Nutrient Accumulation and Distribution Assessment in Response to Potassium Application under Maize–Soybean Intercropping System


Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials and Site Description
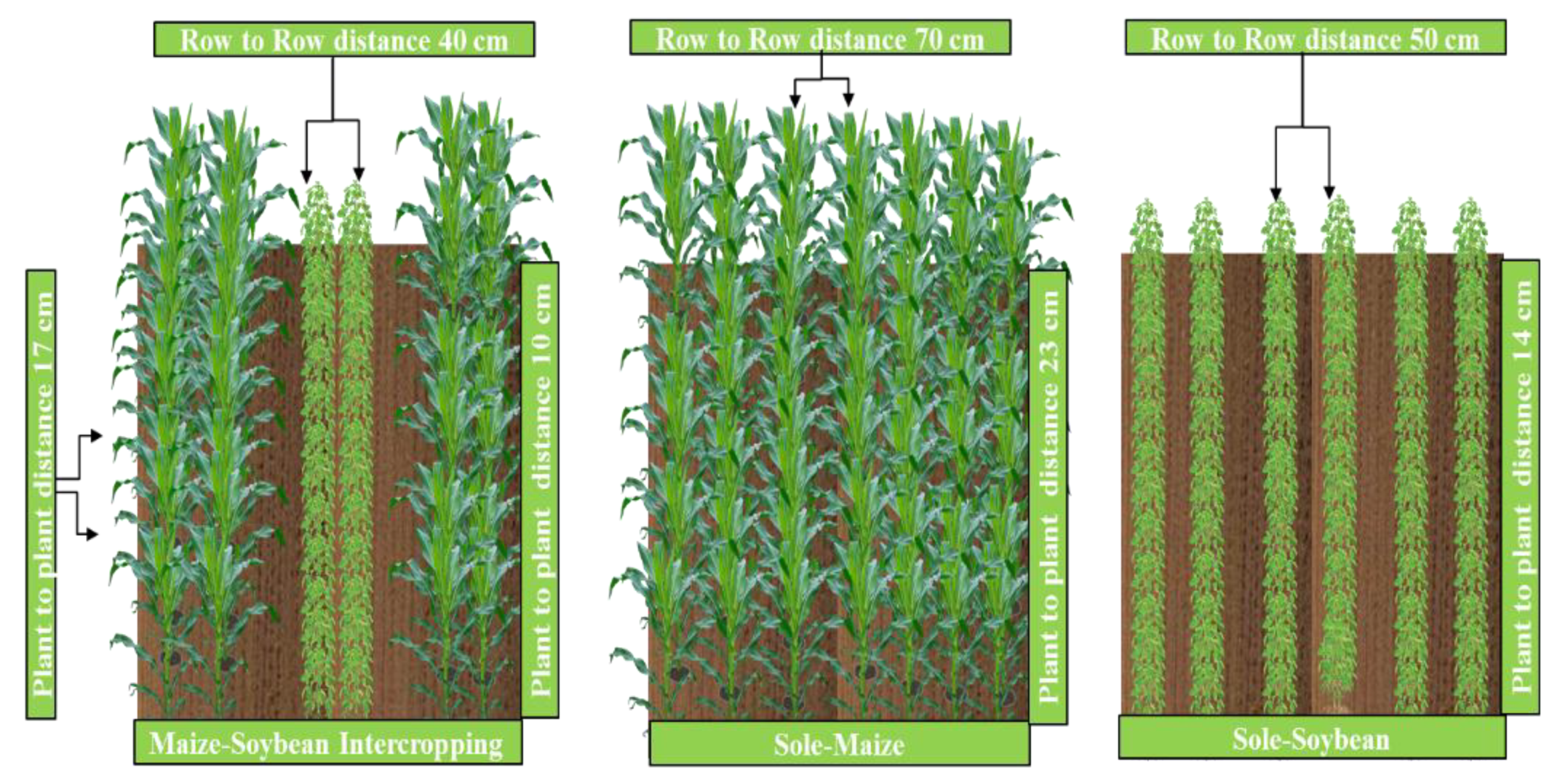
2.2. Experiment Design
2.3. Nutrient Accumulation and Distribution
3. Statistical Analyses
4. Results
4.1. K Effect on Plant Biomass Accumulation
4.2. K Effect on Plant Biomass Distribution
4.3. K Effects on Nitrogen Accumulation
4.4. K Effects on Nitrogen Distribution
4.5. K Effects on Phosphorus (P) Accumulation
4.6. K Effects on Phosphorus (P) Distribution
4.7. K Effects on Potassium (K) Accumulation
4.8. K Effects on Potassium (K) Distribution
4.9. K Effect on Yield and Land Equivalent Ratio
5. Correlation Analysis
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouma, G. Intercropping and its application to banana production in East Africa: A review. J. Plant Breed. Crop Sci. 2009, 1, 13–15. [Google Scholar]
- Zomer, R.J.; Trabucco, A.; Coe, R.; Place, F. Trees on farm: Analysis of global extent and geographical patterns of agroforestry. ICRAF Work. Pap. World Agrofor. Cent. 2009, 89, 63. [Google Scholar]
- Raza, M.A.; Feng, L.Y.; van der Werf, W.; Cai, G.R.; Khalid, M.H.B.; Iqbal, N.; Hassan, M.J.; Meraj, T.A.; Naeem, M.; Khan, I. Narrow-wide-row planting pattern increases the radiation use efficiency and seed yield of intercrop species in relay-intercropping system. Food Energy Secur. 2019, 8, e170. [Google Scholar] [CrossRef]
- Feng, L.; Raza, M.A.; Chen, Y.; Khalid, M.H.B.; Meraj, T.A.; Ahsan, F.; Fan, Y.; Du, J.; Wu, X.; Song, C. Narrow-wide row planting pattern improves the light environment and seed yields of intercrop species in relay intercropping system. PLoS ONE 2019, 14, e0212885. [Google Scholar] [CrossRef]
- Ahmed, S.; Raza, M.A.; Zhou, T.; Hussain, S.; Khalid, M.H.B.; Feng, L.; Wasaya, A.; Iqbal, N.; Ahmed, A.; Liu, W. Responses of soybean dry matter production, phosphorus accumulation, and seed yield to sowing time under relay intercropping with maize. Agronomy 2018, 8, 282. [Google Scholar] [CrossRef] [Green Version]
- Du, J.-B.; Han, T.-F.; Gai, J.-Y.; Yong, T.-W.; Sun, X.; Wang, X.-C.; Yang, F.; Liu, J.; Shu, K.; Liu, W.-G. Maize-soybean strip intercropping: Achieved a balance between high productivity and sustainability. J. Integr. Agric. 2018, 17, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Hussain, S.; Ahmed, Z.; Yang, F.; Wang, X.; Liu, W.; Yong, T.; Du, J.; Shu, K.; Yang, W. Comparative analysis of maize–soybean strip intercropping systems: A review. Plant Prod. Sci. 2019, 22, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Sui, P.; Huang, J.; Wang, D.; Whalen, J.K.; Chen, Y. Global warming potential from maize and maize-soybean as affected by nitrogen fertilizer and cropping practices in the North China Plain. Field Crop. Res. 2018, 225, 117–127. [Google Scholar] [CrossRef]
