Abstract
The ameliorative and antioxidative stress effects of probiotic-enriched fermented oat (FOE) or fermented oat with honey (HFOE) extracts on streptozotocin-induced diabetes in rats were examined. The total phenolic content (TPC) and antioxidant activity (AOA) were increased in FOE and HFOE after 72 h of fermentation, and γ-aminobutyric acid (GABA) reached 7.35 mg 100 g−1 in FOE and 8.49 mg 100 g−1 in HFOE. The β-glucan levels were slightly decreased to 2.45 g 100 g−1 DW in FOE and 2.63 g 100 g−1 DW in HFOE. The antidiabetic and hypolipidemic properties of FOE and HFOE were studied in a designed animal model with seven treated groups for 6 weeks. Groups were treated as follows: group 1 (negative group, NR) and group 2 (diabetic rats, DR) were administered 7 mL distilled water orally per day; group 3 (DR + MET) rats were orally administered 50 mg standard drug Metformin kg−1 daily; group 4 (DR + FOE1) diabetic rats were orally administered 3.5 mL FOE daily; group 5 (DR + FOE2) rats were orally administered 7 mL FOE daily; group 6 (DR + HFOE1) rats were orally administered 3.5 mL HFOE daily; and group 7 (DR + HFOE2) rats were orally administered 7 mL HFOE daily. The HFOE at the high dose had a synergistic effect, lowering random blood glucose (RBG) and fasting blood glucose (FBG). The hypolipidemic potential of HFOE at the high dose was indicated by significant reductions in triglycerides (TG), total cholesterol (CHO), high- and low-density lipoproteins (HDL and LDL), and very-low-density lipoproteins (VLDL). In addition, 7 mL of HFOE improved liver and kidney function more effectively than other fermented extracts or Metformin. As well as the antioxidant enzyme activity, reduced glutathione (GSH), catalase (CAT), superoxide dismutase (SOD), and malonaldehyde (MDA) were significantly enhanced after the administration of HFOE at 7 mL by 68.6%, 71.5%, 55.69%, and 15.98%, respectively, compared to the DR group. In conclusion, administration of L. plantarum-fermented oats supplemented with honey demonstrated antidiabetic effects and a potential approach for controlling glucose levels and lipid profiles, and protecting against oxidative stress.
1. Introduction
The effectiveness of common oats (Avena sativa) in regulating blood glucose levels has been documented since 1903 [1]. Numerous investigations have reported oats’ therapeutic effects on diabetes and multiple other health issues, such as cardiovascular disease, hypertension, and dyslipidemia [2,3]. Soluble dietary fiber, namely β-glucan, plays a significant role in reducing cholesterol, glucose, and insulin levels [4,5,6]. The potent antioxidant capacity of oats, mainly derived from the oats’ unique polyphenols, known as avenanthramides, also delivers multiple health-promoting effects, such as antiatherosclerosis and anti-inflammatory activities [7,8].
Oat fermentation has been shown to significantly increase the antioxidant capacity much more so than non-fermented oat products [9,10], due to the changes and release of multiple nutrients during fermentation [5]. These changes result from the microbial metabolic activities that mainly break down carbohydrate fractions into several end products, such as acids. The presence of multiple end products and the release of several phytochemicals deliver various health-promoting properties, such as the prevention of cardiovascular disease [11], hypertension [12,13,14,15], obesity [16,17], and osteoporosis [18]. In addition, a variety of fermented foods have shown antidiabetic effects both in vitro and in vivo [6] using a wide range of different microorganisms as starter cultures. The well-known Lactobacillus species, such as L. fermentum, L. acidophilus, and L. plantarum, have been widely used as probiotics in the food industry [19]. Various strains have been reported to exert health-promoting activities, such as immunomodulation and antipathogenic activities, and have high cholesterol-lowering efficiencies [20].
Furthermore, a recent in vitro investigation [21] reported that L. plantarum promotes antidiabetic effects. A similar strain was also used in vivo to investigate its antidiabetic effects [22]. Their results showed a reduction in blood glucose levels, and the authors indicated that L. plantarum is a potential therapeutic agent for the management of diabetes. Moreover, a recent investigation has shown that the fermentation of oats with the addition of honey delivers even more desirable results [11], such as maintaining the concentrations of β-glucan at optimum levels. β-glucan is a selective substrate of Lactobacillus [23], and thus, the addition of honey provides an energy source for bacterial growth.
The development of a new dietary intervention for the management of diabetes is highly urgent. The prevalence of diabetes has increased dramatically in recent years. Statistical results showed that more than 6 million deaths were caused directly by diabetes in 2021 [24,25]. In Saudi Arabia, diabetes is the 10th highest cause of death [26]. The prevalence of new cases is also growing day by day. About 50% of the Saudi population are predicted to be diabetic by 2030 [19,27]. Therefore, the prevention of diabetes and seeking new dietary interventions is much needed. This study investigates the antidiabetic efficiency in terms of the ameliorative and antioxidative stress potential of L. plantarum-fermented oat extract (FOE) and fermented oat extract supplemented with honey (HFOE) on several biomarkers in animal diabetic models. To the best of our knowledge, the antidiabetic potential of HFOE using the L. plantarum strain in the form of a functional beverage has not been studied yet.
2. Materials and Methods
2.1. Ingredients, Chemicals, and Strain
Whole oat grains were purchased from the local store of Buraydah, Saudi Arabia (manufactured by Federal Oats Mills, 13400, Butterworth, Malaysia). The nutritional value of oats per 100 g consists of 370 kcal, 12.1 g protein, 8.4 g fat, 56.1 g carbohydrates, 1.0 g sugar, 10.8 g dietary fiber, 4.0 mg iron, 110.0 mg magnesium, 3.0 mg zinc, 52.0 µg folic acid, 1.0 mg, 0.4 mg, and 0.4 µg of vitamin A, B1, and B12, respectively. Sidr honey was obtained from the local market of Riyadh, Saudi Arabia.
A strain of Lactobacillus plantarum (NRRL B-59151) was generously provided by the USDA Agricultural Research Service (ARS) Culture Collection (Peoria, IL, USA), and MRS broth was purchased from Condalab (Madrid, Spain). Streptozotocin (STZ), >97% purity, was purchased from Alfa Aesar, Thermo Fisher Scientific (Kandel, Germany), and Metformin in pure form was purchased from Sigma-Aldrich (Saint Louis, MO, USA). A mixed-linkage β-glucan kit was purchased from Megazyme International (Bray, Ireland), and γ-aminobutyric acid (GABA) analytical standard was purchased from Sigma-Aldrich (Saint Louis, MO, USA).
2.2. Preparation of Fermented Oat (FOE) and Honey-Supplemented Fermented Oat (HFOE) Extracts
The preparation of the fermented extracts was performed following the method of Chen et al. [11], with slight modifications. Briefly, a suspension of 2% Sidr honey in distilled water was pasteurized at 80 °C for 10 min. The suspension was then added to sterilized whole oat flour to yield a concentration of 10% oats (w/v). After cooling down to room temperature, the starter culture of L. plantarum was inoculated at 1% (v/v). The mixture was then fermented at 37 °C for 24, 48, and 72 h in a microbiological incubator. A fermented oat mixture without incorporating honey was prepared following the same method. After fermentation, samples were centrifuged at 12,000× g for 15 min at 4 °C. The collected supernatants were stored at 4–8 °C for 7 days. These extracts were orally administered to the rats, and fresh-made extracts were prepared each week.
2.3. Enumeration of L. plantarum B-59151 Viable Count
The viable count of L. plantarum B-59151 in FO and HFO was enumerated following the standard plate count method according to Vinderola and Reinheimer [28]. Firstly, 10 mL of each sample was suspended in 90 mL of sterile peptone water (Merck, 0.1%), after which it was serially diluted. Aliquots of 1 mL of appropriate dilutions were inoculated on sterile plates of MRS agar. MRS agar-inoculated plates were incubated at 37 °C for 48–72 h in anaerobic jars (2.5 L) with a GasPak system (GasPak System-Oxoid, Basingstoke, Hampshire, UK) according to standard methods [28]. Data are expressed as the logarithm of colony forming units per mL (Log10 CFU mL−1).
