

Forskolin, an Adenylcyclase/cAMP/CREB Signaling Activator Restoring Myelin-Associated Oligodendrocyte Destruction in Experimental Ethidium Bromide Model of Multiple Sclerosis


 , ,
, ,  , and
, and
Abstract
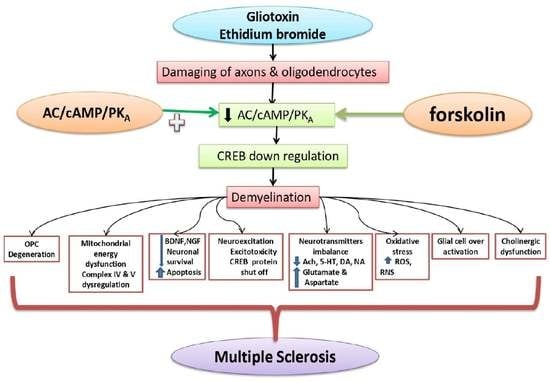
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
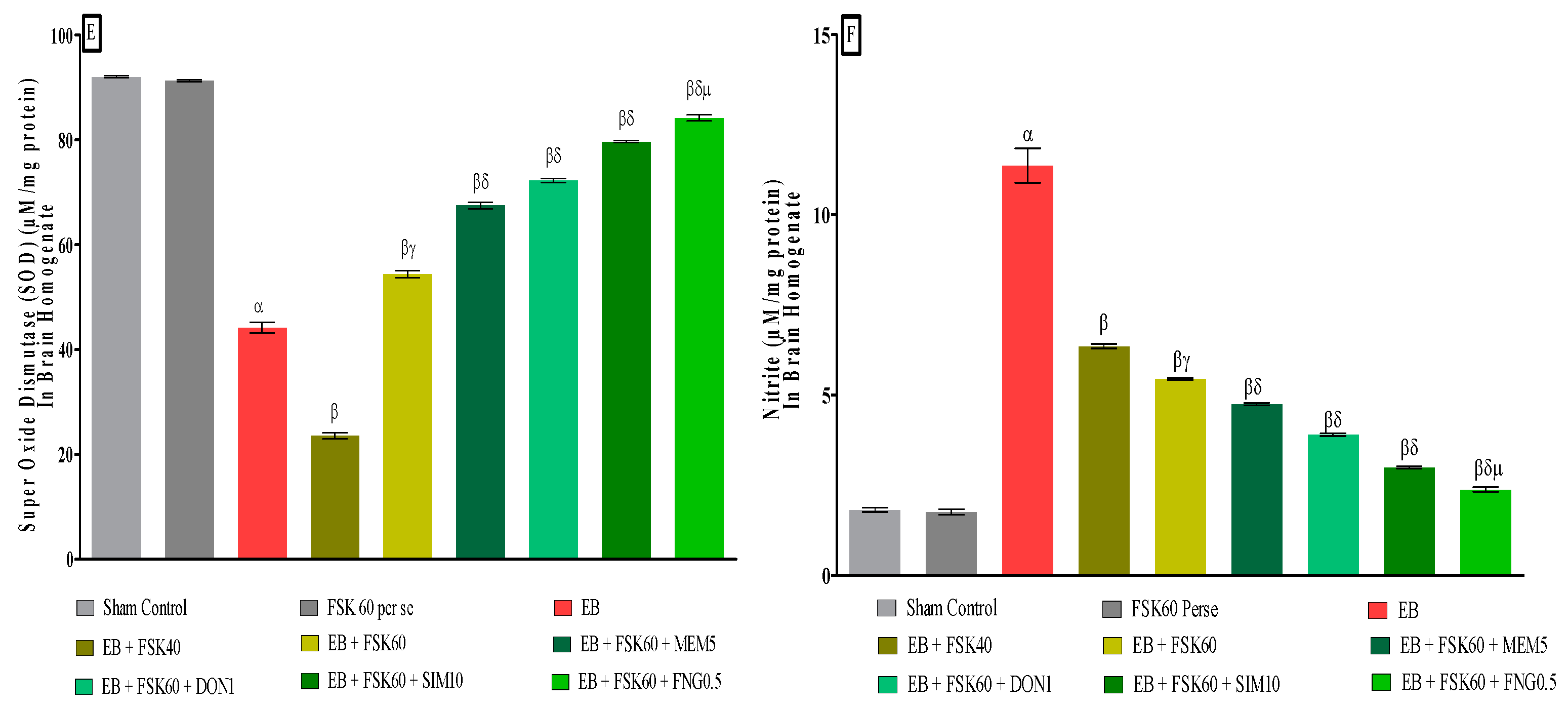{kind=link}
{kind=link}
1. Introduction
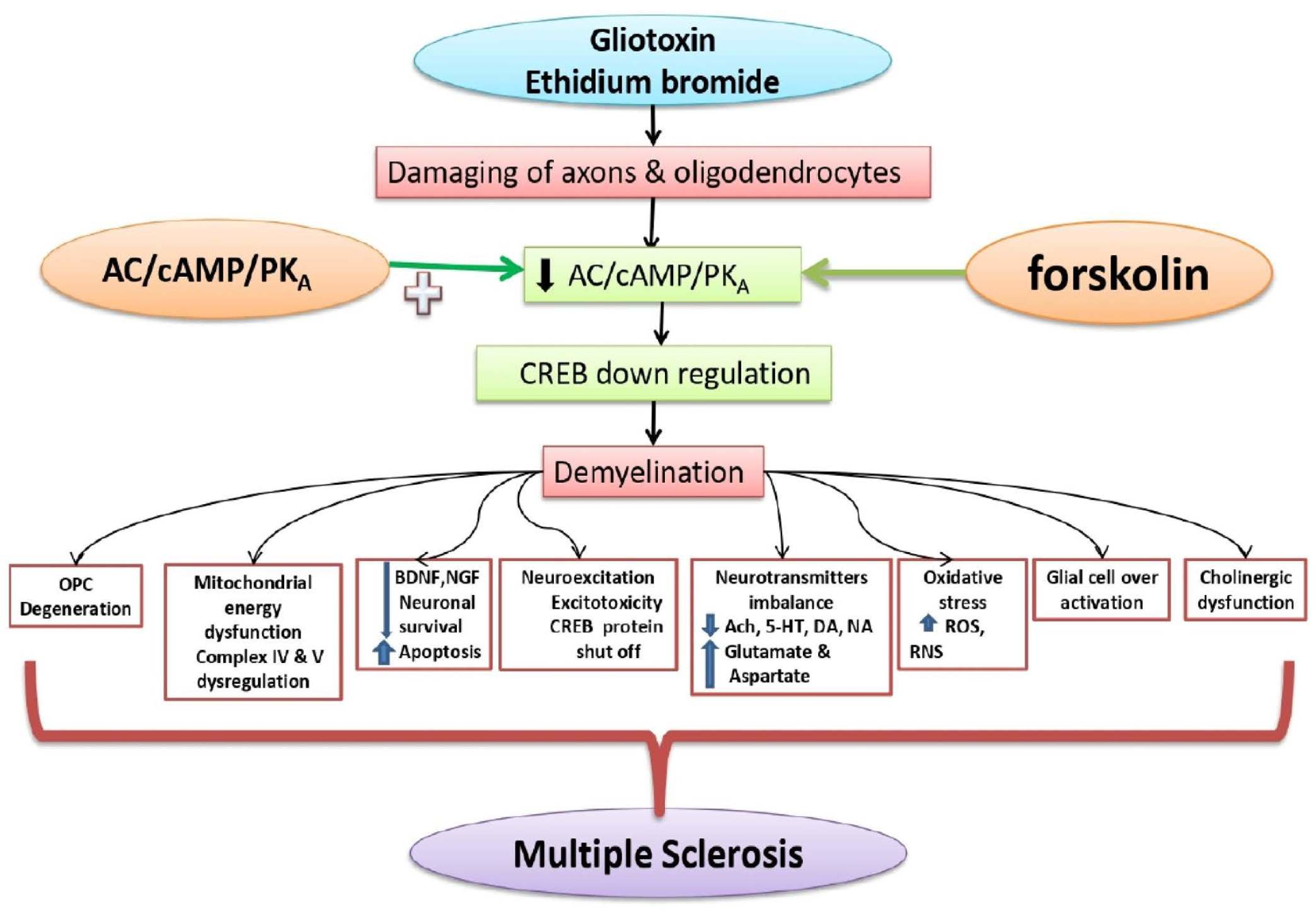
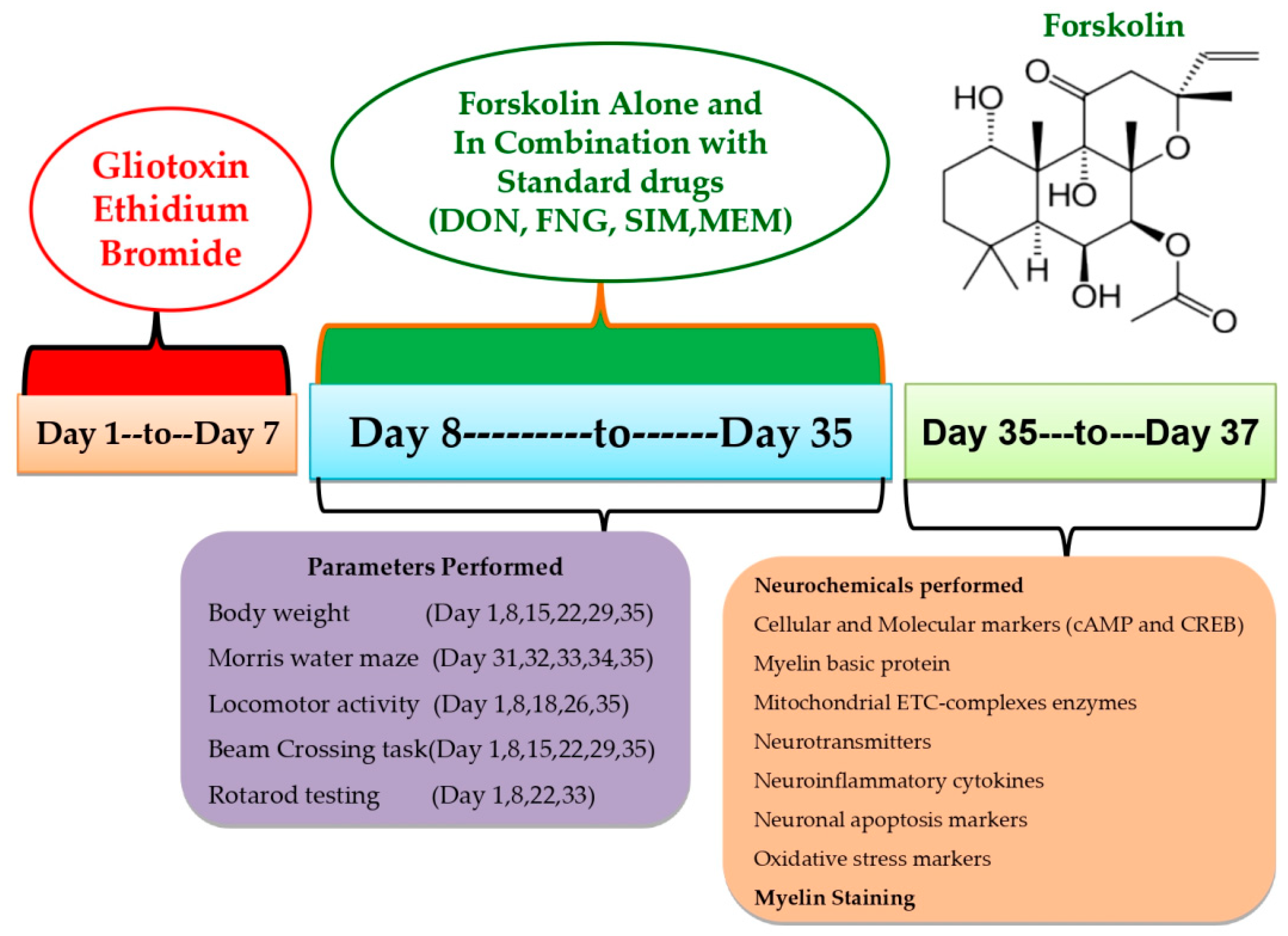
2. Materials and Methods
2.1. Experimental Animals
2.2. Drugs and Chemicals
2.3. Experimental Grouping of Animals
2.4. Demyelinated Multiple Sclerosis Was Induced in Rats with ICP-EB
2.5. Body Weight Measurement
2.6. Behavioral Parameters
2.6.1. Morris Water Maze (MWM) Task
2.6.2. Locomotor Activity (LA)
2.6.3. Beam Crossing Task (BCT)
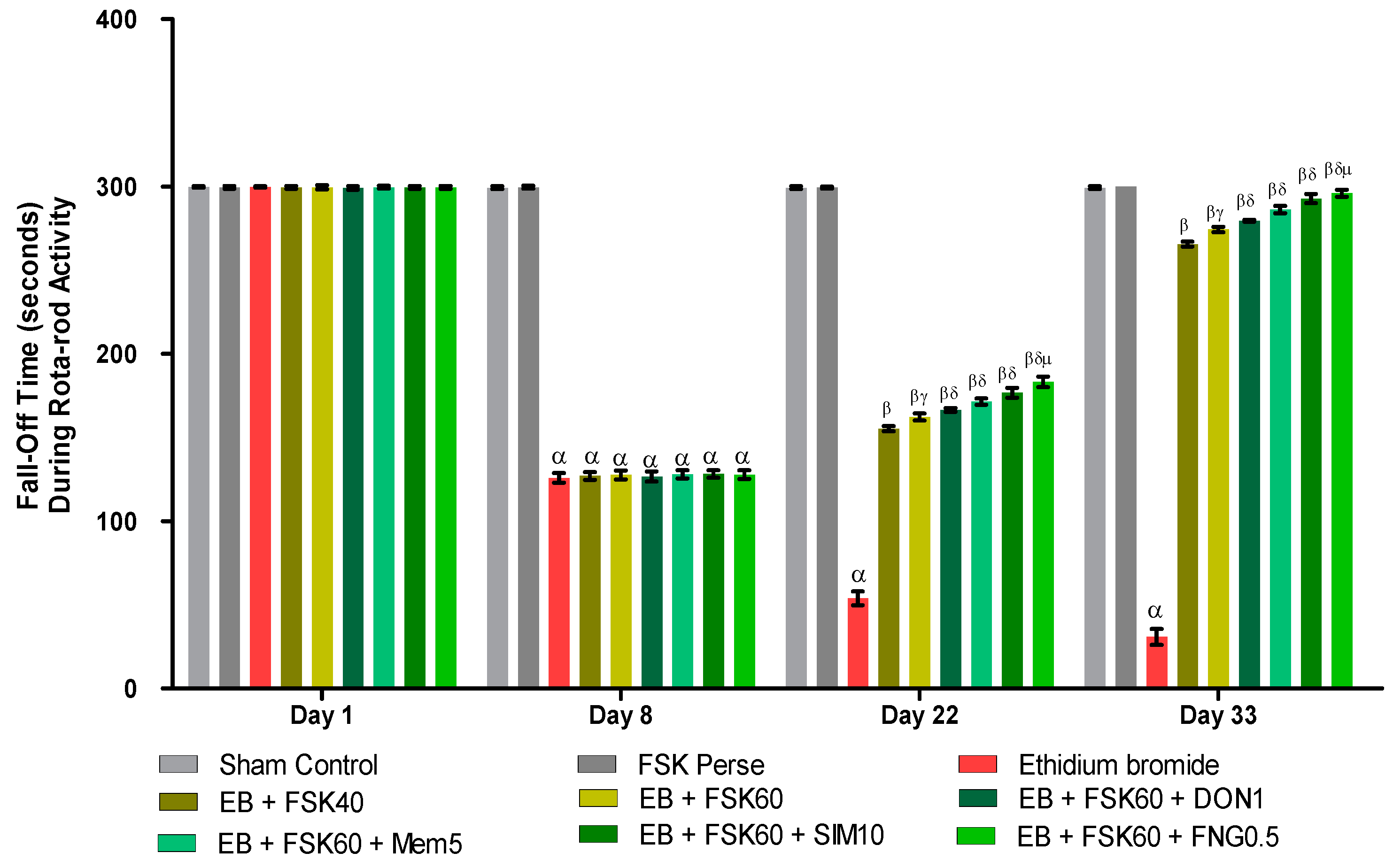
2.6.4. Rotarod Test
2.7. Biological Samples Preparation
Homogenized Brain Preparation
2.8. Analysis of Cellular Markers
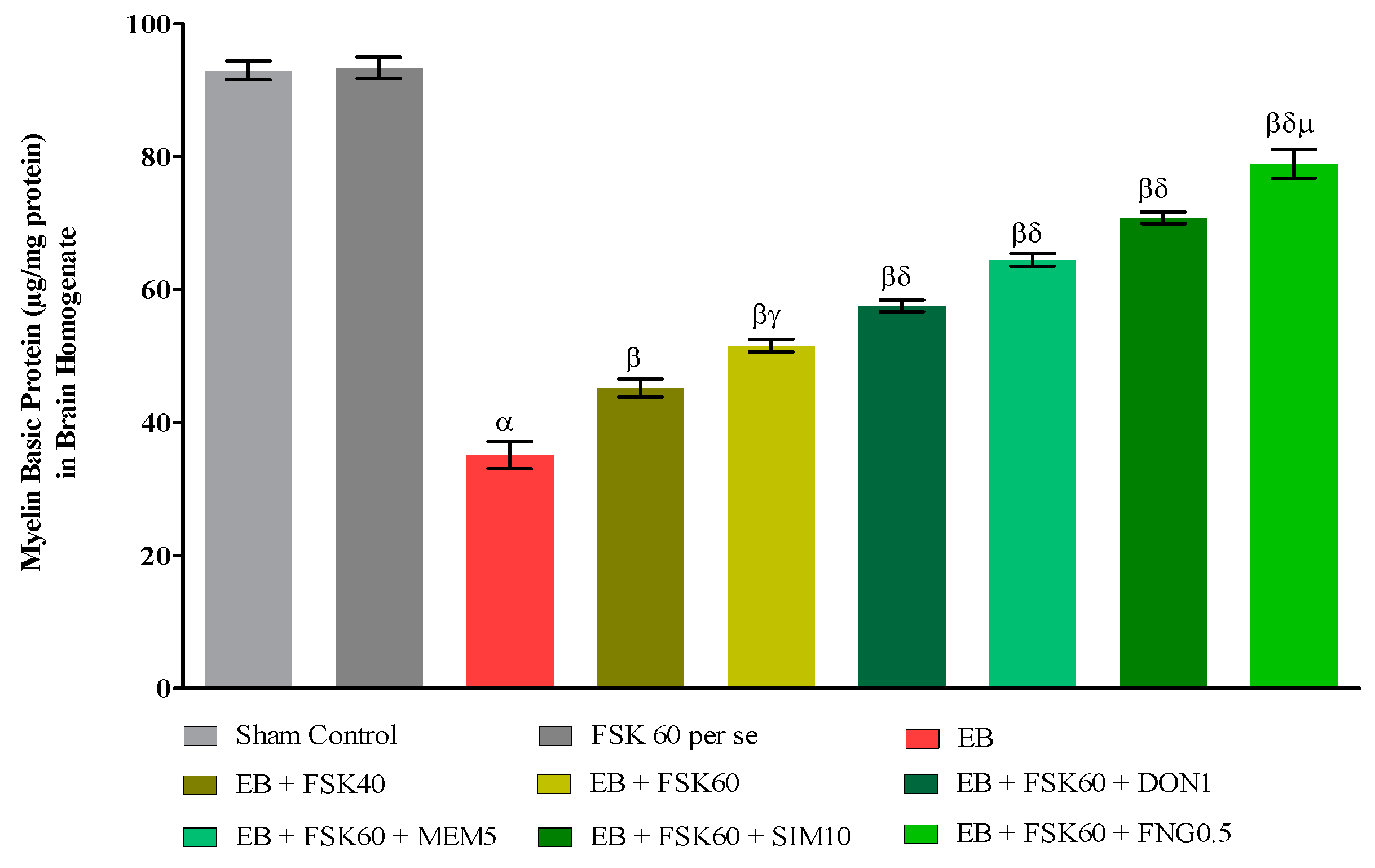
Myelin Basic Protein (MBP) Concentration in Brain
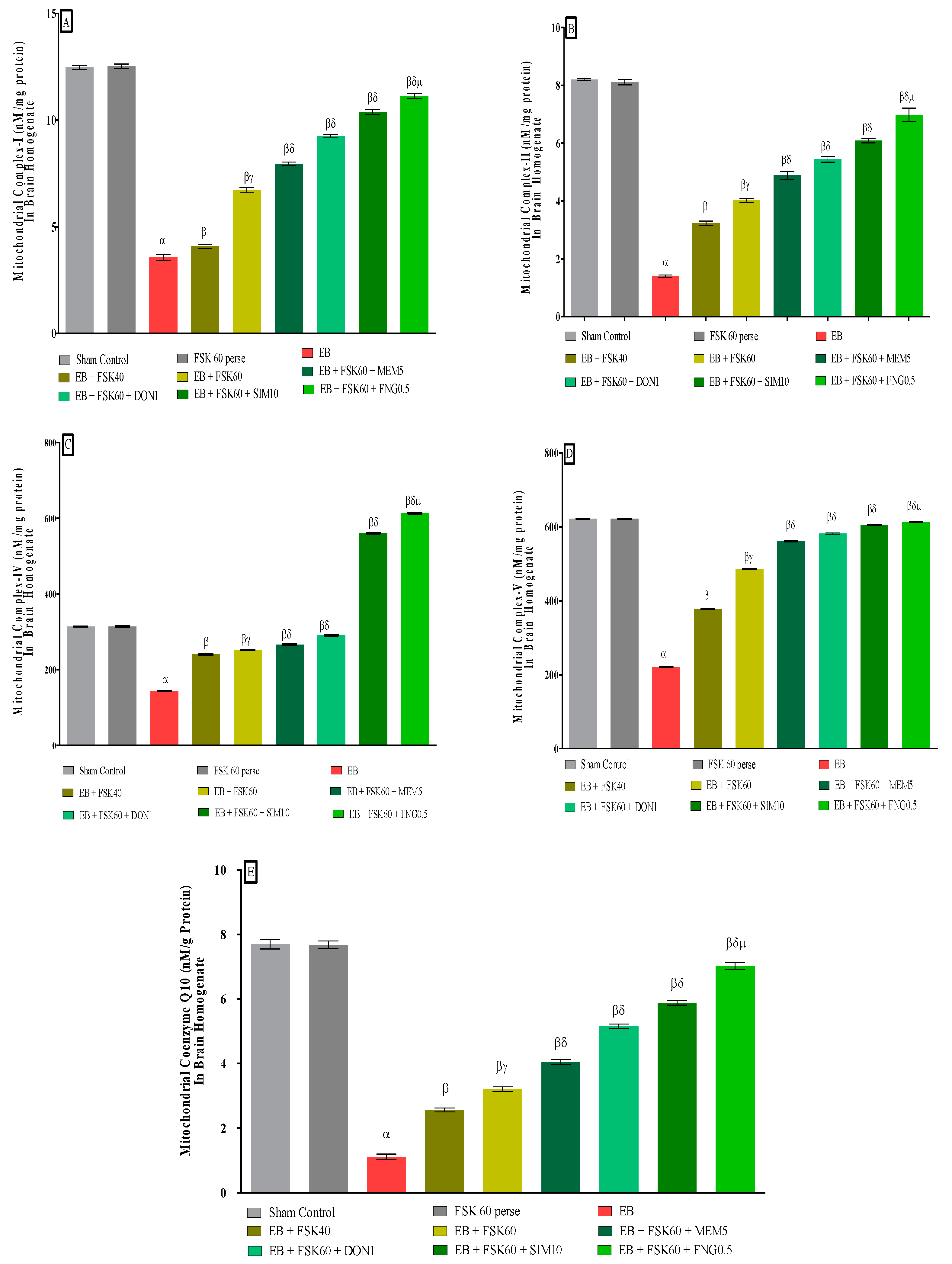
2.9. Analysis of Mitochondrial ETC Complexes Enzyme Concentration in Brain
2.9.1. A Post-Mitochondrial Supernatant (PMS) Prepared from Homogenate of Rat’s Whole Brain
