; Li, D.; Wang, Y.; Zhou, C.; Shao, W.; Peng, J.; You, Y.; Zhang, X.; Shen, X. Cryptotanshinone inhibits human glioma cell proliferation by suppressing STAT3 signaling. Mol. Cell. Biochem. 2013, 381, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhang, S.; Li, C.; Zhou, C.; Li, D.; Liu, P.; Huang, M.; Shen, X. Cryptotanshinone inhibits human glioma cell proliferation in vitro and in vivo through SHP-2-dependent inhibition of STAT3 activation. Cell Death Dis. 2017, 8, e2767. [Google Scholar] [CrossRef]
- Cheng, R.; Huang, Y.; Fang, Y.; Wang, Q.; Yan, M.; Ge, Y. Cryptotanshinone enhances the efficacy of Bcr-Abl tyrosine kinase inhibitors via inhibiting STAT3 and eIF4E signalling pathways in chronic myeloid leukaemia. Pharm. Biol. 2021, 59, 893–903. [Google Scholar] [CrossRef]
- Dong, B.; Liang, Z.; Chen, Z.; Li, B.; Zheng, L.; Yang, J.; Zhou, H.; Qu, L. Cryptotanshinone suppresses key onco-proliferative and drug-resistant pathways of chronic myeloid leukemia by targeting STAT5 and STAT3 phosphorylation. Sci. China Life Sci. 2018, 61, 999–1009. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X.; Wang, L.; Mei, Y.; Yun, Y.; Yao, X.; Chen, Q.; Zhou, J.; Kou, B. Oridonin represses epithelial-mesenchymal transition and angiogenesis of thyroid cancer via downregulating JAK2/STAT3 signaling. Int. J. Med. Sci. 2022, 19, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Huang, G.; Yang, Y.; Gao, R.; Zhang, S.; Kou, B. Oridonin inhibits epithelial-mesenchymal transition of human nasopharyngeal carcinoma cells by negatively regulating AKT/STAT3 signaling pathway. Int. J. Med. Sci. 2021, 18, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhang, J.; Yan, S.; Tao, Z.; Wang, C.; Huang, M.; Zhang, X. Oridonin inhibits the proliferation, migration and invasion of human osteosarcoma cells via suppression of matrix metalloproteinase expression and STAT3 signalling pathway. J. Buon 2019, 24, 1175–1180. [Google Scholar] [PubMed]
- Zhou, J.; Hou, J.; Wang, J.; Wang, J.; Gao, J.; Bai, Y. Brusatol inhibits laryngeal cancer cell proliferation and metastasis via abrogating JAK2/STAT3 signaling mediated epithelial-mesenchymal transition. Life Sci. 2021, 284, 119907. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Rangappa, S.; Mohan, C.D.; Basappa; Sethi, G.; Lin, Z.X.; Rangappa, K.S.; Ahn, K.S. Brusatol, a Nrf2 Inhibitor Targets STAT3 Signaling Cascade in Head and Neck Squamous Cell Carcinoma. Biomolecules 2019, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Ye, W.; Huang, C.; Lou, B.; Zhang, J.; Yu, D.; Huang, X.; Chen, B.; Zhou, M. Brusatol inhibits growth and induces apoptosis in pancreatic cancer cells via JNK/p38 MAPK/NF-κb/Stat3/Bcl-2 signaling pathway. Biochem. Biophys. Res. Commun. 2017, 487, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Mohan, C.D.; Deivasigamani, A.; Jung, Y.Y.; Rangappa, S.; Basappa, S.; Chinnathambi, A.; Alahmadi, T.A.; Alharbi, S.A.; Garg, M.; et al. Brusatol suppresses STAT3-driven metastasis by downregulating epithelial-mesenchymal transition in hepatocellular carcinoma. J. Adv. Res. 2020, 26, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Sung, B.; Aggarwal, B.B. Betulinic acid suppresses STAT3 activation pathway through induction of protein tyrosine phosphatase SHP-1 in human multiple myeloma cells. Int. J. Cancer 2010, 127, 282–292. [Google Scholar] [CrossRef]
- Shin, J.; Lee, H.J.; Jung, D.B.; Jung, J.H.; Lee, H.J.; Lee, E.O.; Lee, S.G.; Shim, B.S.; Choi, S.H.; Ko, S.G.; et al. Suppression of STAT3 and HIF-1 alpha mediates anti-angiogenic activity of betulinic acid in hypoxic PC-3 prostate cancer cells. PLoS ONE 2011, 6, e21492. [Google Scholar] [CrossRef]