2.4. Determination of Total Phenolic Content (TPC) in FOE and HFOE
The TPC of FOE and HFOE was determined using Folin–Ciocalteu reagent, according to Yawadio Nsimba et al. [29]. Briefly, in Eppendorf tubes, 150 µL of the sample was mixed with 300 µL of Folin–Ciocalteu reagent for 5 min. Then, 300 µL of an alkali solution (7.5% sodium carbonate solution, Na2CO3) was added. The mixture was incubated in the dark for 60 min at 23 °C, then centrifuged at 10,000× g for 10 min at 4 °C, and 200 µL of supernatant from each Eppendorf was transferred to a new 96-well plate; the absorbance was then measured at 765 nm using a microplate reader (BioTek, Winooski, VT, USA). Measurements were compared against the standard curve of gallic acid (GA) solution (R2 = 0.99), and TPC content was expressed as milligrams of Gallic acid equivalents (GAE) per gram of dry weight (DW) (mg of GAE g−1 DW).
2.5. Determination of Total Antioxidant Capacity (TAC) of FOE and HFOE
The TAC of FOE and HFOE was measured spectrophotometrically based on the bleaching of DPPH radicals in purple solution according to Yawadio Nsimba et al. [29]. Briefly, 600 µL of DPPH solution was added to 100 µL of sample in Eppendorf tubes. After incubation in the dark for 60 min at 25 °C, tubes were centrifuged at 10,000× g for 5 min at 4 °C, after which, 200 µL of supernatant from each Eppendorf was transferred to a new 96-well plate; the absorbance was then measured at 517 nm using a microplate reader (BioTek, Winooski, VT, USA). The DPPH radical scavenging activity percentage was calculated based on the plotted Trolox calibration curve. The antiradical activity was expressed as micromoles of Trolox equivalents (TE) per gram of dry weight (µmol TE g−1 DW). The ABTS radical scavenging activity of FOE and HFOE against ABTS radicals was tested by the adapted method of Lu et al. [30].
2.6. Determination of the γ-Aminobutyric Acid (GABA) Content
The content of GABA was evaluated spectrophotometrically according to Yeap et al. [31]. Briefly, 0.5 mL of extract was mixed with 0.5 mL of borate buffer, 0.5 mL of 6% phenol reagent, and 1.5 mL of 6% sodium hypochlorite (NaClO); the mixture was then boiled at 100 °C for 10 min and immediately cooled down in a cooling bath. The absorbance was then measured at 630 nm. A GABA standard was used to prepare a standard curve, and the results were expressed as mg 100 g−1 DW.
2.7. Determination of the β-Glucan Content
The β-glucan content in the fermented oat mixtures was quantified using an enzymatic mixed-linkage assay kit (Megazyme International Co., Wicklow, Ireland). According to the manufacturer’s protocol. All measurements were performed twice and in duplicate, following the method of a previous study [11]. β-glucan content was expressed as g 100 g−1 DW.
2.8. Animals and Experimental Design
This study used Wistar rats (56 adult males) weighing between 170 and 190 g. Under standard laboratory conditions, the animals were housed in air-conditioned polypropylene cages and kept at 24 ± 1 °C, 40–45% relative humidity, on a 12 h light/dark cycle, and were fed a commercial standard pellet diet and given water ad libitum [32]. All experiments were approved by the Committee of Research Ethics (Institutional Review Board, IRB) of Qassim University, Saudi Arabia (Approval No. 21-13-19 on 21 March 2022).
After ten days of acclimatization, rats were randomly divided into 7 groups (8 rats/group). The first group, normal rats (NR), received an intraperitoneal injection of saline solution and 5 mL distilled water orally per day. The rest of the groups were fasted overnight and received a single intraperitoneal injection of freshly prepared solution of STZ in 0.1 M citrate buffer (pH = 4.5) at a dose of 45 mg kg−1 BW, to induce diabetes. The confirmation of diabetes was assessed by monitoring the fasting blood glucose (FBG) levels after 48 h of STZ injection using a glucometer (Accu-Check, Roche, Germany). Experimental rats with an FBG level > 200 mg dL−1 were considered diabetic, and were included in the study. Animals were randomized based on their body mass and random blood glucose (RBG) levels, and divided into 6 groups, as shown in Table 1. Metformin was chosen as a reference drug as it is the first-line medication for treating type 2 diabetes [33].

Table 1.
Experimental design of streptozotocin-induced diabetic rats treated with FOE and HFOE daily for 6 weeks.
At the end of the 6th week, 12 h-fasted animals were anesthetized with a mixture of alcohol, chloroform, and ether (1:2:3). Collected blood samples were subjected to serum separation by centrifugation at 4000× g for 30 min under cooling to attain serum used for various biochemical parameters. The biochemical parameters were determined using suitable kits and a blood chemistry analyzer (HumaLyzer 4000, Human Gesellschaft für Biochemica und Diagnostica mbH, Wiesbaden, Germany).
2.8.1. Determination of Fasting Blood Glucose Level (FBG), Lipid Profile, and Liver and Kidney Functions
The GOD–PAP method was used to determine FBG (mg dL−1) using an enzymatic colorimetric test kit. Triglycerides (TG, mg dL−1) and total cholesterol (CHO, mg dL−1) were determined using an enzymatic colorimetric test kit and the GPO–PAP method, respectively. High-density lipoproteins (HDL, mg dL−1) were determined using an enzymatic colorimetric direct homogenous test kit following manufacturer protocols. According to Friedewald et al. [34], low-density lipoproteins (LDL, mg dL−1) and very-low-density lipoproteins (VLDL, mg dL−1) were calculated mathematically using indicated equations. The liver function biomarkers, such as alanine aminotransferase (ALT, UL−1), aspartate aminotransferase (AST, UL−1), alkaline phosphatase (ALP, UL−1), and total bilirubin (T. Bili, mg dL−1) in blood serum were examined using an alanine aminotransferase kit (EC 2.6.1.2), aspartate aminotransferase kit (EC 2.6.1.1), and an alkaline phosphatase kit (2.6.1.4). Kidney function biomarkers, such as total protein (T. Protein, g dL−1), albumin (g dL−1), creatinine (mg dL−1), and urea (mg dL−1) concentrations were respectively determined using photometric and colorimetric test kits applying the Biuret method, photometric and colorimetric test kits applying the BCG method, photometric and colorimetric test kits, and a fully enzymatic test kit applying the GLDH method, according to the instructions of the manufacturer. Globulin concentrations (g dL−1) were calculated by subtracting albumin from T. Protein concentrations. Blood urea nitrogen (BUN, mg dL−1) was calculated by multiplying urea concentration by 0.47. All biochemical examination kits were purchased from Human Co., Wiesbaden, Germany. The atherogenic index (AI) was calculated according to Nwagha et al. [35].
2.8.2. Oxidative Stress Biomarkers
Reduced glutathione (GSH, µg dL−1) was estimated using a GSH colorimetric assay kit (E-BC-K030-S, Elabscience, Houston, TX, USA) according to the method described by Beutler et al. [36]. Lipid peroxidation was assessed using a malondialdehyde (MDA, nmol mL−1) colorimetric assay kit (E-BC-K025-S, Elabscience, Houston, TX, USA) measuring TBARS, and expressed in terms of MDA content according to Ohkawa et al. [37]. MDA, a byproduct of fatty acid peroxidation, reacts with thiobarbituric acid to form a colored complex (TBA). The absorbance of the supernatant was measured at 532 nm and converted to nmol mL−1. Superoxide dismutase (SOD, U L−1) activity was determined using a SOD activity assay kit (E-BC-K022-S, Elabscience, Houston, TX, USA) according to Giannopolitis and Ries [38]. At 550 nm, the color reaction was measured and expressed as U L−1. The activity of catalase (CAT, U L−1) was measured using a CAT activity assay kit (E-BC-K031-S, Elabscience, Houston, TX, USA) and the Aebi method [39]. A blood chemistry analyzer was used to determine all oxidative stress markers (HumaLyzer 4000, Human Gesellschaft für Biochemica und Diagnostica mbH, Wiesbaden, Germany).