- Yan, Y.; Gong, W.; Yang, W.; Wan, Y.; Chen, X.; Chen, Z.; Wang, L. Seed treatment with uniconazole powder improves soybean seedling growth under shading by corn in relay strip intercropping system. Plant Prod. Sci. 2010, 13, 367–374. [Google Scholar] [CrossRef]
- Yang, F.; Huang, S.; Gao, R.; Liu, W.; Yong, T.; Wang, X.; Wu, X.; Yang, W. Growth of soybean seedlings in relay strip intercropping systems in relation to light quantity and red: Far-red ratio. Field Crop. Res. 2014, 155, 245–253. [Google Scholar] [CrossRef]
- Wu, Y.; Gong, W.; Yang, W. Shade inhibits leaf size by controlling cell proliferation and enlargement in soybean. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Raza, M.; Yu, H.; Sun, F.; Zhang, Y.; Lu, F.; Si, L.; Iqbal, N.; Khan, I.; Fu, F. Effect of shade treatments on morphology, photosynthetic and chlorophyll fluorescence characteristics of soybeans (Glycine max L. Merr.). Appl. Ecol. Environ. Res. 2019, 17, 2551–2569. [Google Scholar] [CrossRef]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Khalid, M.H.B.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L. The Influence of Light Intensity and Leaf Movement on Photosynthesis Characteristics and Carbon Balance of Soybean. Front. Plant Sci. 2018, 9, 1952. [Google Scholar] [CrossRef]
- Dai, J.; Dong, H. Intensive cotton farming technologies in China: Achievements, challenges and countermeasures. Field Crop. Res. 2014, 155, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Amin, A.; Nasim, W.; Mubeen, M.; Nadeem, M.; Ali, L.; Hammad, H.M.; Sultana, S.R.; Jabran, K.; ur Rehman, M.H.; Ahmad, S. Optimizing the phosphorus use in cotton by using CSM-CROPGRO-cotton model for semi-arid climate of Vehari-Punjab, Pakistan. Environ. Sci. Pollut. Res. 2017, 24, 5811–5823. [Google Scholar] [CrossRef]
- Divito, G.A.; Sadras, V.O. How do phosphorus, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and pasture legumes? A meta-analysis. Field Crop. Res. 2014, 156, 161–171. [Google Scholar] [CrossRef]
- Ma, Q.; Scanlan, C.; Bell, R.; Brennan, R. The dynamics of potassium uptake and use, leaf gas exchange and root growth throughout plant phenological development and its effects on seed yield in wheat (Triticum aestivum) on a low-K sandy soil. Plant Soil 2013, 373, 373–384. [Google Scholar] [CrossRef]
