
Conjugation of Aspergillus flavipes Taxol with Porphyrin Increases the Anticancer Activity of Taxol and Ameliorates Its Cytotoxic Effects


 ,
,
Abstract
1. Introduction
2. Results and Discussion
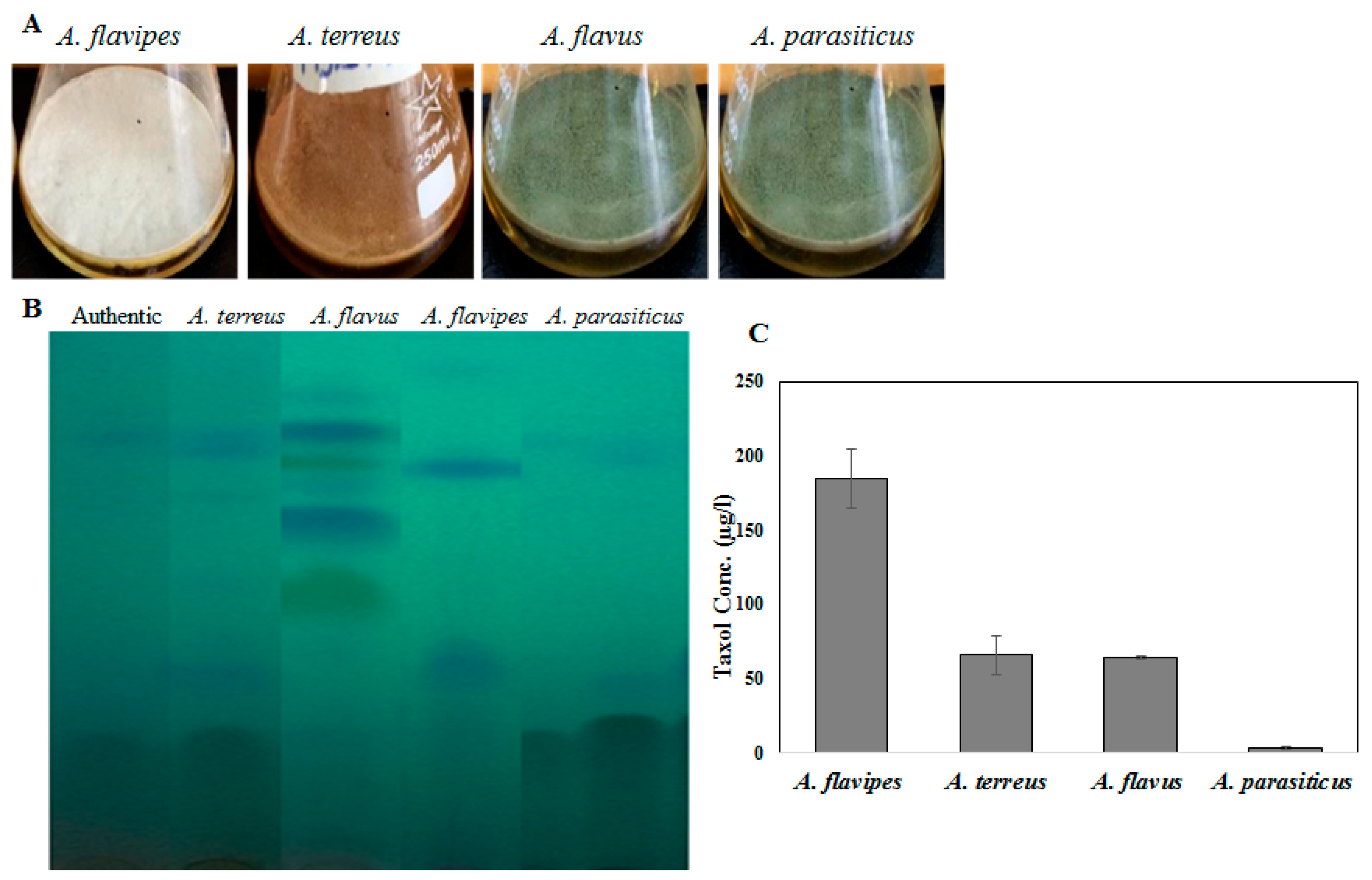
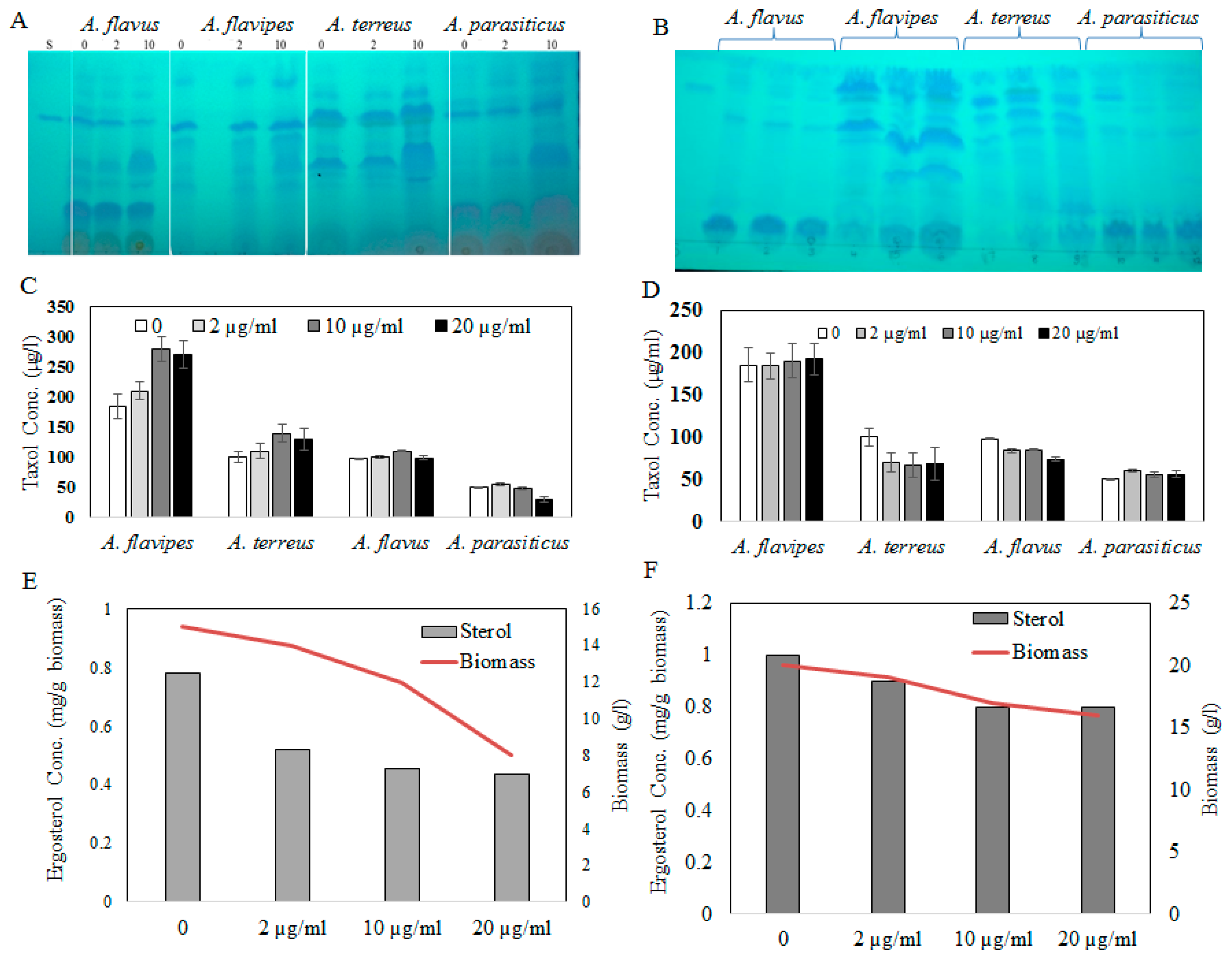
2.1. Taxol Production by the Selected Fungal Isolates in Response to Growth Inhibitors
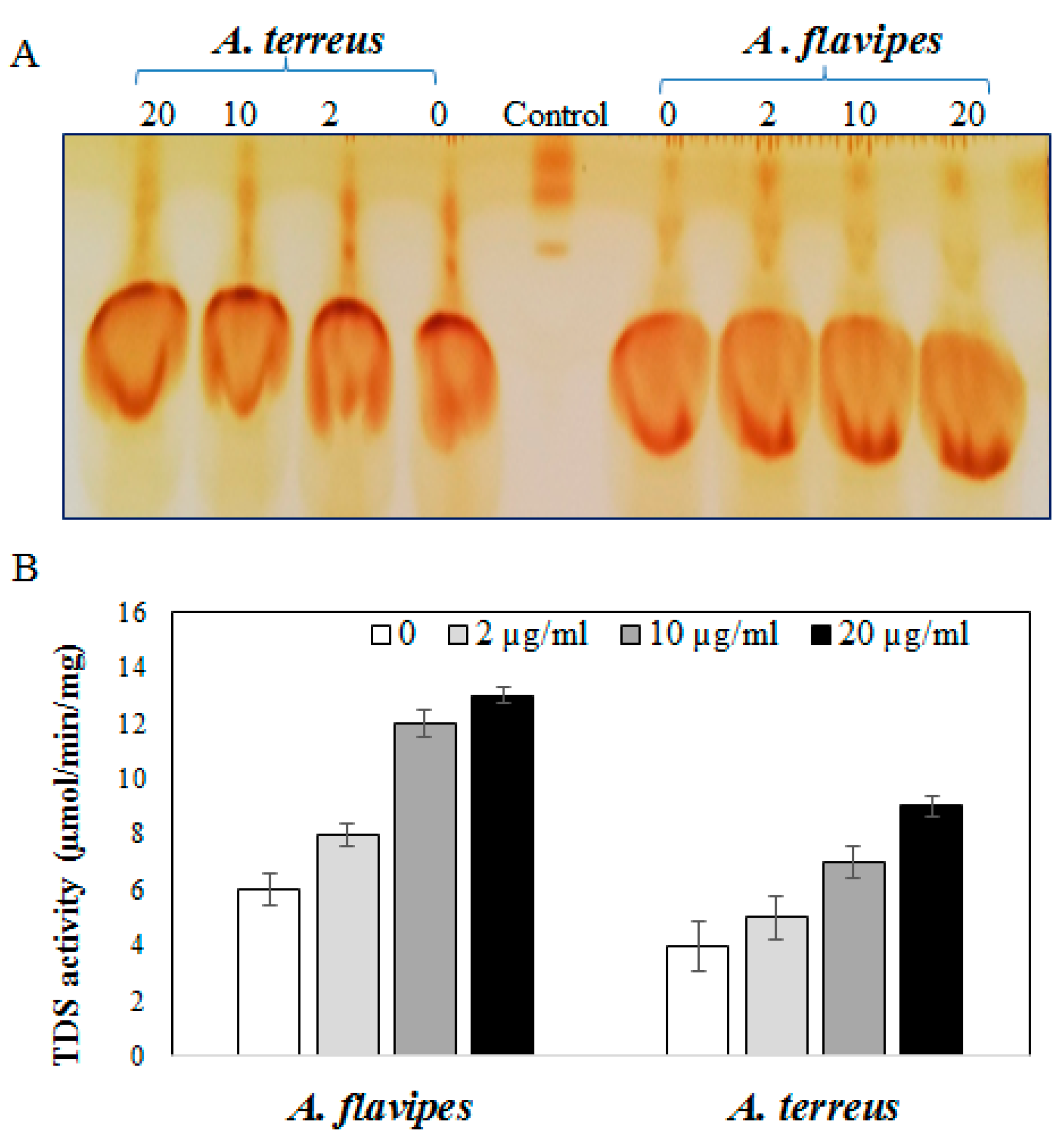
2.2. Activity of Taxadiene Synthase in Response to Sterol Biosynthesis Inhibitor
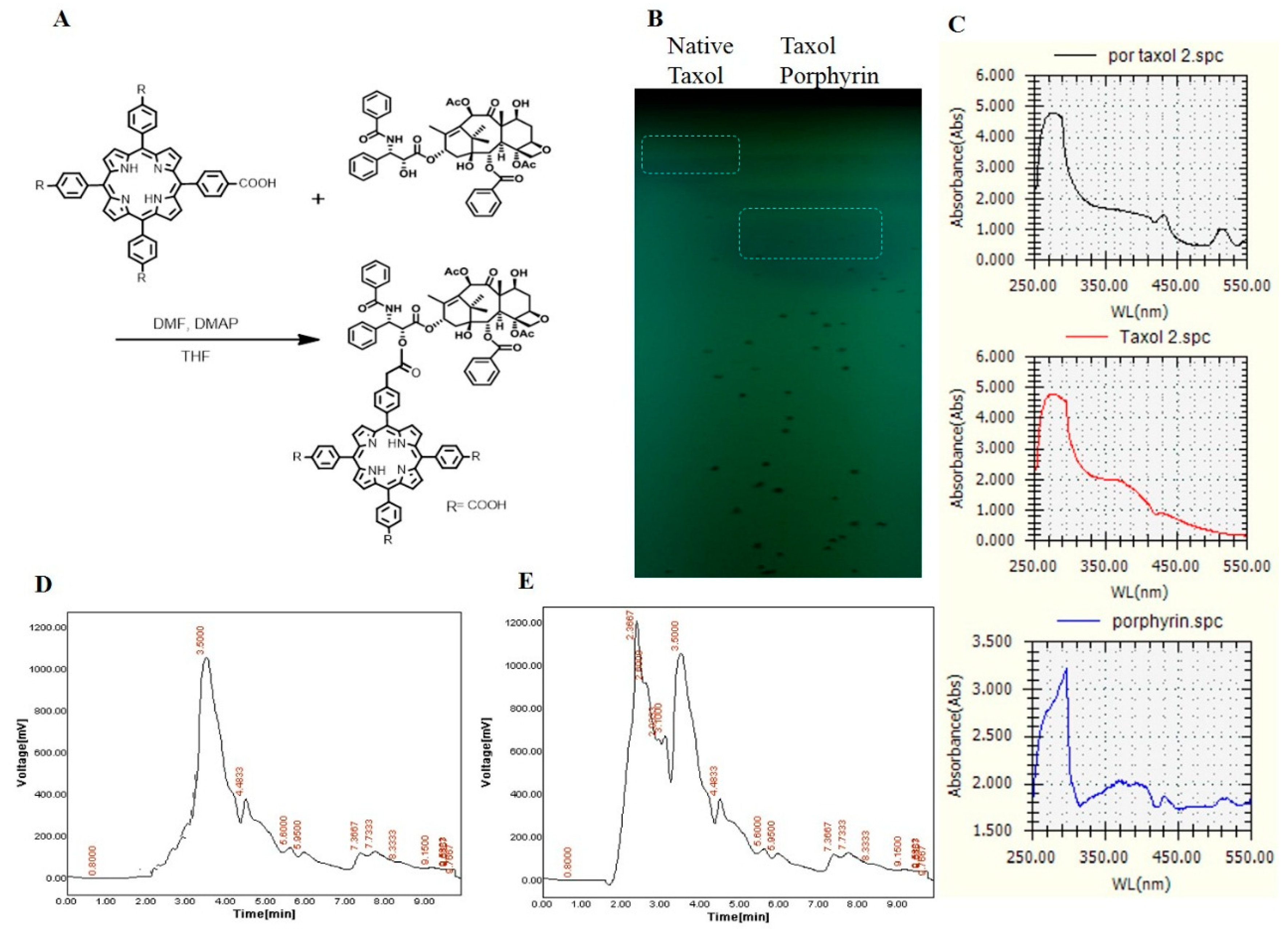
2.3. Conjugation of Taxol with Porphyrin
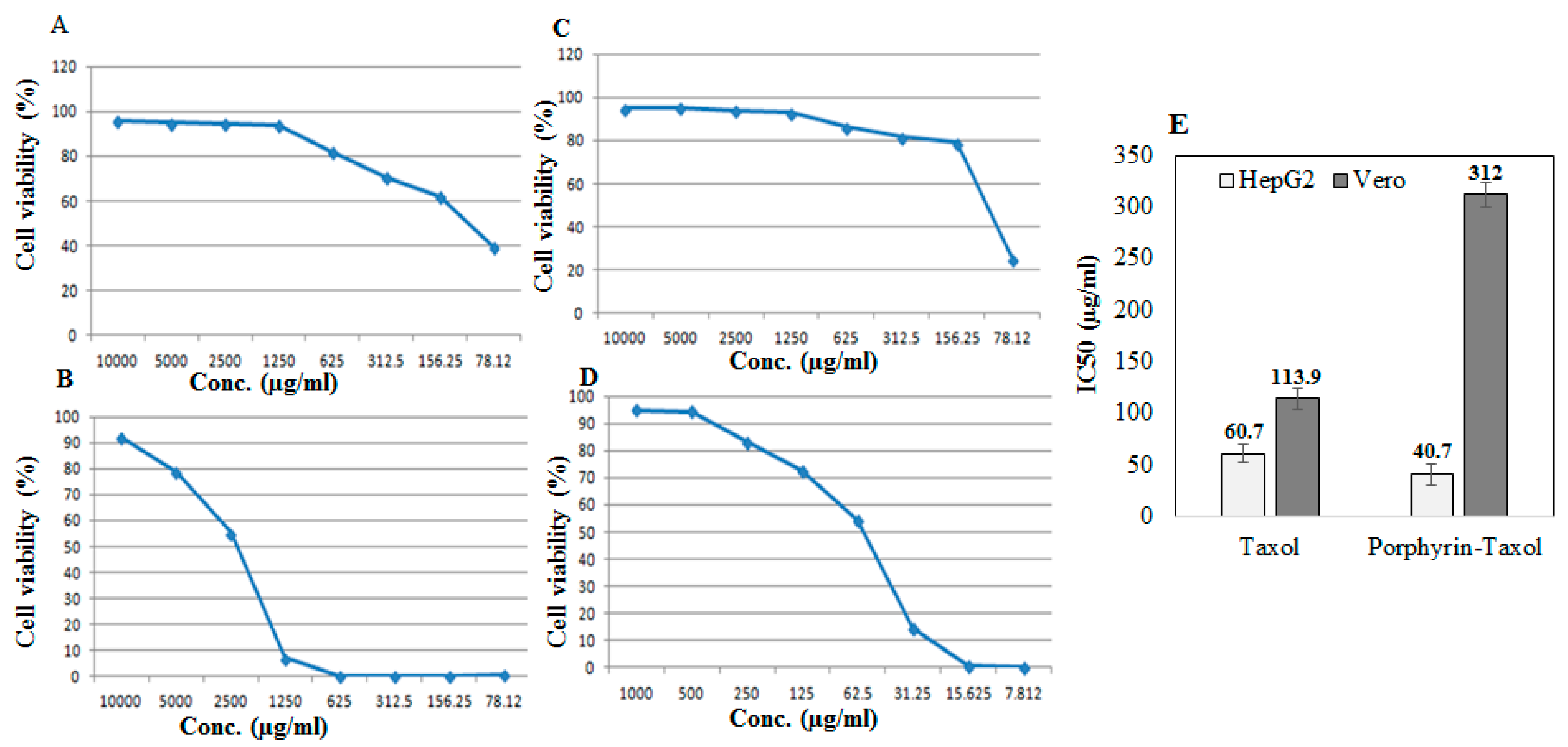
2.4. Antiproliferative Activity of Native Taxol and Taxol–Porphyrin Conjugates
2.5. Biochemical Profile of Mice in Response to Native and Taxol–Porphyrin Conjugates
3. Materials and Methods
3.1. Fungal Isolates, Media Preparation, and Screening for Taxol Production
3.2. Taxol Purification by Preparative-HPLC System
3.3. TLC, UV, and HPLC Analyses
3.4. LC–MS Analyses