2.9.2. Analysis of Mitochondrial ETC Complex-I (NADPH Dehydrogenase) Protein Concentration in Brain
2.9.3. Analysis of Mitochondrial ETC Complex-II (Succinate Dehydrogenase/SDH) Protein Concentration in Brain
2.9.4. Analysis of Mitochondrial ETC Complex-IV (Cytochrome Oxidase) Protein Concentration in Brain
2.9.5. Analysis of Mitochondrial ETC Complex-V (ATP Synthase) Protein Concentration in Brain
2.9.6. Analysis of Mitochondrial CoQ10 Concentration in Brain
2.10. Analysis of Molecular Markers
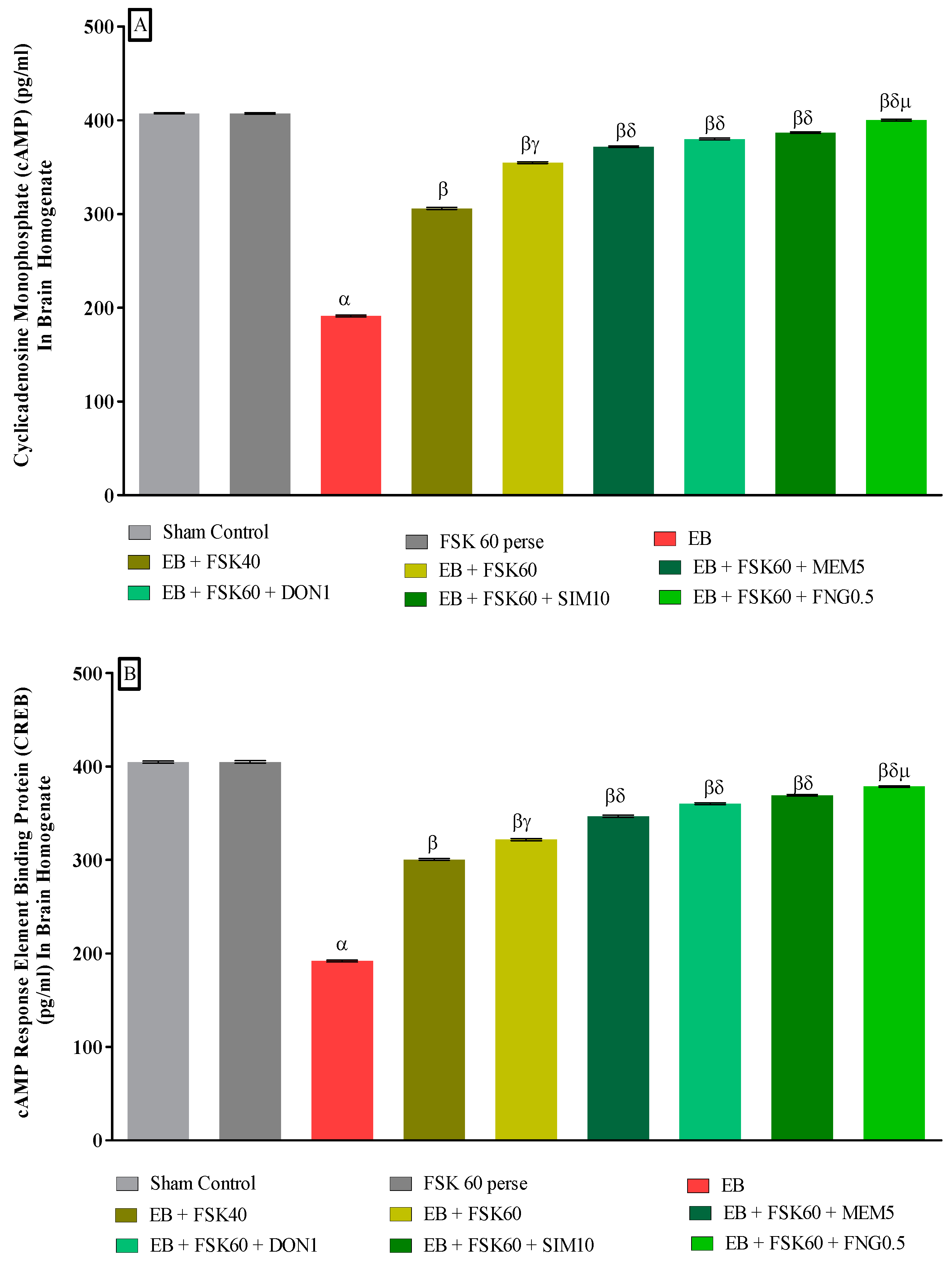
cAMP and CREB Protein Concentration in Brain
2.11. Analysis of Neuroinflammatory Biomarkers
TNF- α and IL-1β Protein Concentration in Brain
2.12. Analysis of Neurotransmitters
2.12.1. Dopamine Concentration in Brain
2.12.2. Glutamate Concentration in Brain
2.13. Analysis of Oxidative Stress Parameters
2.13.1. Protein Estimation
2.13.2. Lactate Dehydrogenase (LDH) Concentration in Brain
2.13.3. Acetylcholinesterase (AChE) Concentration in Brain
2.13.4. Glutathione (GSH) Concentration in Brain
2.13.5. Malondialdehyde (MDA) Concentration in Brain
2.13.6. Superoxide Dismutase (SOD) Concentration in Brain
2.13.7. Nitrite Concentration in Brain
2.14. Luxsol Fast Blue (LFB) Staining
2.15. Statistical Analysis