- Rajendran, P.; Li, F.; Shanmugam, M.K.; Kannaiyan, R.; Goh, J.N.; Wong, K.F.; Wang, W.; Khin, E.; Tergaonkar, V.; Kumar, A.P.; et al. Celastrol suppresses growth and induces apoptosis of human hepatocellular carcinoma through the modulation of STAT3/JAK2 signaling cascade in vitro and in vivo. Cancer Prev. Res. 2012, 5, 631–643. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, Y.; Gong, Y.; Wang, X.; Zhang, L.; Zhao, H.; Li, J.; Zhu, J.; Huang, X.; Zhao, C.; et al. Celastrol elicits antitumor effects by inhibiting the STAT3 pathway through ROS accumulation in non-small cell lung cancer. J. Transl. Med. 2022, 20, 525. [Google Scholar] [CrossRef]
- Kannaiyan, R.; Hay, H.S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Vali, S.; Abbasi, T.; Kapoor, S.; Sharma, A.; Kumar, A.P.; et al. Celastrol inhibits proliferation and induces chemosensitization through down-regulation of NF-κB and STAT3 regulated gene products in multiple myeloma cells. Br. J. Pharmacol. 2011, 164, 1506–1521, Erratum in Br. J. Pharmacol. 2012, 165, 540. [Google Scholar] [CrossRef]
- Thoennissen, N.H.; Iwanski, G.B.; Doan, N.B.; Okamoto, R.; Lin, P.; Abbassi, S.; Song, J.H.; Yin, D.; Toh, M.; Xie, W.D.; et al. Cucurbitacin B induces apoptosis by inhibition of the JAK/STAT pathway and potentiates antiproliferative effects of gemcitabine on pancreatic cancer cells. Cancer Res. 2009, 69, 5876–5884. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, C.; Shan, X.; Yang, X.; Li-Ling, J.; Deng, Y. Inhibition of pancreatic cancer cell growth by cucurbitacin B through modulation of signal transducer and activator of transcription 3 signaling. Pancreas 2010, 39, 923–929. [Google Scholar] [CrossRef]
- Yar Saglam, A.S.; Alp, E.; Elmazoglu, Z.; Menevse, S. Treatment with cucurbitacin B alone and in combination with gefitinib induces cell cycle inhibition and apoptosis via EGFR and JAK/STAT pathway in human colorectal cancer cell lines. Hum. Exp. Toxicol. 2016, 35, 526–543. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, H.; Sun, C.; Shan, X.; Yang, X.; Li-Ling, J.; Deng, Y. Targeted constitutive activation of signal transducer and activator of transcription 3 in human hepatocellular carcinoma cells by cucurbitacin B. Cancer Chemother. Pharmacol. 2009, 63, 635–642. [Google Scholar] [CrossRef]
- Xie, Y.L.; Tao, W.H.; Yang, T.X.; Qiao, J.G. Anticancer effect of cucurbitacin B on MKN-45 cells via inhibition of the JAK2/STAT3 signaling pathway. Exp. Ther. Med. 2016, 12, 2709–2715. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, Y.; Yang, R.; Zhou, T.; Ke, W.; Si, Y.; Yang, S.; Zhang, T.; Liu, X.; Zhang, L.; et al. Cucurbitacin B inhibits gastric cancer progression by suppressing STAT3 activity. Arch. Biochem. Biophys. 2020, 684, 108314. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, M.; Shan, X.; Zhou, X.; Yang, J.; Wang, Y.; Li-Ling, J.; Deng, Y. Inhibitory effect of cucurbitacin E on pancreatic cancer cells growth via STAT3 signaling. J. Cancer Res. Clin. Oncol. 2010, 136, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Lu, B.; Zhang, X.; Zhang, J.; Lai, L.; Li, D.; Wu, Y.; Song, Y.; Luo, J.; Pang, X.; et al. Cucurbitacin E, a tetracyclic triterpenes compound from Chinese medicine, inhibits tumor angiogenesis through VEGFR2-mediated Jak2-STAT3 signaling pathway. Carcinogenesis 2010, 31, 2097–2104, Erratum in Carcinogenesis 2012, 33, 946. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.W.; Yang, J.S.; Lin, M.W.; Chen, P.Y.; Chiou, S.M.; Chueh, F.S.; Lan, Y.H.; Pai, S.J.; Tsuzuki, M.; Ho, W.J.; et al. Cucurbitacin E Induces G2/M Phase Arrest through STAT3/p53/p21 Signaling and Provokes Apoptosis via Fas/CD95 and Mitochondria-Dependent Pathways in Human Bladder Cancer T24 Cells. Evid.-Based Complement. Altern. Med. 2012, 2012, 952762. [Google Scholar] [CrossRef]
- Xu, D.; Shen, H.; Tian, M.; Chen, W.; Zhang, X. Cucurbitacin I inhibits the proliferation of pancreatic cancer through the JAK2/STAT3 signalling pathway in vivo and in vitro. J. Cancer 2022, 13, 2050–2060. [Google Scholar] [CrossRef] [PubMed]
- Üremiş, N.; Üremiş, M.M.; Çiğremiş, Y.; Tosun, E.; Baysar, A.; Türköz, Y. Cucurbitacin I exhibits anticancer efficacy through induction of apoptosis and modulation of JAK/STAT3, MAPK/ERK, and AKT/mTOR signaling pathways in HepG2 cell line. J. Food Biochem. 2022, 46, e14333. [Google Scholar] [CrossRef]