2.9. Statistical Analysis
The statistical analysis was carried out by applying a one-way ANOVA for phytochemicals and antioxidant capacity, γ-aminobutyric acid content, β-glucan content, lipid profile parameters, liver and kidney functions, and antioxidant biomarker data, and a two-way ANOVA for L. plantarum viable count, and pH and RBG data using SPSS (Ver. 22.0 for Windows). Experimental results are expressed as mean ± standard error. Applying Tukey’s test, multiple comparisons were carried out, and the significance level was set at <0.05. Data were treated as a complete randomization design according to Steel et al. [40].
3. Results
3.1. Survival of Lactobacillus plantarum and Related pH Value in FOE and HFOE
The current investigation used sterilized oat flour, obtained from whole oat grains, in water (1:9, w/v) as the fermentation media for L. plantarum (FOE), and (1:9, w/v) with 2% Sidr honey (HFOE). Immediately, L. fermentum was inoculated into both media in the range of 7.45–7.51 (CFU mL−1) using a freshly prepared starter culture. The growth profile, as shown in Figure 1, demonstrated that there was a significant increase in the viable count during the first 24 h (8.25 log CFU mL−1 for FOE and 8.52 log CFU mL−1 for HFOE) as compared to 0 h, with a slight decrease after 48 h (8.16 log CFU mL−1 for FOE) and a slight increase after 48 h (8.65 log CFU mL−1 for HFOE). Indeed, the viable count remained relatively stable during this period, confirming the steady growth of L. plantarum. The viable cell count was 8.36 and 8.57 log CFU mL−1 in FOE and HFOE, respectively, at maximum fermentation time (72 h). This increase in cell growth was accompanied with a significant decrease in pH values from 6.36 to 4.73 after 24 h, 4.63 after 48 h, and a further reduction to 4.60 at 72 h for FOE, and from 6.36 to 4.05 after 24 h, 3.92 after 48 h, and a further decrease to 3.84 at 72 h for HFOE. Interestingly, adding honey to the fermentation media accelerated the growth, reflected in the cell count and pH level.
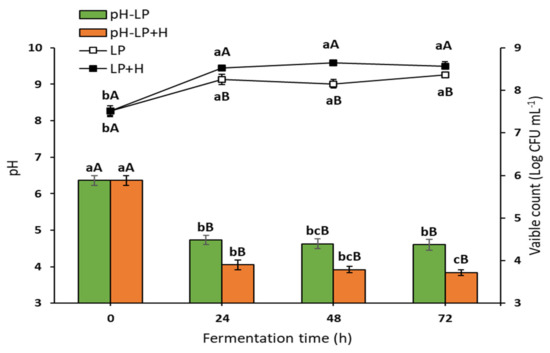
Figure 1.
Viability of L. plantarum (Log CFU mL−1) and pH value during fermentation (mean ± SE), n = 3. LP, L. plantarum count in plain media; LP + H, L. plantarum count in media with 2% Sidr honey; pH-LP, pH value of basic media during L. plantarum fermentation; pH-LP + H, pH value of media with honey during L. plantarum fermentation. a–c Bars or lines during the fermentation period not sharing similar letters are significantly different (p > 0.05). A,B Bars or lines of each treatment not sharing similar letters during the fermentation time are significantly different (p > 0.05).
3.2. Phytochemicals and Antioxidant Capacity of FOE and HFOE
The TPC and relative antioxidant activities using DPPH and ABTS assays of FOE and HFOE were investigated; data are presented in Table 2. TPC was significantly increased with the long fermentation time in both extracts. Interestingly, HFOE showed a significant increase in TPC, much more than FOE. Accordingly, the antioxidant capacity increased with the fermentation time in all subjected extracts; the highest values were recorded in HFOE after 72 h for both DPPH and ABTS radicals.
Table 2.
Total phenolic content (TPC) and potential antioxidant activities in FOE and HFOE during fermentation up to 72 h (mean ± SE), n = 8.
3.3. Gamma-Aminobutyric Acid and β-Glucan Contents during Fermentation
The gamma-aminobutyric acid (GABA) and β-glucan contents in FOE and HFOE were screened (Table 3). The GABA content increased significantly after 48 h to 6.10 ± 0.52 mg 100 g−1 and 6.54 ± 0.21 mg 100 g−1, in FOE and HFOE, respectively. After 72 h, GABA showed the most significant increase (7.35 ± 0.40 mg 100 g−1 in FOE and 8.49 ± 1.13 mg 100 g−1 in HFOE; p < 0.05). Furthermore, monitoring β-glucan as a bioactive component in oats during fermentation was of interest in the present study. There were no significant changes in the β-glucan content during the first 48 h of fermentation in FOE and HFOE. A slight, nonsignificant decrease in FOE and HFOE after 48 h was observed (2.56 ± 0.03 g 100 g−1 DW and 2.67 ± 0.01 g 100 g−1 DW, respectively). After 72 h, β-glucan content was 2.45 ± 0.06 g 100 g−1 DW, which differed significantly compared to its content in FOE during the period of 0–48 h. On the contrary, no significant difference was found in β-glucan content during fermentation of HFOE.
Table 3.
The γ-aminobutyric acid and β-glucan content in FOE and HFOE during fermentation up to 72 h (mean ± SE), n = 8.
3.4. The Hypoglycemic Efficiency of FOE and HFOE
The hypoglycemic efficiency of FOE and HFOE at 3.5 mL and 7 mL, and Metformin at 50 mg kg−1, on STZ-induced diabetic rats was monitored Table 4. STZ injection affected the rats’ RBG directly during the first week. The administration of all fermented oat extracts showed significant improvement in the RBG recorded in the third and the sixth weeks compared to the DR group, as shown in Table 4. The most efficient treatment for improving the rats’ RBG levels was the HFOE2. In addition, FBG measurement confirmed that FOE and HFOE impressively attenuated the levels close to normal rats (NR group). Interestingly, FOE2 and HFOE2 significantly attenuated the serum glucose levels, and there were no significant differences compared to the NR or Metformin groups confirming the potential hypoglycemic efficiency.
Table 4.
Effect of orally administrated FOE and HFOE on RBG and FBG (mg dL−1) in STZ-induced diabetes in rats (mean ± SE), n = 8.
3.5. The Hypolipidemic Efficiency of FOE and HFOE
The hypolipidemic efficiency of FOE and HFOE at 3.5 mL and 7 mL on the STZ-induced diabetic rats was determined and shown in Table 5. A significant increase in TG, CHO, LDL, and VLDL levels, while a significant decrease in HDL levels in the positive control group (DR) compared to the NR group, was noted (p < 0.05). The administration of all fermented oat extracts significantly attenuated the TG, CHO, LDL-CHO, and VLDL-CHO levels compared to the DR group. The high doses, FOE2 and HFOE2, significantly increased the HDL-CHO and decreased VLDL-CHO levels. Interestingly, the HFOE2 showed the most potent effects in improving serum lipids. Moreover, the AI was increased significantly after STZ injection (DR group) compared to the NR group, and the most efficient extracts in attenuating the atherogenicity complication were the high doses of both extracts (FOE2 and HFOE2), which present a superior effect to both the low doses and the Metformin treatment.
Table 5.
Effect of orally administrated FOE and HFOE on lipid profile (mg dL−1) and atherogenic index in STZ-induced diabetes in rats (mean ± SE), n = 8.
3.6. The Liver Functions
STZ injection substantially raised serum ALT, AST, and ALP enzyme levels in DR rats compared to the NR group (p < 0.05); in addition, the T. Bili levels were significantly increased in the DR group, as shown in Table 6. Administration of FOE or HFOE promoted hepatoprotective effects. The high doses exhibited much more effective results than using lower doses; FOE2 and HFOE2 treatments substantially reduced ALT, AST, and ALP to normal levels, even more effectively than Metformin.
Table 6.
Effect of orally administrated FOE and HFOE on liver function biomarkers in STZ-induced diabetes in rats (mean ± SE), n = 8.
