Biofilm Formation Reducing Properties of Manuka Honey and Propolis in Proteus mirabilis Rods Isolated from Chronic Wounds

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Prevalence of P. mirabilis in Wound Infections of Chronic Wounds at Dr. Antoni Jurasz University Hospital No. 1 in Bydgoszcz from 2016 to 2018
2.2. Proteus Mirabilis Examined Clinical Strains
2.3. Reference Strains
2.4. Manuka Honey
2.4.1. Polyphenols Content in Manuka Honey
2.4.2. Antioxidant Activity of Manuka Honey
2.5. New Zealand Propolis Solution Preparation Method
2.6. Minimal Bactericidal Concentration and Biofilm Bactericidal Concentration of Manuka Honey and New Zealand Propolis in Reference Strains
2.7. Minimal Biofilm Inhibitory Concentration, Biofilm Prevention Concentration, and Minimal Biofilm Eradication Concentration of Manuka Honey and New Zealand Propolis in Clinical Strains
2.8. Impact of Manuka Honey and Ethanol Extract of Propolis (EEP) on the Metabolic Activity of P. mirabilis Mature Biofilm
2.8.1. Biofilm Cultivation
2.8.2. Manuka Honey and EEP Impact on Biofilm
2.8.3. Assessment of Mature Biofilm Metabolic Activity after 24 h of Manuka Honey or EEP Treatment
2.9. Statistical Analysis
- A (K+) means positive control absorbance;
- A (x) means absorbance after treatment with honey or EEP in defined concentration.
3. Results
3.1. Prevalence of P. mirabilis in Wound Infections of Chronic Wounds at Dr. Antoni Jurasz University Hospital No. 1 in Bydgoszcz from 2016 to 2018
3.2. Comparison of Minimal Bactericidal Concentration (MBC) and Biofilm Bactericidal Concentration (BBC) Values of Manuka Honey and EEP in Reference Strains
3.3. Comparison of MBIC, BPC, and MBEC Values of Manuka Honey and EEP in Proteus Mirabilis Clinical Strains
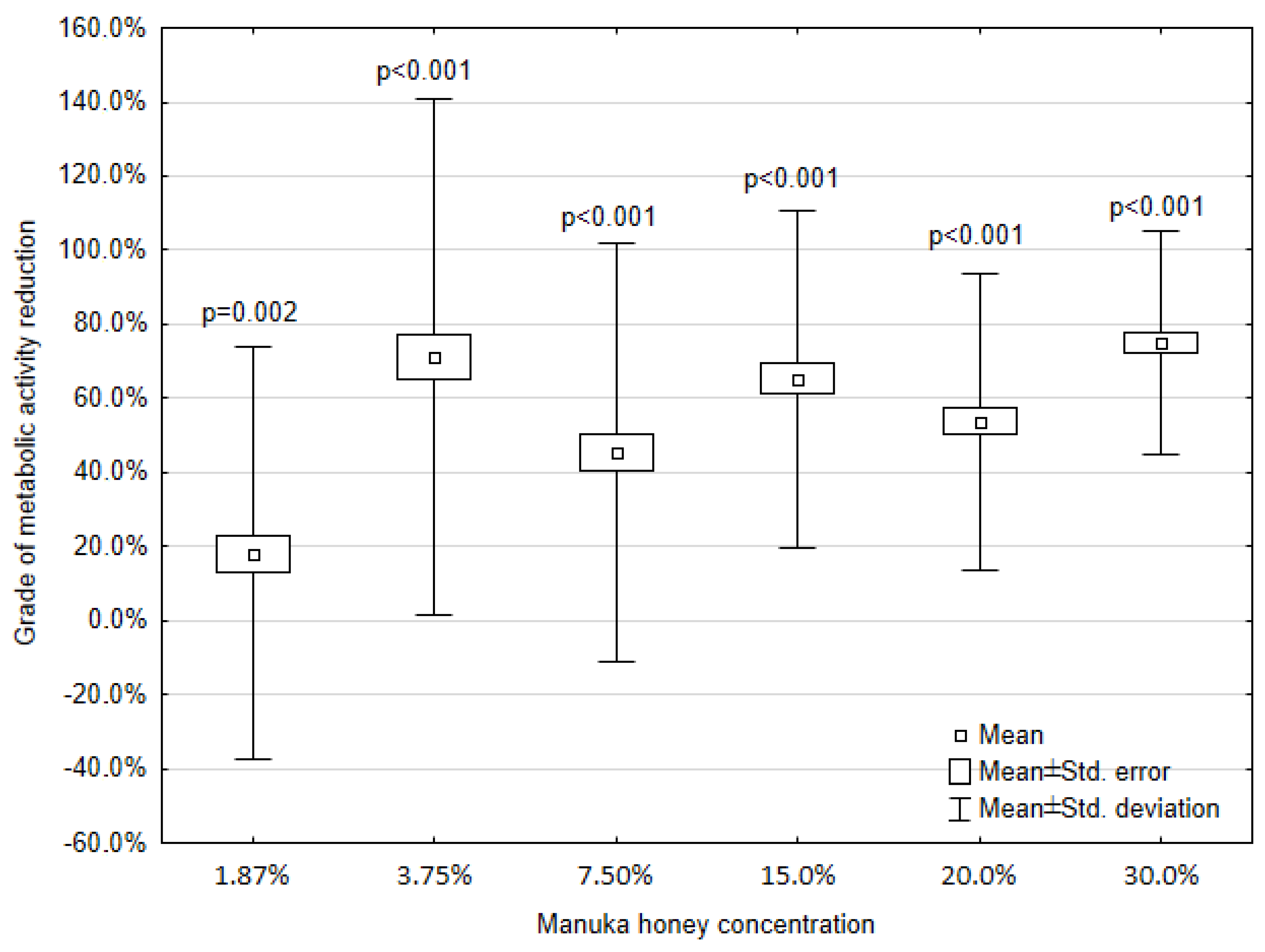
3.4. Presentation of Causality between Manuka Honey or EEP Impact and Metabolic Activity of the Mature Biofilm Derived from Clinical Strains
3.5. Comparison of the EEP and EtOH Impact
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MBIC | Minimal biofilm inhibition concentration, defined as the lowest concentration of a substance that inhibits visible growth of 24-h mature biofilm culture. |
| BPC | Biofilm prevention concentration, defined as the lowest concentration of a substance that inhibits visible growth in the culture where bacterial inoculation and substance exposure occur at the same time. |
| MBEC | Minimal biofilm-eradication concentration, defined according to [29]. |
| BBC | Biofilm bactericidal concentration, defined according to [29]. |
Appendix A

{kind=link}
{kind=link}
| Reference Strain | CFU Per mL | Standard Deviation |
|---|---|---|
| SAU 25923™ | 1.57 × 1010 | ±1.26 × 109 |