- Abbadi, J.; Gerendás, J.; Sattelmacher, B. Effects of potassium supply on growth and yield of safflower as compared to sunflower. J. Plant Nutr. Soil Sci. 2008, 171, 272–280. [Google Scholar] [CrossRef]
- Fageria, N.; Melo, L. Agronomic evaluation of dry bean genotypes for potassium use efficiency. J. Plant Nutr. 2014, 37, 1899–1912. [Google Scholar] [CrossRef]
- Dai, X.; Ouyang, Z.; Li, Y.; Wang, H. Variation in yield gap induced by nitrogen, phosphorus and potassium fertilizer in North China Plain. PLoS ONE 2013, 8, e82147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Liu, X.; He, P.; Johnston, A.M.; Zhao, S.; Qiu, S.; Zhou, W. Yield gap, indigenous nutrient supply and nutrient use efficiency for maize in China. PLoS ONE 2015, 10, e0140767. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, M.; Li, J.; Lu, J.; Ren, T.; Cong, R.; Fahad, S.; Li, X. Effects of fertilization on crop production and nutrient-supplying capacity under rice-oilseed rape rotation system. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Lambers, H.; Poorter, H. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 1992; Volume 23, pp. 187–261. [Google Scholar]
- Garnier, E.; Gobin, O.; Poorter, H. Nitrogen productivity depends on photosynthetic nitrogen use efficiency and on nitrogen allocation within the plant. Ann. Bot. 1995, 76, 667–672. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Shen, J.; Yuan, L.; Zhang, J.; Li, H.; Bai, Z.; Chen, X.; Zhang, W.; Zhang, F. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Dunbabin, V.M.; Diggle, A.J.; Siddique, K.H.; Rengel, Z. Phosphorus starvation boosts carboxylate secretion in P-deficient genotypes of Lupinus angustifolius with contrasting root structure. Crop Pasture Sci. 2013, 64, 588–599. [Google Scholar] [CrossRef]
- Minjian, C.; Haiqiu, Y.; Hongkui, Y.; Chunji, J. Difference in tolerance to potassium deficiency between two maize inbred lines. Plant Prod. Sci. 2007, 10, 42–46. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Amtmann, A.; Hammond, J.P.; Armengaud, P.; White, P.J. Nutrient sensing and signalling in plants: Potassium and phosphorus. Adv. Bot. Res. 2005, 43, 209–257. [Google Scholar]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Lv, X.; Yang, J.; Chen, B.; Zhao, W.; Meng, Y.; Wang, Y.; Zhou, Z.; Oosterhuis, D.M. Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crop. Res. 2016, 191, 139–149. [Google Scholar] [CrossRef]
