
Effect of Stroking on Serotonin, Noradrenaline, and Cortisol Levels in the Blood of Right- and Left-Pawed Dogs

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Animals and Procedures
2.2. Behavioral Tests—Paw Preference Tests
2.2.1. Kong Test
2.2.2. Paw Preference Index (z)
2.3. Blood Analysis—Determination of the Neurotransmitters Level
2.4. Statistical Analysis
3. Results
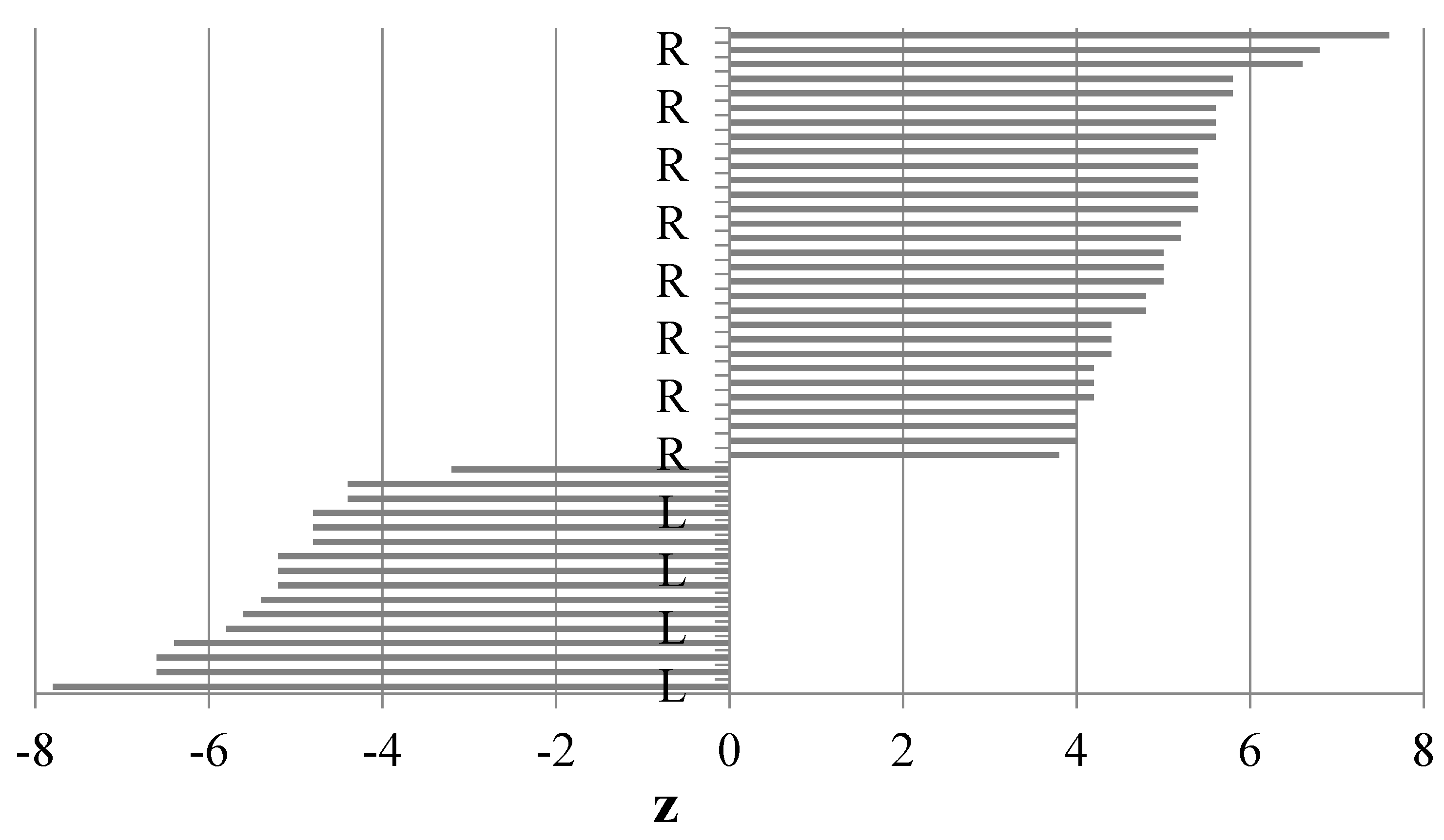
3.1. Paw Preference Index (z)
3.2. Biochemical Blood Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Overall, K.L.; Hamilton, S.P.; Chang, M.L. Understanding the genetic basis of canine anxiety: Phenotyping dogs for behavioral, neurochemical, and genetic assessment. J. Vet. Behav. 2006, 1, 124–141. [Google Scholar] [CrossRef]
- Peremans, K.; Goethals, I.; De vos, F.; Dobbeleir, A.; Ham, H.; Van bree, H.; Van heeringen, C.; Audenaert, K. Serotonin transporter and dopamine transporter imaging in the canine brain. Nucl. Med. Biol. 2006, 33, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Vermeire, S.T.; Audenaert, K.R.; Dobbeleir, A.A.; De meester, R.H.; De vos, F.J.; Peremans, K.Y. Evaluation of the brain 5-HT2A receptor binding index in dogs with anxiety disorders, measured with 123I-5I-R91150 and SPECT. J. Nucl. Med. 2009, 50, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Goleman, M.; Obłodecka, A.; Krupa, W.; Rozempolska-Rucińska, I. Analogies in behavioral and emotional disorders between dogs and humans. Med. Weter. 2019, 75, 146–151. [Google Scholar] [CrossRef]
- Howell., T.J.; King, T.; Bennett, P.C. Puppy parties and beyond: The role of early age socialization practices on adult dog behawior. J. Vet. Med. Res. Rep. 2015, 6, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Leliveld, L.M.C.; Langbein, J.; Puppe, B. The emergence of emotional lateralization: Evidence in non-human vertebrates and implications for farm animals. Appl. Anim. Behav. Sci. 2013, 145, 1–14. [Google Scholar] [CrossRef]
- Barnard, S.; Matthews, L.; Messori, S.; Podaliri-Vulpiani, M.; Ferri, N. Laterality as an indicator of emotional stress in ewes and lambs during a separation test. Anim. Cogn. 2016, 19, 207–214. [Google Scholar] [CrossRef]
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left-right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef]
- MacNeilage, P.; Rogers, L.J.; Vallortigara, G. Origins of the left and right brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar] [CrossRef]