| SAU 29213™ | 1.28 × 109 | ±1.16 × 108 |
| EFA 29212™ | 5.83 × 107 | ±5.44 × 106 |
| ECO 25922™ | 3.33 × 109 | ±5.18 × 108 |
| ECO 35218™ | 1.45 × 108 | ±2.73 × 107 |
| PAE 27853™ | 1.60 × 1010 | ±1.51 × 109 |
References
- Ples, M.; Glik, J.; Misiuga, M.; Skotnicka, J.; Kawecki, M.; Nowak, M. Rany przewlekłe i ich leczenie. Substytuty skóry i przeszczepy allogeniczne. JOTSRR 2016, 1, 48–56. [Google Scholar]
- Kouros, I.; Parham, G. Chronic wounds. Clin. Plast. Surg. 2005, 32, 209–222. [Google Scholar] [CrossRef]
- Potempa, M.; Jonczyk, P.; Janerka, M.; Kucharzewski, M.; Kawczyk-Krupka, A. Rany przewlekłe—Epidemiologia i czynniki wpływające na proces gojenia. Leczenie Ran. 2014, 11, 43–50. [Google Scholar] [CrossRef]
- Dzikowska, M.; Gajda, K.; Merklinger-Soma, M. Analiza występowania odleżyn u pacjentów przebywających w szpitalu specjalistycznym o profilu zachowawczym. Probl. Pielęg. 2011, 19, 162–170. [Google Scholar]
- Hryniewicz, W.; Kulig, J.; Ozorowski, T.; Mól, A.; Kulig, P.; Wąchol, D. Stosowanie Antybiotyków W Wybranych Zakażeniach Skóry i Tkanek Miękkich; NPOA Narodowy Instytut Leków: Warsaw, Poland, 2012. [Google Scholar]
- Cutting, K.F.; White, R.J.; Mahoney, P.; Harding, K.G. Clinical identification of wound infection: A Delphi approach. In European Wound Management Association (EWMA) Position Document: Identifying Criteria for Wound Infections; EWMA, MEP Ltd.: London, UK, 2005. [Google Scholar]
- Mospan, B. Zastosowanie preparatów antyseptycznych zawierających dichlorowodorek oktenidyny w pielęgnacji i leczeniu zakażeń stopy cukrzycowej zgodnie z koncepcją TIME. Leczenie Ran. 2010, 7, 63–69. [Google Scholar]
- Graczyk, M.; Bronszkiewicz, E.; Zgorzelak, B. Profilaktyka i leczenie odleżyn—Ocena pacjentów przebywających na oddziale stacjonarnym Hospicjum im. bł. ks. J. Popiełuszki w Bydgoszczy w latach 2012–2013. Palliat. Med. Pract. 2014, 8, 91–102. [Google Scholar]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the chronic wound microbiota of 2963 patients by 16S rDNA pyrosequencing. Wound Repair Regen. 2016, 163–174. [Google Scholar] [CrossRef]
- Różalski, A.; Kwil, I.; Torzewska, A.; Baranowska, M.; Stączek, P. Proteus bacilli: Features and virulence factors. Post Hig. 2007, 61, 204–219. [Google Scholar]
- Hurlow, J.; Blanz, E.; Gaddy, J.A. Clinical investigation of biofilm in non-healing wounds by high resolution microscopy techniques. J. Wound Care 2016, 25, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Cooper, R.A.; Molan, P.C.; Harding, K.G. Antibacterial activity of honey against strains of Staphylococcus aureus from infected wounds. J. R. Soc. Med. 1999, 92, 283–285. [Google Scholar] [CrossRef] [Green Version]