- Chen, X.; Shi, C.; Yin, Y.; Wang, Z.; Shi, Y.; Peng, D.; Ni, Y.; Cai, T. Relationship between lignin metabolism and lodging resistance in wheat. Acta Agron. Sin. 2011, 37, 1616–1622. [Google Scholar] [CrossRef]
- Wei, J.; Li, C.; Li, Y.; Jiang, G.; Cheng, G.; Zheng, Y. Effects of external potassium (K) supply on drought tolerances of two contrasting winter wheat cultivars. PLoS ONE 2013, 8, e69737. [Google Scholar] [CrossRef] [PubMed]
- Amanullah, A.; Iqbal, A.; Iqbal, M. Impact of potassium rates and their application time on dry matter partitioning, biomass and harvest index of maize (Zea mays) with and without cattle dung application. Emirates J. Food Agric. 2015, 27, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, C.C.; Yan, P.; Chen, X.P.; Yang, J.C.; Zhang, F.S.; Cui, Z.L. Potassium requirement in relation to grain yield and genotypic improvement of irrigated lowland rice in China. J. Plant Nutr. Soil Sci. 2013, 176, 400–406. [Google Scholar] [CrossRef]
- Amanullah, M.H.K. Difference in dry matter accumulation with variable rates of sulphur and potassium application under calcareous soils in Brassica napus vs. B. juncea. J. Oilseed Brassica 2016, 1, 241–248. [Google Scholar]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Damon, P.M.; Rengel, Z. Wheat genotypes differ in potassium efficiency under glasshouse and field conditions. Aust. J. Agric. Res. 2007, 58, 816–825. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2012; Volume 89. [Google Scholar]
- Mondal, A.; Miah, S. Effect of K on bacterial blight (BB) development. Int. Rice Res. Newsl. (Philipp.) 1985, 10, 12–13. [Google Scholar]
- Sindhan, G.; Parashar, R. Influence of some nutrients on phenol and carbohydrate contents related to bacterial blight in rice. Plant Dis. Res. 1986, 1, 65–68. [Google Scholar]
- Gou, F.; Van Ittersum, M.K.; Couëdel, A.; Zhang, Y.; Wang, Y.; van der Putten, P.E.; Zhang, L.; van der Werf, W. Intercropping with wheat lowers nutrient uptake and biomass accumulation of maize, but increases photosynthetic rate of the ear leaf. AoB Plants 2018, 10, ply010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory Methods of Soil and Plant Analysis: A Working Manual, 2nd ed.; Sacred Africa: Nairobi, Kenya, 2002; Volume 21. [Google Scholar]
- Olsen, S.; Sommers, L. Methods of Soil Analysis, Part 2, 2nd ed.; Agron. Monogr. 9; Page, A.L., Ed.; ASA and SSSA: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Huoyan, W.; Cheng, W.; Ting, L.; Jianmin, Z.; Xiaoqin, C. Can nonexchangeable potassium be differentiated from structural potassium in soils? Pedosphere 2016, 26, 206–215. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954; Circular No. 939.
