Impacts of Green Tea on Joint and Skeletal Muscle Health: Prospects of Translational Nutrition


 ,
,  , and
, and 
Abstract
1. Introduction
1.1. Prevalence of Osteoarthritis and Sarcopenia
1.2. Etiology of Osteoarthritis and Sarcopenia
1.2.1. Osteoarthritis
1.2.2. Sarcopenia
1.3. Green Tea
2. Beneficial Effects of Green Tea Extract (GTE) on Joint Health
2.1. Cell and Tissue Explant Studies
2.2. Animal Studies
2.3. Human Studies
3. Beneficial Effects of GTE on Skeletal Muscle Health
3.1. Effect of GTE on Oxidative Damage of Skeletal Muscle Cell Culture Studies
3.1.1. In Vitro Studies
3.1.2. Rodent Studies
3.1.3. Human Studies
3.2. Effect of GTE on Inflammation of Skeletal Muscle Animal studies
3.3. Effect of GTE on Autophagy in Skeletal Muscle
3.4. Effect of GTE on Mitochondria-Related Metabolism in Skeletal Muscle
3.5. Effect of GTE on Satellite Cells, Muscle Damage, and Recovery
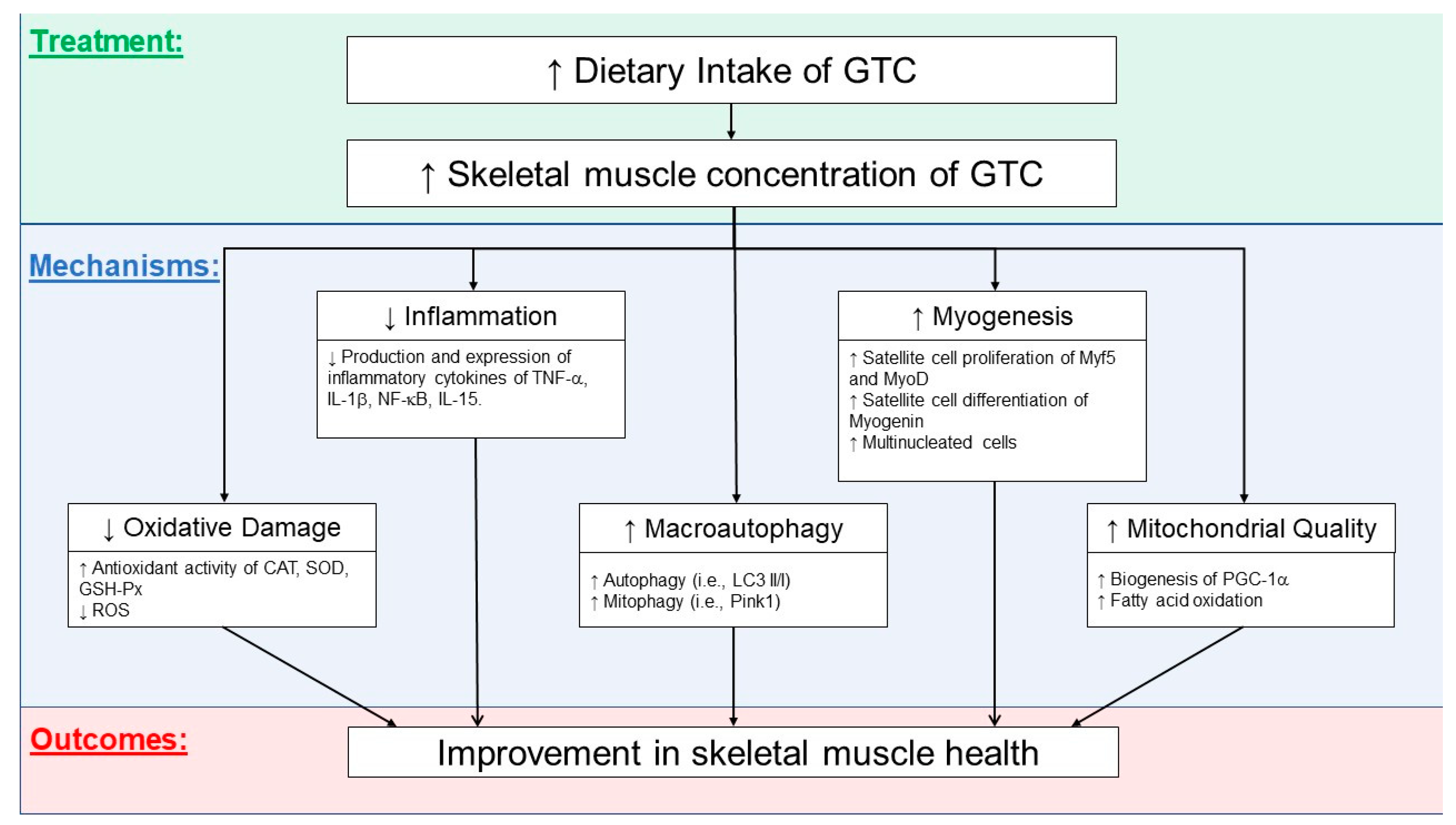
3.6. Possible Mechanisms Related to GTCs’ Effects on Skeletal Health
4. Potential Side Effects of Green Tea
5. Conclusions, Limitations, and Future Directions of Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, L.; Schwartz, T.A.; Helmick, C.G.; Renner, J.B.; Tudor, G.; Koch, G.; Dragomir, A.; Kalsbeek, W.D.; Luta, G.; Jordan, J.M. Lifetime Risk of Symptomatic Knee Osteoarthritis. Arthritis Care Res. Off. J. Am. Coll. Rheumatol. 2008, 59, 1207–1213. [Google Scholar] [CrossRef]
- Murphy, S.L.; Niemiec, S.S.; Lyden, A.K.; Kratz, A.L. Pain, Fatigue, and Physical Activity in Osteoarthritis: The Moderating Effects of Pain-and Fatigue-Related Activity Interference. Arch. Phys. Med. Rehabil. 2016, 97, S201–S209. [Google Scholar] [CrossRef]
- Bhatia, D.; Bejarano, T.; Novo, M. Current Interventions in the Management of Knee Osteoarthritis. J. Pharm. Bioallied Sci. 2013, 5, 30. [Google Scholar] [CrossRef]
- Grumati, P.; Coletto, L.; Schiavinato, A.; Castagnaro, S.; Bertaggia, E.; Sandri, M.; Bonaldo, P. Physical Exercise Stimulates Autophagy in Normal Skeletal Muscles but Is Detrimental for Collagen VI-Deficient Muscles. Autophagy 2011, 7, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Fang, F.; Li, F.; Ren, Y.; Hu, J.; Cao, J. Sarcopenia Is Associated with Hypertension in Older Adults: A Systematic Review and Meta-Analysis. BMC Geriatr. 2020, 20, 279. [Google Scholar] [CrossRef] [PubMed]
- Arango-Lopera, V.; Arroyo, P.; Gutiérrez-Robledo, L.M.; Perez-Zepeda, M.; Cesari, M. Mortality as an Adverse Outcome of Sarcopenia. J. Nutr. Health Aging 2013, 17, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Lexell, J.; Taylor, C.C.; Sjöström, M. What Is the Cause of the Ageing Atrophy? Total Number, Size and Proportion of Different Fiber Types Studied in Whole Vastus Lateralis Muscle from 15- to 83-Year-Old Men. J. Neurol. Sci. 1988, 84, 275–294. [Google Scholar] [CrossRef]
- Von Haehling, S.; Morley, J.E.; Anker, S.D. An Overview of Sarcopenia: Facts and Numbers on Prevalence and Clinical Impact. J. Cachexia Sarcopenia Muscle 2010, 1, 129–133. [Google Scholar] [CrossRef]
- Miljkovic, N.; Lim, J.-Y.; Miljkovic, I.; Frontera, W.R. Aging of Skeletal Muscle Fibers. Ann. Rehabil. Med. 2015, 39, 155. [Google Scholar] [CrossRef]
- Wright, E.A.; Katz, J.N.; Abrams, S.; Solomon, D.H.; Losina, E. Trends in Prescription of Opioids from 2003–2009 in Persons with Knee Osteoarthritis. Arthritis Care Res. 2014, 66, 1489–1495. [Google Scholar] [CrossRef]
- Pickering, M.; Chapurlat, R. Where Two Common Conditions of Aging Meet: Osteoarthritis and Sarcopenia. Calcif. Tissue Int. 2020, 107, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Krasnokutsky, S.; Attur, M.; Palmer, G.; Samuels, J.; Abramson, S. Current Concepts in the Pathogenesis of Osteoarthritis. Osteoarthr. Cartil. 2008, 16, S1–S3. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-Grade Inflammation as a Key Mediator of the Pathogenesis of Osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Sofat, N.; Ejindu, V.; Kiely, P. What Makes Osteoarthritis Painful? The Evidence for Local and Central Pain Processing. Rheumatology 2011, 50, 2157–2165. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M. Inflammation in Osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471. [Google Scholar] [CrossRef]
- Roos, E.M.; Arden, N.K. Strategies for the Prevention of Knee Osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 92. [Google Scholar] [CrossRef]
- Alghasham, A.; Rasheed, Z. Therapeutic Targets for Rheumatoid Arthritis: Progress and Promises. Autoimmunity 2014, 47, 77–94. [Google Scholar] [CrossRef]
- Takiguchi, R.; Komatsu, R.; Kitamura, K.; Watanabe, Y.; Takahashi, A.; Kobayashi, R.; Oshiki, R.; Saito, T.; Kabasawa, K.; Takachi, R. Modifiable Factors Associated with Symptomatic Knee Osteoarthritis: The Murakami Cohort Study. Maturitas 2019, 128, 53–59. [Google Scholar] [CrossRef]
- Sokolove, J.; Lepus, C.M. Role of Inflammation in the Pathogenesis of Osteoarthritis: Latest Findings and Interpretations. Ther. Adv. Musculoskelet. Dis. 2013, 5, 77–94. [Google Scholar] [CrossRef]
- Blanco, F.J.; Rego, I.; Ruiz-Romero, C. The Role of Mitochondria in Osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 161. [Google Scholar] [CrossRef]
- Luo, P.; Gao, F.; Niu, D.; Sun, X.; Song, Q.; Guo, C.; Liang, Y.; Sun, W. The Role of Autophagy in Chondrocyte Metabolism and Osteoarthritis: A Comprehensive Research Review. BioMed Res. Int. 2019, 2019, 5171602. [Google Scholar] [CrossRef] [PubMed]
- Caramés, B.; Taniguchi, N.; Otsuki, S.; Blanco, F.J.; Lotz, M. Autophagy Is a Protective Mechanism in Normal Cartilage, and Its Aging-related Loss Is Linked with Cell Death and Osteoarthritis. Arthritis Rheum. 2010, 62, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Caramés, B.; Olmer, M.; Kiosses, W.B.; Lotz, M.K. The Relationship of Autophagy Defects to Cartilage Damage during Joint Aging in a Mouse Model. Arthritis Rheumatol. 2015, 67, 1568–1576. [Google Scholar] [CrossRef]
- Barranco, C. Activate Autophagy to Prevent Cartilage Degeneration? Nat. Rev. Rheumatol. 2015, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vasheghani, F.; Li, Y.; Blati, M.; Simeone, K.; Fahmi, H.; Lussier, B.; Roughley, P.; Lagares, D.; Pelletier, J.-P. Cartilage-Specific Deletion of MTOR Upregulates Autophagy and Protects Mice from Osteoarthritis. Ann. Rheum. Dis. 2015, 74, 1432–1440. [Google Scholar] [CrossRef]
- Caramés, B.; Hasegawa, A.; Taniguchi, N.; Miyaki, S.; Blanco, F.J.; Lotz, M. Autophagy Activation by Rapamycin Reduces Severity of Experimental Osteoarthritis. Ann. Rheum. Dis. 2012, 71, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F.; Collins, J.A.; Diekman, B.O. Ageing and the Pathogenesis of Osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 412–420. [Google Scholar] [CrossRef]
- Masiero, E.; Agatea, L.; Mammucari, C.; Blaauw, B.; Loro, E.; Komatsu, M.; Metzger, D.; Reggiani, C.; Schiaffino, S.; Sandri, M. Autophagy Is Required to Maintain Muscle Mass. Cell Metab. 2009, 10, 507–515. [Google Scholar] [CrossRef]
- Ali, S.; Garcia, J.M. Sarcopenia, Cachexia and Aging: Diagnosis, Mechanisms and Therapeutic Options-a Mini-Review. Gerontology 2014, 60, 294–305. [Google Scholar] [CrossRef]
- Mankhong, S.; Kim, S.; Moon, S.; Kwak, H.-B.; Park, D.-H.; Kang, J.-H. Experimental Models of Sarcopenia: Bridging Molecular Mechanism and Therapeutic Strategy. Cells 2020, 9, 1385. [Google Scholar] [CrossRef]
- Gianni, P.; Jan, K.J.; Douglas, M.J.; Stuart, P.M.; Tarnopolsky, M.A. Oxidative Stress and the Mitochondrial Theory of Aging in Human Skeletal Muscle. Exp. Gerontol. 2004, 39, 1391–1400. [Google Scholar] [CrossRef]
- Yen, W.-L.; Klionsky, D.J. How to Live Long and Prosper: Autophagy, Mitochondria, and Aging. Physiology 2008, 23, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Marzani, B.; Giardina, S.; Negro, M.; Marzatico, F. Human Skeletal Muscle Aging and the Oxidative System: Cellular Events. Curr. Aging Sci. 2008, 1, 182–191. [Google Scholar] [CrossRef]
- Ranneh, Y.; Ali, F.; Akim, A.M.; Hamid, H.A.; Khazaai, H.; Fadel, A. Crosstalk between Reactive Oxygen Species and Pro-Inflammatory Markers in Developing Various Chronic Diseases: A Review. Appl. Biol. Chem. 2017, 60, 327–338. [Google Scholar] [CrossRef]
- Schakman, O.; Dehoux, M.; Bouchuari, S.; Delaere, S.; Lause, P.; Decroly, N.; Shoelson, S.E.; Thissen, J.-P. Role of IGF-I and the TNFα/NF-ΚB Pathway in the Induction of Muscle Atrogenes by Acute Inflammation. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E729–E739. [Google Scholar] [CrossRef]
- Johnson, M.L.; Robinson, M.M.; Nair, K.S. Skeletal Muscle Aging and the Mitochondrion. Trends Endocrinol. Metab. 2013, 24, 247–256. [Google Scholar] [CrossRef]