- Vallortigara, G.; Chiandetti, C.; Sovrano, V.A. Brain asymmetry (animal). Wiley Interdiscip. Rev. Cogn. Sci. 2010, 2, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Coppola, C.L.; Grandin, T.; Enns, R.M. Human interaction and cortisol: Can human contact reduce stress for shelter dogs? Physiol. Behav. 2006, 87, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Csoltova, E.; Martineau, M.; Boissy, A.; Gilbert, C. Behavioral and physiological reactions in dogs to a veterinary examination: Owner-dog interactions improve canine well-being. Physiol. Behav. 2017, 177, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Buttner, A.P. Neurobiological underpinnings of dogs’ human-like social competence: How interactions between stress response systems and oxytocin mediate dogs’ social skills. Neurosci Biobehav. Rev. 2016, 71, 198–214. [Google Scholar] [CrossRef] [PubMed]
- Serpell, J. Factors influencing human attitudes to animal and their welfare. Anim. Welf. 2004, 13, 145–151. [Google Scholar]
- Stoeckel, L.E.; Palley, L.S.; Gollub, R.L.; Niemi, S.M.; Evins, A.E. Patterns of brain activation when mothers view their own child and dog: An fMRI study. PLoS ONE 2014, 9, e107205. [Google Scholar] [CrossRef]
- Nagasawa, M.; Mitsui, S.; En, S.; Ohtani, N.; Ohta, M.; Sakuma, Y.; Onaka, Y.; Mogi, K.; Kikusui, T. Social evolution. Oxytocin-gaze positive loop and the coevolution of human-dog bonds. Science 2015, 348, 333–336. [Google Scholar] [CrossRef]
- Kikusui, T.; Nagasawa, M.; Nomoto, K.; Kuse-Arata, S.; Mogi, K. Endocrine Regulations in Human-Dog Coexistence through Domestication. Trends Endocrinol. Metab. 2019, 30, 793–806. [Google Scholar] [CrossRef]
- Mariti, C.; Raspanti, E.; Zilocchi, M.; Carlone, B.; Gazzano, A. The assessment of dog welfare in the waiting room of a veterinary clinic. Anim Welf. 2015, 24, 299–305. [Google Scholar] [CrossRef]
- Ogi, A.; Mariti, C.; Baragli, P.; Sergi, V.; Gazzano, A. Effects of stroking on salivary oxytocin and cortisol in guide dogs: Preliminary results. Animals 2020, 10, 708. [Google Scholar] [CrossRef]
- McGreevy, P.D.; Brueckner, A.; Thomson, P.C.; Branson, N.J. Motor laterality in 4 breeds of dogs. J. Vet. Behav. 2010, 5, 318–323. [Google Scholar] [CrossRef]
- Quaranta, A.; Siniscalchi, M.; Frate, A.; Vallortigara, G. Paw preference in dogs: Relations between lateralised behaviour and immunity. Behav. Brain Res. 2004, 153, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.L. Lateralised behaviour in the domestic dog, Canis Fam. Behav. Proc. 2003, 61, 27–35. [Google Scholar] [CrossRef]
- Alves, N.T.; Fukusim, S.S.; Aznar-Casanova, J.A. Models of brain asymmetry in emotional processing. Psychol Neurosci. 2008, 1, 63–66. [Google Scholar] [CrossRef]
- Shobe, E.R. Independent and collaborative contributions of the cerebral hemispheres to emotional processing. Front. Hum. Neurosci. 2014, 8, 230. [Google Scholar] [CrossRef]
- Harmon-Jones, E.; Gable, P.A.; Peterson, C.K. The role of asymmetric frontal cortical activity in emotion-related phenomena: A review and update. Biol. Psychol. 2010, 84, 451–462. [Google Scholar] [CrossRef]