- Ni, Y.; Wu, S.; Wang, X.; Zhu, G.; Chen, X.; Ding, Y.; Jiang, W. Cucurbitacin I induces pro-death autophagy in A549 cells via the ERK-mTOR-STAT3 signaling pathway. J. Cell. Biochem. 2018, 119, 6104–6112. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Blaskovich, M.A.; Jove, R.; Livingston, S.K.; Coppola, D.; Sebti, S.M. Cucurbitacin Q: A selective STAT3 activation inhibitor with potent antitumor activity. Oncogene 2005, 24, 3236–3245, Erratum in Oncogene 2008, 27, 1344. [Google Scholar] [CrossRef]
- Su, Y.; Li, G.; Zhang, X.; Gu, J.; Zhang, C.; Tian, Z.; Zhang, J. JSI-124 inhibits glioblastoma multiforme cell proliferation through G(2)/M cell cycle arrest and apoptosis augment. Cancer Biol. Ther. 2008, 7, 1243–1249. [Google Scholar] [CrossRef]
- Shi, X.; Franko, B.; Frantz, C.; Amin, H.M.; Lai, R. JSI-124 (cucurbitacin I) inhibits Janus kinase-3/signal transducer and activator of transcription-3 signalling, downregulates nucleophosmin-anaplastic lymphoma kinase (ALK), and induces apoptosis in ALK-positive anaplastic large cell lymphoma cells. Br. J. Haematol. 2006, 135, 26–32. [Google Scholar] [CrossRef]
- Kang, D.Y.; Sp, N.; Lee, J.M.; Jang, K.J. Antitumor Effects of Ursolic Acid through Mediating the Inhibition of STAT3/PD-L1 Signaling in Non-Small Cell Lung Cancer Cells. Biomedicines 2021, 9, 297. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Shin, E.A.; Jung, J.H.; Park, J.E.; Kim, D.S.; Shim, B.S.; Kim, S.H. Ursolic Acid Induces Apoptosis in Colorectal Cancer Cells Partially via Upregulation of MicroRNA-4500 and Inhibition of JAK2/STAT3 Phosphorylation. Int. J. Mol. Sci. 2018, 20, 114. [Google Scholar] [CrossRef]
- Liu, T.; Ma, H.; Shi, W.; Duan, J.; Wang, Y.; Zhang, C.; Li, C.; Lin, J.; Li, S.; Lv, J.; et al. Inhibition of STAT3 signaling pathway by ursolic acid suppresses growth of hepatocellular carcinoma. Int. J. Oncol. 2017, 51, 555–562. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Ong, T.H.; Kumar, A.P.; Lun, C.K.; Ho, P.C.; Wong, P.T.; Hui, K.M.; Sethi, G. Ursolic acid inhibits the initiation, progression of prostate cancer and prolongs the survival of TRAMP mice by modulating pro-inflammatory pathways. PLoS ONE 2012, 7, e32476. [Google Scholar] [CrossRef] [PubMed]
- Babaei, G.; Khadem Ansari, M.H.; Aziz, S.G.; Bazl, M.R. Alantolactone inhibits stem-like cell phenotype, chemoresistance and metastasis in PC3 cells through STAT3 signaling pathway. Res. Pharm. Sci. 2020, 15, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Gamallat, Y.; Su, P.; Husain, A.; Rehman, A.U.; Zaky, M.Y.; Bakheet, A.M.H.; Tahir, N.; Xin, Y.; Liang, W. Alantolactone induces apoptosis in THP-1 cells through STAT3, survivin inhibition, and intrinsic apoptosis pathway. Chem. Biol. Drug Des. 2021, 97, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Li, R.J.; Cheng, M.S.; Kim, Y.S. Alantolactone selectively suppresses STAT3 activation and exhibits potent anticancer activity in MDA-MB-231 cells. Cancer Lett. 2015, 357, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yang, L.; Kang, Y.; Chen, M.; Lin, S.; Xiang, Y.; Li, C.; Dai, X.; Huang, X.; Liang, G.; et al. Alantolactone sensitizes human pancreatic cancer cells to EGFR inhibitors through the inhibition of STAT3 signaling. Mol. Carcinog. 2019, 58, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Cho, S.K.; Kapoor, S.; Kumar, A.; Vali, S.; Abbasi, T.; Kim, S.H.; Sethi, G.; Ahn, K.S. β-Caryophyllene oxide inhibits constitutive and inducible STAT3 signaling pathway through induction of the SHP-1 protein tyrosine phosphatase. Mol. Carcinog. 2014, 53, 793–806. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Wang, J.G.; Feng, J.Y.; Huang, G.D.; Luo, C.G. Dihydroartemisinin Affects STAT3/DDA1 Signaling Pathway and Reverses Breast Cancer Resistance to Cisplatin. Am. J. Chin. Med. 2023, 51, 445–459. [Google Scholar] [CrossRef]