- Mavric, E.; Wittmann, S.; Barth, G.; Henle, T. Identification and quantification of methylglyoxal as the dominant antibacterial constituent of Manuka (Leptospermum scoparium) honeys from New Zealand. Mol. Nutr. Food Res. 2008, 52, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Oelschlaegel, S.; Gruner, M.; Wang, P.N.; Boettcher, A.; Koelling-Speer, I.; Speer, K. Classification and characterization of manuka honeys based on phenolic compounds and methylglyoxal. J. Agric. Food Chem. 2012, 60, 7229–7237. [Google Scholar] [CrossRef] [PubMed]
- Kwakman, P.H.; Zaat, S.A.J. Antibacterial components of honey. IUBMB Life 2012, 64, 48–55. [Google Scholar] [CrossRef]
- Schneider, M.; Coyle, S.; Warnock, M.; Gow, I.; Fyfe, L. Anti-microbial acitivity and composition of Manuka and Portobello honey. Phytother. Res. 2013, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J.; Bohova, J.; Prochazka, E.; Klaudiny, J. Methylglyoxal may affect peroxide accumulation in Manuka honey through the inhibition of glucose oxidase. J. Med. Food 2014, 17, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.I.; Alam, N.; Moniruzzaman, M.; Sulaiman, S.A.; Gan, S.H. Phenolic acid composition and antioxidant properties of Malaysian honeys. J. Food Sci. 2011, 76, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Kwakman, P.H.; Te Velde, A.A.; de Boer, L.; Vandenbroucke-Grauls, C.M.J.E.; Zaat, S.A.J. Two major medicinal honeys have different mechanisms of bactericidal acitvity. PLoS ONE 2011, 6, e17709. [Google Scholar] [CrossRef] [Green Version]
- Moniruzzaman, M.; Sulaiman, S.A.; Khalil, M.I.; Gan, S.H. Evaluation of physicochemical and antioxidant properties of sourwood and other Malaysian honeys: A comparison with manuka honey. Chem. Cent. J. 2013, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Pattamayutanon, P.; Angeli, S.; Thakeow, P.; Abraham, J.; Disayathanoowat, T.; Chantawannakul, P. Biomedical activity and related volatile compounds of Thai honeys from 3 different honeybee spieces. J. Food Sci. 2015, 80, M2228–M2240. [Google Scholar] [CrossRef]
- Roshan, N.; Rippers, T.; Locher, C.; Hammer, K.A. Antibacterial activity and chemical characteristics of several Western Australian honeys compared to manuka honey and pasture honey. Arch. Microbiol. 2017, 347–355. [Google Scholar] [CrossRef]
- Stephens, J.M.C. The Factors Responsible for the Varying Levels of UMF(R) in Manuka (Leptospermum Scopiarum) Honey. Ph.D. Thesis, The Univeristy of Waikato, Waikato, New Zealand, 2006. Available online: https://www.umf.org.nz/wp-content/myimages/2017/02/Dr-Stephens-The-factors-responsible-for-UMF-in-Manuka-honey.pdf (accessed on 1 August 2020).