3.7. The Kidney Functions
The nephroprotective efficiency of FOE1, FOE2, HFOE1, HFOE2, and Metformin at a dose of 50 mg kg−1 in the STZ-induced diabetic rats was investigated (Table 7). STZ injection substantially raised serum creatinine, urea, and BUN levels in DR rats compared to normal rats (DR). Contrarily, T. Protein, albumin, and globulin levels were significantly decreased. Treatments with FOE1, FOE2, HFOE1, and HFOE2 considerably improved the creatinine, urea, and BUN concentrations. Additionally, the T. Protein, albumin, and globulin levels were raised to normal levels, as seen in Table 7. The HFOE2 showed the most significant improvement in these levels compared to normal rats, even better than Metformin treatment.
Table 7.
Effect of orally administrated FOE and HFOE on kidney function biomarkers in STZ-induced diabetes in rats (mean ± SE), n = 8.
3.8. Antioxidant Biomarkers
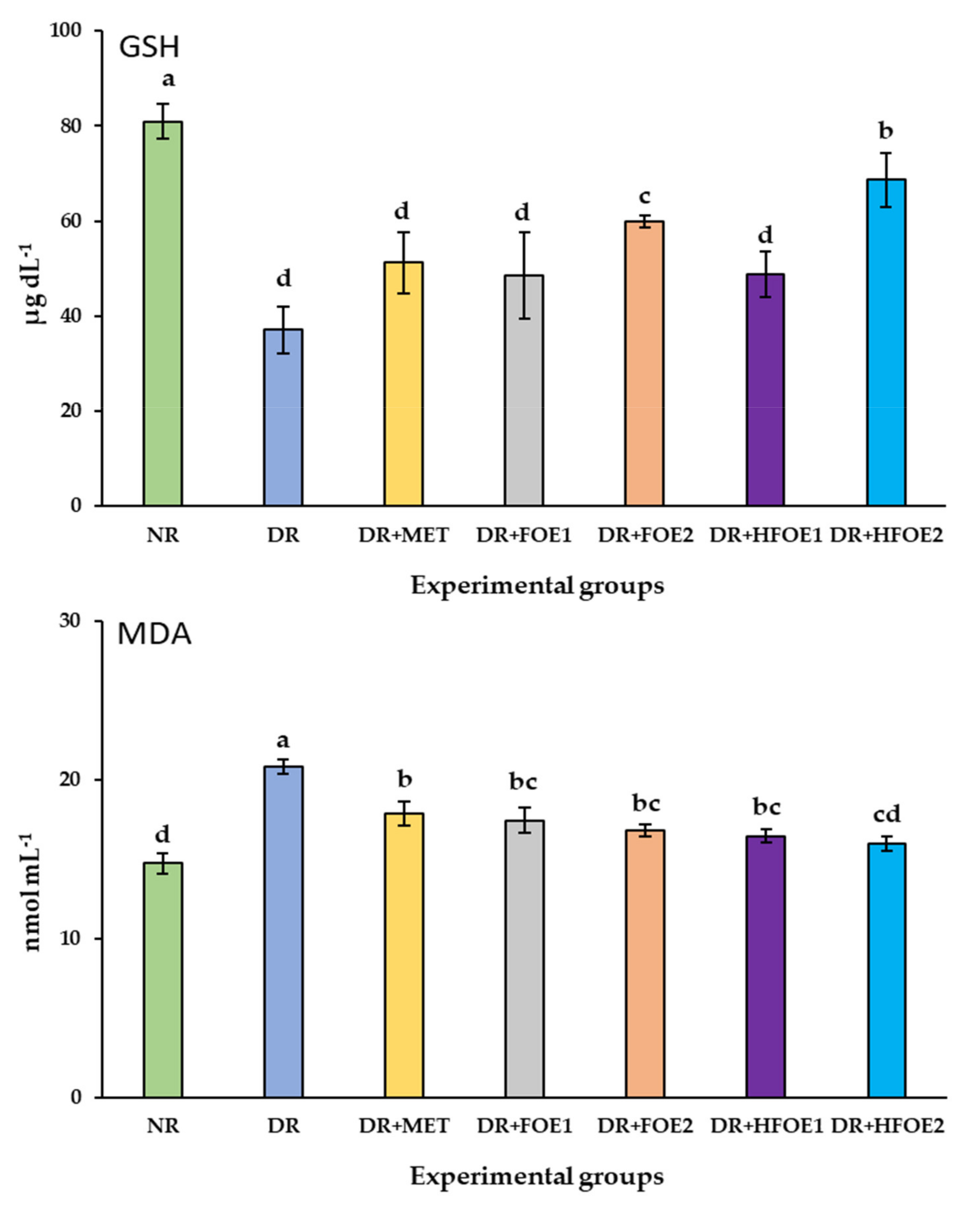
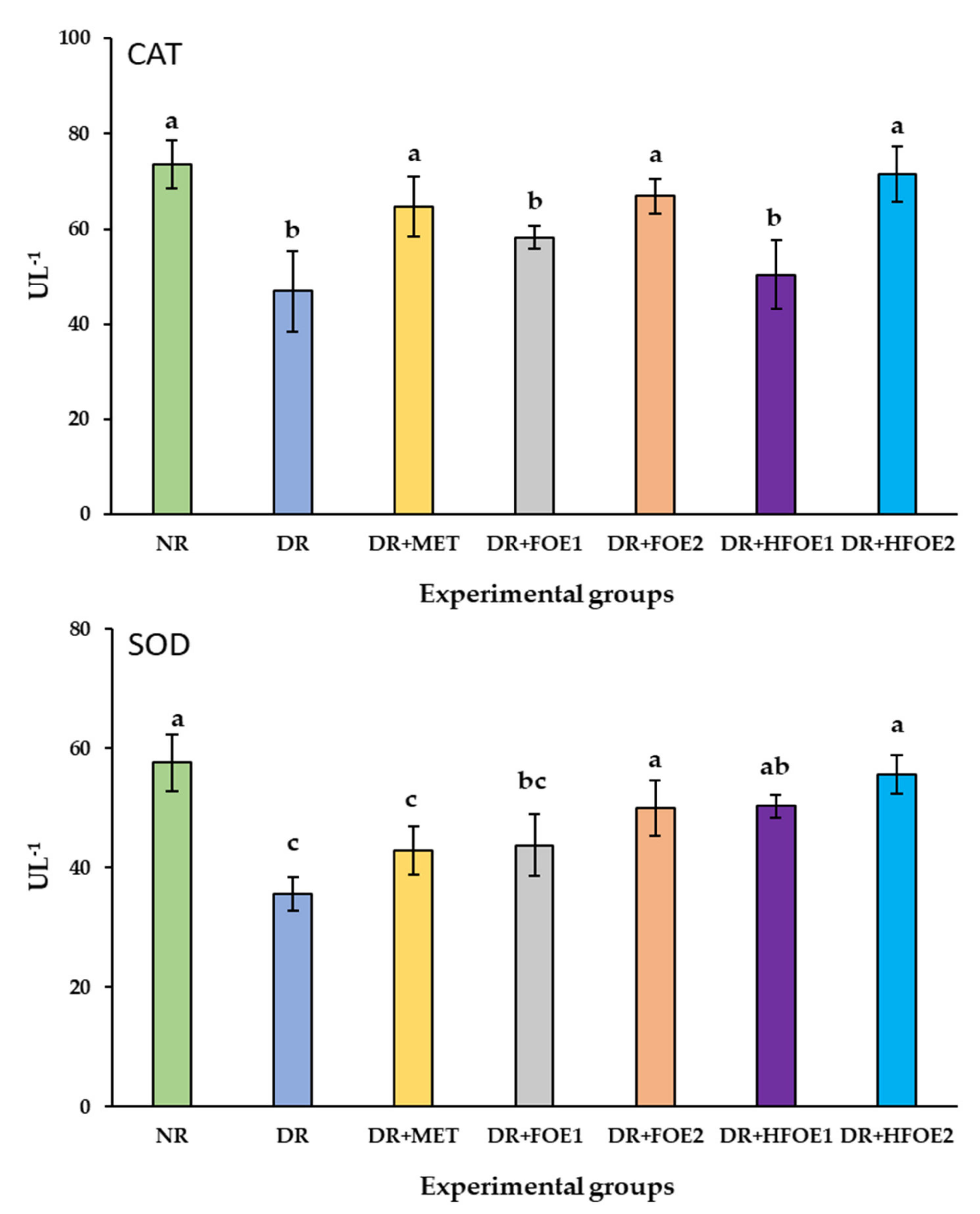
The effect of FOE1, FOE2, HFOE1, HFOE2, and Metformin on antioxidant biomarkers was evaluated. As shown in Figure 2, STZ injection decreased GSH, CAT, and SOD enzyme levels, while it increased MDA levels in DR rats’ blood serum compared to normal rats (NR) (p < 0.05). The administration of all treatments resulted in a considerable increase in the activity of antioxidant enzymes GSH, CAT, and SOD, and a significant decrease in MDA levels. Treatments with all fermented extracts improved the antioxidant biomarkers, and their levels were significantly improved in a dose-dependent manner. Interestingly, HFOE2 dramatically improved the enzymatic defense system compared to all DR rats.