- Yan, X.; Li, P.; Zhan, Y.; Qi, M.; Liu, J.; An, Z.; Yang, W.; Xiao, H.; Wu, H.; Qi, Y.; et al. Dihydroartemisinin suppresses STAT3 signaling and Mcl-1 and Survivin expression to potentiate ABT-263-induced apoptosis in Non-small Cell Lung Cancer cells harboring EGFR or RAS mutation. Biochem. Pharmacol. 2018, 150, 72–85. [Google Scholar] [CrossRef]
- Jia, L.; Song, Q.; Zhou, C.; Li, X.; Pi, L.; Ma, X.; Li, H.; Lu, X.; Shen, Y. Dihydroartemisinin as a Putative STAT3 Inhibitor, Suppresses the Growth of Head and Neck Squamous Cell Carcinoma by Targeting Jak2/STAT3 Signaling. PLoS ONE 2016, 11, e0147157. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhong, B.; Li, Y.; Liu, X. Dihydroartemisinin increases apoptosis of colon cancer cells through targeting Janus kinase 2/signal transducer and activator of transcription 3 signaling. Oncol. Lett. 2018, 15, 1949–1954. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lu, X.; Li, H.; Li, X. Dihydroartemisinin inhibits IL-6-induced epithelial-mesenchymal transition in laryngeal squamous cell carcinoma via the miR-130b-3p/STAT3/β-catenin signaling pathway. J. Int. Med. Res. 2021, 49, 3000605211009494. [Google Scholar] [CrossRef]
- Yu, R.; Jin, L.; Li, F.; Fujimoto, M.; Wei, Q.; Lin, Z.; Ren, X.; Jin, Q.; Li, H.; Meng, F.; et al. Dihydroartemisinin inhibits melanoma by regulating CTL/Treg anti-tumor immunity and STAT3-mediated apoptosis via IL-10 dependent manner. J. Dermatol. Sci. 2020, 99, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lu, H.; Lv, M.; Wang, Q.; Sun, Y. Parthenolide facilitates apoptosis and reverses drug-resistance of human gastric carcinoma cells by inhibiting the STAT3 signaling pathway. Oncol. Lett. 2018, 15, 3572–3579. [Google Scholar] [CrossRef]
- Liu, M.; Xiao, C.; Sun, M.; Tan, M.; Hu, L.; Yu, Q. Parthenolide Inhibits STAT3 Signaling by Covalently Targeting Janus Kinases. Molecules 2018, 23, 1478. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Li, F.; Manu, K.A.; Shanmugam, M.K.; Loo, S.Y.; Kumar, A.P.; Sethi, G. γ-Tocotrienol is a novel inhibitor of constitutive and inducible STAT3 signalling pathway in human hepatocellular carcinoma: Potential role as an antiproliferative, pro-apoptotic and chemosensitizing agent. Br. J. Pharmacol. 2011, 163, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Darakhshan, S.; Bidmeshki Pour, A.; Hosseinzadeh Colagar, A.; Sisakhtnezhad, S. Thymoquinone and its therapeutic potentials. Pharmacol. Res. 2015, 95–96, 138–158. [Google Scholar] [CrossRef] [PubMed]
- Fatima Shad, K.; Soubra, W.; Cordato, D.J. The role of thymoquinone, a major constituent of Nigella sativa, in the treatment of inflammatory and infectious diseases. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1445–1453. [Google Scholar] [CrossRef]
- Mir, P.A.; Mohi-Ud-Din, R.; Banday, N.; Maqbool, M.; Raza, S.N.; Farooq, S.; Afzal, S.; Mir, R.H. Anticancer Potential of Thymoquinone: A Novel Bioactive Natural Compound from Nigella sativa L. Anti-Cancer Agents Med. Chem. 2022, 22, 3401–3415. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Chen, S.R.; Chai, L.; Zhao, J.; Wang, Y.; Wang, Y. Overview of pharmacological activities of Andrographis paniculata and its major compound andrographolide. Crit. Rev. Food Sci. Nutr. 2019, 59 (Suppl. S1), S17–S29. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vannucci, L. Biological properties of andrographolide, an active ingredient of Andrographis Paniculata: A narrative review. Ann. Transl. Med. 2021, 9, 1186. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; Liu, X.; Gao, X. Pharmacological properties of tanshinones, the natural products from Salvia miltiorrhiza. Adv. Pharmacol. 2020, 87, 43–70. [Google Scholar] [CrossRef]