- Kim, Y.A.; Kim, Y.S.; Oh, S.L.; Kim, H.-J.; Song, W. Autophagic Response to Exercise Training in Skeletal Muscle with Age. J. Physiol. Biochem. 2013, 69, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Burd, N.A.; Gorissen, S.H.; Van Loon, L.J. Anabolic Resistance of Muscle Protein Synthesis with Aging. Exerc. Sport Sci. Rev. 2013, 41, 169–173. [Google Scholar] [CrossRef]
- Fan, J.; Kou, X.; Jia, S.; Yang, X.; Yang, Y.; Chen, N. Autophagy as a Potential Target for Sarcopenia. J. Cell. Physiol. 2016, 231, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Petrovski, G.; Das, D.K. Does Autophagy Take a Front Seat in Lifespan Extension? J. Cell. Mol. Med. 2010, 14, 2543–2551. [Google Scholar] [CrossRef]
- Jiao, J.; Demontis, F. Skeletal Muscle Autophagy and Its Role in Sarcopenia and Organismal Aging. Curr. Opin. Pharmacol. 2017, 34, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Shravage, B.V.; Baehrecke, E.H. Regulation and Function of Autophagy during Cell Survival and Cell Death. Cold Spring Harb. Perspect. Biol. 2012, 4, a008813. [Google Scholar] [CrossRef] [PubMed]
- Paré, M.; Baechler, B.; Fajardo, V.; Earl, E.; Wong, E.; Campbell, T.; Tupling, A.; Quadrilatero, J. Effect of Acute and Chronic Autophagy Deficiency on Skeletal Muscle Apoptotic Signaling, Morphology, and Function. Biochim. Biophys. Acta BBA Mol. Cell Res. 2017, 1864, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Carter, H.N.; Kim, Y.; Erlich, A.T.; Zarrin-khat, D.; Hood, D.A. Autophagy and Mitophagy Flux in Young and Aged Skeletal Muscle Following Chronic Contractile Activity. J. Physiol. 2018, 596, 3567–3584. [Google Scholar] [CrossRef]
- Takahashi, H.; Suzuki, Y.; Mohamed, J.S.; Gotoh, T.; Pereira, S.L.; Alway, S.E. Epigallocatechin-3-Gallate Increases Autophagy Signaling in Resting and Unloaded Plantaris Muscles but Selectively Suppresses Autophagy Protein Abundance in Reloaded Muscles of Aged Rats. Exp. Gerontol. 2017, 92, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K. Organellophagy: Eliminating Cellular Building Blocks via Selective Autophagy. J. Cell Biol. 2014, 205, 435–445. [Google Scholar] [CrossRef]
- Joseph, A.-M.; Adhihetty, P.J.; Wawrzyniak, N.R.; Wohlgemuth, S.E.; Picca, A.; Kujoth, G.C.; Prolla, T.A.; Leeuwenburgh, C. Dysregulation of Mitochondrial Quality Control Processes Contribute to Sarcopenia in a Mouse Model of Premature Aging. PLoS ONE 2013, 8, e69327. [Google Scholar] [CrossRef]
- Leduc-Gaudet, J.-P.; Picard, M.; Pelletier, F.S.-J.; Sgarioto, N.; Auger, M.-J.; Vallée, J.; Robitaille, R.; St-Pierre, D.H.; Gouspillou, G. Mitochondrial Morphology Is Altered in Atrophied Skeletal Muscle of Aged Mice. Oncotarget 2015, 6, 17923. [Google Scholar] [CrossRef]
- Alway, S.E.; Mohamed, J.S.; Myers, M.J. Mitochondria Initiate and Regulate Sarcopenia. Exerc. Sport Sci. Rev. 2017, 45, 58. [Google Scholar] [CrossRef]
- He, C.; Klionsky, D.J. Regulation Mechanisms and Signaling Pathways of Autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef]
- García-Prat, L.; Martínez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodríguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L. Autophagy Maintains Stemness by Preventing Senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, J.; Fitzpatrick, J.; Perera, N.K.P.; Orchard, J. Osteoarthritis-a Systematic Review of Long-Term Safety Implications for Osteoarthritis of the Knee. BMC Musculoskelet. Disord. 2019, 20, 151. [Google Scholar] [CrossRef]
- Fortun, P.J.; Hawkey, C.J. Nonsteroidal Antiinflammatory Drugs and the Small Intestine. Curr. Opin. Gastroenterol. 2005, 21, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Landi, F.; Schneider, S.M.; Zúñiga, C.; Arai, H.; Boirie, Y.; Chen, L.-K.; Fielding, R.A.; Martin, F.C.; Michel, J.-P. Prevalence of and Interventions for Sarcopenia in Ageing Adults: A Systematic Review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing 2014, 43, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Burton, L.A.; Sumukadas, D. Optimal Management of Sarcopenia. Clin. Interv. Aging 2010, 5, 217. [Google Scholar]
- Lapane, K.L.; Yang, S.; Jawahar, R.; McAlindon, T.; Eaton, C.B. CAM Use among Overweight and Obese Persons with Radiographic Knee Osteoarthritis. BMC Complement. Altern. Med. 2013, 13, 241. [Google Scholar] [CrossRef] [PubMed]
- Heck, C.I.; De Mejia, E.G. Yerba Mate Tea (Ilex Paraguariensis): A Comprehensive Review on Chemistry, Health Implications, and Technological Considerations. J. Food Sci. 2007, 72, R138–R151. [Google Scholar] [CrossRef]
- Adcocks, C.; Collin, P.; Buttle, D.J. Catechins from Green Tea (Camellia Sinensis) Inhibit Bovine and Human Cartilage Proteoglycan and Type II Collagen Degradation in vitro. J. Nutr. 2002, 132, 341–346. [Google Scholar] [CrossRef]
- Ahmed, S.; Wang, N.; Lalonde, M.; Goldberg, V.M.; Haqqi, T.M. Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCG) Differentially Inhibits Interleukin-1β-Induced Expression of Matrix Metalloproteinase-1 and-13 in Human Chondrocytes. J. Pharmacol. Exp. Ther. 2004, 308, 767–773. [Google Scholar] [CrossRef]
- Bae, J.Y.; Han, D.-W.; Wakitani, S.; Nawata, M.; Hyon, S.H. Biological and Biomechanical Evaluations of Osteochondral Allografts Preserved in Cold Storage Solution Containing Epigallocatechin Gallate. Cell Transplant. 2010, 19, 681–689. [Google Scholar] [CrossRef]
- Vankemmelbeke, M.N.; Jones, G.C.; Fowles, C.; Ilic, M.Z.; Handley, C.J.; Day, A.J.; Knight, C.G.; Mort, J.S.; Buttle, D.J. Selective Inhibition of ADAMTS-1,-4 And-5 by Catechin Gallate Esters. Eur. J. Biochem. 2003, 270, 2394–2403. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.J.; Koh, R.H.; Kim, S.-H.; Kim, K.M.; Park, G.K.; Hwang, N.S. Injectable Anti-Inflammatory Hyaluronic Acid Hydrogel for Osteoarthritic Cartilage Repair. Mater. Sci. Eng. C 2020, 115, 111096. [Google Scholar] [CrossRef]
- Andriamanalijaona, R.; Kypriotou, M.; Baugé, C.; Renard, E.; Legendre, F.; Raoudi, M.; Boumediene, K.; Gatto, H.; Monginoux, P.; Pujol, J.-P. Comparative Effects of 2 Antioxidants, Selenomethionine and Epigallocatechin-Gallate, on Catabolic and Anabolic Gene Expression of Articular Chondrocytes. J. Rheumatol. 2005, 32, 1958–1967. [Google Scholar]
- Huang, G.-S.; Tseng, C.-Y.; Lee, C.-H.; Su, S.-L.; Lee, H.-S. Effects of (−)-Epigallocatechin-3-Gallate on Cyclooxygenase 2, PGE 2, and IL-8 Expression Induced by IL-1β in Human Synovial Fibroblasts. Rheumatol. Int. 2010, 30, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Haqqi, T.M. Epigallocatechin-3-Gallate Suppresses the Global Interleukin-1beta-Induced Inflammatory Response in Human Chondrocytes. Arthritis Res. Ther. 2011, 13, R93. [Google Scholar] [CrossRef] [PubMed]
- Heinecke, L.; Grzanna, M.; Au, A.; Mochal, C.; Rashmir-Raven, A.; Frondoza, C. Inhibition of Cyclooxygenase-2 Expression and Prostaglandin E2 Production in Chondrocytes by Avocado Soybean Unsaponifiables and Epigallocatechin Gallate. Osteoarthr. Cartil. 2010, 18, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Rasheed, N.; Al-Shobaili, H.A. Epigallocatechin-3-O-gallate Up-regulates MicroRNA-199a-3p Expression by Down-regulating the Expression of Cyclooxygenase-2 in Stimulated Human Osteoarthritis Chondrocytes. J. Cell. Mol. Med. 2016, 20, 2241–2248. [Google Scholar] [CrossRef]
- Ahmed, S.; Rahman, A.; Hasnain, A.; Lalonde, M.; Goldberg, V.M.; Haqqi, T.M. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits the IL-1β-Induced Activity and Expression of Cyclooxygenase-2 and Nitric Oxide Synthase-2 in Human Chondrocytes. Free Radic. Biol. Med. 2002, 33, 1097–1105. [Google Scholar] [CrossRef]
- Singh, R.; Ahmed, S.; Islam, N.; Goldberg, V.M.; Haqqi, T.M. Epigallocatechin-3-gallate Inhibits Interleukin-1β–Induced Expression of Nitric Oxide Synthase and Production of Nitric Oxide in Human Chondrocytes: Suppression of Nuclear Factor ΚB Activation by Degradation of the Inhibitor of Nuclear Factor ΚB. Arthritis Rheum. 2002, 46, 2079–2086. [Google Scholar] [CrossRef]
- Singh, R.; Ahmed, S.; Malemud, C.J.; Goldberg, V.M.; Haqqi, T.M. Epigallocatechin-3-gallate Selectively Inhibits Interleukin-1 Β-induced Activation of Mitogen Activated Protein Kinase Subgroup C-Jun N-terminal Kinase in Human Osteoarthritis Chondrocytes. J. Orthop. Res. 2003, 21, 102–109. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Y.; Zhang, M.; Ying, H. Green Tea Polyphenols Attenuate LPS-induced Inflammation through Upregulating MicroRNA-9 in Murine Chondrogenic ATDC5 Cells. J. Cell. Physiol. 2019, 234, 22604–22612. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.Y.; Han, D.-W.; Matsumura, K.; Wakitani, S.; Nawata, M.; Hyon, S.-H. Nonfrozen Preservation of Articular Cartilage by Epigallocatechin-3-Gallate Reversibly Regulating Cell Cycle and Nf-ΚB Expression. Tissue Eng. Part A 2010, 16, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Anbazhagan, A.N.; Akhtar, N.; Ramamurthy, S.; Voss, F.R.; Haqqi, T.M. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits Advanced Glycation End Product-Induced Expression of Tumor Necrosis Factor-α and Matrix Metalloproteinase-13 in Human Chondrocytes. Arthritis Res. Ther. 2009, 11, R71. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, Q.; Liu, L.; Wu, H.; Zheng, L. Effect of Epigallocatechin-3-Gallate on Proliferation and Phenotype Maintenance in Rabbit Articular Chondrocytes in Vitro. Exp. Ther. Med. 2015, 9, 213–218. [Google Scholar] [CrossRef][Green Version]