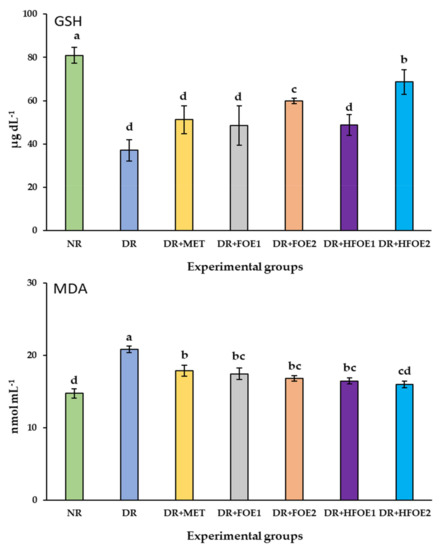
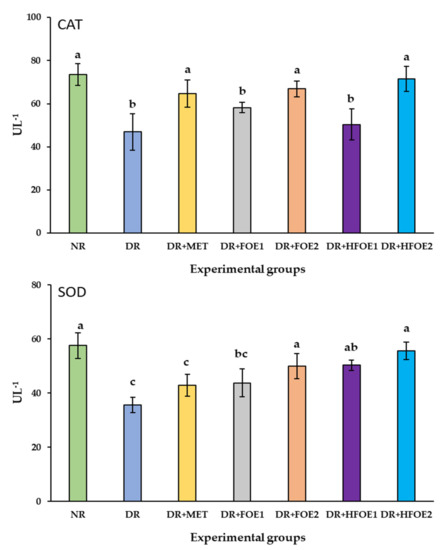
Figure 2.
Effect of orally administrated FOE and HFOE on antioxidant biomarkers in streptozotocin-induced diabetes in rats (mean ± SE), n = 8. Experimental groups, see materials and methods; Section 2.8. GSH, reduced glutathione; MDA, malondialdehyde; SOD, superoxide dismutase; CAT, catalase. a–d Bars not sharing similar letters are significantly different (p > 0.05).
4. Discussion
In recent years, cereal fermentations have been significantly demonstrated as promising functional products for improving or enhancing a foods’ nutritional quality. Several health-promoting effects of fermented products have been reported [10]. Lactic acid bacteria were reported as the most commonly used starter cultures in the food industry [21,22,41]. In the current work, aqueous extracts of L. plantarum-fermented oat and fermented oat supplemented with 2% Sidr honey were beneficial for managing diabetes complications in STZ-induced diabetic rats. The aqueous extracts were further subjected to multiple investigations to assess the changes in the nutritional quality.
Our results show that fermentation by L. plantarum significantly altered the composition of the oats’ aqueous extracts. There was a significant increase in total phenolic compounds and the related antioxidant capacity in a time- and type-dependent manner; the longer the fermentation time, the higher the release of total phenolic compounds. These results agree with multiple studies [11,42]; the long fermentation time (72 h) resulted in the most significant increase in total phenolic compounds. Biologically, phenolic compounds are considered as health-promoting agents [43,44,45,46]. Cereal fermentation can modify the presence of these compounds due to enzymatic activities derived from both the grains and the microbes [23]; enzymes such as glycosidase, hydrolase, esterase, and β-glucosidase produced by different types of bacterial strains can improve the availability of the grains’ phenolic compounds [11]. In addition, fermentation could increase the presence of other bioactive compounds, such as GABA [47], which is a non-protein amino acid that works as a neurotransmitter in the mammalian brain [48]; it has been demonstrated to play several roles in health-promoting activities, such as antihypertension and antidiabetic activities [49]. In the present study, we evaluated the effect of fermentation on the release of GABA; once again, the results were consistently time- and type-dependent. After 72 h of fermentation, we found a significant increase in GABA content by 78.3% and 92% in both FOE and HFOE, respectively. Previous studies reported similar results in fermented oats [50,51].
Interestingly, our results show that the GABA content in HFOE was much higher than that in FOE; such results could be attributed to the prebiotic effects of honey [52]. It was also indicated that GABA production could be affected by multiple factors, including fermentation time and microbe type [47], which supports our findings. The longer fermentation time (72 h) and the use of the L. plantarum strain significantly affected GABA production. The antidiabetic activities of GABA are attributed to its effects on promoting insulin secretion [49]. In the present in vivo investigation, the administration of FOE and HFOE, which were subjected to 72 h of fermentation, showed significant results in improving RBG and FBG. However, further investigation conducting complete GC–MS/MS and HPLC analyses is highly recommended to gain a better overview of the performance of such components during fermentation.
Moreover, oats’ β-glucans are documented as a potent antidiabetic agent [53,54,55]. However, previous studies indicated that β-glucan levels could be reduced during harsh processing [56]. In the interest of the present study, there was a slight decrease in the β-glucan content by 6.5% and 3.3% in FOE and HFOE, respectively, after 72 h of fermentation. Interestingly, a significant difference between both extracts in maintaining the content of β-glucan was observed, which is strongly attributed to the incorporation of honey [11]. Chen et al. [11] support this notion; the incorporation of honey in fermented oat-based beverages showed no significant decrease in the β-glucan content.
On the other hand, prolonged hyperglycemia is a primary cause of most diabetic complications. Indeed, chronic hyperglycemia leads to various metabolic impairments [57]. In the present study, after 6 weeks of oral administration of L. plantarum-fermented oat extracts, our results show significant hypoglycemic effects in STZ-induced diabetic rats in a dose- and type-dependent manner. The incorporation of honey in the fermented oat extract (HFOE2) strongly reduced the RBG and FBG levels compared to FOE1, FOE2, and HFOE1. These results could be related to the prebiotic properties of honey [52]. Moreover, honey was demonstrated to contain tremendous amounts of good-quality phytochemicals, such as flavonoids and phenolic acids, which could significantly improve glucose levels [58,59]. In addition, honey has antimicrobial, anti-inflammatory, and wound-healing properties that have been used in ancient and modern medical practice [60]. These beneficial effects of honey may partly explain the improvements in glycemic control of diabetic animals in this study [1,4]. Therefore, honey is an appropriate ingredient in functional products [61], which could support the functionality of prepared extracts.
Furthermore, imbalances in serum lipids were observed due to the injection of STZ, indicating impairment in fat metabolism due to diabetes complications [4]. The administration of FOE1, FOE2, HFOE1, and HFOE2 significantly enhanced the drastic changes in serum lipids. Interestingly, HFOE2 showed much more significant effects in improving TG, CHO, and LDL-cholesterol levels, which could also be related to the strong presence of phenols and β-glucan [11,61]. Oats were documented in several studies to exert therapeutic effects, improving blood glucose levels and CHO levels [1,62]. In the present study, the desirable results in strengthening hyperglycemia and serum lipid levels demonstrate that oat fermentation supplemented with excellent prebiotics such as honey may increase the potential therapeutic qualities of oats in the management of diabetes and the diabetes-related complications.