- Siniscalchi, M.; Lusito, R.; Vallortigara, G.; Quaranta, A. Seeing left- or right-asymmetric tail wagging produces different emotional responses in dogs. Curr. Biol. 2013, 23, 2270–2282. [Google Scholar] [CrossRef]
- Field, T.; Hernandez-Reif, M.; Diego, M.; Schanberg, S.; Kuhn, C. Cortisol decreases and serotonin and dopamine increase following massage therapy. Int. J. Neurosci. 2005, 115, 1397–1413. [Google Scholar] [CrossRef]
- Okabe, S.; Takayanagi, Y.; Yoshida, M.; Onaka, T. Gentle stroking stimuli induce affiliative responsiveness to humans in male rats. Sci. Rep. 2020, 10, 9135. [Google Scholar] [CrossRef]
- Willen, R.M.; Mutwill, A.; MacDonald, L.J.; Schiml, P.A.; Hennessy, M.B. Factors determining the effects of human interaction on the cortisol levels of shelter dogs. Appl. Anim. Behav. Sci. 2017, 186, 41–48. [Google Scholar] [CrossRef]
- DeVries, A.C.; Glasper, E.R.; Detillion, C.E. Social modulation of stress responses. Physiol. Behav. 2003, 79, 399–407. [Google Scholar] [CrossRef]
- Mariti, C.; Carlone, B.; Protti, M.; Diverio, S.; Gazzano, A. Effects of petting before a brief separation from the owner on dog behavior and physiology: A pilot study. J. Vet. Behav. 2018, 27, 41–46. [Google Scholar] [CrossRef]
- Lewandowski, G.W.; Mattingly, B.A.; Pedreiro, A. Under pressure: The effects of stress on positive and negative relationship behaviors. J. Soc. Psychol. 2014, 154, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Pirrone, F.; Ripamonti, A.; Garoni, E.C.; Stradiotti, S.; Albertini, M. Measuring social synchrony and stress in the handler-dog dyad during animal-assisted activities: A pilot study. J. Vet. Behav. Clin. Appl. Res. 2017, 21, 45–52. [Google Scholar] [CrossRef]
- Butler, R.K.; Oliver, E.M.; Sharko, A.C.; Parilla-Carrero, J.; Kaigler, K.F.; Fadel, J.R.; Wilson, M.A. Activation of corticotropin releasing factor-containing neurons in the rat central amygdala and bed nucleus of the stria terminalis following exposure to two different anxiogenic stressors. Behav. Brain Res. 2016, 304, 92–101. [Google Scholar] [CrossRef][Green Version]
- Major, B.; Rattazzi, L.; Brod, S.; Pilipović, I.; Leposavić, G.; D’Acquisto, F. Massage-like stroking boosts the immune system in mice. Sci. Rep. 2015, 5, 109–113. [Google Scholar] [CrossRef]
- Rosado, B.; García-Belenguer, S.; León, M.; Chacón, G.; Villegas, A.; Palacio, J. Blood concentrations of serotonin, cortisol and dehydroepiandrosterone in aggressive dogs. Appl. Anim. Behav. Sci. 2010, 123, 124–130. [Google Scholar] [CrossRef]
- Wells, D.L.; Hepper, P.G.; Milligan, A.D.S.; Barnard, S. Cognitive bias and paw preference in the domestic dog. Canis Fam. J. Comp. Psychol. 2017, 131, 317–325. [Google Scholar] [CrossRef]
- Winnicka, A. Reference Values of Basic Laboratory Tests in Veterinary Medicine; SGGW: Warsaw, Poland, 2015; p. 118. (In Polish) [Google Scholar]