- Zheng, Y.; Xiao, L.; Yu, C.; Jin, P.; Qin, D.; Xu, Y.; Yin, J.; Liu, Z.; Du, Q. Enhanced Antiarthritic Efficacy by Nanoparticles of (−)-Epigallocatechin Gallate–Glucosamine–Casein. J. Agric. Food Chem. 2019, 67, 6476–6486. [Google Scholar] [CrossRef]
- Elder, S.; Clune, J.; Walker, J.; Gloth, P. Suitability of EGCG as a Means of Stabilizing a Porcine Osteochondral Xenograft. J. Funct. Biomater. 2017, 8, 43. [Google Scholar] [CrossRef]
- Sondag, G.R.; Haqqi, T.M. The Role of MicroRNAs and Their Targets in Osteoarthritis. Curr. Rheumatol. Rep. 2016, 18, 56. [Google Scholar] [CrossRef]
- Rasheed, Z. Green Tea Bioactive Polyphenol Epigallocatechin-3-o-Gallate in Osteoarthritis: Current Status and Future Perspectives. Int. J. Health Sci. 2016, 237, 1–4. [Google Scholar] [CrossRef]
- Akhtar, N.; Rasheed, Z.; Ramamurthy, S.; Anbazhagan, A.N.; Voss, F.R.; Haqqi, T.M. MicroRNA-27b Regulates the Expression of Matrix Metalloproteinase 13 in Human Osteoarthritis Chondrocytes. Arthritis Rheum. 2010, 62, 1361–1371. [Google Scholar] [CrossRef]
- Rasheed, Z.; Al-Shobaili, H.A.; Rasheed, N.; Mahmood, A.; Khan, M.I. MicroRNA-26a-5p Regulates the Expression of Inducible Nitric Oxide Synthase via Activation of NF-ΚB Pathway in Human Osteoarthritis Chondrocytes. Arch. Biochem. Biophys. 2016, 594, 61–67. [Google Scholar] [CrossRef]
- Park, S.J.; Cheon, E.J.; Lee, M.H.; Kim, H.A. MicroRNA-127-5p Regulates Matrix Metalloproteinase 13 Expression and Interleukin-1β–Induced Catabolic Effects in Human Chondrocytes. Arthritis Rheum. 2013, 65, 3141–3152. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Makki, M.S.; Haqqi, T.M. MicroRNA-602 and MicroRNA-608 Regulate Sonic Hedgehog Expression via Target Sites in the Coding Region in Human Chondrocytes. Arthritis Rheumatol. 2015, 67, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Zhang, Z.; Chen, W.; Huang, G.; He, A.; Hou, C.; Long, Y.; Yang, Z.; Liao, W. MicroRNA-320 Regulates Matrix Metalloproteinase-13 Expression in Chondrogenesis and Interleukin-1β-Induced Chondrocyte Responses. Osteoarthr. Cartil. 2016, 24, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Cheon, E.; Kim, H. MicroRNA-558 Regulates the Expression of Cyclooxygenase-2 and IL-1β-Induced Catabolic Effects in Human Articular Chondrocytes. Osteoarthr. Cartil. 2013, 21, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Makki, M.S.; Haseeb, A.; Haqqi, T.M. MicroRNA-9 Promotion of Interleukin-6 Expression by Inhibiting Monocyte Chemoattractant Protein–Induced Protein 1 Expression in Interleukin-1β–Stimulated Human Chondrocytes. Arthritis Rheumatol. 2015, 67, 2117–2128. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Meng, F.; Zhang, Z.; Kang, Y.; Chen, W.; Huang, G.; Fu, M.; Sheng, P.; Zhang, Z.; Liao, W. The Role of MicroRNA-381 in Chondrogenesis and Interleukin-1-β Induced Chondrocyte Responses. Cell. Physiol. Biochem. 2015, 36, 1753–1766. [Google Scholar] [CrossRef]
- Rasheed, Z.; Rasheed, N.; Al-Shaya, O. Epigallocatechin-3-O-Gallate Modulates Global MicroRNA Expression in Interleukin-1β-Stimulated Human Osteoarthritis Chondrocytes: Potential Role of EGCG on Negative Co-Regulation of MicroRNA-140-3p and ADAMTS5. Eur. J. Nutr. 2018, 57, 917–928. [Google Scholar] [CrossRef]
- Haqqi, T.M.; Anthony, D.D.; Gupta, S.; Ahmad, N.; Lee, M.-S.; Kumar, G.K.; Mukhtar, H. Prevention of Collagen-Induced Arthritis in Mice by a Polyphenolic Fraction from Green Tea. Proc. Natl. Acad. Sci. USA 1999, 96, 4524–4529. [Google Scholar] [CrossRef]
- Sobhi, A.; Mohamad, A.; Mehana, E.D.; Abdel-Raheim, M. The Protective Effect of Green Tea Extract against the Oxidative Stress of the Experimental Arthritic Rats. PMJ 2007, 3, 12–18. [Google Scholar]
- Leong, D.J.; Choudhury, M.; Hanstein, R.; Hirsh, D.M.; Kim, S.J.; Majeska, R.J.; Schaffler, M.B.; Hardin, J.A.; Spray, D.C.; Goldring, M.B. Green Tea Polyphenol Treatment Is Chondroprotective, Anti-Inflammatory and Palliative in a Mouse Posttraumatic Osteoarthritis Model. Arthritis Res. Ther. 2014, 16, 1–11. [Google Scholar] [CrossRef]
- Jin, X.; Wang, B.; Wang, X.; Antony, B.; Zhu, Z.; Han, W.; Cicuttini, F.; Wluka, A.; Winzenberg, T.; Blizzard, L. Associations between Endogenous Sex Hormones and MRI Structural Changes in Patients with Symptomatic Knee Osteoarthritis. Osteoarthr. Cartil. 2017, 25, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Hashempur, M.H.; Sadrneshin, S.; Mosavat, S.H.; Ashraf, A. Green Tea (Camellia Sinensis) for Patients with Knee Osteoarthritis: A Randomized Open-Label Active-Controlled Clinical Trial. Clin. Nutr. 2018, 37, 85–90. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Z.; Yang, K.; Shu, G.; Wang, S.; Gao, P.; Zhu, X.; Xi, Q.; Zhang, Y.; Jiang, Q. Epigallocatechin Gallate Reduces Slow-Twitch Muscle Fiber Formation and Mitochondrial Biosynthesis in C2C12 Cells by Repressing AMPK Activity and PGC-1α Expression. J. Agric. Food Chem. 2016, 64, 6517–6523. [Google Scholar] [CrossRef] [PubMed]
- Dorchies, O.M.; Wagner, S.; Buetler, T.M.; Ruegg, U.T. Protection of Dystrophic Muscle Cells with Polyphenols from Green Tea Correlates with Improved Glutathione Balance and Increased Expression of 67LR, a Receptor for (−)-epigallocatechin Gallate. Biofactors 2009, 35, 279–294. [Google Scholar] [CrossRef]
- Babu, G.S.; Ilaiyaraja, N.; Khanum, F.; Anand, T. Cytoprotective Propensity of Green Tea Polyphenols against Citrinin-Induced Skeletal-Myotube Damage in C2C12 Cells. Cytotechnology 2017, 69, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Gervasio, O.L.; Yeung, E.W.; Whitehead, N.P. Calcium and the Damage Pathways in Muscular Dystrophy. Can. J. Physiol. Pharmacol. 2010, 88, 83–91. [Google Scholar] [CrossRef]
- Nakae, Y.; Hirasaka, K.; Goto, J.; Nikawa, T.; Shono, M.; Yoshida, M.; Stoward, P.J. Subcutaneous Injection, from Birth, of Epigallocatechin-3-Gallate, a Component of Green Tea, Limits the Onset of Muscular Dystrophy in Mdx Mice: A Quantitative Histological, Immunohistochemical and Electrophysiological Study. Histochem. Cell Biol. 2008, 129, 489–501. [Google Scholar] [CrossRef]
- Dorchies, O.M.; Wagner, S.; Vuadens, O.; Waldhauser, K.; Buetler, T.M.; Kucera, P.; Ruegg, U.T. Green Tea Extract and Its Major Polyphenol (−)-Epigallocatechin Gallate Improve Muscle Function in a Mouse Model for Duchenne Muscular Dystrophy. Am. J. Physiol. Cell Physiol. 2006, 290, C616–C625. [Google Scholar] [CrossRef]
- Buetler, T.M.; Renard, M.; Offord, E.A.; Schneider, H.; Ruegg, U.T. Green Tea Extract Decreases Muscle Necrosis in Mdx Mice and Protects against Reactive Oxygen Species. Am. J. Clin. Nutr. 2002, 75, 749–753. [Google Scholar] [CrossRef]
- Kumaran, V.S.; Arulmathi, K.; Srividhya, R.; Kalaiselvi, P. Repletion of Antioxidant Status by EGCG and Retardation of Oxidative Damage Induced Macromolecular Anomalies in Aged Rats. Exp. Gerontol. 2008, 43, 176–183. [Google Scholar] [CrossRef]
- Ota, N.; Soga, S.; Haramizu, S.; Yokoi, Y.; Hase, T.; Murase, T. Tea Catechins Prevent Contractile Dysfunction in Unloaded Murine Soleus Muscle: A Pilot Study. Nutrition 2011, 27, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-L.; Cao, J.J.; Dagda, R.Y.; Chanjaplammootil, S.; Lu, C.; Chyu, M.-C.; Gao, W.; Wang, J.-S.; Yeh, J.K. Green Tea Polyphenols Benefits Body Composition and Improves Bone Quality in Long-Term High-Fat Diet–Induced Obese Rats. Nutr. Res. 2012, 32, 448–457. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Y.; Wan, X.; Yang, C.S.; Zhang, J. Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate Triggered Hepatotoxicity in Mice: Responses of Major Antioxidant Enzymes and the Nrf2 Rescue Pathway. Toxicol. Appl. Pharmacol. 2015, 283, 65–74. [Google Scholar] [CrossRef]
- Hong, K.-B.; Lee, H.-S.; Kim, D.H.; Moon, J.M.; Park, Y. Tannase-Converted Green Tea Extract with High (−)-Epicatechin Inhibits Skeletal Muscle Mass in Aged Mice. Evid. Based Complement. Alternat. Med. 2020, 2020, 4319398. [Google Scholar] [CrossRef] [PubMed]
- Hadi, A.; Pourmasoumi, M.; Kafeshani, M.; Karimian, J.; Maracy, M.R.; Entezari, M.H. The Effect of Green Tea and Sour Tea (Hibiscus Sabdariffa L.) Supplementation on Oxidative Stress and Muscle Damage in Athletes. J. Diet. Suppl. 2017, 14, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Jówko, E.; Długołęcka, B.; Makaruk, B.; Cieśliński, I. The Effect of Green Tea Extract Supplementation on Exercise-Induced Oxidative Stress Parameters in Male Sprinters. Eur. J. Nutr. 2015, 54, 783–791. [Google Scholar] [CrossRef]