- Wu, Y.H.; Wu, Y.R.; Li, B.; Yan, Z.Y. Cryptotanshinone: A review of its pharmacology activities and molecular mechanisms. Fitoterapia 2020, 145, 104633. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ouyang, L.; Peng, H.; Zhang, W.Z. Oridonin: Targeting programmed cell death pathways as an anti-tumour agent. Cell Prolif. 2012, 45, 499–507. [Google Scholar] [CrossRef]
- Yao, Z.; Xie, F.; Li, M.; Liang, Z.; Xu, W.; Yang, J.; Liu, C.; Li, H.; Zhou, H.; Qu, L.H. Oridonin induces autophagy via inhibition of glucose metabolism in p53-mutated colorectal cancer cells. Cell Death Dis. 2017, 8, e2633. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, C.T.; Ma, W.; Xie, X.; Huang, Q. Oridonin: A Review of Its Pharmacology, Pharmacokinetics and Toxicity. Front. Pharmacol. 2021, 12, 645824. [Google Scholar] [CrossRef]
- Sobral, P.J.M.; Vicente, A.T.S.; Salvador, J.A.R. Recent advances in oridonin derivatives with anticancer activity. Front. Chem. 2023, 11, 1066280. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.A.; Md Hashim, N.F.; Ammar, A.; Muhamad Zakuan, N. An Insight into the Anti-Angiogenic and Anti-Metastatic Effects of Oridonin: Current Knowledge and Future Potential. Molecules 2021, 26, 775. [Google Scholar] [CrossRef]
- Hu, X.; Huang, S.; Ye, S.; Jiang, J. The Natural Product Oridonin as an Anticancer Agent: Current Achievements and Problems. Curr. Pharm. Biotechnol. 2023. ahead of print. [Google Scholar] [CrossRef]
- Cai, S.J.; Liu, Y.; Han, S.; Yang, C. Brusatol, an NRF2 inhibitor for future cancer therapeutic. Cell Biosci. 2019, 9, 45. [Google Scholar] [CrossRef]
- Hall, I.H.; Lee, K.H.; Eigebaly, S.A.; Imakura, Y.; Sumida, Y.; Wu, R.Y. Antitumor agents. XXXIV: Mechanism of action of bruceoside A and brusatol on nucleic acid metabolism of P-388 lymphocytic leukemia cells. J. Pharm. Sci. 1979, 68, 883–887. [Google Scholar] [CrossRef]
- Jiang, W.; Li, X.; Dong, S.; Zhou, W. Betulinic acid in the treatment of tumour diseases: Application and research progress. Biomed. Pharmacother. 2021, 142, 111990. [Google Scholar] [CrossRef] [PubMed]
- Aswathy, M.; Vijayan, A.; Daimary, U.D.; Girisa, S.; Radhakrishnan, K.V.; Kunnumakkara, A.B. Betulinic acid: A natural promising anticancer drug, current situation, and future perspectives. J. Biochem. Mol. Toxicol. 2022, 36, e23206. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Dai, S.; Zhao, X.; Zhang, Y.; Gong, L.; Fu, K.; Ma, C.; Peng, C.; Li, Y. Celastrol as an emerging anticancer agent: Current status, challenges and therapeutic strategies. Biomed. Pharmacother. 2023, 163, 114882. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Wang, C.; Zhao, X.; Ma, C.; Fu, K.; Liu, Y.; Peng, C.; Li, Y. Cucurbitacin B: A review of its pharmacology, toxicity, and pharmacokinetics. Pharmacol. Res. 2023, 187, 106587. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Kaul, S.C.; Wadhwa, R. Cucurbitacin B and cancer intervention: Chemistry, biology and mechanisms (Review). Int. J. Oncol. 2018, 52, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Zieniuk, B.; Pawełkowicz, M. Recent Advances in the Application of Cucurbitacins as Anticancer Agents. Metabolites 2023, 13, 1081. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, B.; Sharma, U.; Parashar, G.; Parashar, N.C.; Rani, I.; Ramniwas, S.; Kaur, S.; Haque, S.; Tuli, H.S. Apoptotic and antimetastatic effect of cucurbitacins in cancer: Recent trends and advancement. Naunyn Schmiedebergs Arch. Pharmacol. 2023, 396, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.E.M.; Boulos, J.C.; Elhaboub, G.; Rigano, D.; Saab, A.; Loizzo, M.R.; Hassan, L.E.A.; Sugimoto, Y.; Piacente, S.; Tundis, R.; et al. Cytotoxicity of cucurbitacin E from Citrullus colocynthis against multidrug-resistant cancer cells. Phytomedicine 2019, 62, 152945. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Melim, C.; Neves, B.G.; Sharifi-Rad, J.; Calina, D.; Mamurova, A.; Cabral, C. Cucurbitacins as potential anticancer agents: New insights on molecular mechanisms. J. Transl. Med. 2022, 20, 630. [Google Scholar] [CrossRef]