3. Results
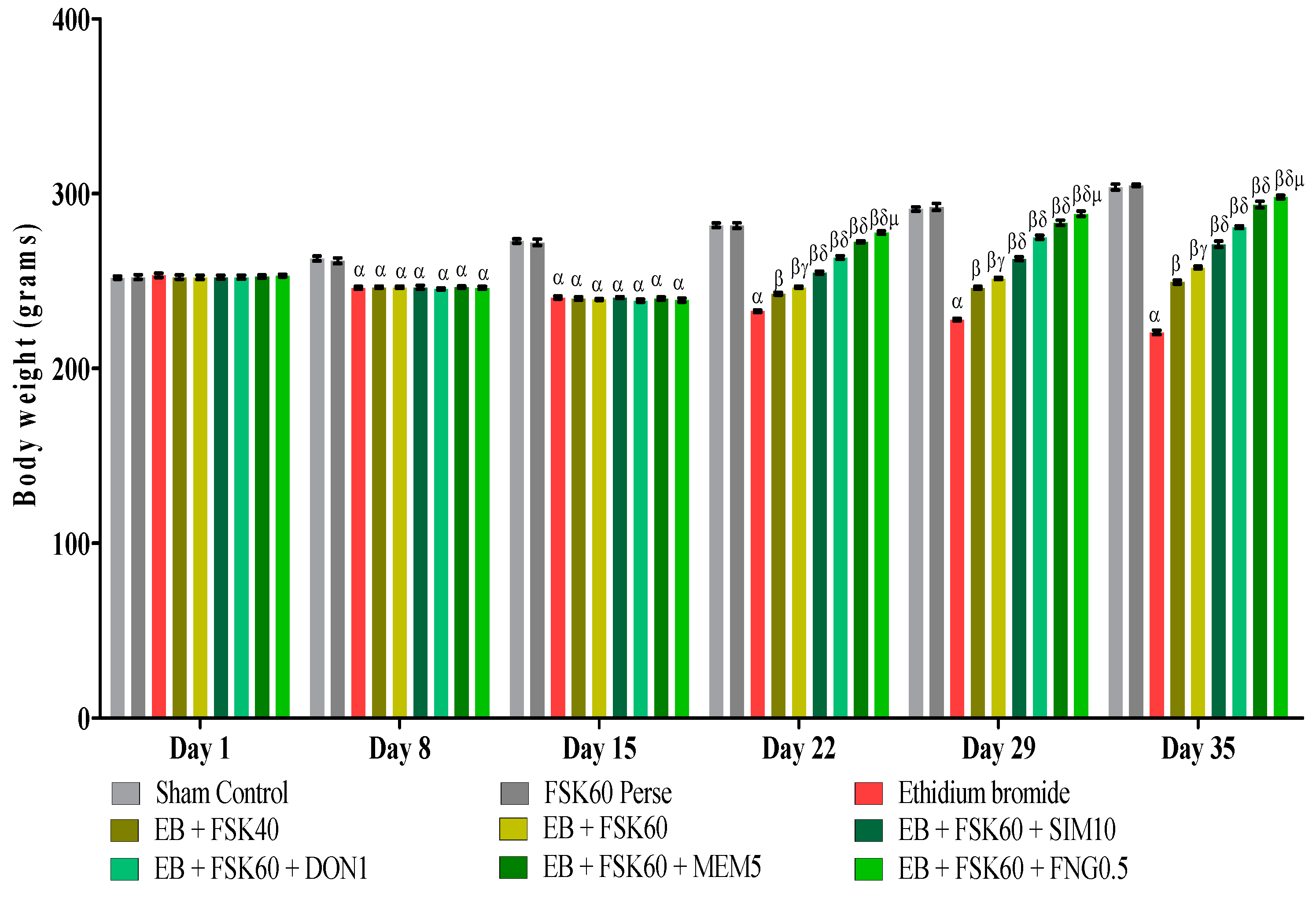
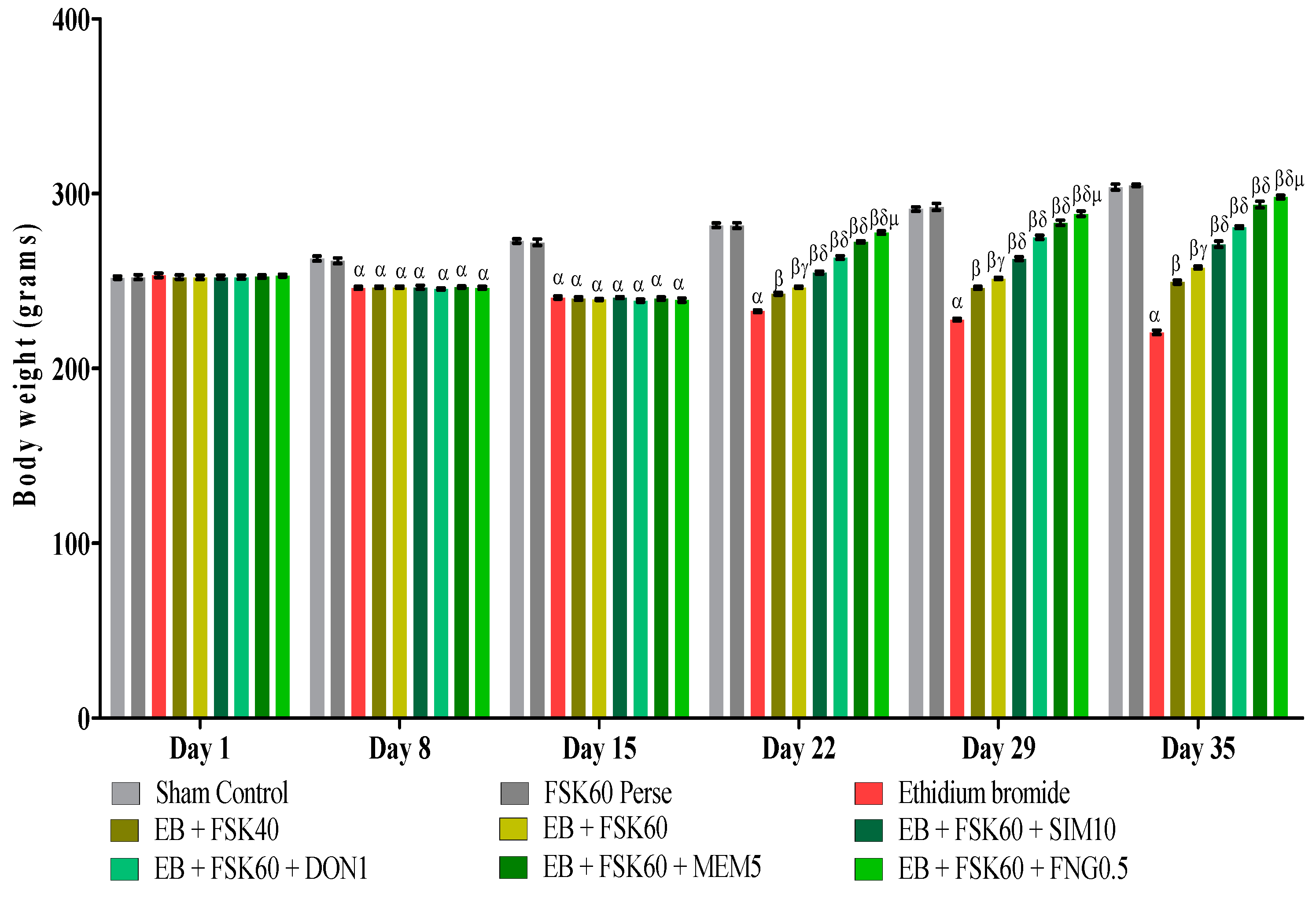
3.1. Protective Role of FSK on Body Weight in Experimental Multiple Sclerosis Rats
3.2. Behavioral Parameters
3.2.1. Protective Role of FSK on Morris Water Maze in Experimental Multiple Sclerosis Rats
Improvement in Spatial Learning and Memory
- a.
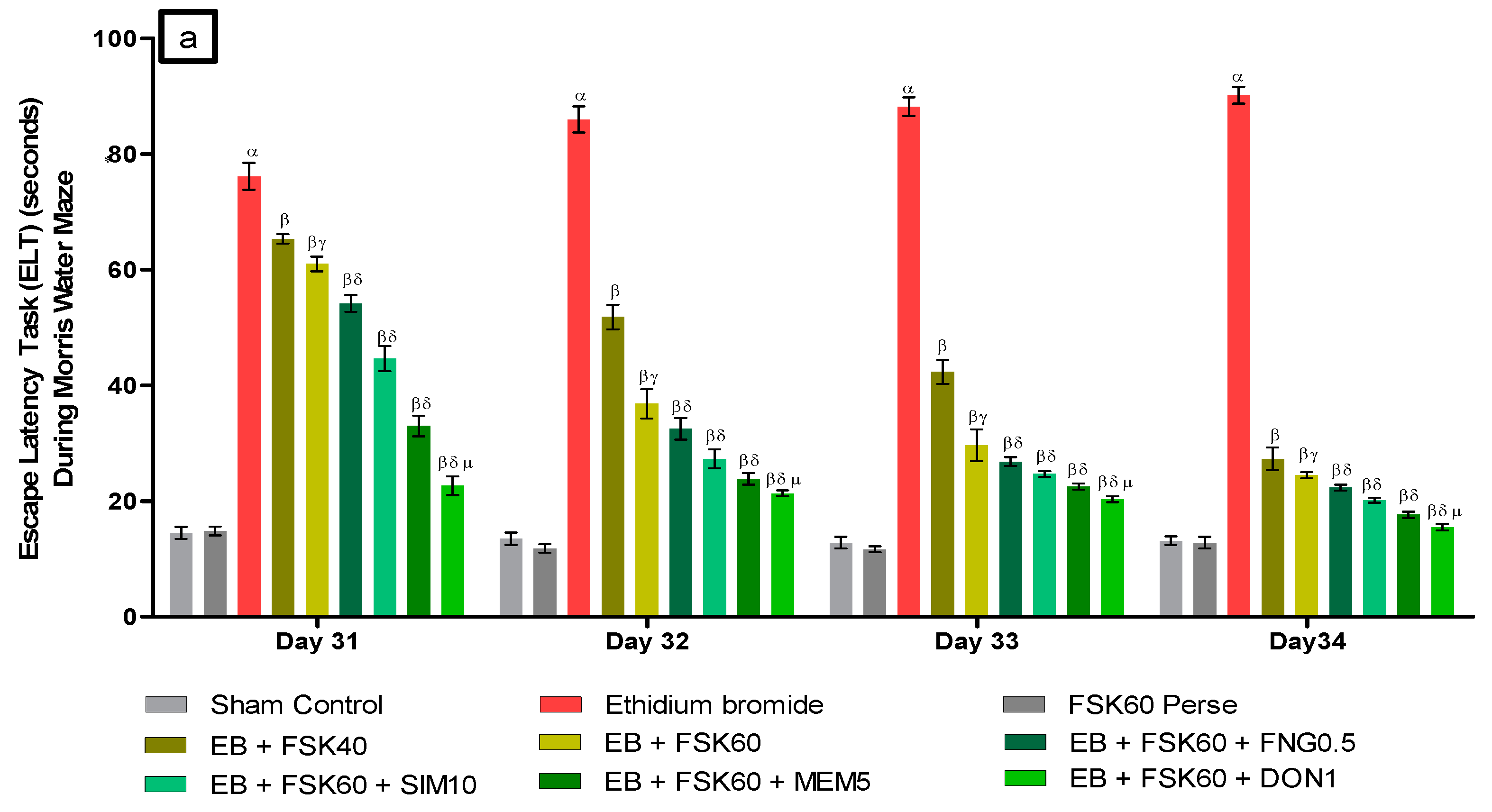
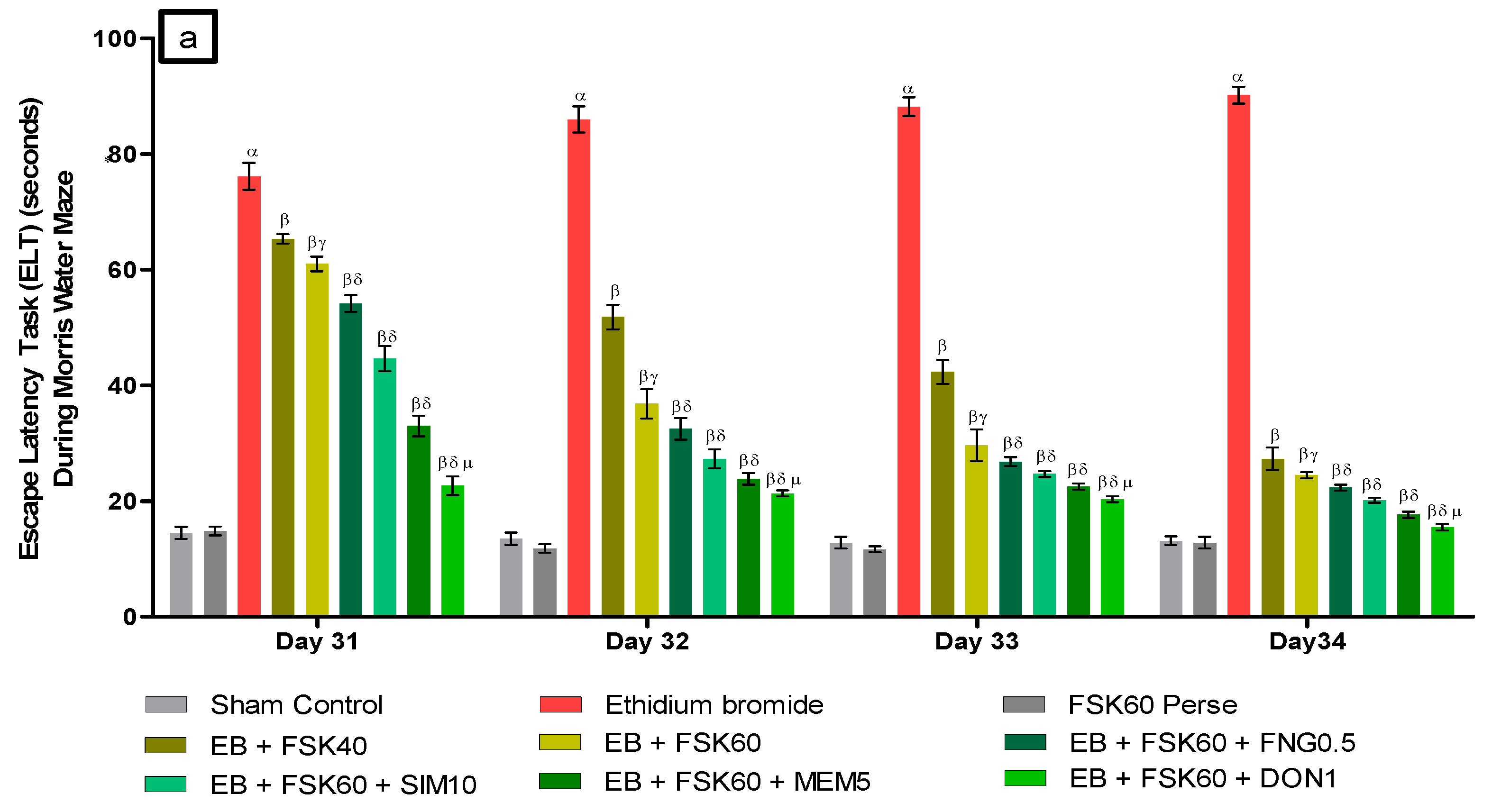
- Decrease in Escape Latency Task (ELT)ELT was performed on the 31st, 32nd, 33rd, and 34th days as per the protocol schedule of the Morris water maze (MWM). The ICP-EB (0.1 percent /10 μL)-unilaterally-treated group exhibited significantly increased ELT. Moreover, chronic administration of FSK40 and FSK60 alone or in combination with conventional medications, such as FNG0.5, SIM10, MEM5 and DON1, considerably decreased the time in ELT as compared to the ICP-EB-treated group in a dose-dependent manner [Two-Way ANOVA:F (24,135) = 195.0, p < 0.01]. However, no significant change was detected in the FSK60 per se treatment group compared to sham control. Among the selected doses of the drug, the high dose of FSK60 and DON1 were found to be more effective in restoring ICP-EB-induced increased ELT (Figure 3a).
- b.