3.5. Eliciting the Taxol Yield of Selected Fungal Isolate with Growth Inhibitors
3.6. Taxadiene Synthase (TDS) Activity Assay
3.7. Total Sterol Assay
3.8. Conjugation of Taxol with Porphyrin
3.9. Antiproliferative Activity of Native Taxol and Taxol–Porphyrin Conjugates
3.10. Biochemical Profile of Mice in Response to Native and Taxol–Porphyrin Conjugates
3.11. Deposition of the Fungal Isolates
3.12. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- El-Sayed, A.S.A.; Ali, D.M.I.; Yassin, M.A.; Zayed, R.A.; Ali, G.S. Sterol inhibitor “Fluconazole” enhance the Taxol yield and molecular expression of its encoding genes cluster from Aspergillus flavipes. Process Biochem. 2019, 76, 55–67. [Google Scholar] [CrossRef]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A.T. Plant antitumor agents. VI. The isolation and strcture of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef] [PubMed]
- Suffness, M.; Cordell, G.A. Antitumor alkaloids. Alkaloids 1985, 25, 280–288. [Google Scholar]
- Rowinski, E.K.; Cazenave, L.A.; Donehower, F.C. Taxol: A novel investigational antimicrotubule agent. J. Natl. Cancer Inst. 1990, 82, 1247–1259. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Abdel-Ghany, S.E.; Ali, G.S. Genome editing approaches: Manipulating of lovastatin and taxol synthesis of filamentous fungi by CRISPR/Cas9 system. Appl. Microbiol. Biotechnol. 2017, 101, 3953–3976. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Safan, S.; Mohamed, N.Z.; Shaban, L.; Ali, G.S.; Sitohy, M.Z. Induction of Taxol biosynthesis by Aspergillus terreus, endophyte of Podocarpus gracilior Pilger upon intimate interaction with the plant endogenous microbes. Process Biochem. 2018, 71, 31–40. [Google Scholar] [CrossRef]
- Barboni, L.; Gariboldi, P.; Torregiani, E.; Appendino, G.; Gabetta, B.; Zini, G.; Bombardelli, E. Taxanes from the needles of Taxus wallichiana. Phytochemistry 1993, 33, 145–150. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Mohamed, N.M.; Safan, S.; Yassin, M.A.; Shaban, L.; Shindia, A.; Ali, G.S.; Sitohy, M. Restoring the Biosynthetic Machinery of Taxol of Aspergillus terreus via cocultivation with the endophytic microbiome of Podocarpus gracilior Pilger. Sci. Rep. 2019, 9, 11534. [Google Scholar] [CrossRef]
- Holton, R.A.; Somoza, C.; Kim, H.B. First total synthesis of taxol.1. Functionalization of the B ring. J. Am. Chem. Soc. 1994, 116, 1597–1604. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Yang, Z.; Liu, J.J.; Ueno, H.; Nantermet, P.G.; Guy, R.K.; Claiborne, C.F.; Renaud, J.; Couladouros, E.A.; Paulvannan, K. Total synthesis of taxol. Nature 1994, 367, 593–594. [Google Scholar] [CrossRef]
- Borman, S. Scientists mobilize to increase supply of the anticancer drug taxol. Chem. Eng. News 1994, 2, 11–18. [Google Scholar] [CrossRef]
- Borman, S. Total synthesis of anticancer agent taxol achieved by two different routes. Chem. Eng. News 1994, 2, 32–34. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D.; Grothaus, P.; Bignami, G. The search for a taxol producing microorganism among the endophytic Fungi of the Pacific Yew, Taxus brevifolia. J. Nat. Prod. 1995, 58, 1315–1324. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Baldi, A.; Farkya, S.; Jain, A.; Gupta, N.; Mehra, R.; Datta, V. Enhanced production of podophyllotoxins by co-culture of transformed Linum album cells with plant growth-promoting fungi. Pure Appl. Chem. 2009, 82, 227–241. [Google Scholar] [CrossRef]
- Li, J.Y.; Sidhu, R.S.; Ford, E.; Hess, W.M.; Strobel, G.A. The induction of taxol production in the endophytic fungus- Periconia sp. from Torreya grandifolia. J. Ind. Microbiol. 1998, 20, 259–264. [Google Scholar]
- Zheng, X.; Li, Z.; Chen, L.; Xie, Z.; Jing, X. Self-Assembly of Porphyrin–Paclitaxel Conjugates Into Nanomedicines: Enhanced Cytotoxicity due to Endosomal Escape. Chem. Asian J. 2016, 11, 1780–1784. [Google Scholar] [CrossRef]
- Dougherty, T.J.; Gomer, C.J.; Henderson, B.W.; Jori, G.; Kessel, D.; Korbelik, M.; Johan, M.; Qian, P. Photodynamic therapy. J. Natl. Cancer Inst. 1998, 90, 889–905. [Google Scholar] [CrossRef]
- Huynh, E.; Lovell, J.F.; Helfield, B.L.; Jeon, M.; Kim, C.; Goertz, D.E.; Wilson, B.C.; Zheng, G. Porphyrin shell microbubbles with intrinsic ultrasound and photoacoustic properties. J. Am. Chem. Soc. 2012, 134, 16464–16467. [Google Scholar] [CrossRef]
- Exposito, O.; Bonfill, M.; Moyano, E.; Onrubia, M.; Mirjalili, M.H.; Cusido, R.M.; Palazon, J. Biotechnological production of taxol and related taxoids: Current state and prospects. Anticancer Agents Med. Chem. 2009, 9, 109–121. [Google Scholar] [CrossRef]
- Nims, E.; Dubois, C.P.; Roberts, S.C.; Walker, E.L. Expression profiling of genes involved in paclitaxel biosynthesis for targeted metabolic engineering. Metab. Eng. 2006, 8, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Bringi, V.; Kakrade, P.G.; Prince, C.L.; Roach, B. Enhanced Production of Taxanes by Cell Cultures of Taxus Species. Patent WO1997044476A1, 27 November 1997. Available online: https://patents.google.com/patent/WO1997044476A1/en (accessed on 9 January 2020).