- Panza, V.S.P.; Wazlawik, E.; Schütz, G.R.; Comin, L.; Hecht, K.C.; da Silva, E.L. Consumption of Green Tea Favorably Affects Oxidative Stress Markers in Weight-Trained Men. Nutrition 2008, 24, 433–442. [Google Scholar] [CrossRef]
- Kuo, Y.-C.; Lin, J.-C.; Bernard, J.R.; Liao, Y.-H. Green Tea Extract Supplementation Does Not Hamper Endurance-Training Adaptation but Improves Antioxidant Capacity in Sedentary Men. Appl. Physiol. Nutr. Metab. 2015, 40, 990–996. [Google Scholar] [CrossRef]
- Da Silva, W.; Machado, Á.S.; Souza, M.A.; Mello-Carpes, P.B.; Carpes, F.P. Effect of Green Tea Extract Supplementation on Exercise-Induced Delayed Onset Muscle Soreness and Muscular Damage. Physiol. Behav. 2018, 194, 77–82. [Google Scholar] [CrossRef]
- Cao, H.; Kelly, M.A.; Kari, F.; Dawson, H.D.; Urban, J.F.; Coves, S.; Roussel, A.M.; Anderson, R.A. Green Tea Increases Anti-Inflammatory Tristetraprolin and Decreases pro-Inflammatory Tumor Necrosis Factor MRNA Levels in Rats. J. Inflamm. 2007, 4, 1. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, M.; Wang, R.; Li, M.; Li, D.; Xie, Z. Dietary Supplement of Yunkang 10 Green Tea and Treadmill Exercise Ameliorate High Fat Diet Induced Metabolic Syndrome of C57BL/6 J Mice. Nutr. Metab. 2020, 17, 1–15. [Google Scholar]
- Haramizu, S.; Ota, N.; Hase, T.; Murase, T. Catechins Suppress Muscle Inflammation and Hasten Performance Recovery after Exercise. Med. Sci. Sports Exerc. 2013, 45, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lai, Y.-J.; Chan, Y.-L.; Li, T.-L.; Wu, C.-J. Epigallocatechin-3-Gallate Effectively Attenuates Skeletal Muscle Atrophy Caused by Cancer Cachexia. Cancer Lett. 2011, 305, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.P.; Call, J.A.; Bassaganya-Riera, J.; Robertson, J.L.; Grange, R.W. Green Tea Extract Decreases Muscle Pathology and NF-ΚB Immunostaining in Regenerating Muscle Fibers of Mdx Mice. Clin. Nutr. 2010, 29, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.Q.; Kumar, A.V.; Mills, J.; Lapierre, L.R. Autophagy in Aging and Longevity. Hum. Genet. 2020, 139, 277–290. [Google Scholar] [CrossRef]
- Carnio, S.; LoVerso, F.; Baraibar, M.A.; Longa, E.; Khan, M.M.; Maffei, M.; Reischl, M.; Canepari, M.; Loefler, S.; Kern, H. Autophagy Impairment in Muscle Induces Neuromuscular Junction Degeneration and Precocious Aging. Cell Rep. 2014, 8, 1509–1521. [Google Scholar] [CrossRef]
- Alway, S.E. Antioxidants and Polyphenols Mediate Mitochondrial Mediated Muscle Death Signaling in Sarcopenia. In Nutrition and Skeletal Muscle; Elsevier: New York, NY, USA, 2019; pp. 439–494. [Google Scholar]
- Bartholome, A.; Kampkötter, A.; Tanner, S.; Sies, H.; Klotz, L.-O. Epigallocatechin Gallate-Induced Modulation of FoxO Signaling in Mammalian Cells and C. Elegans: FoxO Stimulation Is Masked via PI3K/Akt Activation by Hydrogen Peroxide Formed in Cell Culture. Arch. Biochem. Biophys. 2010, 501, 58–64. [Google Scholar] [CrossRef]
- Leone, M.; Zhai, D.; Sareth, S.; Kitada, S.; Reed, J.C.; Pellecchia, M. Cancer Prevention by Tea Polyphenols Is Linked to Their Direct Inhibition of Antiapoptotic Bcl-2-Family Proteins. Cancer Res. 2003, 63, 8118–8121. [Google Scholar]
- Townsend, J.R.; Stout, J.R.; Jajtner, A.R.; Church, D.D.; Beyer, K.S.; Riffe, J.J.; Muddle, T.W.; Herrlinger, K.L.; Fukuda, D.H.; Hoffman, J.R. Polyphenol Supplementation Alters Intramuscular Apoptotic Signaling Following Acute Resistance Exercise. Physiol. Rep. 2018, 6, e13552. [Google Scholar] [CrossRef]
- Mirza, K.A.; Pereira, S.L.; Edens, N.K.; Tisdale, M.J. Attenuation of Muscle Wasting in Murine C 2 C 12 Myotubes by Epigallocatechin-3-Gallate. J. Cachexia Sarcopenia Muscle 2014, 5, 339–345. [Google Scholar] [CrossRef]
- Hood, D.A.; Memme, J.M.; Oliveira, A.N.; Triolo, M. Maintenance of Skeletal Muscle Mitochondria in Health, Exercise, and Aging. Annu. Rev. Physiol. 2019, 81, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.D.; Duan, J.; Samuelson, A.T.; Gaffrey, M.J.; Merrihew, G.E.; Egertson, J.D.; Wang, L.; Bammler, T.K.; Moore, R.J.; White, C.C. Improving Mitochondrial Function with SS-31 Reverses Age-Related Redox Stress and Improves Exercise Tolerance in Aged Mice. Free Radic. Biol. Med. 2019, 134, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.C.; Gonzalo-Benito, H.; Jové, M.; Fourcade, S.; Cassanyé, A.; Boada, J.; Delgado, M.A.; Espinel, A.E.; Pamplona, R.; Portero-Otín, M. Dietary Intake of Green Tea Polyphenols Regulates Insulin Sensitivity with an Increase in AMP-activated Protein Kinase α Content and Changes in Mitochondrial Respiratory Complexes. Mol. Nutr. Food Res. 2013, 57, 459–470. [Google Scholar] [CrossRef]
- Sae-Tan, S.; Grove, K.A.; Kennett, M.J.; Lambert, J.D. (−)-Epigallocatechin-3-Gallate Increases the Expression of Genes Related to Fat Oxidation in the Skeletal Muscle of High Fat-Fed Mice. Food Funct. 2011, 2, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Bezzina, R.; Hinch, E.; Lewandowski, P.A.; Cameron-Smith, D.; Mathai, M.L.; Jois, M.; Sinclair, A.J.; Begg, D.P.; Wark, J.D. Green Tea, Black Tea, and Epigallocatechin Modify Body Composition, Improve Glucose Tolerance, and Differentially Alter Metabolic Gene Expression in Rats Fed a High-Fat Diet. Nutr. Res. 2009, 29, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Meador, B.; Mirza, K.; Tian, M.; Skelding, M.; Reaves, L.; Edens, N.; Tisdale, M.; Pereira, S. The Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCg) Attenuates Skeletal Muscle Atrophy in a Rat Model of Sarcopenia. J. Frailty Aging 2015, 4, 209. [Google Scholar]
- Sae-tan, S.; Rogers, C.J.; Lambert, J.D. Voluntary Exercise and Green Tea Enhance the Expression of Genes Related to Energy Utilization and Attenuate Metabolic Syndrome in High Fat Fed Mice. Mol. Nutr. Food Res. 2014, 58, 1156–1159. [Google Scholar] [CrossRef]
- Murase, T.; Haramizu, S.; Ota, N.; Hase, T. Tea Catechin Ingestion Combined with Habitual Exercise Suppresses the Aging-Associated Decline in Physical Performance in Senescence-Accelerated Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R281–R289. [Google Scholar] [CrossRef]
- Murase, T.; Haramizu, S.; Shimotoyodome, A.; Nagasawa, A.; Tokimitsu, I. Green Tea Extract Improves Endurance Capacity and Increases Muscle Lipid Oxidation in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R708–R715. [Google Scholar] [CrossRef]
- Murase, T.; Haramizu, S.; Shimotoyodome, A.; Tokimitsu, I.; Hase, T. Green Tea Extract Improves Running Endurance in Mice by Stimulating Lipid Utilization during Exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1550–R1556. [Google Scholar] [CrossRef]
- Call, J.A.; Voelker, K.A.; Wolff, A.V.; McMillan, R.P.; Evans, N.P.; Hulver, M.W.; Talmadge, R.J.; Grange, R.W. Endurance Capacity in Maturing Mdx Mice Is Markedly Enhanced by Combined Voluntary Wheel Running and Green Tea Extract. J. Appl. Physiol. 2008, 105, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, L.; Ramirez-Sanchez, I.; Perkins, G.A.; Murphy, A.; Taub, P.R.; Ceballos, G.; Villarreal, F.J.; Hogan, M.C.; Malek, M.H. (–)-Epicatechin Enhances Fatigue Resistance and Oxidative Capacity in Mouse Muscle. J. Physiol. 2011, 589, 4615–4631. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-W.; Chang, C.-C.; Liao, S.-F.; Liao, Y.-H.; Hou, C.-W.; Tsao, J.-P.; Cheng, I.-S. Effect of Green Tea Extract Supplementation on Glycogen Replenishment in Exercised Human Skeletal Muscle. Br. J. Nutr. 2017, 117, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.C.; Rudnicki, M.A. Satellite Cells: The Architects of Skeletal Muscle. In Current Topics in Developmental Biology; Elsevier: New York, NY, USA, 2014; Volume 107, pp. 161–181. [Google Scholar]
- Collins-Hooper, H.; Woolley, T.E.; Dyson, L.; Patel, A.; Potter, P.; Baker, R.E.; Gaffney, E.A.; Maini, P.K.; Dash, P.R.; Patel, K. Age-related Changes in Speed and Mechanism of Adult Skeletal Muscle Stem Cell Migration. Stem Cells 2012, 30, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.-B.; Lee, H.-S.; Hong, J.S.; Kim, D.H.; Moon, J.M.; Park, Y. Effects of Tannase-Converted Green Tea Extract on Skeletal Muscle Development. BMC Complement. Med. Ther. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Kim, A.R.; Kim, K.M.; Byun, M.R.; Hwang, J.-H.; Park, J.I.; Oh, H.T.; Kim, H.K.; Jeong, M.G.; Hwang, E.S.; Hong, J.-H. Catechins Activate Muscle Stem Cells by Myf5 Induction and Stimulate Muscle Regeneration. Biochem. Biophys. Res. Commun. 2017, 489, 142–148. [Google Scholar] [CrossRef]