- Lin, X.; Farooqi, A.A. Cucurbitacin mediated regulation of deregulated oncogenic signaling cascades and non-coding RNAs in different cancers: Spotlight on JAK/STAT, Wnt/β-catenin, mTOR, TRAIL-mediated pathways. Semin. Cancer Biol. 2021, 73, 302–309. [Google Scholar] [CrossRef]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2021, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bian, L.; Duan, X.; Zhuang, X.; Sui, Y.; Yang, L. Alantolactone: A sesquiterpene lactone with diverse pharmacological effects. Chem. Biol. Drug Des. 2021, 98, 1131–1145. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Gao, K.; Peng, B.; Xu, Z.; Peng, J.; Li, J.; Chen, X.; Zeng, S.; Hu, K.; Yan, Y. Alantolactone: A Natural Plant Extract as a Potential Therapeutic Agent for Cancer. Front. Pharmacol. 2021, 12, 781033. [Google Scholar] [CrossRef]
- Babaei, G.; Gholizadeh-Ghaleh Aziz, S.; Rajabi Bazl, M.; Khadem Ansari, M.H. A comprehensive review of anticancer mechanisms of action of Alantolactone. Biomed. Pharmacother. 2021, 136, 111231. [Google Scholar] [CrossRef] [PubMed]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Liu, X.; Cao, J.; Huang, G.; Zhao, Q.; Shen, J. Biological Activities of Artemisinin Derivatives Beyond Malaria. Curr. Top. Med. Chem. 2019, 19, 205–222. [Google Scholar] [CrossRef]
- Slezakova, S.; Ruda-Kucerova, J. Anticancer Activity of Artemisinin and its Derivatives. Anticancer. Res. 2017, 37, 5995–6003. [Google Scholar] [CrossRef]
- Sztiller-Sikorska, M.; Czyz, M. Parthenolide as Cooperating Agent for Anti-Cancer Treatment of Various Malignancies. Pharmaceuticals 2020, 13, 194. [Google Scholar] [CrossRef]
- Pang, K.L.; Mai, C.W.; Chin, K.Y. Molecular Mechanism of Tocotrienol-Mediated Anticancer Properties: A Systematic Review of the Involvement of Endoplasmic Reticulum Stress and Unfolded Protein Response. Nutrients 2023, 15, 1854. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Class | Phytochemical | Cancer | Model | Molecular Target | Mechanism | References |
|---|---|---|---|---|---|---|
| Monoterpene | Thymoquinone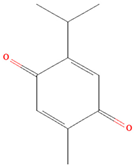 | Gastric cancer | HGC27, BGC823, SGC7901 cells; xenograft tumor mouse model | JAK2, STAT3, Src | Reduced tumor growth, induced apoptosis | [61] |
| Colon cancer | HCT116 cells | JAK2, STAT3, Src | Inhibited cell proliferation, induced apoptosis | [62] | ||
| Leukemia | K562 cells | JAK2, STAT3, STAT5 | Inhibited cell proliferation, induced apoptosis | [63] | ||
| Acute myeloid leukemia | HL60 cells | c-Myc, PI3K, AKT, mTOR, JAK2, STAT3, STAT5a, STAT5b | Inhibited cell proliferation, induced apoptosis | [64] | ||
| Breast cancer | Mice bearing solid Ehrlich tumors | STAT3, caspase-3/9 | Attenuated tumor growth, induced apoptosis, chemosensitivity | [65] | ||
| Melanoma | SK-MEL-28 cells, SK-MEL-28 tumor xenografts | Jak2, STAT3 | Induced apoptotic cell death | [66] | ||
| Renal cell carcinoma | Caki-1 cells, tumor xenograft mice | Jak2, STAT3, cyclin D2 | Attenuated tumor growth, induced apoptosis | [67] | ||
| Skin cancer | A431 cells, tumor xenograft mice | STAT3, Src, cyclin D1 | Attenuated tumor growth, induced apoptosis | [68] | ||
| Myeloma | U266 and RPMI 8226 cells | STAT3, c-Src, JAK2 | Attenuated cell growth, induced apoptosis, G1 cell cycle arrest, chemosensitivity | [69] | ||
| Diterpene | Andrographolide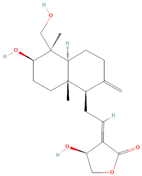 | Non-small-cell lung cancer (NSCLC) | H1975 and H1299 cell lines | STAT3, PARP, PD-L1, P62 | Inducted autophagy, antitumor immune response | [70] |
| Multiple cancer cell types | HCT116, MDA-MB-231, HepG2, HeLa, TPC-1 | STAT3, Bcl-xl, cyclin D1 | Induced cell death, chemosensitivity | [71] | ||
Cryptotanshinone | Prostate cancer | DU145 cells | JAK2, STAT3 | Retarded cell proliferation, induced apoptosis | [72] | |
| Ovarian cancer | Hey and A2780 cells | HIF- 1α, STAT3 | Retarded cell proliferation and glucose metabolism | [73] | ||
| Pancreatic cancer | BxPC-3 cells | JAK2, STAT3, mTOR, Akt | Retarded cell proliferation, induced apoptosis | [74] | ||