- Jackson, M. Soil Chemical Analysis; 2nd Indian Print; Prentice-Hall of India Pvt. Ltd.: New Delhi, India, 1973; pp. 38–336. [Google Scholar]
- Pietsch, D.; Mabit, L. Terrace soils in the Yemen Highlands: Using physical, chemical and radiometric data to assess their suitability for agriculture and their vulnerability to degradation. Geoderma 2012, 185, 48–60. [Google Scholar] [CrossRef]
- Liu, W.; Deng, Y.; Hussain, S.; Zou, J.; Yuan, J.; Luo, L.; Yang, C.; Yuan, X.; Yang, W. Relationship between cellulose accumulation and lodging resistance in the stem of relay intercropped soybean [Glycine max (L.) Merr.]. Field Crop. Res. 2016, 196, 261–267. [Google Scholar] [CrossRef]
- Yang, F.; Liao, D.; Wu, X.; Gao, R.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; Liu, J.; et al. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercropping systems. Field Crop. Res. 2017, 203, 16–23. [Google Scholar] [CrossRef]
- Li, L.; Sun, J.; Zhang, F.; Li, X.; Yang, S.; Rengel, Z. Wheat/maize or wheat/soybean strip intercropping: I. Yield advantage and interspecific interactions on nutrients. Field Crop. Res. 2001, 71, 123–137. [Google Scholar] [CrossRef]
- Xia, H.-Y.; Wang, Z.-G.; Zhao, J.-H.; Sun, J.-H.; Bao, X.-G.; Christie, P.; Zhang, F.-S.; Li, L. Contribution of interspecific interactions and phosphorus application to sustainable and productive intercropping systems. Field Crop. Res. 2013, 154, 53–64. [Google Scholar] [CrossRef]
- Raza, M.A.; Feng, L.Y.; Manaf, A.; Wasaya, A.; Ansar, M.; Hussain, A.; Khalid, M.H.B.; Iqbal, N.; Xi, Z.J.; Chen, Y.K. Sulphur application increases seed yield and oil content in sesame seeds under rainfed conditions. Field Crop. Res. 2018, 218, 51–58. [Google Scholar] [CrossRef]
- Raza, M.A.; Feng, L.Y.; Iqbal, N.; Manaf, A.; Khalid, M.H.B.; Ur Rehman, S.; Wasaya, A.; Ansar, M.; Billah, M.; Yang, F. Effect of sulphur application on photosynthesis and biomass accumulation of sesame varieties under rainfed conditions. Agronomy 2018, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Zhao, D.; Oosterhuis, D.; Bednarz, C. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Reddy, K.R.; Zhao, D. Interactive effects of elevated CO2 and potassium deficiency on photosynthesis, growth, and biomass partitioning of cotton. Field Crop. Res. 2005, 94, 201–213. [Google Scholar] [CrossRef]
- Pettigrew, W. Effects of K on plant metabolism. Physiol. Plant 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Saur, E.; Constantin, J.; Porté, A.; Jordan-Meille, L. Effect of carbon assimilation on dry weight production and partitioning during vegetative growth. Plant Soil 2009, 324, 329–343. [Google Scholar] [CrossRef]
- Pettigrew, W.; Gerik, T. Cotton leaf photosynthesis and carbon metabolism. Adv. Agron. 2007, 94, 209–236. [Google Scholar]