- Kim, A.R.; Kim, K.M.; Byun, M.R.; Hwang, J.-H.; Park, J.I.; Oh, H.T.; Jeong, M.G.; Hwang, E.S.; Hong, J.-H. (-)-Epigallocatechin-3-Gallate Stimulates Myogenic Differentiation through TAZ Activation. Biochem. Biophys. Res. Commun. 2017, 486, 378–384. [Google Scholar] [CrossRef]
- Alway, S.E.; Bennett, B.T.; Wilson, J.C.; Sperringer, J.; Mohamed, J.S.; Edens, N.K.; Pereira, S.L. Green Tea Extract Attenuates Muscle Loss and Improves Muscle Function during Disuse, but Fails to Improve Muscle Recovery Following Unloading in Aged Rats. J. Appl. Physiol. 2015, 118, 319–330. [Google Scholar] [CrossRef]
- Onishi, S.; Ishino, M.; Kitazawa, H.; Yoto, A.; Shimba, Y.; Mochizuki, Y.; Unno, K.; Meguro, S.; Tokimitsu, I.; Miura, S. Green Tea Extracts Ameliorate High-Fat Diet–Induced Muscle Atrophy in Senescence-Accelerated Mouse Prone-8 Mice. PLoS ONE 2018, 13, e0195753. [Google Scholar] [CrossRef]
- Bhattacharya, T.K.; Pence, B.D.; Ossyra, J.M.; Gibbons, T.E.; Perez, S.; McCusker, R.H.; Kelley, K.W.; Johnson, R.W.; Woods, J.A.; Rhodes, J.S. Exercise but Not (–)-Epigallocatechin-3-Gallate or β-Alanine Enhances Physical Fitness, Brain Plasticity, and Behavioral Performance in Mice. Physiol. Behav. 2015, 145, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Alway, S.E.; Bennett, B.T.; Wilson, J.C.; Edens, N.K.; Pereira, S.L. Epigallocatechin-3-Gallate Improves Plantaris Muscle Recovery after Disuse in Aged Rats. Exp. Gerontol. 2014, 50, 82–94. [Google Scholar] [CrossRef]
- Mirza, K.A.; Luo, M.; Pereira, S.; Voss, A.; Das, T.; Tisdale, M.J. In Vitro Assessment of the Combined Effect of Eicosapentaenoic Acid, Green Tea Extract and Curcumin C3 on Protein Loss in C 2 C 12 Myotubes. Vitro Cell. Dev. Biol. Anim. 2016, 52, 838–845. [Google Scholar] [CrossRef]
- Chen, S.; Minegishi, Y.; Hasumura, T.; Shimotoyodome, A.; Ota, N. Involvement of Ammonia Metabolism in the Improvement of Endurance Performance by Tea Catechins in Mice. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Wimbley, T.D.; Graham, D.Y. Diagnosis and Management of Iron Deficiency Anemia in the 21st Century. Ther. Adv. Gastroenterol. 2011, 4, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Harder, S.; Fuhr, U.; Staib, A.H.; Wolff, T. Ciprofloxacin-Caffeine: A Drug Interaction Established Using in Vivo and in Vitro Investigations. Am. J. Med. 1989, 87, S89–S91. [Google Scholar] [CrossRef]
- Carrillo, J.A.; Benitez, J. Clinically Significant Pharmacokinetic Interactions between Dietary Caffeine and Medications. Clin. Pharmacokinet. 2000, 39, 127–153. [Google Scholar] [CrossRef] [PubMed]
- Hägg, S.; Spigset, O.; Mjörndal, T.; Dahlqvist, R. Effect of Caffeine on Clozapine Pharmacokinetics in Healthy Volunteers. Br. J. Clin. Pharmacol. 2000, 49, 59–63. [Google Scholar] [CrossRef]
- Taylor, J.R.; Wilt, V.M. Probable Antagonism of Warfarin by Green Tea. Ann. Pharmacother. 1999, 33, 426–428. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, M.; Yamashiro, N.; Kobayashi, F.; Nagasaka, T.; Takiyama, Y. A Case of Hypokalemic Myopathy Induced by Excessive Drinking of a Beverage Containing Green Tea Extract. Rinsho Shinkeigaku 2013, 53, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.K.; Salwan, P.; Salwan, S. Various Possible Toxicants Involved in Thyroid Dysfunction: A Review. J. Clin. Diagn. Res. JCDR 2016, 10, FE01. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.J.; Coates, A.M.; Kohler, M.; Banks, S. Caffeine Consumption and Sleep Quality in Australian Adults. Nutrients 2016, 8, 479. [Google Scholar] [CrossRef] [PubMed]
- Murao, T.; Sakurai, K.; Mihara, S.; Marubayashi, T.; Murakami, Y.; Sasaki, Y. Lifestyle Change Influences on GERD in Japan: A Study of Participants in a Health Examination Program. Dig. Dis. Sci. 2011, 56, 2857–2864. [Google Scholar] [CrossRef]
- Cho, Y.; Ko, I.; Kim, S.; Hwan, L.; Shin, M.; Kim, C.; Kim, S.; Jin, J.; Chung, J.; Kim, K. Caffeine Enhances Micturition through Neuronal Activation in Micturition Centers. Mol. Med. Rep. 2014, 10, 2931–2936. [Google Scholar] [CrossRef] [PubMed]
- Karadas, M.; Seven, N. The Effect of Different Drinks on Tooth Color after Home Bleaching. Eur. J. Dent. 2014, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Steves, C.J.; Bird, S.; Williams, F.M.; Spector, T.D. The Microbiome and Musculoskeletal Conditions of Aging: A Review of Evidence for Impact and Potential Therapeutics. J. Bone Miner. Res. 2016, 31, 261–269. [Google Scholar] [CrossRef]
- Berthelot, J.-M.; Sellam, J.; Maugars, Y.; Berenbaum, F. Cartilage-Gut-Microbiome Axis: A New Paradigm for Novel Therapeutic Opportunities in Osteoarthritis. RMD Open 2019, 5, e001037. [Google Scholar] [CrossRef] [PubMed]
- Szychlinska, M.A.; Di Rosa, M.; Castorina, A.; Mobasheri, A.; Musumeci, G. A Correlation between Intestinal Microbiota Dysbiosis and Osteoarthritis. Heliyon 2019, 5, e01134. [Google Scholar] [CrossRef] [PubMed]
- Boer, C.G.; Radjabzadeh, D.; Medina-Gomez, C.; Garmaeva, S.; Schiphof, D.; Arp, P.; Koet, T.; Kurilshikov, A.; Fu, J.; Ikram, M.A. Intestinal Microbiome Composition and Its Relation to Joint Pain and Inflammation. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut Microbiota Contribute to Age-Related Changes in Skeletal Muscle Size, Composition, and Function: Biological Basis for a Gut-Muscle Axis. Calcif. Tissue Int. 2018, 102, 433–442. [Google Scholar] [CrossRef]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging Gut Microbiota at the Cross-Road between Nutrition, Physical Frailty, and Sarcopenia: Is There a Gut–Muscle Axis? Nutrients 2017, 9, 1303. [Google Scholar] [CrossRef]
- Le Bras, A. A Gut Microbiota–Skeletal Muscle Axis. Sci. Transl. Med. 2019, 11, eaan5662. [Google Scholar] [CrossRef]
- De Sire, R.; Rizzatti, G.; Ingravalle, F.; Pizzoferrato, M.; Petito, V.; Lopetuso, L.; Graziani, C.; De Sire, A.; Mentella, M.C.; Mele, M.C. Skeletal Muscle-Gut Axis: Emerging Mechanisms of Sarcopenia for Intestinal and Extra Intestinal Diseases. Minerva Gastroenterol. Dietol. 2018, 64, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tang, L.; Shen, C.-L.; Wang, J.-S. Green Tea Polyphenols Modify Gut-Microbiota Dependent Metabolisms of Energy, Bile Constituents and Micronutrients in Female Sprague–Dawley Rats. J. Nutr. Biochem. 2018, 61, 68–81. [Google Scholar] [CrossRef]
- Wang, J.; Tang, L.; Zhou, H.; Zhou, J.; Glenn, T.C.; Shen, C.-L.; Wang, J.-S. Long-Term Treatment with Green Tea Polyphenols Modifies the Gut Microbiome of Female Sprague-Dawley Rats. J. Nutr. Biochem. 2018, 56, 55–64. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Green Tea | Black Tea | Yerba Mate Tea | |
|---|---|---|---|
| Processing of tea leaves | Steamed or pan-fried; not oxidized. | Withered and fermented and not blanched before drying; oxidized. | Flash heated over an open flame; semioxidized. |
| Types of polyphenols | Caffeic acid, caffeine, catechin, coumaric acid, EGCG, EC, EGC, ECG, gallic acid, gallocatechin gallate, kaempferol, myricetin, quercetin, quinic acid, rutin. | Caffeine, catechin, catechin gallate, ECG, gallic acid, gallocatechin gallate, kaempferol, myricetin, procyanidin, quercetin, rutin, theaflavin, theobromine. | Caffeic acid, caffeine, caffeoyl derivatives, caffeoylshikimic acid, chlorogenic acid, feruloylquinic acid, kaempferol, quercetin, quinic acid, rutin, theobromine. |
| First Author, Year of Study, Citation | Experimental Design and Treatments | Effects of Green Tea or EGCG |
|---|---|---|
| Cellular Studies | ||
| Adcocks, 2002, [58] | Model: Bovine cartilage explants cotreated with TNF-α (3 nM) Treatments: EGCG (0.2, 2, 20, or 200 µM) for 5 days Model: Human cartilage from an OA knee joint cotreated with IL-1β (3 nM), TNF-α (6 nM) Treatment: EGCG (20 µM) for 9 days | Compared to the control group, the EGCG groups showed: ↓ Proteoglycan and type II collagen degradation |
| Ahmed, 2002, [68] | Model: Human primary chondrocytes cotreated with IL-1β (5 ng/mL) Treatments: Control, EGCG (20, 50, 100, or 200 μM) for 24 h | Compared to the control group, the EGCG groups showed: ↓ iNOS expression and NO production ↓ COX-2 expression and PGE2 production ↓ LDH release |
| Ahmed, 2004, [59] | Model: Human primary chondrocytes or human cartilage explants cotreated with IL-1β (50 or 10 ng/mL) Treatments: EGCG (20, 50, 100, or 200 μM) for 24 or 72 h | Compared to the control group, the EGCG groups showed: ↓ mRNA and protein expression of MMP-1 and MMP-13 in chondrocytes ↓ GAG release from human cartilage explants ↓ Transcription activity of NF-κB and AP-1 |
| Akhtar, 2011, [65] | Model: Human primary chondrocytes with IL-1β (5 ng/mL) after EGCG Treatments: Control, EGCG (10–100 μM) for 24 h | Compared to the IL-1β group, the IL-1β + EGCG groups showed: ↓ ENA-78, GM-CSF, GRO, GRO-α, IL-6, IL-8, MCP1, MCP-3, MIP-1β, GCP-2, MIP-3α, IP-10, NAP-2, LIF via ↓ activation of NF-κB and JNK-MAPK |