- Chong, H.S.; Campbell, L.; Padula, M.P.; Hill, C.; Harry, E.; Li, S.S.; Wilkins, M.R.; Herbert, B.; Carter, D. Time-course proteome analysis reveals the dynamic response of Cryptococcus gattii cells to Fluconazole. PloS ONE 2012. [Google Scholar] [CrossRef]
- Fett-Neto, A.G.; Melanson, S.J.; Nicholson, S.A.; Pennington, J.J.; DiCosmo, F. Improved taxol yield by aromatic carboxylic acid and amino acid feeding to cell cultures of Taxus cuspidata. Biotechnol. Bioeng. 1994, 44, 967–971. [Google Scholar] [CrossRef]
- Siegel, M.R. Sterol-Inhibiting Fungicides: Effects on Sterol Biosynthesis and sites of action. Plant Dis. 1981, 65, 986–989. [Google Scholar] [CrossRef]
- Nanda, R.; Sasmal, A.; Nayak, P.L. Preparation and characterization of chitosan–polylactide composites blended with Cloisite 30B for control release of the anticancer drug paclitaxel. Carbohydr. Polym. 2011, 83, 988–994. [Google Scholar] [CrossRef]
- Brigger, I.; Dubernet, C.; Couvreur, P. Nanoparticles in cancer therapy and diagnosis. Adv. Drug Deliv. Rev. 2013, 64, 24–36. [Google Scholar] [CrossRef]
- Li, Y.; Shi, J. Hollow-structured mesoporous materials: Chemical synthesis, functionalization and applications. Adv. Mater. 2014, 26, 3176–3205. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.M.; Chen, Y.; Chen, J.-T.; Liu, Y. Polysaccharide-based Noncovalent Assembly for Targeted Delivery of Taxol. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Raper, K.B.; Fennell, D.I. The Genus Aspergillus; Williams and Wilkins: Philadelphia, PA, USA, 1965. [Google Scholar]
- El-Sayed, A.S.A.; Khalaf, S.A.; Abdel Hamid, G.; El-Batrik, M.I. Screening, morphological and molecular identification of cystathionine γ-lyase producing fungi. Acta Biol. Hung. 2015, 66, 119–132. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Iqrar, I.; Ali, R.; Norman, D.; Brennan, M.; Ali, G.S. A glucanolytic Pseudomonas sp. associated with Smilax bona-nox L. displays strong activity against Phytophthora parasitica. Microbiol. Res. 2018, 207, 140–152. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Ruff, L.E.; Ghany, S.E.A.; Ali, S.E.A.; Esener, S. Molecular and spectroscopic characterization of Aspergillus flavipes and Pseudomonas putida L-Methionine γ-Lyase in vitro. Appl. Biochem. Biotechnol. 2017, 181, 1513–1532. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Shindia, A.A.; AbouZaid, A.A.; Yassin, A.M.; Ali, G.S.; Sitohy, M. Biochemical characterization of peptidylarginine deiminase-like orthologs from thermotolerant Emericella dentata and Aspergillus nidulans. Enzym. Microb. Technol. 2019, 124, 41–53. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; George, N.M.; Bolbol, A.A.; Mohamed, M.S. Purification and biochemical characterization of Aspergillus terreus ornithine decarboxylase: Curcumin is a potent enzyme inhibitor. Molecules 2019, 24, 2756. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Ali, G.S. Aspergillus flavipes is a novel efficient biocontrol agent of Phytophthora parasiticus. Biol. Control 2020, 140, 104072. [Google Scholar] [CrossRef]