| Esophageal cancer | EC109 and CAES17 cells, athymic nude mice | JAK2, STAT3 | Retarded cell proliferation, induced apoptosis | [75] | ||
| Renal cell carcinoma | A498, 786-O, ACHN | STAT3, cyclin D1, Bcl-2 | Retarded cell proliferation, induced apoptosis | [76] | ||
| Glioma | T98G and U87 cells | STAT3, cyclin D1, survivin | Suppressed cell viability, induced cell cycle arrest and apoptosis | [77] | ||
| Glioma | C6, U251, T98G, U87; nude xenograft mice | STAT3, SHP2, cyclin D1 | Inhibited cell proliferation | [78] | ||
| Chronic myeloid leukemia | K562-R cells, xenografts in nude mice | STAT3, eIF4E | Inhibited tumor growth, induced apoptosis | [79] | ||
| Chronic myeloid leukemia | K562 cells | STAT3, STAT5 | Suppressed key oncogenic proliferation, drug resistance | [80] | ||
Oridonin | Thyroid cancer | TPC-1 and BCPAP cells | JAK2, STAT3 | Inhibited metastatic, angiogenesis, and modulated EMT | [81] | |
| Nasopharyngeal carcinoma | CNE-2Z and HNE-1 cells | Akt, STAT3 | Inhibited metastatic, angiogenesis, and modulated EMT | [82] | ||
| Osteosarcoma | U2OS cells | STAT3, MMP-2, 3, 9 | Suppressed cell viability, induced apoptosis | [83] | ||
| Triterpene | Brusatol | Laryngeal cancer | Hep-2 cells, xenograft laryngeal tumor | JAK2, STAT3 | Inhibited viability, migration, and invasion ability | [84] |
| Head and neck squamous cell carcinoma | UMSCC 47, UD SCC2, JMAR, Tu167, LN686, FaDu | JAK1, JAK2, STAT3, Src | Reduced cell growth, induced apoptosis | [85] | ||
| Pancreatic cancer | PANC-1 and PATU-8988 cells | NF-κb, Stat3 | Reduced cell growth, induced apoptosis | [86] | ||
| Hepatocellular carcinoma | HCCLM3 cell line | STAT3 | Inhibited cell migration and invasion | [87] | ||
Betulinic acid | Multiple myeloma | U266 and MM.1S cells | JAK1, JAK2, STAT3, Src | Induced apoptotic cancer cell death | [88] | |
| Prostate cancer | PC3 | HIF-1α, STAT3 | Exert anti-angiogenic activity | [89] | ||
Celastrol  | Hepatocellular carcinoma | C3A, HepG2, Hep3B, PLC/PRF5; athymic nu/nu mice | JAK1, JAK2, STAT3, Src | Inhibited cell migration and invasion, induced apoptosis | [90] | |
| NSCLC | H460, PC-9, H520, BEAS-2B, PC-9 cells; thymic BALB/c nude mice | STAT3, Bcl-2 | Reduced cell growth, proliferation, and metastasis | [91] | ||
| Multiple myeloma | U266, RPMI 8226 | IκBα kinase, STAT3 | Suppressed cell viability, induced cell cycle arrest and apoptosis | [92] | ||
Cucurbitacin B | Pancreatic cancer | MiaPaCa-2, AsPC-1 | JAK2, STAT3, and STAT5 activation | Cytotoxicity | [93] | |
| Pancreatic cancer | PANC-1 | STAT3 | Growth and cell cycle inhibition, induced apoptosis | [94] | ||
| Colorectal cancer | HT-29, HCT-116 | STAT3 | Growth and cell cycle inhibition, induced apoptosis | [95] | ||
| Hepatocellular carcinoma | HepG2 | STAT3 | Cell cycle arrest, retarded cell growth | [96] | ||
| Gastric cancer | MKN-45 | STAT3 | Cell cycle arrest, retarded cell growth | [97] | ||
| Gastric cancer | SGC7901, BGC823, MGC803, MKN74; human gastric cancer xenograft | STAT3, c-Myc, Bcl-xL | Suppressed invasion, induced apoptosis | [98] | ||
Cucurbitacin E  | Pancreatic cancer | PANC-1 | STAT3 | Cell cycle arrest, retarded cell growth | [99] | |
| Prostate cancer | PC-3, xenograft | STAT3, JAK2 | Inhibited angiogenesis, proliferation, survival, and migration | [100] | ||
| Human bladder cancer | T24 cells | STAT3, CDK1, cyclin B | Induced G(2)/M phase arrest and apoptosis | [101] | ||
Cucurbitacin I  | Pancreatic cancer | ASPC-1, BXPC-3, CFPAC-1, SW 1990; orthotopic xenograft mice | STAT3, JAK2 | Inhibited proliferation | [102] | |
| Hepatocellular carcinoma | HepG2 cells | STAT3, JAK2 | Induced antiproliferation and G2/M phase of cell cycle | [103] | ||
| Lung cancer | A549 cells | ERK, mTOR, STAT3 | Decreased cell viability, inhibited colony formation, induced apoptosis | [104] | ||
| Lung cancer | A549; nude mouse tumor xenograft model | STAT3 | Suppressed tumor growth, induced apoptosis | [105] | ||