| Andriamanalijaona, 2005, [63] | Model: Bovine primary articular chondrocytes Treatments: Pretreated with EGCG (20 or 50 µM) for 24 h and then cotreated with by IL-1β (10 ng/mL) for 24 h | Compared to the control group, the EGCG groups showed: ↑ Type II collagen and aggrecan core protein expression ↑ mRNA expression of TGF-β1, TGF-β2, TGF-βR1, and TGF-βII ↓ mRNA levels of MMP-1, MMP-3, MMP-13, aggrecanase-1, aggrecanase-2, iNOS, COX-1, COX-2 (anti-inflammatory effect) ↓ MAPK (Erk1/Erk2, p38 kinase), NF-κB, and AP-1 activity |
| Bae, 2010a, [72] | Model: Primary human articular cartilage with or without OA treated with EGCG. Then, cartilage is implanted to cartilage defects in rabbits Treatments: Control (no EGCG) and EGCG (1 mM) at 4 °C for 1–4 weeks | Compared to the control group, the EGCG group showed: Preserving and repairing articular cartilage by reversibly regulating cell cycle at G0/G1 phase and NF-κB expression |
| Bae, 2010b, [60] | Model: Human articular cartilage Treatments: Control (no RGCG), cotreated EGCG (1 mM), and pretreated EGCG for 2 weeks | Compared to the control group, the EGCG group showed: ↑ GAG content and total collagen ↓ Denaturation of extracellular matrix |
| Elder, 2017, [76] | Model: Porcine osteochondral xenograft Treatments: Control (no EGCG), EGCG at 0.04%, 0.2%, and 1% w/v | Compared to the control group, the EGCG group showed: ↑ Compressive resistance of decellularized porcine cartilage ↑ Fixation on the decellularized porcine cartilage coefficient of friction against glass ↑ Strength of cartilage–bone interference in shear ↑ Mechanical properties |
| Heinecke, 2010, [66] | Model: Equine primary articular cartilage Treatments: Pretreated with EGCG (4, 40, or 400 ng/mL) for 24 h and then cotreated with IL-1β (10 ng/mL) and TNF-α (1 ng/mL) for 24 h | Compared to the control group, the EGCG groups showed: ↓ COX-2 expression and PGE2 production ↓ NF-κB translocation to the nucleus |
| Huang, 2010, [64] | Model: Human primary osteoarthritic synovial adherent cells Treatments: Pretreated with EGCG (10, 20, or 50 μM) for 12 h and then cotreated with IL-1β for 12 h | Compared to the control group, the EGCG groups showed: ↓ COX-2 upregulation ↓ PGE2 and IL-8 production ↓ Phosphorylation of IKKβ |
| Huang, 2015, [74] | Model: Primary rabbit articular chondrocytes Treatments: Control, EGCG (5–20 µM) for 2, 4, and 6 days | Compared to the control group, the EGCG groups showed: ↑ Chondrocyte growth and synthesis of the cartilage extracellular matrix ↑ Expression of aggrecan, collagen II, and SOX9 genes ↓ Gene expression of collagen I ↓ Dedifferentiation of chondrocytes |
| Jin, 2020, [62] | Model: Primary porcine 3D encapsulated articular chondrocytes stimulated with IL-1β (10 ng/mL) Treatments: Control (no HA), EGCG (50 µM) + 2%HA, EGCG + 5%HA | Compared to the no HA group, the EGCG + 5%HA group showed: ↓ Expression of proinflammatory genes (COX2, TNF-α, MMP1, MMP13, ADMTS5) ↑ Chondrogenic regeneration via ↑ gene expression of COL2, SOX9, ACAN |
| Rasheed, 2009, [73] | Model: Human primary chondrocytes stimulated with ACE for 8 h Treatments: Pretreated with EGCG (25, 75, or 150 µM) for 2 h | Compared to the control group, the EGCG groups showed: ↓ Gene expression and production of TNF-α and MMP-13 via ↓ p38-MAPK and JNK activation ↓ IκBα protein degradation in the cytoplasm, followed via ↓ activation and translocation of NF-κB to the nucleus |
| Rasheed, 2016, [67] | Model: Human primary chondrocytes with and without IL-1β (5 ng/mL) Transfected with miRNA inhibitors Treatments: Control, IL-1β, IL-1β + EGCG (20–50 μM) for 24 h | Compared to the IL-1β group, the IL-1β + EGCG groups showed: ↓ COX-2 expression, PGE2 production via ↑ hsa-miRNA-199a-3p expression |
| Rasheed, 2018, [87] | Model: Human primary chondrocytes with and without IL-1β (5 ng/mL) Treatments: Control, IL-1β, IL-1β + EGCG (20–50 μM) for 24 h | Compared to the IL-1β group, the IL-1β + EGCG groups showed: ↑ Expression 19 miRNAs, ↓ 17 miRNAs, and no change in 1311 miRNAs ↓ ADAMTS5 expression ↑ hsa-miRNA-140-3p expression |
| Singh, 2002, [69] | Model: Human primary chondrocytes cotreated with IL-1β (2 ng/mL) Treatments: Control, EGCG (1, 10, 50, or 100 μM) for 12 or 24 h | Compared to the control group, the EGCG groups showed: ↓ iNOS expression and NO production ↓ IκBα protein degradation in the cytoplasm, followed by activation and translocation of NF-κB to the nucleus |
| Singh, 2003, [70] | Model: Human primary chondrocytes cotreated with IL-1β (2 ng/mL) Treatments: Control, EGCG (100 µM) for 30 min | Compared to the control group, the EGCG groups showed: ↓ Phosphorylation of JNK isoforms ↓ Accumulation of hosphor-c-Jun and DNA binding activity of AP-1 ↔ Activation of extracellular signal-regulated kinase p44/p42 (ERKp44/p42) or p38-MAPK |
| Zhang, 2019, [71] | Model: LPS-stimulated ATDC5 cells to mimic an inflammatory response during OA Treatments: Control, pre-GTP (100 µg/mL) for 24 hour, LPS, and GTP + LPS | Compared to the LPS group, the GTP + LPS group showed: ↓ Cell damage ↓ MAPK and NF-κB cascades via positively regulating miRNA-9 |
| Zheng, 2019, [75] | Model: Human fibroblast-like synoviocyte-OA cells Treatments: EGCG (4 mg/mL), GA, EGCG + GA, EGCG + GA-NPs for 72 h | Compared to the EGCG group, the EGCG + GA-NPs group showed: ↓ Arthritic activity |
| Animal Studies | ||
| Haqqi, 1999, [88] | Model: A mouse model of collagen-induced arthritis Treatments: Control, GTP (0.2% in water) for 7 weeks | Compared to the control group, the GTP group showed: ↓ Arthritic incidence ↓ Total IgG and type II collagen-specific IgG levels in serum and arthritic joints ↓ Expression of proinflammatory genes (COX2, TNF-α, IFN-γ) in arthritic joints ↓ Joint infiltration by TNF-α and IFN-γ-producing cells in arthritic joints ↑ Neutral endopeptidase activity in arthritic joints |
| Jin, 2020, [62] | Model: A rat model of surgically induced OA Treatments: Control (no HA), EGCG + 5%HA for 3 weeks | Compared to the no HA group, the EGCG + HA group showed: ↓ Cartilage loss (GAG, type II collagen) ↓ Expression of collagen I and X |
| Leong, 2014, [90] | Model: A mouse post-traumatic OA model using the destabilization of the medial meniscus (DMM) Male C57BL/6 mice (5–6 months old) Treatments: Sham, sham + EGCG (25 mg/kg), DMN, DMN + EGCG (25 mg/kg) for 4 and 8 weeks | Compared to the DMN group, the DMN + EGCG group showed: ↓ Progression in early and midstage OA as shown by decreased Safranin O staining and OARSI scores ↓ OA-associated pain symptoms as shown by higher locomotor behavior (distance traveled) ↓ Degradation of both type II collagen and aggrecan in the articular cartilage matrix ↓ Staining of MMP-13 and ADAMTS5 in the articular cartilage ↓ Gene expression of MMP-1, MMP-3, MMP-8, MMP-13, ADAMTS5, IL-1β, and TNF-α in the articular cartilage ↑ Gene expression of MMP-repressing transcriptional regulator CITED2 in the articular cartilage ↓ CCR2 and IL-1β and TNF-α in the DRG |
| Sobhi, 2007, [89] | Model: A mouse model of intra-articular carrageenan-induced OA Rats (150–200 g) Treatments: Control, OA, OA + GTE (1.5% w/v) for 3 weeks | Compared to the OA group, the OA + GTE group showed: ↓ Plasma lipid peroxides, NO, and total thiols ↓ Numbers of inflammatory cells infiltrating the synovial membrane at arthritic joint ↓ Erosive necrosis, chondrodysplasia, congestion, edema, mineralization, and multinucleated giant cells infiltration No cartilage and bone erosion |
| Human Studies | ||
| Hashempur, 2018, [92] | Model: Randomized open-label active-controlled clinical trial Adults (n = 55) with knee OA Treatments: Control group (diclofenac at 100 mg/d) and intervention group (GTE at 1500 mg/d plus diclofenac) for 4 weeks | Compared to the control group, the intervention group showed: ↓ VAS pain, total WOMAC, WOMAC-physical function scores ↔WOMAC-pain, WOMAC-stiffness |
| Takiguchi, 2019, [18] | Model: 5-year cohort study (n = 11,091) (age range 40–72 years) with no history of knee OA | In men only, but not in women, older age, higher BMI, higher METs score, less smoking, and lower green tea consumption were associated with incident knee OA |
| First Author, Year of Study, Citation/Cell Type | Effect of EGCG on Selected miRNA Expression |
|---|---|
| Rasheed, 2018 [87], human osteoarthritic chondrocytes | ↑ Upregulated expression of the following miRNA: hsa-let-7a-5p; hsa-let-7b-5p; hsa-let-7c; hsa-let-7d-5p; hsa-let-7f-5p; hsa-let-7i-5p; hsa-miR-100-5p; hsa-miR-140-3p; hsa-miR-193a-3p; hsa-miR-199a-3p; hsa-miR-27b-3p; hsa-miR-29a-3p; hsa-miR-320b; hsa-miR-34a-5p; hsa-miR-3960; hsa-miR-4284; hsa-miR-4454; hsa-miR-497-5p; hsa-miR-5100 ↓ Downregulated expression of the following miRNA: hsa-let-7e-5p; hsa-miR-103a-3p; hsa-miR-125b-5p; hsa-miR-151a-5p; hsa-miR-195-5p; hsa-miR-222-3p; hsa-miR-23a-3p; hsa-miR-23b-3p; hsa-miR-26a-5p; hsa-miR-27a-3p; hsa-miR-29b-3p; hsa-miR-3195; hsa-miR-3651; hsa-miR-4281; hsa-miR-4459; hsa-miR-4516; hsa-miR-762 |