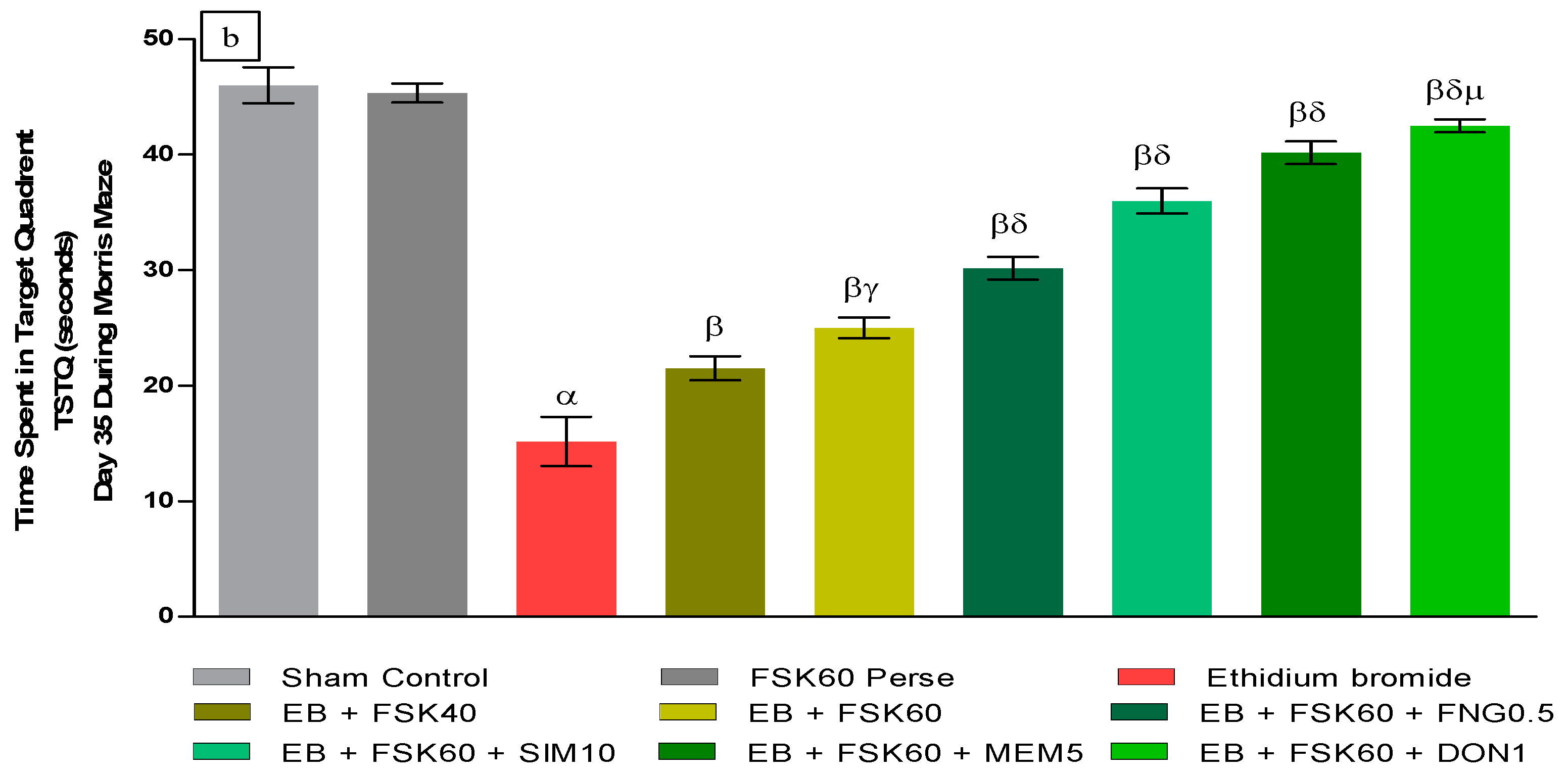
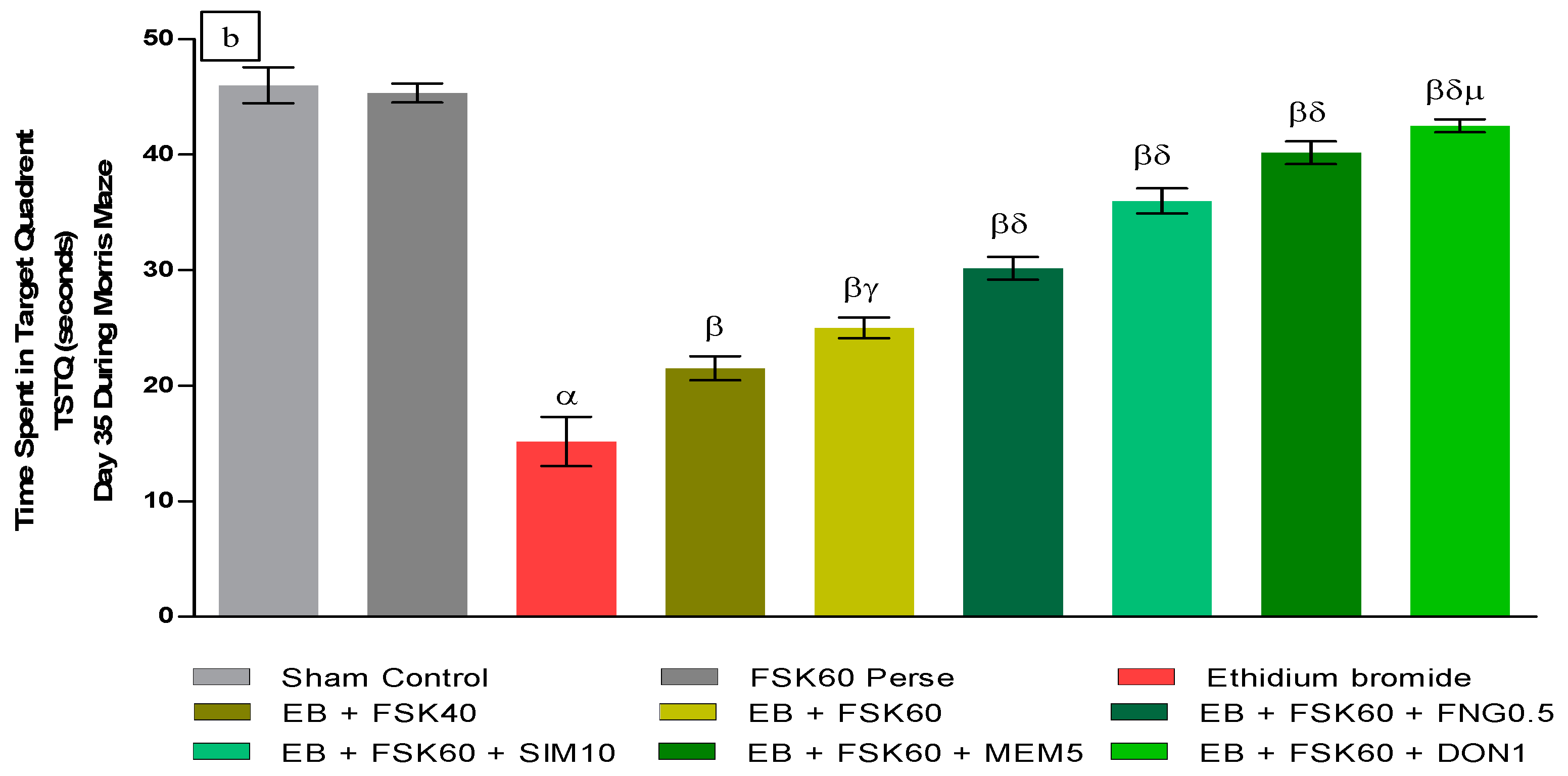
- Increase in Time Spent in Target Quadrant (TSTQ)TSTQ was performed on day 35 as per the protocol schedule on MWM. The ICP-EB (0.1 percent/10 μL)-unilaterally-treated group had a remarkably decreased TSTQ. Moreover, chronic administration of FSK40 and FSK60 alone or in combination with conventional medications, such as FNG0.5, SIM10, MEM5, and DON1, considerably increased TSTQ as compared to ICP-EB-treated groups in a dose-dependent manner [One-Way ANOVA:F (8,40) = 2.046, p < 0.01]. However, no significant change was detected in the FSK60 per se treatment group compared to sham control. Among the selected doses of the drug, the high doses of FSK60 and DON1 were found to be more effective in restoring ICP-EB-induced decreased TSTQ (Figure 3b).
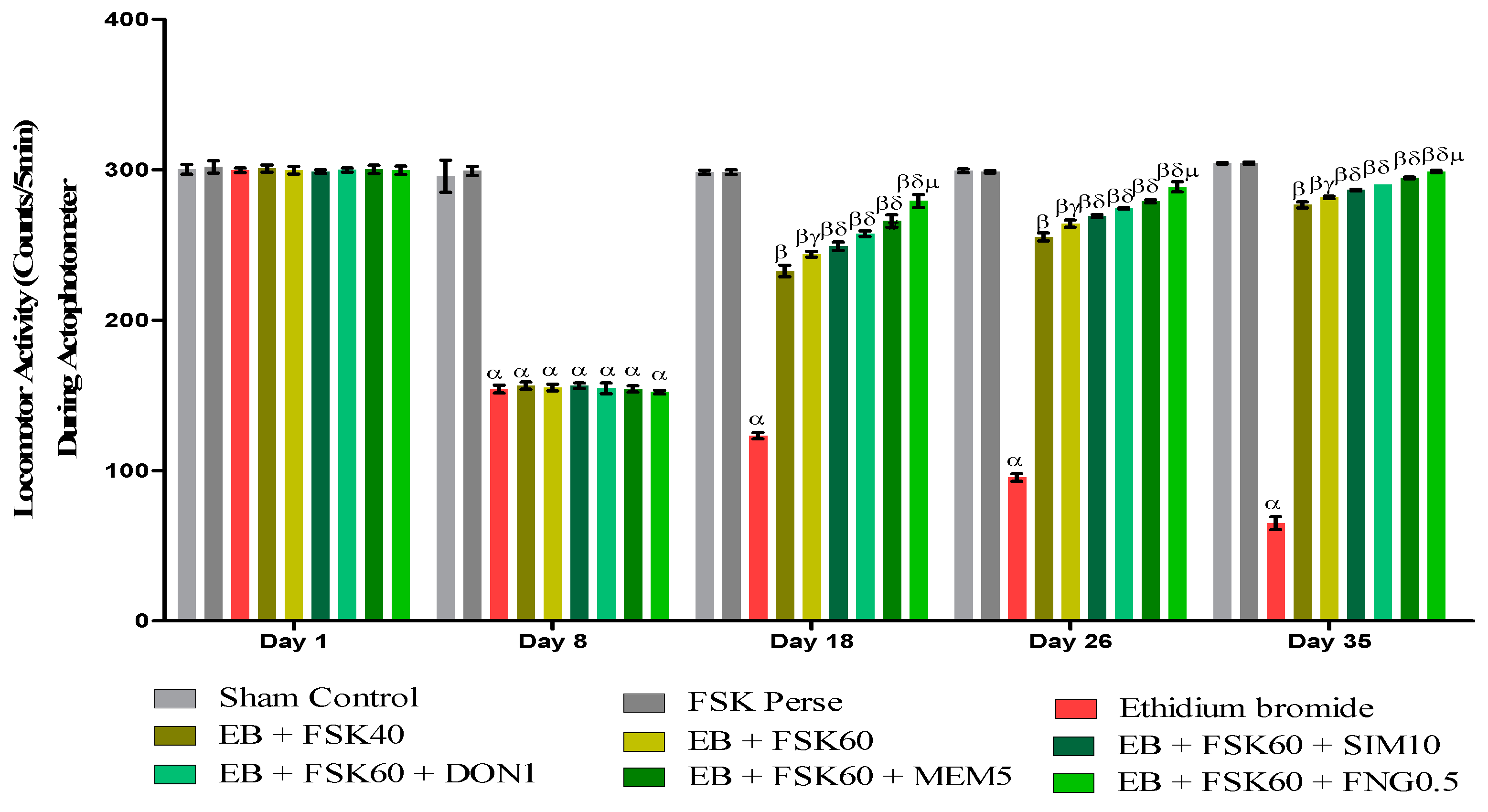
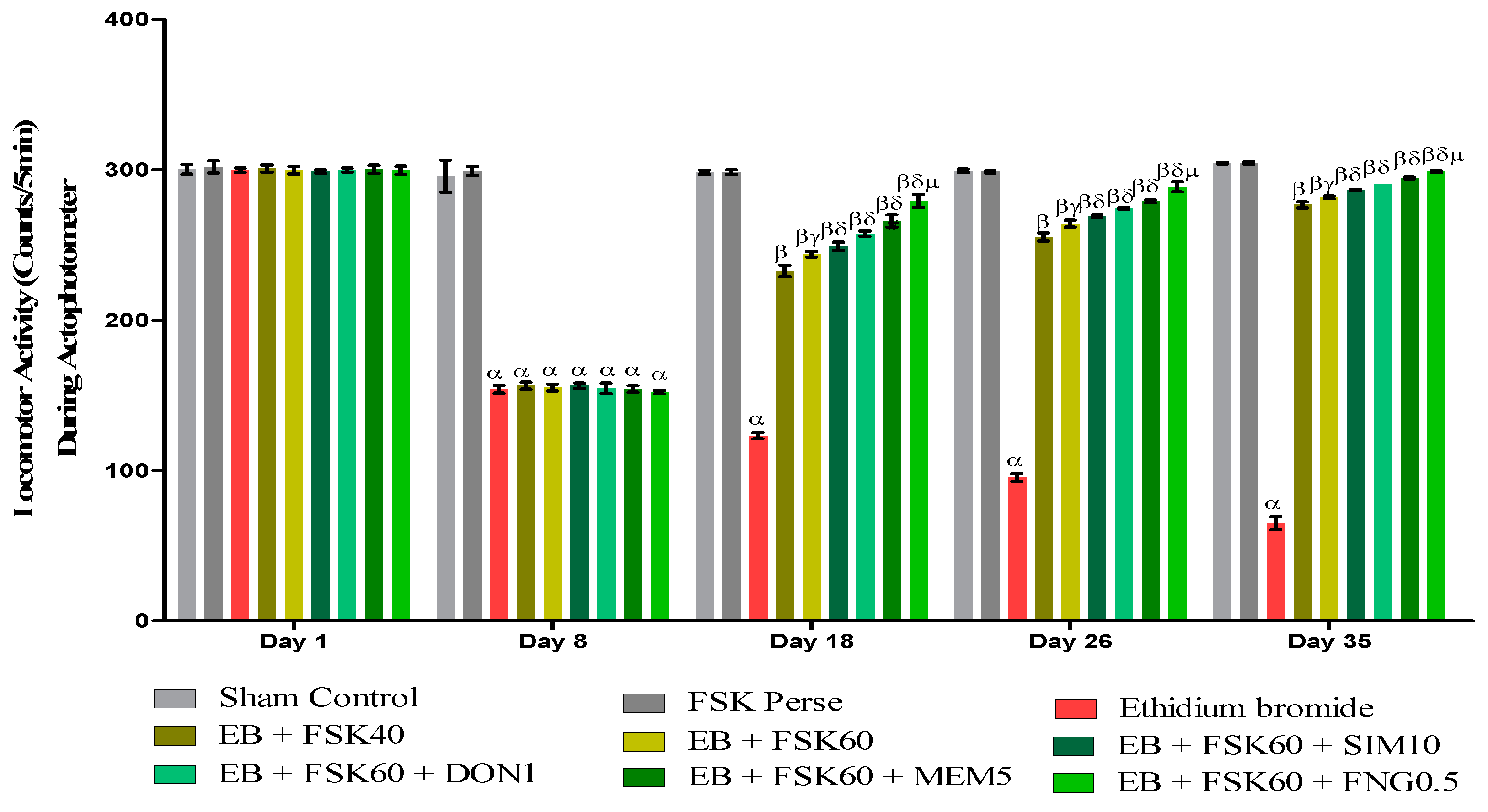
3.2.2. Protective Role of FSK on Locomotor Activity in Experimental Multiple Sclerosis Rats
Increase in Locomotion Movement
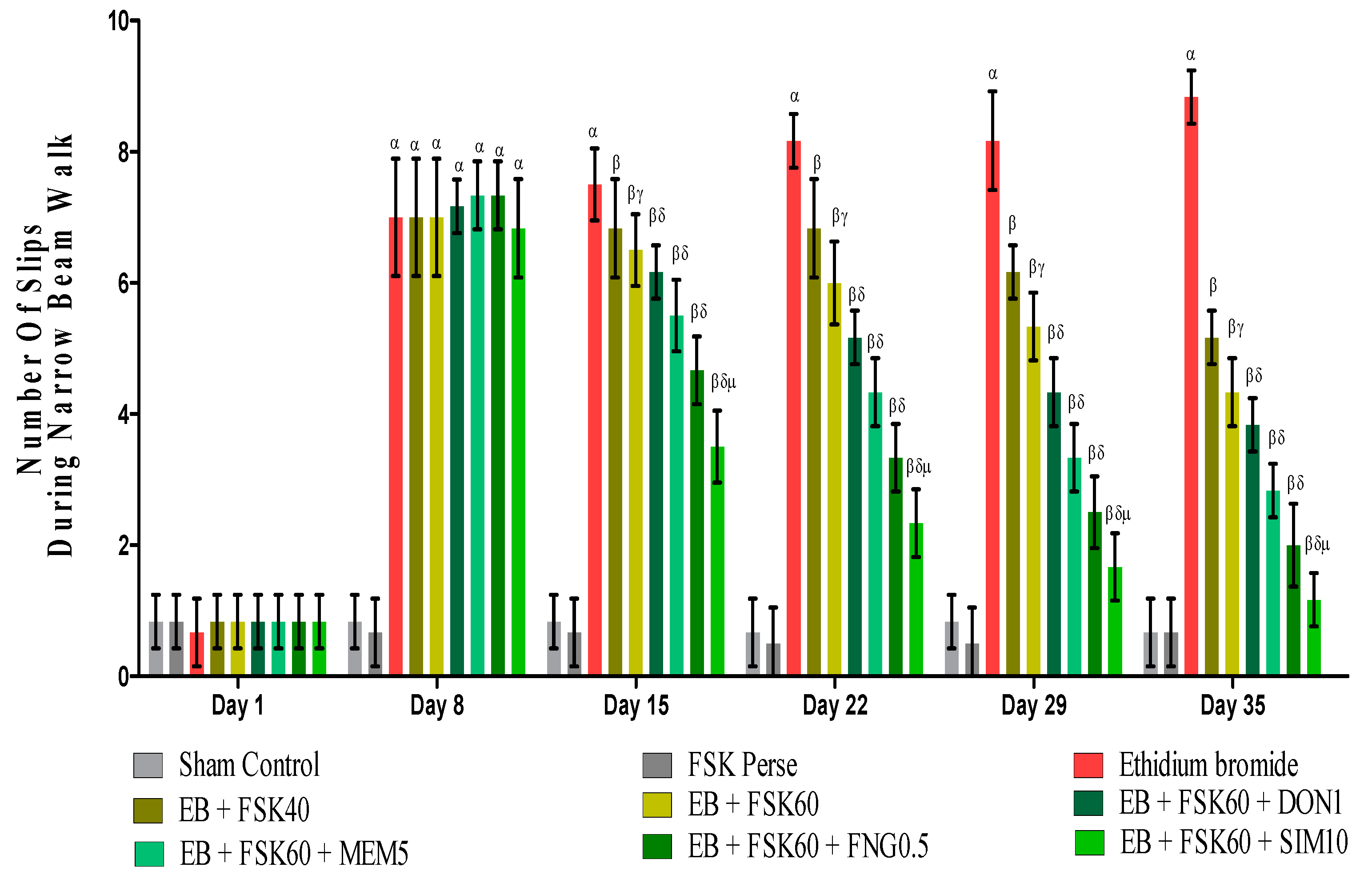
3.2.3. Protective Role of FSK on Beam Crossing Task in Experimental Multiple Sclerosis Rats
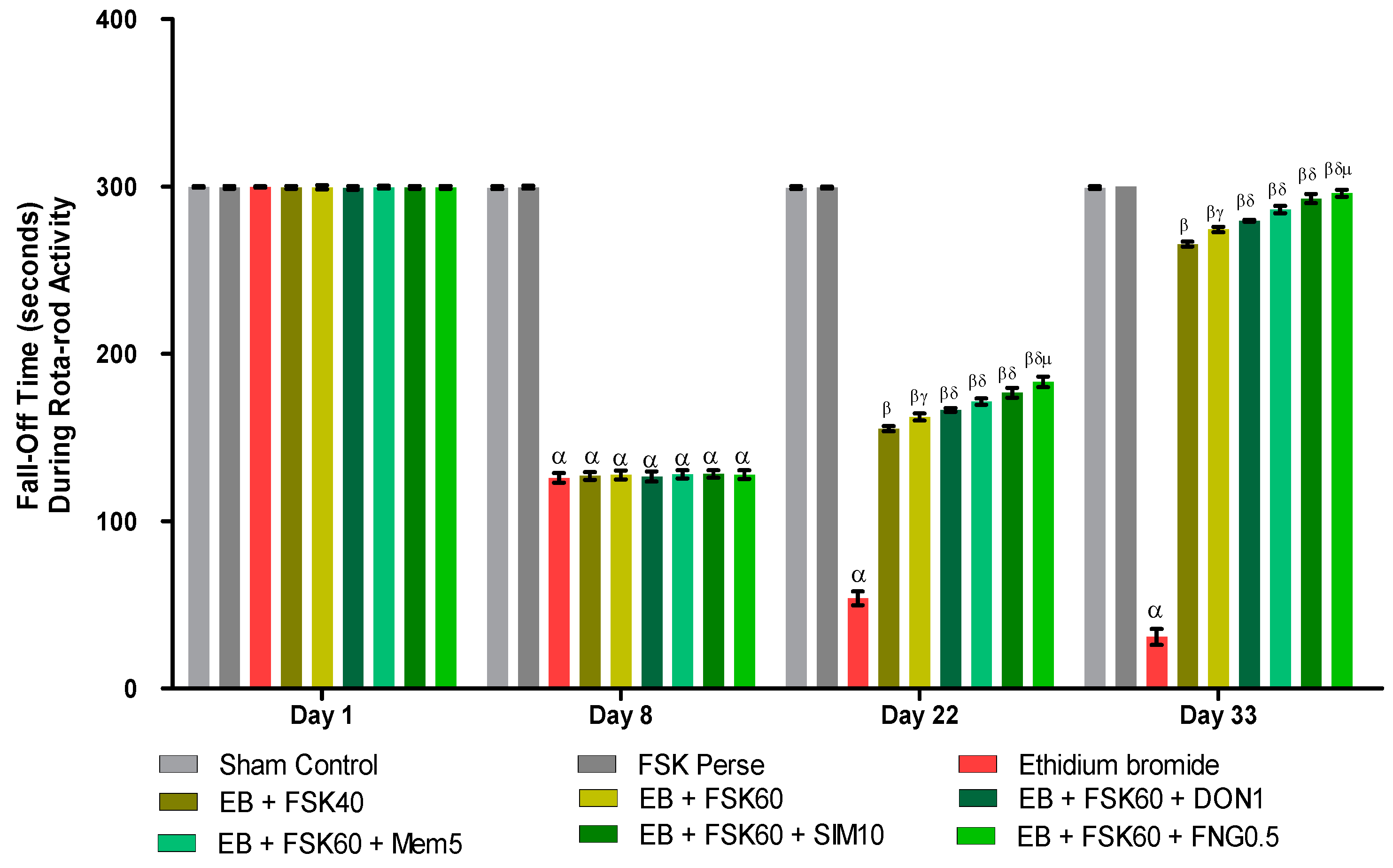
3.2.4. Protective Role of FSK on Grip Strength in Experimental Multiple Sclerosis Rats
3.3. Neurochemical Parameters
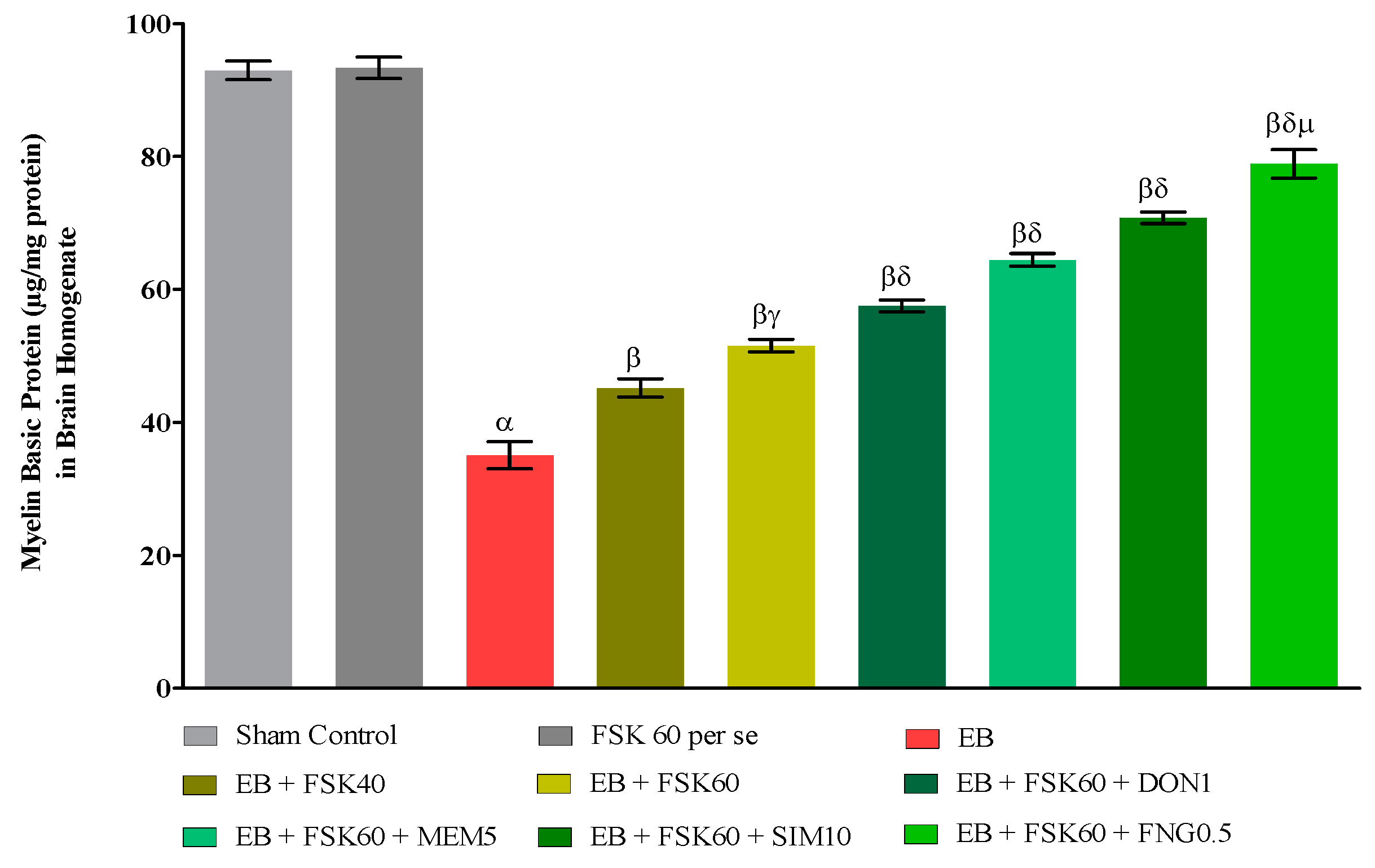
3.3.1. Protective Role of FSK on the Restoration of Myelination in Experimental Multiple Sclerosis Rats
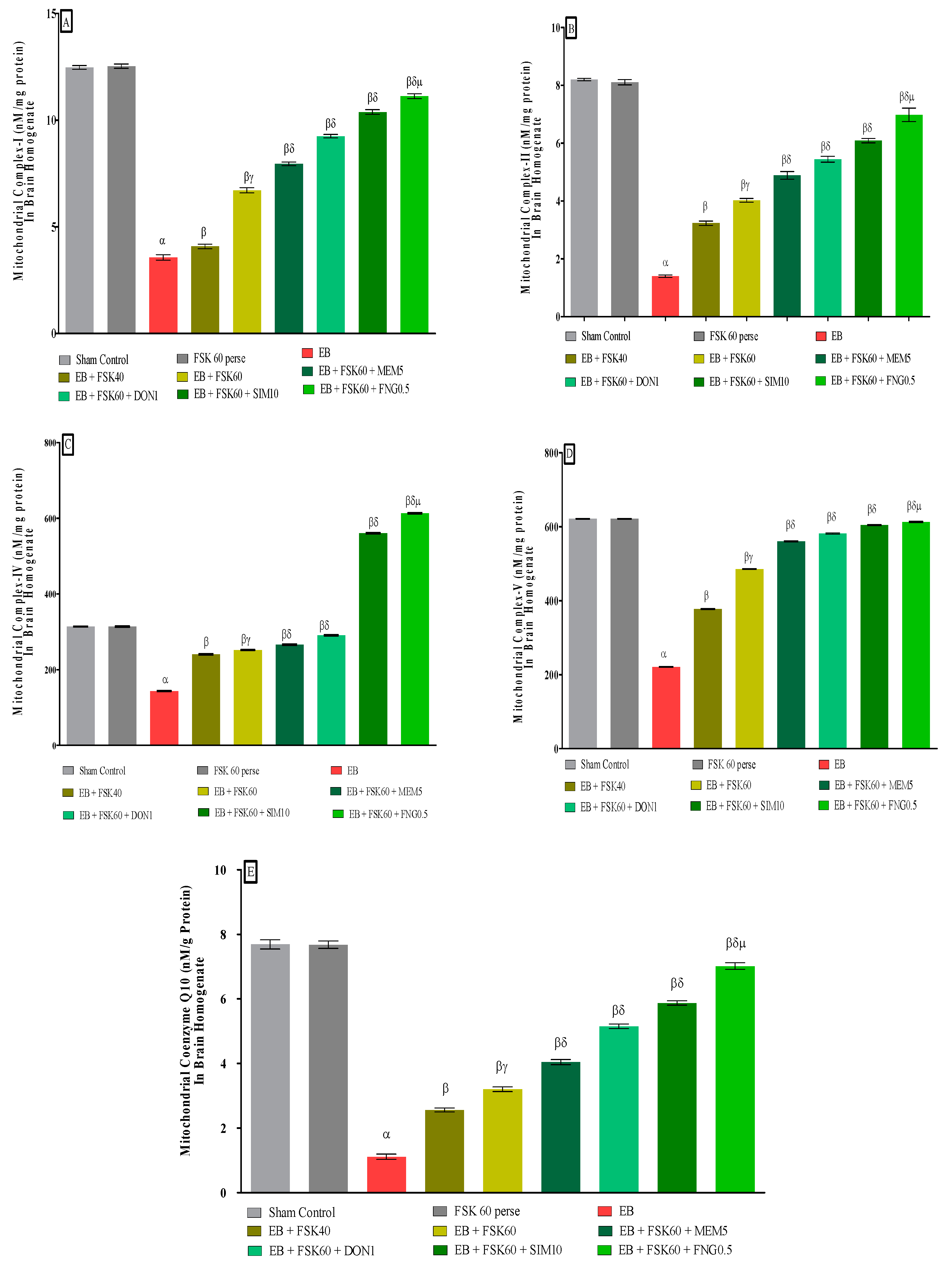
3.3.2. Protective Role of FSK in Restoring Mitochondrial ETC Complex Enzymes in Experimental Multiple Sclerosis Rats
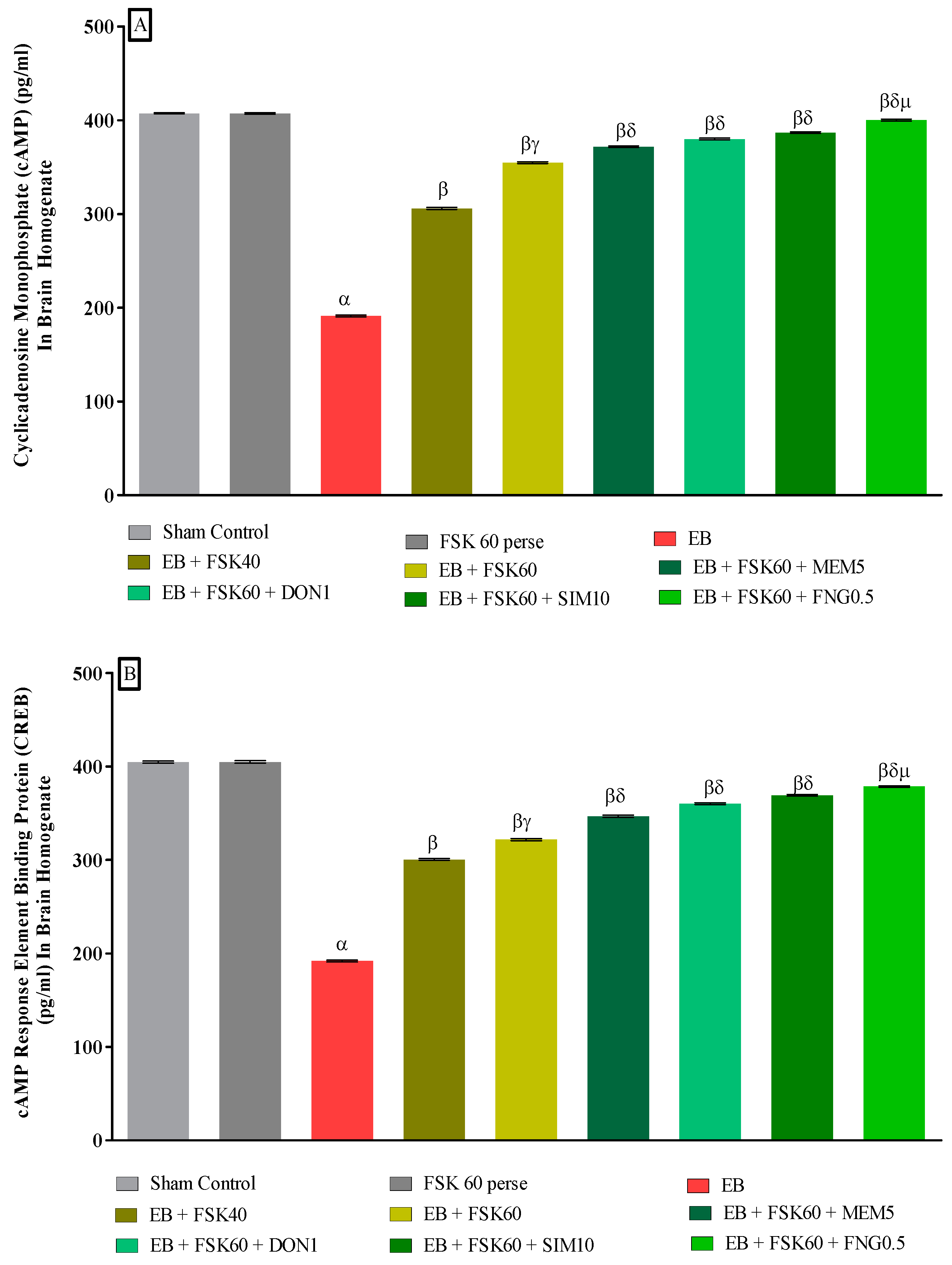
3.3.3. Protective Role of FSK in Upregulation of cAMP and CREB Levels in Experimental Multiple Sclerosis Rats
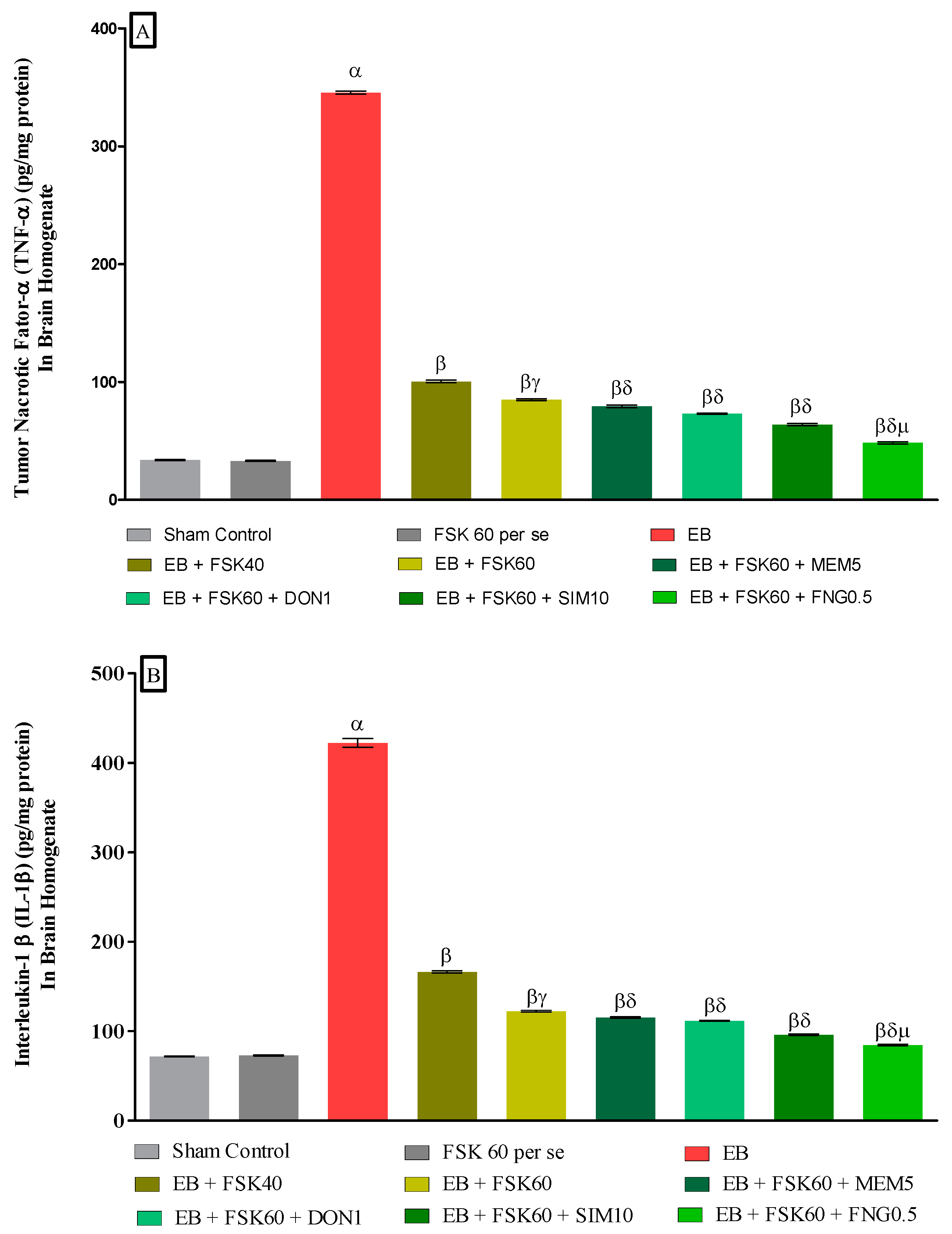
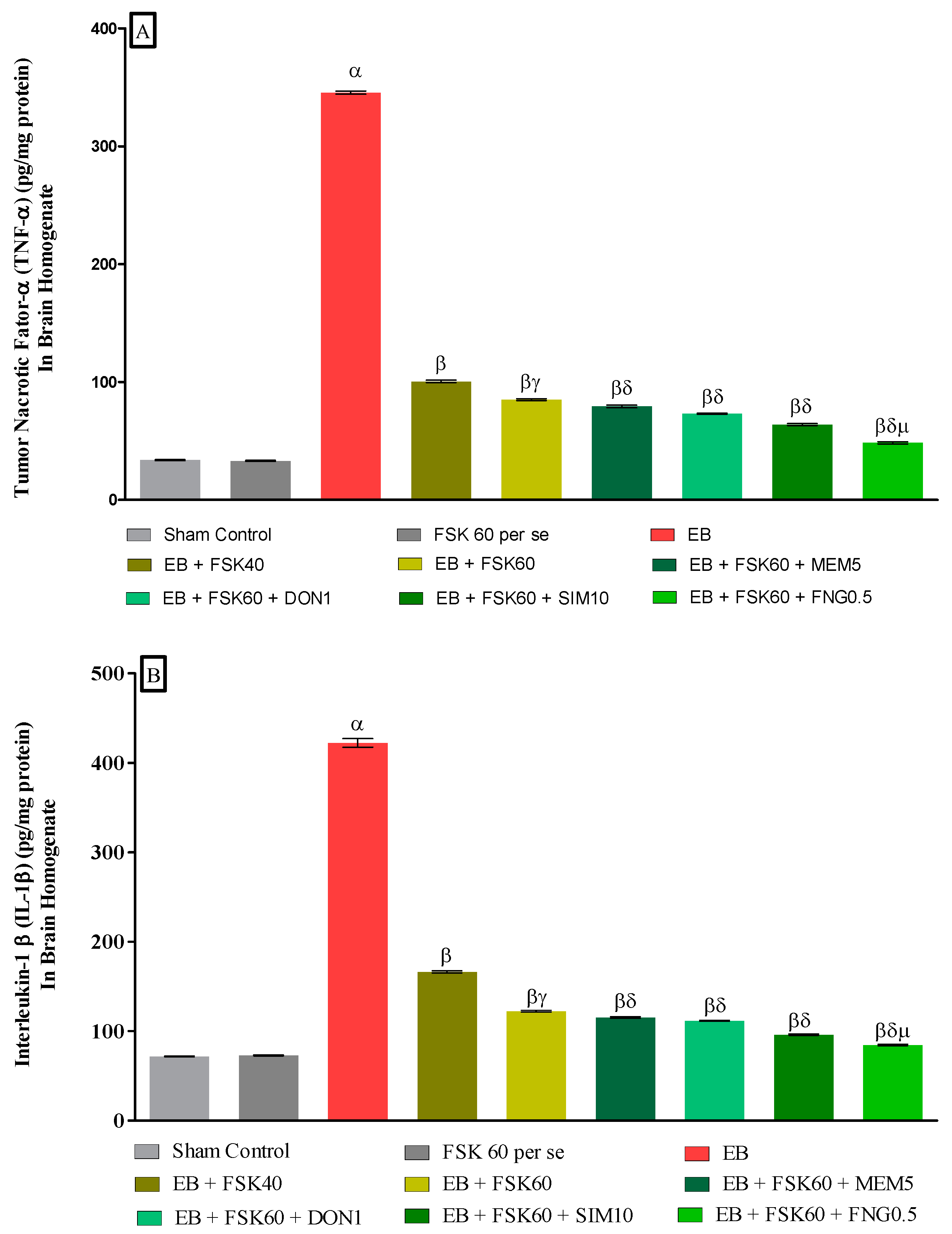
3.3.4. Protective Role of FSK on Amelioration of Inflammatory Cytokines Levels in Experimental Multiple Sclerosis Rats
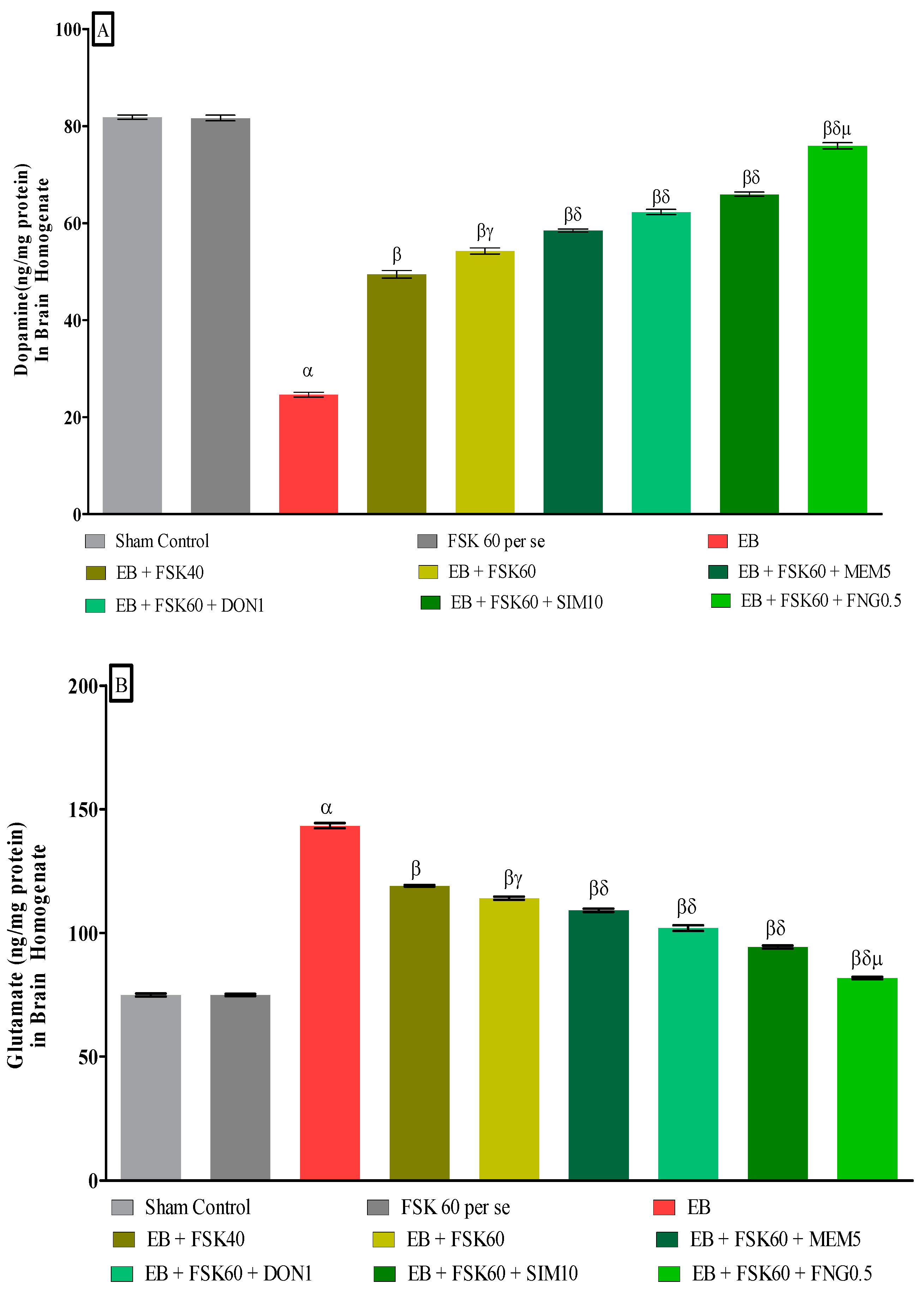
3.3.5. Protective Role of FSK on Ameliorating Neurotransmitter Alteration Levels in Experimental Multiple Sclerosis Rats
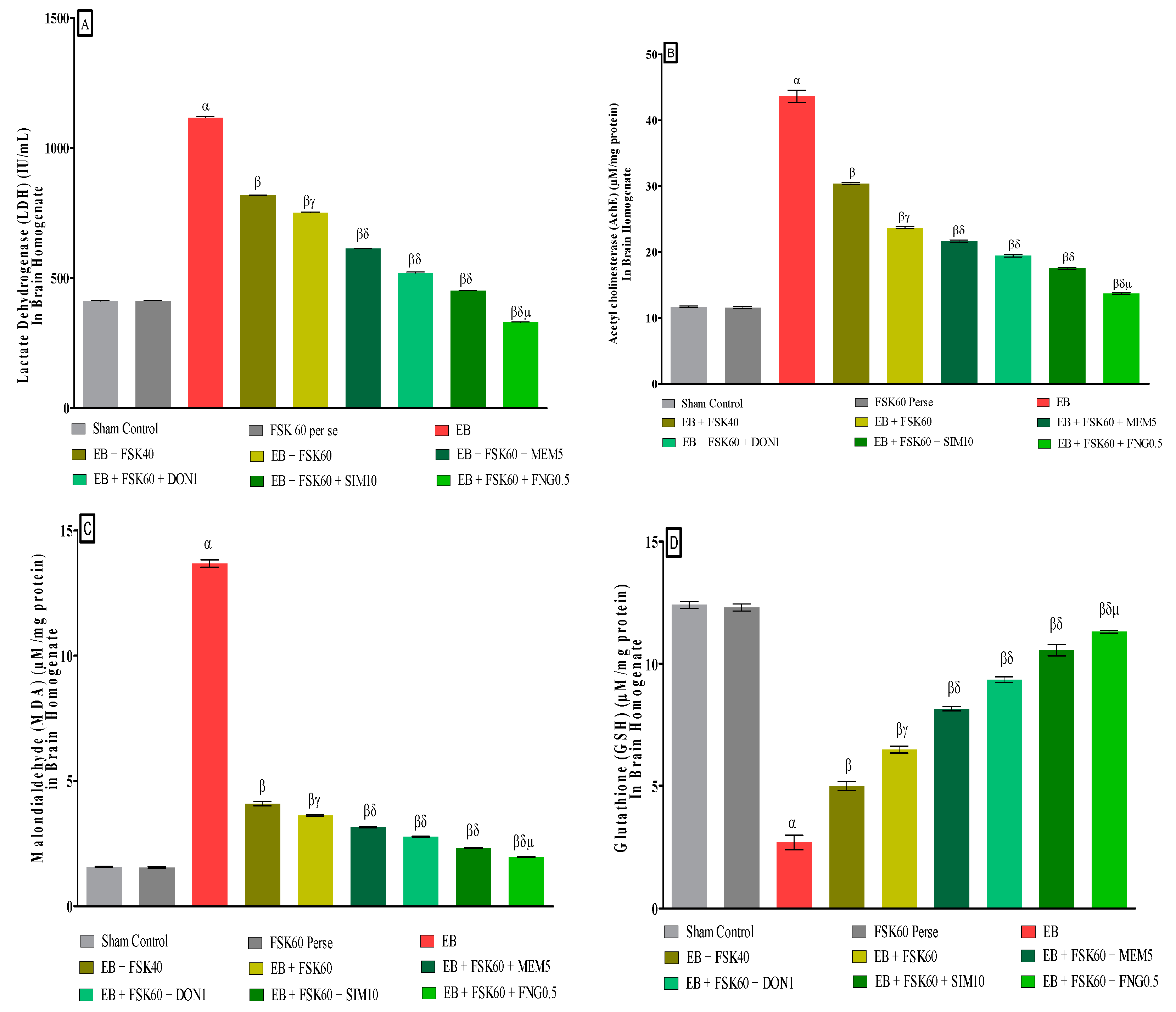
3.3.6. Protective Role of FSK on Reduction in Oxidative Stress Levels in Experimental Multiple Sclerosis Rats
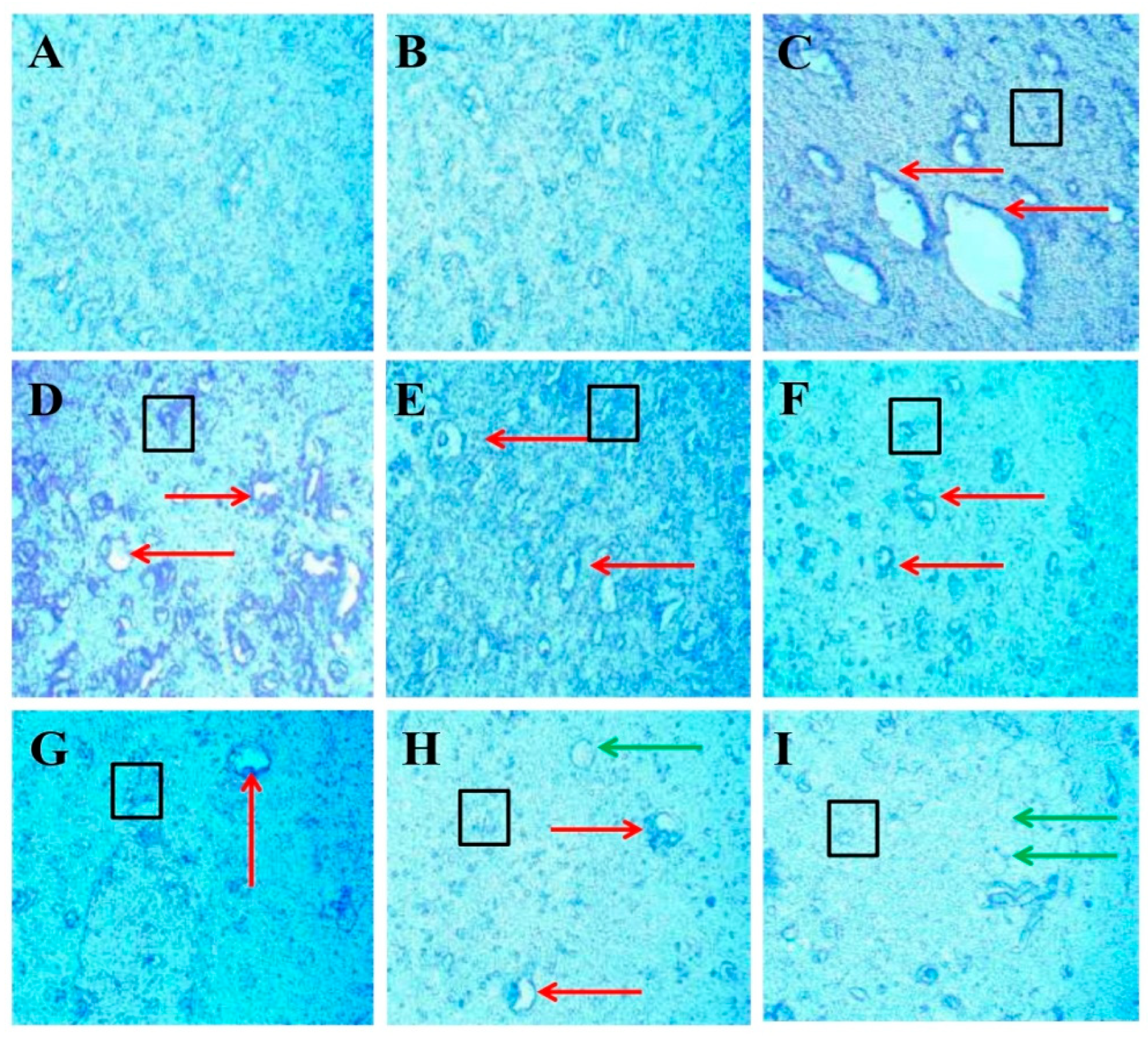
3.3.7. Protective Role of FSK on LFB Stain in Experimental Multiple Sclerosis Rats
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | Adenylylcyclase |
| Ach | Acetyl choline |
| Ach | Acetyl Cholinesterase |
| ALS | Amyotrophic Lateral Sclerosis |
| ANOVA | Analysis of Variance |
| ATP | Adenosine Triphosphate |
| BCT | Beam Crossing Task |
| BDNF | Brain-Derived Neurotrophic Factor |
| BHT | Butylated Hydroxytoluene |
| b-ME | b-Mercaptoethanol |
| cAMP | Cyclic Adenosine Monophosphate |
| CoQ10 | Coenzyme Q10 |
| CREB | cAMP Response Element Binding Protein |
| DNA | Deoxyribonucleic Acid |
| DON | Donepezil |
| DTNB | 5,5-dithiobis-(2-nitrobenzoic acid) |
| EB | Ethidium Bromide |
| ECD | Electrochemical Detector |
| EDTA | Ethylenediaminetetraacetic Acid |
| ELT | Escape Latency Task |
| ERK | Extracellular Signal-Regulated Kinase |
| ETC | Electron Transport Chain |
| FDA | Food and Drug Administration |
| FNG | Fingolimod |
| FSK | Forskolin |
| HD | Huntington’s Diseases |
| HMG-CoA | Hydroxymethylglutaryl-Coenzyme A |
| HPLC | High Performance Liquid Chromatography |
| IACE | Institutional Animal Ethics Committee |
| ICP | Intracerebropeduncle |
| IL-1β | Interleukin-1β |
| i.p. | Intraperitoneal injection |
| LA | Locomotor Activity |
| LDH | Lactate Dehydrogenase |
| LFB | Luxol Fast Blue |
| MBP | Myelin Basic Protein |
| MDA | Malondialdehyde |
| MEM | Memantine |
| MS | Multiple Sclerosis |
| mtDNA | Mitochondrial DNA |
| MWM | Morris Water Maze |
| NaCl | Sodium Chloride |
| NAD | Nicotinamide Adenosine Dinucleotide |
| NADH | Nicotinamide Adenosine Dinucleotide + Hydrogen |
| NADPH | Nicotinamide Adenosine Dinucleotide Phosphate |
| NaH2PO4 | Sodium Dihydrogen Phosphate |
| NaOH | Sodium Hydroxide |
| NBW | Narrow Beam Walk |
| NGF | Nerve Growth Factor |
| NMDA | N-methyl D-apartate |