- Atrott, J.; Henle, T. Methylglyoxal in Manuka honey—Correlation with antibacterial properties. Czech. J. Food Sci. 2009, 27, S163–S165. [Google Scholar] [CrossRef] [Green Version]
- Zabaiou, N.; Fouache, A.; Trousson, A.; Baron, S.; Zellagui, A.; Lahouel, M.; Lobacarro, J.M.A. Biological properties of propolis extracts: Something new from an ancient product. Chem. Phys. Lipids 2017, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic properties of bioactive compounds from different honey products. Front. Pharmacol. 2017, 8, 412. [Google Scholar] [CrossRef] [PubMed]
- Markham, K.; Mitchell, K.; Wilkins, A.; Daldy, J.; Lu, Y. HPLC and GC-MS indetification of the major organic constituents in New Zealand propolis. Phytochemistry 1996, 42, 205–211. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Quality and nutritional value of strawberry fruit under long term salt stress. Food Chem. 2008, 107, 1413–1420. [Google Scholar] [CrossRef]
- Macià, M.D.; Rojo-Molinero, E.; Oliver, A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin. Microbiol. Infect. 2014, 20, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.N.; Wysocki, A.; Specht-Glick, D.D.; Rooney, A.; Feldman, R.A.; St Amand, A.L.; Pace, N.R.; Trent, J.D. Microbial diversity in chronic open wounds. Wound Repair Regen. 2009, 17, 163–172. [Google Scholar] [CrossRef]
- Abad, C.; Safdar, N. From ulcer to infection: An update on clinical practice and adjunctive treatments of diabetic foot ulcers. Curr. Infect. Dis. Rep. 2012, 14, 540–550. [Google Scholar] [CrossRef]
- Szewczyk, M.T.; Gospodarek, E.; Mościcka, P.; Jawień, A.; Cwajda-Białasik, J.; Cierzniakowska, K.; Hancke, E. Infections of chronic wounds—A serious medical problem. Pielęg. Chir. Angiol. 2015, 2015, 1–6. [Google Scholar]
- Okhiria, O.A.; Henriques, A.F.M.; Burton, N.F.; Peters, A.; Cooper, R.A. Honey modulates biofilm of Pseudomonas aeruginosa in a time and dose dependent manner. JAAS 2009, 1, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Jervis-Brady, J.; Foreman, A.; Bray, S.; Tan, L.; Wormald, P.J. Methylglyoxal-infused honey mimics the anti-Staphylococcus aureus biofilm acitivty of Manuka honey: Potential implication in chronic rhinositusitis. Laryngoscope 2011, 121, 1104–1107. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Turnbull, L.; Burke, C.M.; Liu, M.; Carter, D.A.; Schlothauer, R.C.; Whitchurch, C.B.; Harry, E.J. Manuka-type honeys can eradicate biofilms produced by Staphylococcus aureus with different biofilm-forming abilities. PeerJ 2014, 2, e326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddocks, S.E.; Jenkins, R.E.; Rowlands, R.S.; Purdy, K.J.; Cooper, R.A. Manuka honey inhibits adhesion and invasion of medically important wound bacteria in vitro. Future Microbiol. 2013, 8, 1523–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emineke, S.; Cooper, A.J.; Fouch, S.; Birch, B.R.; Lwaleed, B.A. Diluted honey inhibits biofilm formation: Potential application in urinacry catheter management? Clin. Pathol. 2017, 70, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Tze Tan, H.; Rahman, R.A.; Hua Gan, S.; Halim, A.S.; Asma’Hassan, S.; Sulaiman, S.A.; Kurnpla-Kaur, B.S. The antimicrobial properties of Malaysian tualang honey against wound and enteric microorganisms in comparison to manuka honey. BMC Complement. Altern. Med. 2009, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Majtan, J.; Bohova, J.; Horniackova, M.; Klaudiny, J.; Majtan, V. Anti-biofilm effects of honey against wound pathogens Proteus mirabilis and Enterobacter cloacae. Phytother. Res. 2013, 28, 69–75. [Google Scholar] [CrossRef]