| Zhang, 2019, [71], murine chondrogenic ATDC5 cells | ↑ Upregulated expression of miRNA-9 |
| Rasheed, 2016, [67], human osteoarthritic chondrocytes | ↑ Upregulated expression of hsa-miR-199a-3p |
| First Author, Year, Citation | Experimental Design and Treatments | Effects of Green Tea or EGCG |
|---|---|---|
| Cellular Studies | ||
| Babu, 2017, [95] | Model: C2C12 skeletal muscle cells Treatment: Pretreated with or without GTE (20, 40, and 80 μg/mL) for 2 h followed by with or without citrinin treatment (0, 25, 50, 75, and 100 μM) for 24 h | Compared to citrinin the treated without GTE group, the citrinin-treated GTE group showed: ↑ Cell viability dose-dependently ↓ Citrinin-induced LDH release ↑ Myotube integrity and cell morphology ↓ ROS 3-fold at 80 μg/mL ↓ Citrinin-induced antioxidant enzyme activity |
| Dorchies, 2009, [94] | Model: Primary culture muscle cells from an mdx mouse model treated with H2O2 Treatment: Control (no EGCG) or EGCG (10 mg/mL) for 48 h | Compared to the control group, the EGCG group showed: ↑ Glutathione content ↑ Cell survival ↑ 67LR (a receptor for EGCG) |
| Hong, 2020, [137] | Model: C2C12 skeletal muscle cells with or without 100 μM of H2O2 for 48 h Treatment: Control (no EGCG), GTE or tannase-converted GTE (1, 5, 10, 15, and 20 μg/mL) for 24 h | Compared to the control group, the H2O2-treated tannase-converted GTE group showed: ↑ Myotube density and fusion ↑ mRNA expression of myogenin, Myf5, MyoD ↑ FOXO1 and FOXO3 transcription levels ↓ Oxidative-stress-induced ↓ AMPK |
| Kim, 2017, [139] | Model: Satellite cells isolated from 8-week-old C57BL/6 mice Treatment: Control (no EGCG) and EGCG (10 μM) for 72 h | Compared to the control group, the EGCG group showed: ↑ MHC (myogenic factor) ↑ TAZ localization TAZ knockdown inhibits EGCG-induced myogenic differentiation |
| Kim, 2017, [138] | Model: C2C12 myoblasts Treatment: Control (no EGCG) and EGCG (10 μM) for 24 h | Compared to the control group, the EGCG group showed: ↑ MHC content ↑ Myogenin and muscle creatine kinase gene expression ↑ Number of multinucleated cells |
| Mirza, 2014, [121] | Model: C2C12 skeletal muscle cells with or without starvation media and TNF-α Treatment: Control (no EGCG) or EGCG (10–150 μM) for 24 h | Compared to the control group, the EGCG group showed: ↓ Protein degradation at 10 μM ↑ Protein degradation at >10 μM ↑ pAkt and pFoxO3a expression at >10 μM ↔ pAkt and pFoxO3a expression at 10 μM |
| Mirza, 2016, [144] | Model: C2C12 skeletal muscle cells treated with or without PIF (4.2 nM) or TNF-α (25 ng/mL) Treatment: Control (no EGCG) and EGCG (10 μM) for 24 h | Compared to the control group, the protein-degradation-induced EGCG group showed: ↓ PIF- or TNF-α-induced protein degradation ↔ Protein synthesis rates |
| Wang, 2016, [93] | Model: C2C12 skeletal muscle cells Treatment: Control (no EGCG) or EGCG (0, 5, 25, and 50 μM) with or without H2O2 (300μM) for 48 h | Compared to the control group, the EGCG group (at 25 μM) showed: ↓ SDH ↑ CAT and GSH-Px activity ↓ PGC-1α and NRF-1 protein content ↓ Intracellular ROS levels Compared to the EGCG group, the EGCG and H2O2 groups showed: ↑ p-AMPKα/AMPKα ↑ MyHC 1 |
| Animal Studies | ||
| Alway, 2014, [143] | Model: Brown Norway inbred aged (34 months) rats Treatment: Fed a control diet (no GTE) or GTE (50 mg/kg) for 7 days prior and throughout. Rats either received 14 days hindlimb suspension or 14 days hindlimb suspension and 14 days ambulation | Compared to the control group, the GTE with ambulation group showed: ↑ Activation of myogenic progenitor cells ↑ Muscle fiber area ↓ Bax and FADD |
| Alway, 2015, [140] | Model: Fischer 344 brown Norway aged (34 months) rats Treatment: Fed control (no GTE) or GTE (50 mg/kg) for 7 days prior and throughout. Rats either received 14 days hindlimb suspension or 14 days hindlimb suspension and 14 days ambulation | Compared to the control group or GTE with suspension group, the GTE with suspension and ambulation group showed: ↓ Suspension-induced muscle mass loss, cross-sectional area, and tetanic force ↔ Muscle mass and force during ambulation ↑ Number of nuclei adjacent to basal lamina (SC proliferation) ↑ Number of nuclei inside sarcolemma (SC differentiation) ↓ Protein carbonyl levels |
| Bhattacharya, 2015, [142] | Model: Young adult male BALB/c mice Treatment: Fed a control diet (no EGCG) or EGCG (250 mg/kg/d) for 39 days and given access to a running wheel | Compared to the control group, the EGCG group showed: ↑ Lean body mass (trend) |
| Buetler, 2002, [99] | Model: Mdx mouse treated with or without tert-butylhydroperoxide Treatment: Fed standard diet supplemented with or without 0.01% and 0.05% (by wt) GTE for 4 weeks | Compared to the control group, the GTE group showed: ↓ Necrosis in the fast-twitch muscle elongator digitorum longus ↓ Oxidative stress induced by tert-butylhydroperoxide treatment |
| Call, 2008, [132] | Model: C57BL/6J mice treated with a high-fat diet Treatment: Fed a control diet (no GTE) or GTE (0.5%, 45% EGCG) and endurance exercised (voluntary wheel running) for 3 weeks | Compared to the control group, the GTE group showed: ↑ Total distance running (128%) ↑ Citrate synthase activity ↓ Serum creatine kinase |
| Cao, 2007, [110] | Model: Male Wistar Rat Treatment: Rats were fed a high-fructose diet (induced insulin resistance and oxidative stress) and green tea solid extract (EGCG 12.75%) (1 or 2 g/kg) diet for 6 weeks | Compared to the control (high-fat alone), the GTE group showed: ↑ mRNA expression of anti-inflammatory tristetraproline family in liver and muscle ↓ mRNA expression of proinflammatory genes in liver and muscle |
| Chen, 2020, [145] | Model: Male BALB/c mice Treatment: Fed a control diet (no GTE) or GTE (0.2 g/kg) and/or endurance exercised (treadmill running) for 8 weeks Model: C2C12 skeletal muscle cells Treatment: Pretreated with or without (control group) GTE (0.01%) and then exposed to ammonium chloride (5 mM) | Compared to the control group, the GTE with exercise group showed: ↑ Endurance capacity and urea-cycle-related gene expression Compared to the control group, the GTE group showed: ↓ Hyperammonemia-induced reduced mitochondrial respiration |
| Dorchies, 2006, [98] | Model: Mdx mice Treatment: Fed a standard diet with or without 0.25% GTE (EGCG, 0.1%) for 5 weeks | Compared to the control group, the GTE group showed: ↑ Phasic and tetanic tensions almost to matched control values ↑ Residual force by 30–50% |
| Evans, 2010, [114] | Model: Mdx mice Treatment: Fed a control diet (no GTE), 0.25%, or 0.5% GTE (EGCG >45%) for 21 days | Compared to the control group, the GTE group showed: ↓ Serum creatine kinase ↑ Area of normal fiber morphology ↓ Area of regenerating fibers ↓ NF-κB staining in regenerating muscle fibers |
| Haramizu, 2013, [112] | Model: Male ICR mice Treatment: Fed a control diet (no GTE) or GTE (0.5%) for 3 weeks. Mice were then downhill running exercised (induce muscle damage) | Compared to the control group, the GTE group showed: ↓ Muscle-damage-induced reduction in voluntary wheel-running activity, tetanic force ↓ Muscle-damage-induced reduction in plasma creatine phosphokinase levels ↓ TNF-α, IL-1β, MCP-1 |
| Hong, 2020, [104] | Model: Male ICR aged mice (24 months) Treatment: Fed a control diet, epicatechin (2 mg/kg), or tannase-converted GTE (20 or 40 mg/kg) diet | Compared to the control group, the tannase-converted GTE group or epicatechin group showed: ↑ Lean mass at 40 mg/kg for the tannase-converted GTE group ↑ MyoD, myogenin for both groups ↓ Myostatin for both groups ↑ S6K, follistatin at 40 mg/kg for the tannase-converted GTE group ↑ SOD, CAT for both groups ↓ FoxO3a, ↑ mTOR at 40 mg/kg for the tannase-converted GTE group ↓ MuRF-1 and atrogin-1 for the tannase-converted GTE group |
| Kumaran, 2008, [100] | Model: Male albino Wistar aged (34 months) rats Treatment: Fed a control diet (no EGCG) or EGCG (100 mg/kg/d) for 30 days | Compared to the control group, the EGCG group showed: ↓ Lipid peroxidation and protein carbonyl content |
| Meador, 2015, [127] | Model: Sprague–Dawley aged (20 months) rats Treatment: Fed a control diet (no EGCG) or EGCG (200 mg/kg) for 8 weeks | Compared to the control group, the EGCG group showed: ↑ Muscle mass, cross-sectional area ↑ IL-15, IGF-1 |
| Murase, 2005, [130] | Model: BALB/c mice Treatment: Fed a control diet (no GTE) or GTE (0.2–0.5%) for 10 weeks and then endurance exercised (swimming until exhaustion) | Compared to the control group, the GTE group showed: ↑ Swimming times to exhaustion (8–24%) ↑ β-oxidation activity, fat oxidation, and plasma free fatty acid concentration ↓ Respiratory quotient and plasma lactate concentration ↑ Fatty acid translocase/CD36 mRNA expression |