| NMDAR | NMDA receptor |
| NO | Nitric Oxide |
| NTs | Neurotransmitters |
| OPA | o-phthaldehyde |
| OPC | Oligodendrocytes Precursor Cells |
| PKA | Protein kinase A |
| PMS | Post-Mitochondrial Supernatant |
| rpm | Revolution Per Minute |
| SD | Standard Deviation |
| SDH | Succinate Dehydrogenase |
| S1P | Spingosine 1-Phosphate |
| TNF- α | Tumor Nacrotic Factor- α |
| TSTQ | Time Spent in Target Quadrant |
References
- Wootla, B.; Eriguchi, M.; Rodriguez, M. Is multiple sclerosis an autoimmune disease? Autoimmune Dis. 2012, 2012, 969657. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Vidal-Jordana, A.; Montalban, X. Multiple sclerosis: Clinical aspects. Curr. Opin. Neurol. 2018, 31, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Walton, C.; King, R.; Rechtman, L.; Kaye, W.; Leray, E.; Marrie, R.A.; Robertson, N.; La Rocca, N.; Uitdehaag, B.; Van Der Mei, I.; et al. Rising prevalence of multiple sclerosis worldwide: Insights from the Atlas of MS, third edition. Mult. Scler. J. 2020, 26, 1816–1821. [Google Scholar] [CrossRef] [PubMed]
- Harbo, H.F.; Gold, R.; Tintoré, M. Sex and gender issues in multiple sclerosis. Ther. Adv. Neurol. Disord. 2013, 6, 237–248. [Google Scholar] [CrossRef]
- Bhatia, R.; Bali, P.; Chowdhary, R. Epidemiology and genetic aspects of multiple sclerosis in India. Ann. Indian Acad. Neurol. 2015, 18 (Suppl. 1), S6. [Google Scholar] [CrossRef] [PubMed]
- Upadhayay, S.; Mehan, S. Targeting Nrf2/HO-1 anti-oxidant signaling pathway in the progression of multiple sclerosis and influences on neurological dysfunctions. Brain Disord. 2021, 3, 100019. [Google Scholar] [CrossRef]
- Mehan, S.; Kapoor, T. Neuroprotective Methodologies in the Treatment of Multiple Sclerosis Current Status of Clinical and Pre-clinical Findings. Curr. Drug Discov. Technol. 2021, 18, 31–46. [Google Scholar]
- Singh, A.; Upadhayay, S.; Mehan, S. Inhibition of c-JNK/p38MAPK signaling pathway by Apigenin prevents neurobehavioral and neurochemical defects in Ethidium Bromide-induced experimental model of multiple sclerosis in rats: Evidence from CSF, blood plasma and brain samples. Phytomed. Plus 2021, 1, 100139. [Google Scholar] [CrossRef]
- Sharma, N.; Upadhayay, S.; Shandilya, A.; Sahu, R.; Singh, A.; Rajkhowa, B.; Mehan, S. Neuroprotection by solanesol against Ethidium Bromide-induced multiple sclerosis-like neurobehavioral, molecular, and neurochemical alterations in experimental rats. Phytomed. Plus 2021, 1, 100051. [Google Scholar] [CrossRef]
- Kumar, N.; Sharma, N.; Khera, R.; Gupta, R.; Mehan, S. Guggulsterone ameliorates Ethidium Bromide-induced experimental model of multiple sclerosis via restoration of behavioral, molecular, neurochemical and morphological alterations in rat brain. Metab. Brain Dis. 2021, 36, 911–925. [Google Scholar] [CrossRef]
- Wang, H.; Xu, J.; Lazarovici, P.; Quirion, R.; Zheng, W. cAMP Response Element-Binding Protein (CREB): A Possible Signaling Molecule Link in the Pathophysiology of Schizophrenia. Front. Mol. Neurosci. 2018, 11, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirò, G.; Iacono, S.; Ragonese, P.; Aridon, P.; Salemi, G.; Balistreri, C.R. A Brief Overview on BDNF-Trk Pathway in the Nervous System: A Potential Biomarker or Possible Target in Treatment of Multiple Sclerosis? Front. Neurol. 2022, 13, 917527. [Google Scholar] [CrossRef] [PubMed]
- Lee, D. Global and local missions of cAMP signaling in neural plasticity, learning, and memory. Front. Pharmacol. 2015, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Vakilzadeh, G.; Khodagholi, F.; Ghadiri, T.; Darvishi, M.; Ghaemi, A.; Noorbakhsh, F.; Gorji, A.; Sharifzadeh, M. Protective effect of a cAMP analogue on behavioral deficits and neuropathological changes in cuprizone model of demyelination. Mol. Neurobiol. 2015, 52, 130–141. [Google Scholar] [CrossRef]
- Schenk, P.W.; Snaar-Jagalska, B.E. Signal perception and transduction: The role of protein kinases. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 1999, 1449, 1–24. [Google Scholar] [CrossRef]
- Kaur, R.; Parveen, S.; Mehan, S.; Khanna, D.; Kalra, S. Neuroprotective effect of ellagic acid against chronically scopolamine induced Alzheimer’s type memory and cognitive dysfunctions: Possible behavioural and biochemical evidences. Int. J. Prev. Med. Res. 2015, 1, 45–64. [Google Scholar]