- Ambi, A.; Bryan, J.; Borbon, K.; Centeno, D.; Liu, T.; Chen, T.P.; Cattabiani, T.; Traba, C. Are Russian propolis ethanol extracts the future for the prevention of medical and biomedical implant contaminations? Phytomedicine 2017, 30, 50–58. [Google Scholar] [CrossRef]
- Sherlock, O.; Dolan, A.; Athman, R.; Power, A.; Gethin, G.; Cowman, S.; Humpreys, H. Comparison of the antimicrobial activity of Ulmo honey from Chile and Manuka honey against methicillin-resistant Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2010, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kwakman, P.H.S.; te Velde, A.A.; de Boer, L.; Speijer, D.; Vandenbroucke-Grauls, C.M.; Zaat, S.A. How honey kills bacteria. FASEB J. 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [Green Version]
- Hixon, K.R.; Lu, T.; Carletta, M.N.; McBridge-Gagyi, S.H.; Janowiak, B.E.; Sell, S.A. A preliminary in vitro evaluation of the bioactive potential of cryogel scaffolds incorporated with Manuka honey for the treatment of chronic bone infections. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 106, 1918–1933. [Google Scholar] [CrossRef]
- Schuhladen, K.; Swathi, N.V.R.; Liverani, L.; Neščákowá, Z.; Boccaccini, A.R. Production of a novel poly(ε-caprolactone)-methylcellulose electrospun wound dressing by incorporating bioactive glass and Manuka honey. J. Biomed. Mater. Res. B Appl. Biomater. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Ward, J.; Choy, K. L Nature-inspired bacterial cellulose/methylglyoxal (BC/MGO) nanocomposite for broad-spectrum antimicrobial wound dressing. Macromol. Biosci. 2020, 20, 200070. [Google Scholar] [CrossRef] [PubMed]
- Vas, P.; Rayman, G.; Dhatariya, K.; Driver, V.; Hartemann, A.; Londahl, M.; Piagessi, A.; Apelqvist, J.; Attinger, C.; Game, F. Effectiveness of interventions to enhance healing of chronic foot ulcers in diabetes: A systematic review. Diabetes Metab. Res. Rev. 2020, 36, e3284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albietz, J.M.; Schmid, K.L. Randomised cotrolled trial of topical antibacterial Manuka (Leptospermum species) honey for evaporative dry eye due to meibomian gland dysfunction. Clin. Exp. Optom. 2017, 100, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, V.S.; Humpreys, I.M.; Purcell, P.L.; Greg, E.D. Manuka honey sinus irrigation for the treatment of chronic rhinosinusitis: A randomized controlled trial. Int. Forum Allergy Rhinol. 2017, 7, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Alencar, S.M.; Oldoni, T.L.; Castro, M.L.; Cabral, I.S.; Costa-Neto, C.M.; Cury, J.A.; Rosalen, P.L.; Ikegaki, M. Chemical composition and biological activity of a new type of Brazilian propolis: Red propolis. J. Ethnopharmacol. 2007, 113, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Wojtyczka, R.; Dziedzic, A.; Idzik, D.; Kępa, M.; Kubina, R.; Kabała-Dzik, A.; Smoleń-Dzirba, J.; Stojko, J.; Sajewicz, M.; Wąsik, T. Susceptibility of Staphylococcus aureus clinical isolates to propolis extract alone or in combination with antimicrobial drugs. Molecules 2013, 18, 9623–9640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouidhi, B.; Zmantar, T.; Bakhrouf, A. Anti-cariogenic and anti-biofilms activity of Tunisian propolis extract and its potential protective effect against cancer cells proliferation. Anaerobe 2010, 16, 566–571. [Google Scholar] [CrossRef]