Furthermore, significant differences in liver and kidney diagnostic markers were observed in the treated groups compared to the positive control group. The significant increase in ALT, AST, ALP, and T. Bili enzyme activities in the positive control group due to the STZ injection represents normal deterioration associated with diabetic liver injury [44,46]. The intervention of all subjected extracts resulted in significant improvements in balancing the liver enzyme levels in a dose- and type-dependent manner. Bioactive compounds derived from the activity of probiotic strains during fermentation could explain these desirable hepatoprotective effects [61,63]. Probiotic supplementation could prevent hepatic steatosis by improving and treating metabolic disorders, balancing the lipid profile, and suppressing the inflammatory state [63,64,65]. Consistently, imbalances in kidney enzymes were observed due to significant hyperglycemia. The higher the blood glucose levels, the more damage to the kidney filtering units, resulting in kidney failure [66,67]. This has led to diabetes becoming one of the leading causes of end-stage kidney disease, also known as diabetic nephropathy [66]. In our investigation, the recovery of all biomarkers of kidney functions in the diabetic rats was observed after the intervention of all fermented oat extracts in a dose- and type-dependent manner.
Interestingly enough, an increase in T. Protein and albumin levels, concomitant with decreases in urea and BUN levels, were highly significant after administering HFOE2 compared to all other groups. In a previous study [66], kidney function parameters were recovered to a normal state after the intervention of oats [68]. Oat extract was related to a reduction in the liver’s glucose production and increased insulin secretion, which is associated with improved kidney functions [64,67]. On the other hand, the injection of STZ also resulted in significant imbalances in oxidative stress markers. A decrease in serum GSH, SOD, and CAT levels, and an increase in MDA levels, was observed. In hyperglycemia, various mechanisms generate free radicals, leading to the oxidation of lipids, proteins, and DNA, resulting in a significant state of oxidative stress [69]. The increase in free radicals reduces the production of biological antioxidative enzymes such as CAT, SOD, and GSH [70], making the tissues more vulnerable to oxidative stress [71]. The intervention of all fermented oat extracts in the present study recovered the altered activity of antioxidant enzymes in diabetic rats. Consistently, we found that HFOE2 was more effective than the other subjected extracts and even more effective than Metformin treatment at preventing oxidative stress. These effects are strongly related to a decrease in lipid peroxidation [71], which could be attributed to the combination of the antioxidant activities of oats, honey, and L. plantarum metabolites [58,66,71]. Therefore, our results support the hypothesis that a good-quality cereal-based symbiotic product possesses potent antidiabetic activities with antioxidative stress potential.
5. Conclusions
In this study, L. plantarum-fermented oat or fermented oat combined with Sidr honey exhibited potential antidiabetic effects, including ameliorative and antioxidative stress effects. We innovatively investigated the antidiabetic and antioxidative potential of FOE and HFOE in the form of aqueous extracts. Our results show a significant enhancement in the total phenolic compounds, the antioxidant capacity, and the gamma-aminobutyric acid in a type- and time-dependent manner. Fermentation up to 72 h and the incorporation of honey showed much more desirable effects. Moreover, honey proved to be a good prebiotic source, which was reflected in maintaining the oats’ β-glucan levels. Furthermore, the oral administration of HFOE for 6 weeks effectively improved diabetic complications in diabetic rats by improving blood glucose levels, lipidemia, liver and kidney functions, and oxidative stress markers. This study demonstrates that incorporating honey in L. plantarum-fermented oats has high potential as a symbiotic product for the management of diabetes. Future strategies for producing functional supplements based on these data are highly recommended.
Author Contributions
Conceptualization, R.A. and H.F.A.; methodology, R.A., H.F.A. and H.B.; investigation, R.A., H.F.A. and H.B.; Funding acquisition, R.A., H.F.A. and H.B.; data curation H.B.; Formal analysis, H.B.; writing—original draft preparation, H.F.A. and H.B.; review, and editing; R.A., H.F.A. and H.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was approved by the committee of Research Ethics, Deanship of Scientific Research, Qassim University (21-13-19 on 21 March 2022), SA, which is governed by the Control and Supervision of Experiments on Animals (CPCSEA) Committee of the National Committee of BioEthics (NCBE), which implements regulations related to the ethics of research on living creatures.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
The researchers would like to thank the Deanship of Scientific Research, Qassim University, for funding the publication of this project.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ABTS, 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid); ALT, Alanine aminotransferase; AST, Aspartate aminotransferase; CAT, Catalase; CHO, Total cholesterol; DPPH: 1,1-diphenyl-2-picryl hydrazine; DW: Dry weight; FBG, Fasting blood glucose; FOE, Fermented oat; GA: Gallic acid; GAE: Gallic acid equivalent; GSH: Reduced-glutathione; HDL, High-density lipoproteins; HFOE, honey Fermented oat; LDL, Low-density lipoproteins; MDA: Malonaldehyde; RSA: Radical scavenging activity; SE: Standard error; SOD: Superoxide dismutase; TBA, Thiobarbituric acid; TE: Trolox equivalents; TG, Triglycerides; TPC, Total phenolic content; VLDL, Very-Low-density lipoproteins.
References
- Wehrli, F.; Taneri, P.E.; Bano, A.; Bally, L.; Blekkenhorst, L.C.; Bussler, W.; Metzger, B.; Minder, B.; Glisic, M.; Muka, T.; et al. Oat Intake and Risk of Type 2 Diabetes, Cardiovascular Disease and All-Cause Mortality: A Systematic Review and Meta-Analysis. Nutrients 2021, 13, 2560. [Google Scholar] [CrossRef]
- Sangwan, S.; Singh, R.; Tomar, S.K. Nutritional and Functional Properties of Oats: An Update. J. Innov. Biol. 2014, 1, 3–14. [Google Scholar]
- Ahmad, M.; Dar, Z.A.; Habib, M. A Review on Oat (Avena sativa L.) as A Dual-Purpose Crop. Sci. Res. Essays 2014, 9, 52–59. [Google Scholar]
- Zhu, Y.; Dong, L.; Huang, L.; Shi, Z.; Dong, J.; Yao, Y.; Shen, R. Effects of Oat β-glucan, Oat Resistant Starch, and the Whole Oat Flour on Insulin Resistance, Inflammation, and Gut Microbiota in High-Fat-Diet-Induced Type 2 Diabetic Rats. J. Funct. Foods 2020, 69, 103939. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, X.; Dong, Y.; Shi, L.; Xu, T.; Wu, F. The Anti-obesity Effect of Fermented Barley Extracts with Lactobacillus plantarum dy-1 and Saccharomyces cerevisiae in Diet-Induced Obese Rats. Food Funct. 2017, 8, 1132–1143. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Prasanth, M.I.; Chaiyasut, C. A Mini Review on Antidiabetic Properties of Fermented Foods. Nutrients 2018, 10, 1973. [Google Scholar] [CrossRef] [Green Version]