- Singh, L.; Rana, S.; Mehan, S. Role of adenylyl cyclase activator in controlling experimental diabetic nephropathy in rats. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 144. [Google Scholar]
- Alam, M.M.; Minj, E.; Yadav, R.K.; Mehan, S. Neuroprotective potential of adenylcyclase/cAMP/CREB and mitochondrial CoQ10 activator in amyotrophic lateral sclerosis rats. Curr. Bioact. Compd. 2021, 17, 53–69. [Google Scholar] [CrossRef]
- Mehan, S.; Khera, H.; Sharma, R. Neuroprotective Strategies of Blood-Brain Barrier Penetrant “Forskolin” (AC/cAMP/PK) to Ameliorate Mitochondrial Dysfunctioning in Neurotoxic Experimental Model of Autism. In Recent Advances in Neurodegeneration; IntechOpen: London, UK, 2019. [Google Scholar]
- Mehan, S.; Parveen, S.; Kalra, S. Adenylcyclase activator forskolin protects against Huntington’s disease-like neurodegenerative disorders. Neural Regen. Res. 2017, 12, 290. [Google Scholar] [CrossRef]
- Du, H.; Guo, L.; Wu, X.; Sosunov, A.A.; McKhann, G.M.; Chen, J.X.; Yan, S.S. Cyclophilin D deficiency rescues Aβ-impaired PKA/CREB signaling and alleviates synaptic degeneration. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 2517–2527. [Google Scholar] [CrossRef]
- Fanara, S.; Aprile, M.; Iacono, S.; Schirò, G.; Bianchi, A.; Brighina, F.; Dominguez, L.J.; Ragonese, P.; Salemi, G. The Role of Nutritional Lifestyle and Physical Activity in Multiple Sclerosis Pathogenesis and Management: A Narrative Review. Nutrients 2021, 25, 3774. [Google Scholar] [CrossRef] [PubMed]
- Groves, A.; Kihara, Y.; Chun, J. Fingolimod: Direct CNS effects of sphingosine 1-phosphate (S1P) receptor modulation and implications in multiple sclerosis therapy. J. Neurol. Sci. 2013, 328, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Pournajaf, S.; Dargahi, L.; Javan, M.; Pourgholami, M.H. Molecular Pharmacology and Novel Potential Therapeutic Applications of Fingolimod. Front. Pharmacol. 2022, 13, 807639. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, A.; Khanghahi, A.M.; Baharvand, H.; Javan, M. Fingolimod enhances oligodendrocyte differentiation of transplanted human induced pluripotent stem cell-derived neural progenitors. Iran. J. Pharm. Res. IJPR 2018, 17, 1444. [Google Scholar]
- Adlimoghaddam, A.; Neuendorff, M.; Roy, B.; Albensi, B.C. A review of clinical treatment considerations of donepezil in severe Alzheimer’s disease. CNS Neurosci. Ther. 2018, 24, 876–888. [Google Scholar] [CrossRef]
- Han, X.R.; Wen, X.; Wang, Y.J.; Wang, S.; Shen, M.; Zhang, Z.F.; Fan, S.H.; Shan, Q.; Wang, L.; Li, M.Q.; et al. Effects of CREB1 gene silencing on cognitive dysfunction by mediating PKA-CREB signaling pathway in mice with vascular dementia. Mol. Med. 2018, 24, 18. [Google Scholar] [CrossRef]
- Johnson, J.W.; Kotermanski, S.E. Mechanism of action of memantine. Curr. Opin. Pharmacol. 2006, 6, 61–67. [Google Scholar] [CrossRef]
- Saint Paul, L.P.; Creveuil, C.; Heinzlef, O.; De Seze, J.; Vermersch, P.; Castelnovo, G.; Cabre, P.; Debouverie, M.; Brochet, B.; Dupuy, B.; et al. Efficacy and safety profile of memantine in patients with cognitive impairment in multiple sclerosis: A randomized, placebo-controlled study. J. Neurol. Sci. 2016, 363, 69–76. [Google Scholar] [CrossRef]
- Stein, A.; Stroobants, S.; Gieselmann, V.; D’hooge, R.; Matzner, U. Anti-inflammatory therapy with simvastatin improves neuroinflammation and CNS function in a mouse model of metachromatic leukodystrophy. Mol. Ther. 2015, 23, 1160–1168. [Google Scholar] [CrossRef]
- Gupta, R.; Mehan, S.; Sethi, P.; Prajapati, A.; Alshammari, A.; Alharbi, M.; Al-Mazroua, H.A.; Narula, A.S. Smo-Shh agonist Purmorphamine prevents neurobehavioral and neurochemical defects in 8-OH-DPAT-induced experimental model of obsessive-compulsive disorder. Brain Sci. 2022, 12, 342. [Google Scholar] [CrossRef]
- Duggal, P.; Mehan, S. Neuroprotective approach of anti-cancer microtubule stabilizers against tauopathy associated dementia: Current status of clinical and preclinical findings. J. Alzheimer’s Dis. Rep. 2019, 3, 179–218. [Google Scholar] [CrossRef]