- Dreccer, M.; Grashoff, C.; Rabbinge, R. Source-sink ratio in barley (Hordeum vulgare L.) during grain filling: Effects on senescence and grain protein concentration. Field Crop. Res. 1997, 49, 269–277. [Google Scholar] [CrossRef]
- Hao, M.B.; Wang, K.J.; Dong, S.T.; Zhang, J.W.; Li, D.H.; Liu, P.; Yang, J.S.; Liu, J.G. Leaf redundancy of high-yielding maize (Zea may L.) and its effects on maize yield and photosynthesis. Yingyong Shengtai Xuebao 2010, 21, 344–350. [Google Scholar]
- Chen, P.; Du, Q.; Liu, X.; Zhou, L.; Hussain, S.; Lei, L.; Song, C.; Wang, X.; Liu, W.; Yang, F. Effects of reduced nitrogen inputs on crop yield and nitrogen use efficiency in a long-term maize-soybean relay strip intercropping system. PLoS ONE 2017, 12, e0184503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Ramna, S. Response of sorghum to nitrogen and potassium in Alfisol. J. Potash. Res. 1993, 9, 171–175. [Google Scholar]
- Hall, D.O.; Scurlock, J.; Bolhàr-Nordenkampf, H.; Leegood, R.C.; Long, S. Photosynthesis and Production in a Changing Environment: A Field and Laboratory Manual; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Liu, G.; Hou, P.; Xie, R.; Ming, B.; Wang, K.; Liu, W.; Yang, Y.; Xu, W.; Chen, J.; Li, S. Nitrogen uptake and response to radiation distribution in the canopy of high-yield maize. Crop Sci. 2019, 59, 1236–1247. [Google Scholar] [CrossRef]
- Høgh-Jensen, H.; Pedersen, M.B. Morphological plasticity by crop plants and their potassium use efficiency. J. Plant Nutr. 2003, 26, 969–984. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Kawakami, E.M.; Pettigrew, W.T. The physiology of potassium in crop production. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2014; Volume 126, pp. 203–233. [Google Scholar]
- Du, Q.; Zhao, X.; Jiang, C.; Wang, X. Effect of potassium deficiency on root growth and nutrient uptake in Maize (Zea mays L.). Agric. Sci. 2017, 8, 1263–1277. [Google Scholar] [CrossRef] [Green Version]
- Bednarz, C.; Oosterhuis, D.; Evans, R. Leaf photosynthesis and carbon isotope discrimination of cotton in response to potassium deficiency. Environ. Exp. Bot. 1998, 39, 131–139. [Google Scholar] [CrossRef]
- Li, L.; Sun, J.; Zhang, F.; Li, X.; Rengel, Z.; Yang, S. Wheat/maize or wheat/soybean strip intercropping: II. Recovery or compensation of maize and soybean after wheat harvesting. Field Crop. Res. 2001, 71, 173–181. [Google Scholar] [CrossRef]
- Ahmad, M.; Riaz, A.; Ishaque, M.; Malik, A. Response of maize hybrids to varying potassium application in Pakistan. Pak. J. Agric. Sci. 2009, 46, 179–184. [Google Scholar]
- Sadiq, S.; Jan, A. Effect of Graded Application of Potash on Kharif Maize Sown at Different fErtility Levels. Master’s Thesis, Department of Agronomy, The University of Agriculture, Peshawar, Pakistan, 2001. [Google Scholar]