- Batt, L.; Batt, M.; McGreevy, P. Two tests for motor laterality in dogs. J. Vet. Behav. 2007, 2, 47–51. [Google Scholar] [CrossRef]
- Batt, L.S.; Batt, M.S.; Baguley, J.A.; McGreevy, P.D. Factors associated with success in guide dog training. J. Vet. Behav. 2008, 3, 143–151. [Google Scholar] [CrossRef]
- Marshall-Pescini, S.; Barnard, S.; Branson, N.J.; Valsecchi, P. The effect of preferential paw usage on dogs’ (Canis familiaris) performance in a manipulative problem-solving task. Behav. Proc. 2013, 100, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Plueckhahn, T.; Schneider, L.A.; Delfrabbo, P.H. Assessing lateralization in domestic dogs: Performance by Canis familiaris on the Kong test. J. Vet. Behav. 2016, 15, 25–30. [Google Scholar] [CrossRef]
- Wells, D.L.; Hepper, P.G.; Milligan, A.D.S.; Barnard, S. Stability of motor bias in the domestic dog. Canis Fam. Behav. Process. 2018, 149, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Beerda, B.; Schilder, M.B.; van Hooff, J.A.; DeVries, H.W.; Mol, J.A. Behavioural and hormonal indicators of enduring environmental stress in dogs. Anim. Welf. 2000, 9, 49–62. [Google Scholar]
- Hekman, J.; Karas, A.; Dreschel, N. Salivary cortisol concentrations and behavior in a population of healthy dogs hospitalized for elective procedures. Appl. Anim. Behav. Sci. 2012, 141, 149–157. [Google Scholar] [CrossRef]
- Cozzi, A.; Mariti, C.; Ogi, A.; Sighieri, C.; Gazzano, A. Behavioral modification in sheltered dogs. Dog Behav. 2016, 2, 1–12. [Google Scholar] [CrossRef]
- Chen, Y.K.; Richter, H.M., 3rd; Go, V.L.; Tyce, G.M. Free and conjugated catecholamines and serotonin in canine thoracic duct lymph: Effects of feeding. Am. J. Physiol. 1993, 265, 84–89. [Google Scholar] [CrossRef]
- Ferrara, A.; Zinner, M.J.; Jaffe, B.M. Intraluminal release of serotonin, substance P, and gastrin in the canine small intestines. Dig. Dis. Sci. 1987, 32, 289–294. [Google Scholar] [CrossRef]
- LaRosa, C.A.; Sherlock, D.; Kimura, K.; Pimpl, W.; Money, S.R.; Jaffe, B.M. The role of serotonin in the canine secretory response to cholera toxin in vivo. J. Pharmacol. Exp. Ther. 1989, 251, 71–76. [Google Scholar]
- Fukuwatari, T.; Shibata, K. Nutritional Aspect of Tryptophan Metabolism. Int. J. Tryptophan Res. 2013, 6, 3–8. [Google Scholar] [CrossRef]
- Hart, E.C.; Charkoudian, N.; Miller, V.M. Sex, Hormones and Neuroeffector Mechanisms. Acta Physiol. 2011, 203, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Berman, M.E.; Tracy, J.I.; Coccaro, E.F. The serotonin hypothesis of aggression revisited. Clin. Psychol. Rev. 1997, 17, 651–665. [Google Scholar] [CrossRef]