- Sharma, A.; Mehan, S. Targeting PI3K-AKT/mTOR signaling in the prevention of autism. Neurochem. Int. 2021, 147, 105067. [Google Scholar] [CrossRef] [PubMed]
- Sahu, R.; Mehan, S.; Kumar, S.; Prajapati, A.; Alshammari, A.; Alharbi, M.; Assiri, M.A.; Narula, A.S. Effect of alpha-mangostin in the prevention of behavioural and neurochemical defects in methylmercury-induced neurotoxicity in experimental rats. Toxicol. Rep. 2022, 9, 977–998. [Google Scholar] [CrossRef] [PubMed]
- Khera, R.; Mehan, S.; Bhalla, S.; Kumar, S.; Alshammari, A.; Alharbi, M.; Sadhu, S.S. Guggulsterone mediated JAK/STAT and PPAR-gamma modulation prevents neurobehavioral and neurochemical abnormalities in propionic acid-induced experimental model of autism. Molecules 2022, 27, 889. [Google Scholar] [CrossRef] [PubMed]
- Shandilya, A.; Mehan, S.; Kumar, S.; Sethi, P.; Narula, A.S.; Alshammari, A.; Alharbi, M.; Alasmari, A.F. Activation of IGF-1/GLP-1 Signalling via 4-Hydroxyisoleucine Prevents Motor Neuron Impairments in Experimental ALS-Rats Exposed to Methylmercury-Induced Neurotoxicity. Molecules 2022, 27, 3878. [Google Scholar] [CrossRef] [PubMed]
- Rajdev, K.; Siddiqui, E.M.; Jadaun, K.S.; Mehan, S. Neuroprotective potential of solanesol in a combined model of intracerebral and intraventricular hemorrhage in rats. IBRO Rep. 2020, 8, 101–114. [Google Scholar] [CrossRef]
- Mehan, S.; Rahi, S.; Tiwari, A.; Kapoor, T.; Rajdev, K.; Sharma, R.; Khera, H.; Kosey, S.; Kukkar, U.; Dudi, R. Adenylatecyclase activator forskolin alleviates intracerebroventricular propionic acid-induced mitochondrial dysfunction of autistic rats. Neural Regen. Res. 2020, 15, 1140. [Google Scholar] [CrossRef]
- Sharma, R.; Rahi, S.; Mehan, S. Neuroprotective potential of solanesol in intracerebroventricular propionic acid induced experimental model of autism: Insights from behavioral and biochemical evidence. Toxicol. Rep. 2019, 6, 1164–1175. [Google Scholar] [CrossRef]
- Minj, E.; Upadhayay, S.; Mehan, S. Nrf2/HO-1 signaling activator acetyl-11-keto-beta Boswellic acid (AKBA)-Mediated Neuroprotection in Methyl Mercury-induced experimental model of ALS. Neurochem. Res. 2021, 46, 2867–2884. [Google Scholar] [CrossRef]
- Siddiqui, E.M.; Mehan, S.; Upadhayay, S.; Khan, A.; Halawi, M.; Halawi, A.A.; Alsaffar, R.M. Neuroprotective efficacy of 4-Hydroxyisoleucine in experimentally induced intracerebral hemorrhage. Saudi J. Biol. Sci. 2021, 28, 6417–6431. [Google Scholar] [CrossRef]
- Rahi, S.; Gupta, R.; Sharma, A.; Mehan, S. Smo-Shh signaling activator purmorphamine ameliorates neurobehavioral, molecular, and morphological alterations in an intracerebroventricular propionic acid-induced experimental model of autism. Hum. Exp. Toxicol. 2021, 40, 1880–1898. [Google Scholar] [CrossRef]
- Yadav, R.K.; Mehan, S.; Sahu, R.; Kumar, S.; Khan, A.; Makeen, H.A.; Al Bratty, M. Protective effects of apigenin on methylmercury-induced behavioral/neurochemical abnormalities and neurotoxicity in rats. Hum. Exp. Toxicol. 2022, 41, 09603271221084276. [Google Scholar] [CrossRef] [PubMed]
- Maharaj, D.S.; Walker, R.B.; Glass, B.D.; Daya, S. 6-Hydroxymelatonin protects against cyanide induced oxidative stress in rat brain homogenates. J. Chem. Neuroanat. 2003, 26, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Bala, R.; Khanna, D.; Mehan, S.; Kalra, S. Experimental evidence for the potential of lycopene in the management of scopolamine induced amnesia. RSC Adv. 2015, 5, 72881–72892. [Google Scholar] [CrossRef]
- Deshmukh, R.; Sharma, V.; Mehan, S.; Sharma, N.; Bedi, K.L. Amelioration of intracerebroventricularstreptozotocin induced cognitive dysfunction and oxidative stress by vinpocetine—A PDE1 inhibitor. Eur. J. Pharmacol. 2009, 620, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Dudi, R.; Mehan, S. Neuroprotection of brain permeable Forskolin ameliorates behavioral, biochemical and histopatho-logical alterations in rat model of intracerebral hemorrhage. Pharmaspire 2018, 10, 68–86. [Google Scholar]
- Ghaiad, H.R.; Nooh, M.M.; El-Sawalhi, M.M.; Shaheen, A.A. Resveratrol promotes remyelination in cuprizone model of multiple sclerosis: Biochemical and histological study. Mol. Neurobiol. 2017, 54, 3219–3229. [Google Scholar] [CrossRef]