- Boz, H. Phenolic Amides (Avenanthramides) in Oats—A review. Czech J. Food Sci. 2015, 33, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Sang, S.; Chu, Y. Whole Grain Oats, More than Just A Fiber: Role of Unique Phytochemicals. Mol. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.-B. Fermentation and Germination Improve Nutritional Value of Cereals and Legumes Through Activation of Endogenous Enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health Benefits of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, D.; Schlundt, J.; Conway, P.L. Development of a Dairy-Free Fermented Oat-Based Beverage With Enhanced Probiotic and Bioactive Properties. Front. Microbiol. 2020, 11, 609734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, W.; Xue, J.; Yang, J.; Chen, X.; Shao, Y.; Kwok, L.-Y.; Bilige, M.; Mang, L.; Zhang, H. Angiotensin-Converting Enzyme Inhibitory Activity of Lactobacillus helveticus Strains from Traditional Fermented Dairy Foods and Antihypertensive Effect of Fermented Milk of Strain H9. J. Dairy Sci. 2014, 97, 6680–6692. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Naramoto, K.; Koyama, M. Blood-Pressure-Lowering Effect of Fermented Buckwheat Sprouts in Spontaneously Hypertensive Rats. J. Funct. Foods 2013, 5, 406–415. [Google Scholar] [CrossRef]
- Pouliot-Mathieu, K.; Gardner-Fortier, C.; Lemieux, S.; St-Gelais, D.; Champagne, C.P.; Vuillemard, J.-C. Effect of Cheese Containing Gamma-aminobutyric Acid-producing Lactic Acid Bacteria on Blood Pressure in Men. PharmaNutrition 2013, 1, 141–148. [Google Scholar] [CrossRef]
- Yoshinaga, M.; Toda, N.; Tamura, Y.; Terakado, S.; Ueno, M.; Otsuka, K.; Numabe, A.; Kawabata, Y.; Uehara, Y. Japanese Traditional Miso Soup Attenuates Salt-Induced Hypertension and its Organ Damage in Dahl Salt-Sensitive Rats. Nutrition 2012, 28, 924–931. [Google Scholar] [CrossRef]
- Guan, Q.; Ding, X.-W.; Zhong, L.-Y.; Zhu, C.; Nie, P.; Song, L.-H. Beneficial Efects of Lactobacillus-fermented Black Barley on High Fat Diet-Induced Fatty Liver in Rats. Food Funct. 2021, 12, 6526–6539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiao, X.; Dong, Y.; Zhou, X. Fermented Barley Extracts with Lactobacillus plantarum dy-1 Changes Serum Metabolomic Profiles in Rats with High-Fat Diet-Induced Obesity. Int. J. Food Sci. Nutr. 2019, 70, 303–310. [Google Scholar] [CrossRef]
- Lee, C.S.; Kim, B.K.; Lee, I.O.; Park, N.H.; Kim, S.H. Prevention of Done Loss by Using Lactobacillus-Fermented Milk Products in A Rat Model of Glucocorticoid-Induced Secondary Osteoporosis. Int. Dairy J. 2020, 109, 104788. [Google Scholar] [CrossRef]
- Robert, A.A.; Al Dawish, A.M. The Worrying Trend of Diabetes Mellitus in Saudi Arabia: An Urgent Call to Action. Curr. Diabetes Rev. 2020, 16, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Vries, M.; Vaughan, E.; Kleerebezem, M.; De Vos, W. Lactobacillus plantarum-Survival, Functional and Potential Probiotic Properties in the Human Intestinal Tract. Int. Dairy J. 2006, 16, 1018–1028. [Google Scholar] [CrossRef]
- Rathore, S.; Salmerón, I.; Pandiella, S.S. Production of Potentially Probiotic Beverages Using Single and Mixed Cereal Substrates Fermented with Lactic acid Bacteria Cultures. Food Microbiol. 2012, 30, 239–244. [Google Scholar] [CrossRef]
- Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved Bioavailability of Dietary Phenolic Acids in Whole Grain Barley and Oat Groat following Fermentation with Probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [Google Scholar] [CrossRef]
- Dordević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of Fermentation on Antioxidant Properties of some Cereals and Pseudo Cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- International Diabetes Federation. IDF Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2021; Volume 102, pp. 147–148. [Google Scholar]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.-F. Global, Regional, and National Durden and Trend of Diabetes in 195 Countries and Territories: An Analysis from 1990 to 2025. Sci. Rep. 2020, 10, 1–11. [Google Scholar]
- CDC. United States Department of Health and Human Services. 2019. Available online: https://www.cdc.gov/ (accessed on 16 May 2022).
- Aldossari, K.K.; Aldiab, A.; Al-Zahrani, J.M.; Al-Ghamdi, S.H.; Abdelrazik, M.; Batais, M.A.; Javad, S.; Nooruddin, S.; Razzak, H.A.; El-Metwally, A. Prevalence of Prediabetes, Diabetes, and Its Associated Risk Factors among Males in Saudi Arabia: A Population-Based Survey. J. Diabetes Res. 2018, 2018, 2194604. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Reinheimer, J.A. Culture media for the Enumeration of Bifidobacterium bifidum and Lactobacillus acidophilus in the Presence of Yoghurt Bacteria. Int. Dairy J. 1999, 9, 497–505. [Google Scholar] [CrossRef]
- Yawadio Nsimba, R.; Kikuzaki, H.; Konishi, Y. Antioxidant activity of Various Extracts and Fractions of Chenopodium quinoa and Amaranthus spp. Seeds. Food Chem. 2008, 106, 760–766. [Google Scholar] [CrossRef]
- Lu, J.; Zhao, H.; Chen, J.; Fan, W.; Dong, J.; Kong, W.; Sun, J.; Cao, Y.; Cai, G. Evolution of Phenolic Compounds and Antioxidant Activity During Malting. J. Agric. Food Chem. 2007, 55, 10994–11001. [Google Scholar] [CrossRef]
- Yeap, S.K.; Mohd Ali, N.; Mohd Yusof, H.; Alitheen, N.B.; Beh, B.K.; Ho, W.Y.; Koh, S.P.; Long, K. Antihyperglycemic Effects of Fermented and Nonfermented Mung Bean Extracts on Alloxan-Induced-Diabetic Mice. J. Biotechnol. Biomed. 2012, 2012, 285430. [Google Scholar] [CrossRef] [Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition ad hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Jia, Y.; Lao, Y.; Zhu, H.; Li, N.; Leung, S.W. Is Metformin Still the Most Efficacious First-Line Oral Hypoglycaemic Drug in Treating Type 2 Diabetes? A Network Meta-Analysis of Randomized Controlled Trials. Obes. Rev. 2019, 20, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-density Lipoprotein Cholesterol in Plasma, without use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Nwagha, U.; Ikekpeazu, E.; Ejezie, F.; Neboh, E.; Maduka, I. Atherogenic Index of Plasma as Useful Predictor of Cardiovascular Risk Among Postmenopausal Women in Enugu, Nigeria. Afr. Health Sci. 2010, 10, 248–252. [Google Scholar] [PubMed]
- Beutler, E. Improved Method for the Determination of Blood Glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Steel, R.G. Pinciples and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw-Hill: Boston, MA, USA, 1997. [Google Scholar]
- Mubarak, A.; Hodgson, J.M.; Considine, M.J.; Croft, K.D.; Matthews, V.B. Supplementation of a High-Fat Diet with Chlorogenic Acid Is Associated with Insulin Resistance and Hepatic Lipid Accumulation in Mice. J. Agric. Food Chem. 2013, 61, 4371–4378. [Google Scholar] [CrossRef] [PubMed]
- Balli, D.; Bellumori, M.; Pucci, L.; Gabriele, M.; Longo, V.; Paoli, P.; Melani, F.; Mulinacci, N.; Innocenti, M. Does Fermentation Really Increase the Phenolic Content in Cereals? A Study on Millet. Foods 2020, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Al-Qabba, M.M.; El-Mowafy, M.A.; Althwab, S.A.; Alfheeaid, H.A.; Aljutaily, T.; Barakat, H. Phenolic Profile, Antioxidant Activity, and Ameliorating Efficacy of Chenopodium quinoa Sprouts against CCl4-Induced Oxidative Stress in Rats. Nutrients 2020, 12, 2904. [Google Scholar] [CrossRef]