- Çakiroglu, D.; Meral, Y.; Sancak, A.A.; Cifti, G. Relationship between the serum concentrations of serotonin and lipids and aggression in dogs. Vet. Rec. 2007, 161, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Diego, M.A.; Field, T. Moderate pressure massage elicits a parasympathetic nervous system response. Int. J. Neurosci. 2009, 119, 630–638. [Google Scholar] [CrossRef]
- Field, T.; Diego, M. Vagal activity, early growth and emotional development. Infant Behav. 2008, 361–373. [Google Scholar] [CrossRef]
- Pilipović, I.; Radojević, K.; Perišić, M.; Kosec, D.; Nacka-Aleksić, M.; Djikić, J.; Leposavić, G. Catecholaminergic signalling through thymic nerve fibres, thymocytes and stromal cells is dependent on both circulating and locally synthesized glucocorticoids. Exp. Physiol. 2012, 97, 1211–1223. [Google Scholar] [CrossRef]
- Radojević, K.; Rakin, A.; Pilipović, I.; Kosec, D.; Djikić, J.; Bufan, B.; Vujnović, I.; Leposavić, G. Effects of catecholamines on thymocyte apoptosis and proliferation depend on thymocyte microenvironment. J. Neuroimmunol. 2014, 272, 16–28. [Google Scholar] [CrossRef]
- Leposavić, G.; Pilipović, I.; Radojević, K.; Pesić, V.; Perisić, M.; Kosec, D. Catecholamines as immunomodulators: A role for adrenoceptor-mediated mechanisms in fine tuning of T-cell development. Auton. Neurosci. 2008, 15, 1–12. [Google Scholar] [CrossRef]
- Vrontou, S.; Wong, A.; Rau, K.; Koerber, R.; Anderson, D. Genetic identification of C fibres that detect massage-like stroking of hairy skin in vivo. Nature 2013, 493, 669–673. [Google Scholar] [CrossRef]
- Bangasser, D.A.; Wicks, B. Sex-specific mechanisms for responding to stress. J. Neurosci. Res. 2017, 95, 75–82. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Purves-Tyson, T.D.; Handelsman, D.J.; Double, K.L.; Owens, S.J.; Bustamante, S.; Weickert, C.S. Testosterone regulation of sex steroid-related mRNAs and dopamine-related mRNAs in adolescent male rat substantia nigra. BMC Neurosci. 2012, 13, 95. [Google Scholar] [CrossRef]
- Tan, Ü. Paw preferences in dogs. Int. J. Neurosci. 1987, 32, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Siniscalchi, M.; D’Ingeo, S.; Quaranta, A. Lateralized functions in the dog brain. Symmetry 2017, 9, 71. [Google Scholar] [CrossRef]
- Siniscalchi, M.; Sasso, R.; Pepe, A.M.; Vallortigara, G.; Quaranta, A. Dogs turn left to emotional stimuli. Behav. Brain Res. 2010, 208, 516–521. [Google Scholar] [CrossRef]
- Siniscalchi, M.; d’Ingeo, S.; Fornelli, S.; Quaranta, A. Relationship between visuospatial attention and paw preference in dogs. Sci. Rep. 2016, 6, 31682. [Google Scholar] [CrossRef]
- Branson, N.J.; Rogers, L.J. Relationship between paw prefer- ence strength and noise phobia in Canis familiaris. J. Comp. Psychol. 2006, 120, 176–183. [Google Scholar] [CrossRef]
- Schneider, L.A.; Delfabbro, P.H.; Burns, N.R. Temperament and lateralisation in the domestic dog (Canis familiaris). J. Vet. Behav. 2013, 8, 124–134. [Google Scholar] [CrossRef]