- Uddin, S.; Sarkar, M.; Rahman, M. Effect of nitrogen and potassium on yield of dry direct seeded rice cv. NERICA 1 in aus season. Int. J. Agron. Plant Prod. 2013, 4, 69–75. [Google Scholar]
- Cheema, M.A.; Wahid, M.; Sattar, A.; Rasul, F.; Saleem, M. Influence of different levels of potassium on growth, yield and quality of canola (Brassica napus L.) cultivars. Pak. J. Agric. Sci. 2012, 49, 163–168. [Google Scholar]
- Yang, F.; Wang, X.; Liao, D.; Lu, F.; Gao, R.; Liu, W.; Yong, T.; Wu, X.; Du, J.; Liu, J. Yield response to different planting geometries in maize–soybean relay strip intercropping systems. Agron. J. 2015, 107, 296–304. [Google Scholar] [CrossRef]
- Agegnehu, G.; Ghizaw, A.; Sinebo, W. Yield performance and land-use efficiency of barley and faba bean mixed cropping in Ethiopian highlands. Eur. J. Agron. 2006, 25, 202–207. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Treatment | TDM (t ha−1) | TNA (g plant−1) | TPA (g plant−1) | TKA (g plant−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Maize | Soybeans | Maize | Soybeans | Maize | Soybeans | Maize | Soybeans | ||
| 2018 | T0 | 11.40 ± 0.05 d | 3.03 ± 0.16 d | 1.52 ± 0.02 c | 0.98 ± 0.04 d | 0.36 ± 0.02 d | 0.17 ± 0.01 d | 2.93 ± 0.03 d | 0.58 ± 0.01 d |
| T1 | 13.07 ± 0.11 c | 3.77 ± 0.05 c | 1.87 ± 0.04 b | 1.29 ± 0.05 c | 0.41 ± 0.01 c | 0.19 ± 0.05 c | 3.09 ± 0.07 c | 0.71 ± 0.02 c | |
| T2 | 15.28 ± 0.18 a | 4.46 ± 0.08 b | 2.58 ± 0.01 a | 1.58 ± 0.06 b | 0.52 ± 0.01 a | 0.23 ± 0.08 b | 3.92 ± 0.06 a | 0.85 ± 0.01 b | |
| SM | 14.51 ± 0.07 b | - | 1.90 ± 0.03 b | - | 0.49 ± 0.06 b | - | 3.43 ± 0.04 b | - | |
| SS | - | 5.97 ± 0.04 a | - | 1.89 ± 0.03 a | - | 0.29 ± 0.03 a | - | 1.01 ± 0.04 a | |
| LSD | 0.40 | 0.29 | 0.11 | 0.17 | 0.046 | 0.14 | 0.2 | 0.05 | |
| 2019 | T0 | 11.81 ± 0.06 d | 3.12 ± 0.02 d | 1.58 ± 0.01 d | 1.04 ± 0.01 d | 0.39 ± 0.01 d | 0.19 ± 0.01 d | 2.97 ± 0.03 d | 0.63 ± 0.02 d |
| T1 | 13.32 ± 0.03 c | 3.85 ± 0.03 c | 2.01 ± 0.08 c | 1.42 ± 0.02 c | 0.46 ± 0.02 c | 0.21 ± 0.06 c | 3.29 ± 0.02 c | 0.77 ± 0.04 c | |
| T2 | 15.73 ± 0.04 a | 4.54 ± 0.01 b | 2.72 ± 0.02 a | 1.65 ± 0.02 b | 0.56 ± 0.03 a | 0.26 ± 0.02 b | 4.19 ± 0.02 a | 0.93 ± 0.02 b | |
| SM | 15.51 ± 0.20 b | - | 2.17 ± 0.02 b | - | 0.52 ± 0.02 b | - | 3.81 ± 0.01 b | - | |
| SS | - | 6.05 ± 0.07 a | - | 1.97 ± 0.03 a | - | 0.34 ± 0.04 a | - | 1.12 ± 0.03 a | |
| LSD | 0.33 | 0.13 | 0.13 | 0.61 | 0.043 | 0.32 | 0.09 | 0.05 | |
| Years | Treatments | Kernels plant−1 | Seed Number plant−1 | Seed Weight plant−1 (g) | Seed Index (1000 Seed Weight) | Grain-Yield t ha−1 | LER | Total LER | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Maize | Soybeans | Maize | Soybeans | Maize | Soybeans | Maize | Soybeans | |||||