- Hassan, B.; Tariq, I.A. Phenolic Compounds and Hepatoprotective Potential of Anastatica hierochuntica Ethanolic and Aqueous Extracts Against CCl4-induced Hepatotoxicity in Rats. J. Tradit. Chin. Med. 2020, 40, 947–955. [Google Scholar] [CrossRef]
- Almundarij, T.I.; Alharbi, Y.M.; Abdel-Rahman, H.A.; Barakat, H. Antioxidant Activity, Phenolic Profile, and Nephroprotective Potential of Anastatica hierochuntica Ethanolic and Aqueous Extracts against CCl4-Induced Nephrotoxicity in Rats. Nutrients 2021, 13, 2973. [Google Scholar] [CrossRef]
- Alharbi, Y.M.; Sakr, S.S.; Albarrak, S.M.; Almundarij, T.I.; Barakat, H.; Hassan, M.F.Y. Antioxidative, Antidiabetic, and Hypolipidemic Properties of Probiotic-Enriched Fermented Camel Milk Combined with Salvia officinalis Leaves Hydroalcoholic Extract in Streptozotocin-Induced Diabetes in Rats. Antioxidants 2022, 11, 668. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of GABA (γ-aminobutyric acid) by Microorganisms: A Review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [Green Version]
- Mohd Ali, N.; Mohd Yusof, H.; Long, K.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Koh, S.P.; Abdullah, M.P.; Alitheen, N.B. Antioxidant and Hepatoprotective Effect of Aqueous Extract of Germinated and Fermented Mung Bean on Ethanol-Mediated Liver Damage. Biomed. Res. Int. 2013, 2013, 693613. [Google Scholar] [CrossRef] [PubMed]
- Soltani, N.; Qiu, H.; Aleksic, M.; Glinka, Y.; Zhao, F.; Liu, R.; Li, Y.; Zhang, N.; Chakrabarti, R.; Ng, T. GABA Exerts Protective and Regenerative Effects on Islet Beta Cells and Reverses Diabetes. Proc. Natl. Acad. Sci. USA 2011, 108, 11692–11697. [Google Scholar] [CrossRef] [Green Version]
- Coda, R.; Rizzello, C.G.; Gobbetti, M. Use of Sourdough Fermentation and Pseudo-Cereals and Leguminous Flours for the Making of A Functional Bread Enriched of γ-aminobutyric Acid (GABA). Int. J. Food Microbiol. 2010, 137, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Gao, F.; Zhang, X.; Wang, O.; Wu, W.; Zhu, S.; Zhang, D.; Zhou, F.; Ji, B. Evaluation of γ-Aminobutyric Acid, Phytate and Antioxidant Activity of Tempeh-Like Fermented Oats (Avena sativa L.) Prepared with Different Filamentous Fungi. J. Food Sci. Technol. 2014, 51, 2544–2551. [Google Scholar] [CrossRef] [Green Version]
- Miguel, M.; Antunes, M.D.; Faleiro, M.L. Honey As A Complementary Medicine. Integr. Med. Insights 2017, 12, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, E.F.; Lobato, R.V.; de Araújo, T.V.; Zangerônimo, M.G.; de Sousa, R.V.; Pereira, L.J. Effect of Beta-Glucans in the Control of Blood Glucose Levels of Diabetic Patients: A Systematic Review. Nutr. Hosp. 2015, 31, 170–177. [Google Scholar]
- Bozbulut, R.; Sanlier, N. Promising Effects of β-glucans on Glyceamic Control in Diabetes. Trends Food Sci. Technol. 2019, 83, 159–166. [Google Scholar] [CrossRef]
- Chen, J.; Raymond, K. Beta-Glucans in the Treatment of Diabetes and Associated Cardiovascular Risks. Vasc. Health Risk Manag. 2008, 4, 1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Du, B.; Xu, B. A Critical Review on Production and Industrial Applications of Beta-Glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 8267234. [Google Scholar] [CrossRef] [Green Version]
- Hossen, M.S.; Ali, M.Y.; Jahurul, M.H.A.; Abdel-Daim, M.M.; Gan, S.H.; Khalil, M.I. Beneficial Roles of Honey Polyphenols Against some Human Degenerative Diseases: A Review. Pharmacol. Rep. 2017, 69, 1194–1205. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Bravo Lamas, L.; Martínez Flórez, S.; Agudo Toyos, P. Phenolic Compounds in Honey and their Associated Health Benefits: A Review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef] [Green Version]
- Al-Waili, N.S.; Salom, K.; Butler, G.; Al Ghamdi, A.A. Honey and Microbial Infections: A Review Supporting the Use of Honey for Microbial Control. J. Med. Food 2011, 14, 1079–1096. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Datta, S.; Mukherjee, S.; Bose, S.; Ghosh, S.; Dhar, P. Evaluation of Antioxidative, Antibacterial and Probiotic Growth Stimulatory Activities of Sesamum indicum Honey Containing Phenolic Compounds and Lignans. LWT Food Sci. Technol. 2015, 61, 244–250. [Google Scholar] [CrossRef]
- Hou, Q.; Li, Y.; Li, L.; Cheng, G.; Sun, X.; Li, S.; Tian, H. The Metabolic Effects of Oats Intake in Patients with Type 2 Diabetes: A Systematic Review and Meta-analysis. Nutrients 2015, 7, 10369–10387. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-C.; Huang, C.-N.; Yeh, D.-M.; Wang, S.-J.; Peng, C.-H.; Wang, C.-J. Oat Prevents Obesity and Abdominal Fat Distribution, and Improves Liver Function in Humans. Plant Foods Hum. Nutr. 2013, 68, 18–23. [Google Scholar] [CrossRef]
- Li, C.; Nie, S.-P.; Zhu, K.-X.; Ding, Q.; Li, C.; Xiong, T.; Xie, M.-Y. Lactobacillus plantarum NCU116 Improves Liver Function, Oxidative Stress and Lipid Metabolism in Rats with High Fat Diet Induced Non-Alcoholic Fatty Liver Disease. Food Funct. 2014, 5, 3216–3223. [Google Scholar] [CrossRef]
- Huang, L.; Zhao, Z.; Duan, C.; Wang, C.; Zhao, Y.; Yang, G.; Gao, L.; Niu, C.; Xu, J.; Li, S. Lactobacillus plantarum C88 Protects Against Aflatoxin B(1)-induced Liver Injury in Mice via Inhibition of NF-κB-Mediated Inflammatory Responses and Excessive Apoptosis. BMC Microbiol. 2019, 19, 170. [Google Scholar] [CrossRef] [Green Version]
- Al-Waili, N.; Al-Waili, H.; Al-Waili, T.; Salom, K. Natural Antioxidants in the Treatment and Prevention of Diabetic Nephropathy; A Potential Approach that Warrants Clinical Trials. Redox Rep. 2017, 22, 99–118. [Google Scholar] [CrossRef]
- Pecoits-Filho, R.; Abensur, H.; Betônico, C.C.R.; Machado, A.D.; Parente, E.B.; Queiroz, M.; Salles, J.E.N.; Titan, S.; Vencio, S. Interactions Between Kidney Disease and Diabetes: Dangerous Liaisons. Diabetol. Metab. Syndr. 2016, 8, 50. [Google Scholar] [CrossRef]
- Liu, S.; Yin, X.; Hou, C.; Liu, X.; Ma, H.; Zhang, X.; Xu, M.; Xie, Y.; Li, Y.; Wang, J. As a Staple Food Substitute, Oat and Buckwheat Compound Has Health-Promoting Effects for Diabetic Rats. Front. Nutr. 2021, 8, 762277. [Google Scholar] [CrossRef]
- Bokhary, K.; Aljaser, F.; Abudawood, M.; Tabassum, H.; Bakhsh, A.; Alhammad, S.; Aleyadhi, R.; Almajed, F.; Alsubki, R. Role of Oxidative Stress and Severity of Diabetic Retinopathy in Type 1 and Type 2 Diabetes. Ophthalmic Res. 2021, 64, 613–621. [Google Scholar] [CrossRef]
- Das, R.R.; Rahman, M.A.; Al-Araby, S.Q.; Islam, M.S.; Rashid, M.M.; Babteen, N.A.; Alnajeebi, A.M.; Alharbi, H.F.H.; Jeandet, P.; Rafi, M.K.J.; et al. The Antioxidative Role of Natural Compounds from a Green Coconut Mesocarp Undeniably Contributes to Control Diabetic Complications as Evidenced by the Associated Genes and Biochemical Indexes. Oxidative Med. Cell. Longev. 2021, 2021, 060803. [Google Scholar] [CrossRef]
- Al-Malki, A.L. Oat Protects Against Diabetic Nephropathy in Rats via Attenuating Advanced Glycation End Products and Nuclear Factor Kappa B. Evid. Based Complement. Altern. Med. 2013, 2013, 609745. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).