- De Oliveira Dembogurski, D.S.; Silva Trentin, D.; Boaretto, A.G.; Rigo, G.V.; da Silva, R.C.; Tasca, T.; Macedo, A.J.; Carollo, C.A.; Silva, D.B. Brown propolis-metabolomic innovative approach to determine compounds capable of killing Staphylococcus aureus biofilm and Trichomonas vaginalis. Food Res. Int. 2018, 111, 661–673. [Google Scholar] [CrossRef]
- Krasnodebski, J.; Scheller, S.; Suchy, H. Use of an ethanol extract of propolis in the treatment of vaginitis and conditions after electrooagulation of cervical erosion. Ginekol. Pol. 1986, 57, 471–478. [Google Scholar]
- Rojczyk, E.; Klama-Baryła, A.; Łabuś, W.; Wilemska-Kucharzewska, K.; Kucharzewski, M. Historical and modern research on propolis and its application in wound healing and other fields of medicine and contribution. J. Ethnopharmacol. 2020, 262, 113159. [Google Scholar] [CrossRef]
- Kucharzewski, M.; Kubacka, S.; Urbanek, T.; Wilemska-Kucharzewska, K.; Morawiec, T. Stan Scheller: The forerunner of clinical studies on using propolis for poor and chronic nonhealig wounds. Evid. Based Complement. Altern. Med. 2013, 2013, 456859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magro Filho, O.; de Carvalho, A.C. Application of propolis to dental sockets and skin wounds. J. Nihon Univ. Sch. Dent. 1990, 32, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khachaturov, A.A.; Gudkov, A.I. Propolis therapy of certain dermatoses and burns in the far north. Vestn. Dermatol. Venerol. 1969, 43, 63–65. [Google Scholar] [PubMed]
- Catchpole, O.; Mitchell, K.; Bloor, S.; Davis, P.; Suddes, A. Antiproliferative activity of New Zealand propolis and phenolic compounds vs human colorectal adenocarcinoma cells. Fitoterapia 2015, 106, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, K.; Singh, S.; Burke, T.R., Jr.; Grunberger, D.; Aggarwal, B.B. Caffeic acid phenethyl ester is a potent and specific inhibitor of activation of nuclear transcription factor NF-kappa B. Proc. Natl. Acad. Sci. USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romana-Souza, B.; Dos Santos, J.S.; Monte-Alto-Costa, A. Caffeic acid phenethyl ester promotes wound healing of mice pressure ulcers affecting NF-κB, NOS2 and NRF2 expression. Life Sci. 2018, 207, 158–165. [Google Scholar] [CrossRef]
- Gheib, N.; Farzam, A.; Habibian, Z.; Samiee-Rad, F. The effect of oral consumption of propolis alone and in combination with silver nanoparticles on wound healing in male Wistar rats. Wound Manag. Prev. 2020, 66, 38–46. [Google Scholar] [CrossRef]
- Ong, T.H.; Chitra, E.; Ramamurthy, S.; Siddalingam, R.P.; Yuen, K.H.; Ambu, S.P.; Davamani, F. Chitosan-propolis nanoparticle formulation demonstrates anti-bacterial activity against Enterococcus faecalis biofilms. PLoS ONE 2017, 12, e0174888. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Mello, D.; Vasconcellos, L.; Valente, A.J.M.; Borges, A. Chitosan-based coacervate polymers for propolis encapsulation: Release and cutotoxicity studies. Int. J. Mol. Sci. 2020, 21, 4561. [Google Scholar] [CrossRef]
- Marquele-Oliveira, F.; da Silva Barud, H.; Torres, E.C.; Machado, R.T.A.; Caetano, G.F.; Leite, M.N.; Frade, M.A.C.; Ribeiro, S.J.L.; Berretta, A.A. Development, characterization and pre-clinical trials of an innovative wound healing dressing based on propolis (EPP-AF®)-containing self-microemulsifying formulation incorporated in biocellulose membranes. Int. J. Biol. Macromol. 2019, 136, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Eskandarinia, A.; Kefayat, A.; Rafienia, M.; Agheb, M.; Navid, S.; Ebrahimpour, K. Cornstarch-based wound dressing incorporated with hyaluronic acid and propolis: In vitro and in vivo studies. Carbohydr. Polym. 2019, 216, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Eskandarinia, A.; Kefayat, A.; Gharakhloo, M.; Agheb, M.; Khodabakhshi, D.; Khorshidi, M.; Sheikhmoradi, V.; Rafienia, M.; Salehi, H. A propolis enriched polyurethane-hyaluronic acid nanofibrous wound dressing with remarkable antibacterial and wound healing activities. Int. J. Biol. Macromol. 2020, 149, 467–476. [Google Scholar] [CrossRef] [PubMed]