| Murase, 2006, [131] | Model: BALB/c mice Treatment: Exercised with or without GTE (0.2–0.5%) for 10 weeks and then endurance exercised (treadmill running until exhaustion) | Compared to the control (exercise only) group, the GTE group showed: ↑ Running times to exhaustion 30% ↓ Respiratory exchange ratio, malonyl-CoA content, and plasma lactate concentration ↑ β-oxidation activity, muscle glycogen content, and plasma free fatty acid concentration |
| Murase, 2008, [129] | Model: Senescence-accelerated prone mice Treatment: Fed a control diet (no GTE) or GTE (0.35%, EGCG 41%) for 10 weeks and/or endurance exercised (treadmill running) | Compared to the control group, the GTE group showed: Maintained endurance capacity ↓ Oxidative stress Compared to the control group, the GTE with the exercise group showed: ↑ Oxygen consumption, fatty acid β-oxidation, mitochondria-related mRNA expression |
| Nakae, 2008, [97] | Model: Mdx mice Treatment: Fed vehicle (control group) or EGCG (180 mg/kg/d) for 5 weeks | Compared to the control group, the EGCG group showed: ↓ Development of dystrophic muscle lesions |
| Onishi, 2018, [141] | Model: Senescence-accelerated mouse prone-8 Treatment: Fed a control diet, an HFD diet, or HFD with 0.5% GTEs (71.68% EGCG) (GTE group) diet for 4 months | Compared to the HFD group, the GTE group showed: ↑ Muscle mass ↑ Akt and S6K phosphorylation |
| Ota, 2011, [101] | Model: BALB/c mice Treatment: Fed a control diet (no GTE) or a diet containing 0.5% GTE (81% catechins) for 14 days, and then mice were subjected to continuous tail suspension for 10 days | Compared to the control group, the GTE group showed: ↓ Unloading induced muscle tetanic force loss ↑ Total antioxidant potential ↓ Carbonylated protein levels |
| Sae-Tan, 2014, [128] | Model: C57bl/6J mice treated with a high-fat diet Treatment: Fed control (high-fat) diet or 0.32% EGCG for 16 weeks | Compared to the GTE without exercise group, the GTE with exercise group showed: ↓ Body mass and visceral fat ↑ PGC-1α, Cytb, CO III (mitochondrial metabolism enzymes) |
| Serrano, 2013, [124] | Model: Male C57BL6 mice Treatment: Fed a control diet or tea beverage (EGCG 37.8%) for 3 months | Compared to the control group, the EGCG group showed: ↓ Oxidative damage ↑ AMP-activated protein kinase α levels ↓ UCP-2 and UCP-4 ↑ Porin ↓ Mitochondrial DNA to nuclear DNA ratio |
| Shen, 2012, [102] | Model: Female Sprague–Dawley rats Treatment: Fed a high-fat diet for 4 months and then fed with or without GTE (0.5%) for additional 4 months | Compared to the control (high-fat without GTE) group, the GTE group showed: ↑ Glutathione peroxidase (reduces ROS) |
| Takahashi, 2017, [45] | Model: Brown Norway inbred aged (34 months) rats Treatment: Fed a control diet (no EGCG) or EGCG (50 mg/kg). Rats either received 14 days hindlimb suspension or 14 days hindlimb suspension and 14 days ambulation | Compared to the control group, the EGCG without ambulation group showed: ↑ ATG16L2, SNCA, TM9SF1, Pink1, PIM-2 gene expression ↑ ATG12, ↓ Beclin-1 protein content Compared to the control group, the EGCG group with ambulation showed: ↓ Beclin1 and LC3-II/I protein content |
| Wang, 2015, [103] | Model: Male Kunming mice Treatment: Intraperitoneally injected daily vehicle (control group) or EGCG at the dose of 55, 75, 100, and 200 mg/kg for 5 consecutive days | Compared to the control group, the EGCG group showed: ↓ SOD, catalase, glutathione peroxidase at 75 mg/kg ↔ Endogenous antioxidant activity Hepatotoxicity triggered at 200 mg/kg |
| Wang (2011), [113] | Model: Male C57BL/6 mice Treatment: Mice were injected with LLC cells (induced LLC tumor) and then fed EGCG (0.2 mg/mouse/d, prevention or 0.6, treatment mg/mouse/d) for 12 days | Compared to the control group, the EGCG group showed: ↓ Leukocyte infiltration ↓ NF-κB, and MuRF1, MAFbx (E3-ligases) ↓ Tumor volume and mass |
| Zhang (2020), [111] | Model: C57BL/6J mice Treatment:Mice were fed with HFD for 10 weeks to induce obesity. Obese mice were fed with continuous HFD, HFD with GTE (EGCG 12.5%), HFD with Ex, and HFD with both GTE and Ex for 8 weeks | Compared to the control group, the GTE + EX showed: ↓ blood glucose, serum total cholesterol, triglyceride, insulin, and alanine aminotransferase activity GTE, GTE + EX, EX ↓ proinflammatory gene expression GTE, GTE + EX ↑ IkBα (NF-κB inhibitor) activity GTE + EX ↑ glucose transport genes mRNA |
| Human Studies | ||
| Hadi, 2017, [105] | Model: Men (n = 18; 20.94 ± 1.43 years) soccer players Treatment: Supplemented with or without (placebo group) GTE (450 mg/d) for 6 weeks Exercise: Maintained the same exercise schedule throughout the study | Compared to the placebo group, the GTE group showed: ↓ MDA levels |
| Jowko, 2015, [106] | Model: Male (n = 16; 21.6 ± 1.5 years) sprinters Treatment: Supplemented with or without (placebo group) GTE (980 mg/d) for 4 weeks Exercise: Repeated cycle sprint test | Compared to the placebo group, the GTE group showed: ↓ MDA and SOD levels ↑ Total antioxidant capacity |
| Panza, 2008, [107] | Model: Weight-trained men (n = 14; 19–30 years) Treatment: Supplement with water (placebo) or with GTE (200 mL) 3 times per day for 8 days. Exercise: Resistance exercise (Ex) protocol consisted of a warm-up followed by 4 sets of 10, 8, 6, and 4 repetitions, with 75%, 80%, 85%, and 90% of 1-RM, respectively | Compared to the placebo group, the GTE group showed: ↓ LH, aspartate aminotransferase, and uric acid ↑ GSH-Px ↓ Ex-induced increase in CK and xanthine oxidase |
| Silva, 2018, [109] | Model: Untrained men (n = 20; 23 ± 5 years) Treatment: Supplemented with or without (placebo group) GTE (500 mg/d) for 15 days and exercised (induced DOMS in triceps sural) before and after supplementation Exercise: Calf raising exercise trial one (maximal voluntary repetitions) and trial two (75% of maximal repetitions with repeated sets until >50% maximal of maximal repetitions could be achieved) | Compared to the placebo group, the GTE group showed: ↓ Creatine kinase |
| Townsend, 2018, [120] | Model: Untrained men (n = 38; 18–35 years) Treatment: Supplemented with, without (control group), or with placebo, aqueous proprietary polyphenol blend (40% catechins, 3–8% EGCG) for 28 days. Then subjects completed acute low body resistance exercise protocol (induced muscle damage) Exercise: Squat (6 sets of 10 repetitions) and leg press, leg extension (4 sets of 10 repetitions) exercises all at 70% 1-RM | Compared to the control group, the EGCG group showed: ↓ Muscle-damage-induced increase in Bcl-2, BAD |
| Tsai, 2017, [134] | Model: Healthy men (n = 8; 22 ± 1 years) with regular recreational physical activities 3/week Treatment: Supplemented with or without (control group) GTE (500 mg/d) for 8 weeks and then performed cycling exercise Exercise: 60 min cycling exercise at 75% VO2max after a 5 min warm-up exercise (50 W), 24 h postsupplement | Compared to the control group, the GTE group showed: ↑ Exercise-induced muscle GLUT4 protein content ↑ Fat oxidation energy reliance |
| Kuo 2015, [108] | Model: Forty untrained men (age: 20 ± 1 years) Treatment: Assigned to placebo (control), GTE, endurance training (EX), or endurance training with GTE (GTE + EX). GTE groups orally took GTE (250 mg/day) for 4 weeks Exercise: 75% oxygen uptake reserve for three 20 min sessions per week | Compared to the control group, the EX and GTE + EX groups showed: ↑ Exhaustive run time ↑ Maximal oxygen uptake ↓ Serum creatine kinase, MDA levels |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luk, H.-Y.; Appell, C.; Chyu, M.-C.; Chen, C.-H.; Wang, C.-Y.; Yang, R.-S.; Shen, C.-L. Impacts of Green Tea on Joint and Skeletal Muscle Health: Prospects of Translational Nutrition. Antioxidants 2020, 9, 1050. https://doi.org/10.3390/antiox9111050
Luk H-Y, Appell C, Chyu M-C, Chen C-H, Wang C-Y, Yang R-S, Shen C-L. Impacts of Green Tea on Joint and Skeletal Muscle Health: Prospects of Translational Nutrition. Antioxidants. 2020; 9(11):1050. https://doi.org/10.3390/antiox9111050
Chicago/Turabian StyleLuk, Hui-Ying, Casey Appell, Ming-Chien Chyu, Chung-Hwan Chen, Chien-Yuan Wang, Rong-Sen Yang, and Chwan-Li Shen. 2020. "Impacts of Green Tea on Joint and Skeletal Muscle Health: Prospects of Translational Nutrition" Antioxidants 9, no. 11: 1050. https://doi.org/10.3390/antiox9111050
APA StyleLuk, H.-Y., Appell, C., Chyu, M.-C., Chen, C.-H., Wang, C.-Y., Yang, R.-S., & Shen, C.-L. (2020). Impacts of Green Tea on Joint and Skeletal Muscle Health: Prospects of Translational Nutrition. Antioxidants, 9(11), 1050. https://doi.org/10.3390/antiox9111050