- Riet-Correa, G.; Fernandes, C.G.; Pereira, L.A.; Graça, D.L. Ethidium Bromide-induced demyelination of the sciatic nerve of adult Wistar rats. Braz. J. Med. Biol. Res. 2002, 35, 99–104. [Google Scholar] [CrossRef]
- Goudarzvand, M.; Choopani, S.; Shams, A.; Javan, M.; Khodaii, Z.; Ghamsari, F.; Naghdi, N.; Piryaei, A.; Haghparast, A. Focal injection of Ethidium Bromide as a simple model to study cognitive deficit and its improvement. Basic Clin. Neurosci. 2016, 7, 63. [Google Scholar]
- Markianos, M.; Evangelopoulos, M.E.; Koutsis, G.; Davaki, P.; Sfagos, C. Body mass index in multiple sclerosis: Associations with CSF neurotransmitter metabolite levels. Int. Sch. Res. Not. 2013, 2013, 981070. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Fulk, G.D. Locomotor training and virtual reality-based balance training for an individual with multiple sclerosis: A case report. J. Neurol. Phys. Ther. 2005, 29, 34–42. [Google Scholar] [CrossRef]
- Wajda, D.A.; Sosnoff, J.J. Cognitive-motor interference in multiple sclerosis: A systematic review of evidence, correlates, and consequences. BioMed Res. Int. 2015, 2015, 720856. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.N.; Schmidt, R.L.; Wilson, A.R.; Freedman, M.S.; Grenache, D.G. Cerebrospinal fluid myelin basic protein is frequently ordered but has little value: A test utilization study. Am. J. Clin. Pathol. 2012, 138, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, I.P.; Troxell, R.M.; Graves, J.S. Mitochondrial dysfunction and multiple sclerosis. Biology 2019, 11, 37. [Google Scholar] [CrossRef]
- Caminero, A.; Comabella, M.; Montalban, X. Tumor necrosis factor alpha (TNF-α), anti-TNF-α and demyelination revisited: An ongoing story. J. Neuroimmunol. 2011, 234, 1–6. [Google Scholar] [CrossRef]
- Dujmovic, I.; Mangano, K.; Pekmezovic, T.; Quattrocchi, C.; Mesaros, S.; Stojsavljevic, N.; Nicoletti, F.; Drulovic, J. The analysis of IL-1 beta and its naturally occurring inhibitors in multiple sclerosis: The elevation of IL-1 receptor antagonist and IL-1 receptor type II after steroid therapy. J. Neuroimmunol. 2009, 15, 101–106. [Google Scholar] [CrossRef]
- Noorzehi, G.; Pasbakhsh, P.; Borhani-Haghighi, M.; Kashani, I.R.; Madadi, S.; Tahmasebi, F.; Nekoonam, S.; Azizi, M. Microglia polarization by methylprednizolone acetate accelerates cuprizone induced demyelination. J. Mol. Histol. 2018, 49, 471–479. [Google Scholar] [CrossRef]



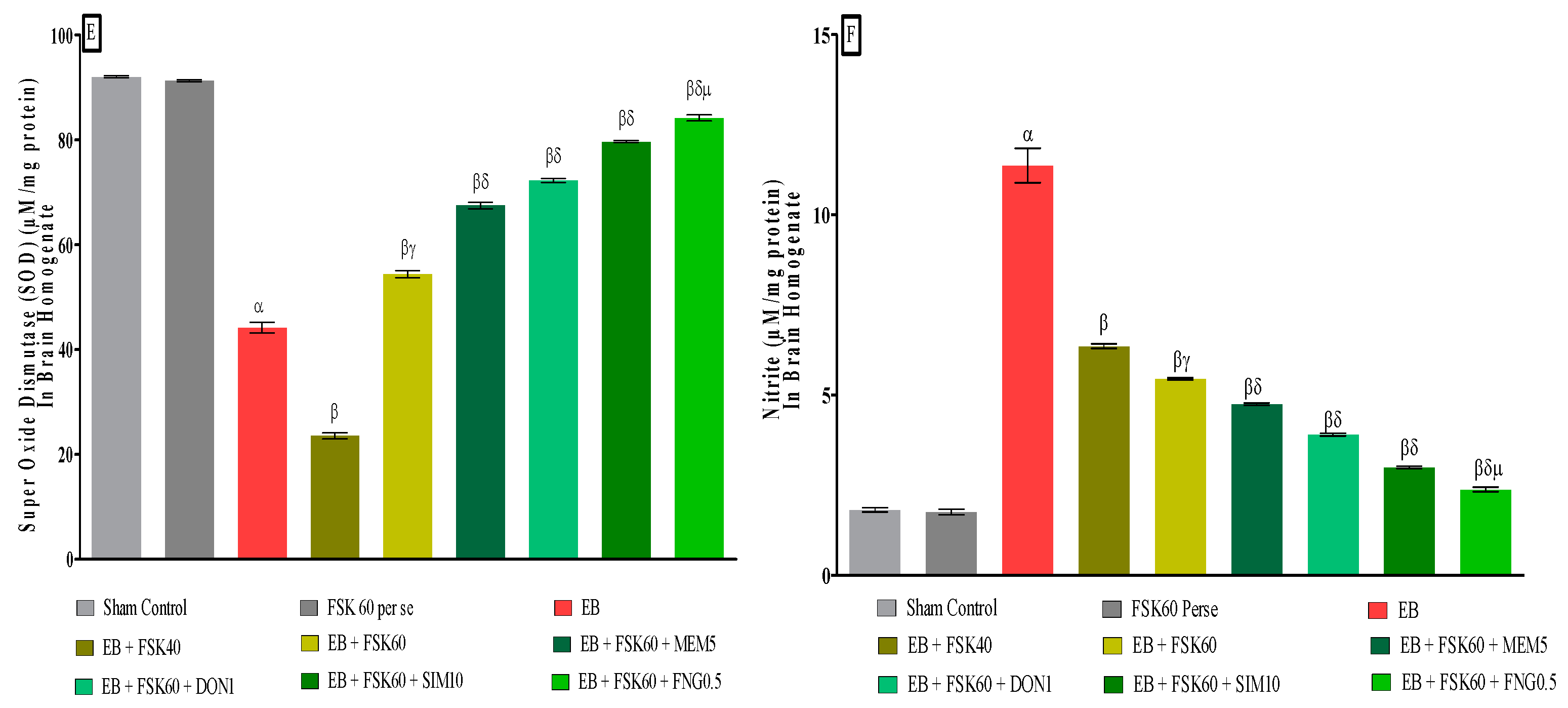

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapoor, T.; Mehan, S.; Suri, M.; Sharma, N.; Kumar, N.; Narula, A.S.; Alshammari, A.; Alasmari, A.F.; Alharbi, M.; Assiri, M.A.; et al. Forskolin, an Adenylcyclase/cAMP/CREB Signaling Activator Restoring Myelin-Associated Oligodendrocyte Destruction in Experimental Ethidium Bromide Model of Multiple Sclerosis. Cells 2022, 11, 2771. https://doi.org/10.3390/cells11182771
Kapoor T, Mehan S, Suri M, Sharma N, Kumar N, Narula AS, Alshammari A, Alasmari AF, Alharbi M, Assiri MA, et al. Forskolin, an Adenylcyclase/cAMP/CREB Signaling Activator Restoring Myelin-Associated Oligodendrocyte Destruction in Experimental Ethidium Bromide Model of Multiple Sclerosis. Cells. 2022; 11(18):2771. https://doi.org/10.3390/cells11182771
Chicago/Turabian StyleKapoor, Tarun, Sidharth Mehan, Manisha Suri, Nidhi Sharma, Nitish Kumar, Acharan S. Narula, Abdulrahman Alshammari, Abdullah F. Alasmari, Metab Alharbi, Mohammed A. Assiri, and et al. 2022. "Forskolin, an Adenylcyclase/cAMP/CREB Signaling Activator Restoring Myelin-Associated Oligodendrocyte Destruction in Experimental Ethidium Bromide Model of Multiple Sclerosis" Cells 11, no. 18: 2771. https://doi.org/10.3390/cells11182771
APA StyleKapoor, T., Mehan, S., Suri, M., Sharma, N., Kumar, N., Narula, A. S., Alshammari, A., Alasmari, A. F., Alharbi, M., Assiri, M. A., & Kalfin, R. (2022). Forskolin, an Adenylcyclase/cAMP/CREB Signaling Activator Restoring Myelin-Associated Oligodendrocyte Destruction in Experimental Ethidium Bromide Model of Multiple Sclerosis. Cells, 11(18), 2771. https://doi.org/10.3390/cells11182771