| Reference Strains | MH (v/v%) | EEP (v/v%) | ||||
|---|---|---|---|---|---|---|
| MBC | >BBC | MBC | >BBC | |||
| 0.5 MF | 1:100 Dilution a | 0.5 MF | 1:100 Dilution a | |||
| SAU 25923™ | 20.0 | 20.0 | 15.0 | 5.0 | 20.0 | 40.0 |
| SAU 29213™ | 20.0 | 25.0 | 25.0 | 5.0 | 40.0 | 40.0 |
| EFA 29212™ | 20.0 | 25.0 | 25.0 | 5.0 | 20.0 | 20.0 |
| ECO 25922™ | 15.0 | 10.0 | 10.0 | 5.0 | 40.0 | 40.0 |
| ECO 35218™ | 15.0 | 10.0 | 10.0 | 5.0 | 20.0 | 40.0 |
| PAE 27853™ | 25.0 | >25.0 | 15.0 | 10.0 | 40.0 | 40.0 |
| MH Concentration (v/v%) | Number of P. mirabilis Strains (%) | ||
|---|---|---|---|
| MBIC b 0 h | BPC c 0 h | MBEC c 24 h | |
| >30.0 | 0 (0.0) | 11 (35.5) | 31 (100.0) |
| 30.0 | 28 (90.3) | 20 (64.5) | 0 (0.0) |
| 20.0 | 3 (9.7) | 0 (0.0) | 0 (0.0) |
| 15.0 | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Total number of strains | 31 (100.0) | 31 (100.0) | 31 (100.0) |
| C [v/v%] | Number of P. mirabilis Strains | |||||
|---|---|---|---|---|---|---|
| MBIC b 0 h (%) | BPC c 0 h (%) | MBEC c 24 h (%) | ||||
| EEP | EtOH | EEP | EtOH | EEP | EtOH | |
| >40.0% | - | - | - | - | - | 3 (9.67) |
| 40.0 | - | - | - | - | - | 6 (19.3) |
| 20.0 | - | - | 5 (16.1) | 3 (9.7) | 3 (9.7) | 2 (6.45) |
| 10.0 | - | 1 (3.2) | 12 (38.7) | 14 (45.2) | 21 (67.7) | 6 (19.3) |
| 5.0 | 4 (12.9) | 22 (71.0) | 12 (38.7) | 13 (41.9) | 7 (22.6) | 4 (12.9) |
| 2.5 | 17 (54.8) | 8 (25.8) | 2 (6.5) | - | - | 7 (22.6) |
| 1.0 | 4 (12.9) | - | - | 1 (3.2) | - | 3 (9.7) |
| <1.0 | 6 (19.4) | - | - | - | - | - |
| Total number of strains | 31 (100.0) | |||||
| Manuka Honey | |||
|---|---|---|---|
| Strain | UMF 10+ a | UMF 25+ b | Values Maintained in This Study (UMF 13+) |
| S. aureus ATCC® 25923™ | MBC 25.0% | - | MBC 20.0% |
| P. aeruginosa ATCC® 27853™ | MBC 22.5% | MBC 12.5% | MBC 25.0% |
| E. coli ATCC® 25922™ | MBC 15.0% | - | MBC 15.0% |
| E. coli ATCC® 35218™ | - | MBC 12.5% | MBC 15.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiecińska-Piróg, J.; Przekwas, J.; Majkut, M.; Skowron, K.; Gospodarek-Komkowska, E. Biofilm Formation Reducing Properties of Manuka Honey and Propolis in Proteus mirabilis Rods Isolated from Chronic Wounds. Microorganisms 2020, 8, 1823. https://doi.org/10.3390/microorganisms8111823
Kwiecińska-Piróg J, Przekwas J, Majkut M, Skowron K, Gospodarek-Komkowska E. Biofilm Formation Reducing Properties of Manuka Honey and Propolis in Proteus mirabilis Rods Isolated from Chronic Wounds. Microorganisms. 2020; 8(11):1823. https://doi.org/10.3390/microorganisms8111823
Chicago/Turabian StyleKwiecińska-Piróg, Joanna, Jana Przekwas, Michał Majkut, Krzysztof Skowron, and Eugenia Gospodarek-Komkowska. 2020. "Biofilm Formation Reducing Properties of Manuka Honey and Propolis in Proteus mirabilis Rods Isolated from Chronic Wounds" Microorganisms 8, no. 11: 1823. https://doi.org/10.3390/microorganisms8111823
APA StyleKwiecińska-Piróg, J., Przekwas, J., Majkut, M., Skowron, K., & Gospodarek-Komkowska, E. (2020). Biofilm Formation Reducing Properties of Manuka Honey and Propolis in Proteus mirabilis Rods Isolated from Chronic Wounds. Microorganisms, 8(11), 1823. https://doi.org/10.3390/microorganisms8111823