- Aydinlioglu, A.; Arslan, K.; Cengiz, N.; Ragbetli, M.; Erdogan, E. The relationships of dog hippocampus to sex and paw preference. Int. J. Neurosci. 2006, 116, 77–88. [Google Scholar] [CrossRef]


{kind=link}
| Item | z | HI (Absolute Value) | ||
|---|---|---|---|---|
| Mean | Range | Mean | Range | |
| Left paw n = 16 | −5.4 | −7.8–3.2 | 0.54 | 0.3–0.8 |
| Right paw n = 24 | 5.1 | 3.8–7.6 | 0.51 | 0.4–0.8 |
| Items | Noradrenaline | Serotonin | Cortisol | |
|---|---|---|---|---|
| pg/mL | ng/mL | ng/mL | ||
| F + NS | 238.48 | 457.73 | 51.83 | |
| M + NS | 278.52 | 602.51 | 59.16 | |
| F + SS | 344.18 | 507.08 | 46.16 | |
| M + SS | 451.33 | 578.41 | 48.97 | |
| SEM | 0.039 | 0.073 | 0.012 | |
| Effect of sex (EP) | F | 291.33 b | 482.41 b | 48.99 b |
| M | 364.33 a | 590.46 a | 54.07 a | |
| Effect of stroking (EB) | NS | 258.50 y | 530.12 y | 55.49 x |
| SS | 397.76 x | 542.75 x | 47.57 y | |
| p-value | ||||
| EP effect | 0.042 | 0.032 | 0.042 | |
| EB effect | 0.011 | 0.023 | 0.031 | |
| EP x EB interaction | 0.235 | 0.218 | 0.089 | |
| Item | Noradrenaline | Serotonin | Cortisol | |
|---|---|---|---|---|
| pg/mL | ng/mL | ng/mL | ||
| L + NS | 147.38 | 501.95 | 59.65 | |
| R + NS | 369.61 | 568.28 | 51.34 | |
| L + SS | 367.25 | 527.65 | 43.5 | |
| R + SS | 428.25 | 607.85 | 51.63 | |
| SEM | 0.029 | 0.037 | 0.009 | |
| Effect of laterality (EL) | L | 257.32 b | 514.80 b | 51.58 |
| R | 398.93 a | 573.07 a | 51.49 | |
| Effect of stroking (EB) | NS | 258.50 y | 541.75 y | 55.50 x |
| SS | 397.75 x | 575.77 x | 47.57 y | |
| p-value | ||||
| EL effect | 0.042 | 0.033 | 0.533 | |
| EB effect | 0.049 | 0.042 | 0.026 | |
| EL x EB interaction | 0.232 | 0.156 | 0.838 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpiński, M.; Ognik, K.; Garbiec, A.; Czyżowski, P.; Krauze, M. Effect of Stroking on Serotonin, Noradrenaline, and Cortisol Levels in the Blood of Right- and Left-Pawed Dogs. Animals 2021, 11, 331. https://doi.org/10.3390/ani11020331
Karpiński M, Ognik K, Garbiec A, Czyżowski P, Krauze M. Effect of Stroking on Serotonin, Noradrenaline, and Cortisol Levels in the Blood of Right- and Left-Pawed Dogs. Animals. 2021; 11(2):331. https://doi.org/10.3390/ani11020331
Chicago/Turabian StyleKarpiński, Mirosław, Katarzyna Ognik, Aleksandra Garbiec, Piotr Czyżowski, and Magdalena Krauze. 2021. "Effect of Stroking on Serotonin, Noradrenaline, and Cortisol Levels in the Blood of Right- and Left-Pawed Dogs" Animals 11, no. 2: 331. https://doi.org/10.3390/ani11020331
APA StyleKarpiński, M., Ognik, K., Garbiec, A., Czyżowski, P., & Krauze, M. (2021). Effect of Stroking on Serotonin, Noradrenaline, and Cortisol Levels in the Blood of Right- and Left-Pawed Dogs. Animals, 11(2), 331. https://doi.org/10.3390/ani11020331