| 2018 | T0 | 394 ± 0.02 d | 74.22 ± 0.13 d | 82.88 ± 0.17 d | 9.25 ± 0.42 d | 246.05 ± 0.26 d | 178.42 ± 0.22 d | 4.973 ± 0.09 d | 0.925 ± 0.02 d | 0.81 ± 0.02 c | 0.46 ± 0.04 c | 1.28 ± 0.01 c |
| T1 | 402 ± 0.01 c | 80.59 ± 0.31 c | 85.82 ± 0.07 c | 10.87 ± 0.57 c | 253.57 ± 0.17 c | 185.12 ± 0.01 c | 5.149 ± 0.03 c | 1.087 ± 0.07 c | 0.84 ± 0.03 b | 0.54 ± 0.02 b | 1.39 ± 0.02 b | |
| T2 | 432 ± 0.06 a | 91.72 ± 0.07 b | 104.87 ± 0.20 a | 12.65 ± 0.59 b | 288.48 ± 0.22 a | 194.75 ± 0.15 b | 6.292 ± 0.02 a | 1.265 ± 0.06 b | 1.03 ± 0.49 a | 0.63 ± 0.02 a | 1.66 ± 0.02 a | |
| SM | 409 ± 0.08 b | - | 101.77 ± 0.05 b | - | 261.29 ± 0.05 b | - | 6.106 ± 0.04 b | - | - | - | - | |
| SS | - | 102.08 ± 0.17 a | - | 20.02 ± 0.43 a | - | 203.12 ± 0.18 a | - | 2.002 ± 0.036 a | - | - | - | |
| LSD | 0.15 | 0.36 | 0.52 | 0.09 | 0.18 | 0.61 | 0.15 | 0.03 | 0.08 | 0.02 | 0.06 | |
| 2019 | T0 | 407 ± 0.10 d | 70.91 ± 0.11 d | 89.75 ± 0.20 d | 10.13 ± 0.59 d | 252.32 ± 0.46 d | 181.27 ± 0.14 d | 5.385 ± 0.05 d | 1.013 ± 0.02 d | 0.87 ± 0.03 c | 0.51 ± 0.3 c | 1.41 ± 0.02 c |
| T1 | 419 ± 0.06 c | 77.69 ± 0.15 c | 91.62 ± 0.01 c | 11.09 ± 0.72 c | 269.07 ± 0.11 c | 189.61 ± 0.20 c | 5.497 ± 0.30 c | 1.109 ± 0.01 c | 0.89 ± 0.15 b | 0.56 ± 0.02 b | 1.44 ± 0.03 b | |
| T2 | 447 ± 0.59 a | 85.29 ± 0.17 b | 107.42 ± 0.27 a | 12.71 ± 0.25 b | 295.56 ± 0.20 a | 196.77 ± 0.05 b | 6.445 ± 0.06 a | 1.271 ± 0.05 b | 1.04 ± 0.03 a | 0.64 ± 0.05 a | 1.68 ± 0.01 a | |
| SM | 429 ± 0.08 b | - | 103.33 ± 0.17 b | - | 288.48 ± 0.34 b | 6.200 ± 0.04 b | - | - | - | - | ||
| SS | - | 97.86 ± 0.15 a | - | 15.90 ± 0.60 a | - | 199.65 ± 0.18 a | - | 1.993 ± 0.08 a | - | - | - | |
| LSD | 0.98 | 0.55 | 0.56 | 0.19 | 0.66 | 0.57 | 0.19 | 0.04 | 0.05 | 0.01 | 0.03 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.; Aftab, S.; Hussain, S.; Nazir Cheema, H.; Liu, W.; Yang, F.; Yang, W. Nutrient Accumulation and Distribution Assessment in Response to Potassium Application under Maize–Soybean Intercropping System. Agronomy 2020, 10, 725. https://doi.org/10.3390/agronomy10050725
Ahmed A, Aftab S, Hussain S, Nazir Cheema H, Liu W, Yang F, Yang W. Nutrient Accumulation and Distribution Assessment in Response to Potassium Application under Maize–Soybean Intercropping System. Agronomy. 2020; 10(5):725. https://doi.org/10.3390/agronomy10050725
Chicago/Turabian StyleAhmed, Aftab, Samina Aftab, Sadam Hussain, Hafsa Nazir Cheema, Weigou Liu, Feng Yang, and Wenyu Yang. 2020. "Nutrient Accumulation and Distribution Assessment in Response to Potassium Application under Maize–Soybean Intercropping System" Agronomy 10, no. 5: 725. https://doi.org/10.3390/agronomy10050725