| Glioblastoma | U251 and A172 cells | STAT3, cyclin B1, cdc2 | Decreased cell viability, induced G2/M cell cycle arrest, induced apoptosis | [106] | ||
| Lymphoma | ALK+ ALCL cell lines | STAT3, JAK3, NPM-ALK | Decreased cell viability, induced G2/M cell cycle arrest, induced apoptosis | [107] | ||
Ursolic Acid | Lung cancer | A549 and H460 cells | STAT3, MMP2, PD-L1, VEGF | Attenuated cell growth, invasion, migration, angiogenesis induced apoptosis, G1 cell cycle arrest, chemosensitivity | [108] | |
| Colorectal cancer | HCT116 and HT29 cells | STAT3 | Induced apoptotic cell death | [109] | ||
| Hepatocellular carcinoma | Hep3B, HEPG2, SSMC-7721, and Huh7 cells; mouse xenograft tumor model | STAT3 | Suppressed cell viability, cell migration, and colony formation | [110] | ||
| Prostate cancer | TRAMP mice | NF-κB, STAT3, AKT, IKKα/β | Reduced tumor growth | [111] | ||
| Sesquiterpene | Alantolactone | Prostate cancer | PC3 | SOX2, Oct-4, Nanog, CD133, CD44, STAT3 | Antimetastatic potential | [112] |
| Leukemia | THP-1 cells | STAT3, survivin | Decreased cell viability, increased cell death and apoptosis | [113] | ||
| Breast cancer | MDA-MB-231 cells | STAT3, MAPKs, NF-κB | Inhibition of migration, invasion, and adhesion | [114] | ||
| Pancreatic cancer | BxPC-3, AsPC1, and PANC-1 cells | STAT3, Bcl-2 | Reduced cell growth, induced cell death | [115] | ||
β-Caryophyllene oxide  | Human multiple myeloma (MM) cell lines, human prostate carcinoma, human breast carcinoma | U266, MM1.S, DU145, MDAMB-231 | JAK1, JAK2, STAT3, Src | Reduced cell proliferation and invasion, induced apoptosis | [116] | |
Dihydroartemisinin  | Breast cancer | MDA-MB-231 cells | STAT3, DDA1 | Repressed cell proliferation, induced apoptosis | [117] | |
| Lung cancer | H1975, HCC827, H1650, H3255, A549, H727 | STAT3, Mcl-1, survivin | Bax-dependent apoptosis | [118] | ||
| Head and neck Carcinoma | Fadu, Cal-27, and Hep-2 cells; xenograft model | Jak2, STAT3 | Apoptosis induction, attenuation of cell migration | [119] | ||
| Colon cancer | HCT116 | Jak2, STAT3 | Suppressed cell viability, induced apoptosis | [120] | ||
| Laryngeal squamous cell carcinoma | AMC-HN-8 and Tu212 cells | IL-6/STAT3, β-catenin | Repressed EMT and invasion | [121] | ||
| Melanoma | B16F10 cells; BALB/c xenograft mice | STAT3, IL-10, IL-6 | Induced apoptotic cell death | [122] | ||
Parthenolide  | Gastric cancer | SGC-7901/DDP cell | JAK2, STAT3, cyclin D1 | Decreased cell viability, induced G1 cell cycle arrest, induced apoptosis | [123] | |
| Multiple cancer subtypes | HepG2, MDA-MB-231, MDA-MB-468, HCT116, HT-29, Lovo, NCI-H1299, Colo205, BGC, H460, and Du145 cells | JAK1, STAT3, | Suppressed cellular growth and migration | [124] | ||
γ-Tocotrienol  | Hepatocellular carcinoma | HepG2, C3A, SNU-387, and PLC/PRF5 cells | STAT3, cyclin D1, Bcl-2, Bcl-xL, survivin, Mcl-1, VEGF | Inhibited proliferation, induced apoptosis, chemosensitivity | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, F.; Pandey, P.; Verma, M.; Upadhyay, T.K. Terpenoid-Mediated Targeting of STAT3 Signaling in Cancer: An Overview of Preclinical Studies. Biomolecules 2024, 14, 200. https://doi.org/10.3390/biom14020200
Khan F, Pandey P, Verma M, Upadhyay TK. Terpenoid-Mediated Targeting of STAT3 Signaling in Cancer: An Overview of Preclinical Studies. Biomolecules. 2024; 14(2):200. https://doi.org/10.3390/biom14020200
Chicago/Turabian StyleKhan, Fahad, Pratibha Pandey, Meenakshi Verma, and Tarun Kumar Upadhyay. 2024. "Terpenoid-Mediated Targeting of STAT3 Signaling in Cancer: An Overview of Preclinical Studies" Biomolecules 14, no. 2: 200. https://doi.org/10.3390/biom14020200
APA StyleKhan, F., Pandey, P., Verma, M., & Upadhyay, T. K. (2024). Terpenoid-Mediated Targeting of STAT3 Signaling in Cancer: An Overview of Preclinical Studies. Biomolecules, 14(2), 200. https://doi.org/10.3390/biom14020200