- Bilgrami, K.S.; Verma, R.N. Physiology of Fungi, 2nd ed.; Vikas Publishing. PVT. Ltd.: New Delhi, India, 1981; pp. 23–27. [Google Scholar]
- Heinig, U.; Scholz, S.; Jennewein, S. Getting to the bottom of Taxol biosynthesis by fungi. Fungal Divers. 2013, 60, 161–170. [Google Scholar] [CrossRef]
- Li, J.Y.; Strobel, G.; Sidhu, R.; Hess, W.M.; Ford, E.J. Endophytic taxol-producing fungi from bald cypress, Taxodium distichum. Microbiology 1996, 142, 2223–2226. [Google Scholar] [CrossRef] [PubMed]
- Hezari, M.; Lewis, N.G.; Croteau, R. Purification and characterization of taxa-4(5), 11(12)-diene synthase from Pacific yew (Taxus brevifolia) that catalyzes the first committed step of taxol biosynthesis. Arch. Biochem. Biophys. 1995, 322, 437–444. [Google Scholar] [CrossRef]
- Artz, J.; Wernimont, A.K.; Dunford, J.E.; Schapira, M.; Dong, A.; Zhao, Y.; Lew, J.; Russell, R.G.G.; Ebetino, F.H.; Oppermann, U.; et al. Molecular characterization of a novel Geranylgeranyl Pyrophosphate Synthase from Plasmodium parasites. J. Biol. Chem. 2011, 286, 3315–3322. [Google Scholar] [CrossRef]
- Breivik, O.N.; Owades, J.L. Spectrophotometric semi-microdetermination of ergosterol in yeast. Agric. Food Chem. 1957, 5, 360–363. [Google Scholar] [CrossRef]
- Fathalla, M.; Strutt, N.L.; Sampath, S.; Katsiev, K.; Hartlieb, K.J.; Bakr, O.M.; Stoddart, J.F. Porphyrinic supramolecular daisy chains incorporating pillar[5]arene-viologen host-guest interactions. Chem. Commun. 2015, 4, 10455–10458. [Google Scholar] [CrossRef]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the Porphyrin-Taxol conjugates are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | 12 h | 24 h | 48 h | ||||
|---|---|---|---|---|---|---|---|
| Control | Por–Taxol | Taxol | Por–Taxol | Taxol | Por–Taxol | Taxol | |
| Alanine amino transferase (ALT) (U/L) | 221.5 | 252 | 248 | 220 | 231 | 185 | 150 |
| Aspartate aminotransferase (AST) (U/L) | 310 | 334 | 178 | 616 | 323 | 216 | 125 |
| Alkaline phosphatase (ALP) (U/L) | 276 | 871 | 714 | 1067 | 682 | 214 | 702 |
| Total protein (g/dl) | 8.2 | 5.67 | 5.46 | 9.2 | 9.5 | 7.9 | 5.14 |
| Albumin (g/dl) | 5 | 4.73 | 3.9 | 5.72 | 6.2 | 5.58 | 3.01 |
| Globulin (g/dl) | 3.2 | 2.94 | 1.56 | 3.48 | 3.3 | 1.32 | 2.13 |
| Albumin/Globulin ratio | 1.6 | 0.93 | 2.5 | 1.64 | 1.88 | 1.95 | 1.41 |
| Urea (mg/dl) | 12.8 | 11.3 | 26 | 11.1 | 28.4 | 16.2 | 20.9 |
| Creatinine (mg/dl) | 0.8 | 0.84 | 0.46 | 0.99 | 2.09 | 0.6 | 0.85 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Sayed, A.S.A.; Fathalla, M.; Yassin, M.A.; Zein, N.; Morsy, S.; Sitohy, M.; Sitohy, B. Conjugation of Aspergillus flavipes Taxol with Porphyrin Increases the Anticancer Activity of Taxol and Ameliorates Its Cytotoxic Effects. Molecules 2020, 25, 263. https://doi.org/10.3390/molecules25020263
El-Sayed ASA, Fathalla M, Yassin MA, Zein N, Morsy S, Sitohy M, Sitohy B. Conjugation of Aspergillus flavipes Taxol with Porphyrin Increases the Anticancer Activity of Taxol and Ameliorates Its Cytotoxic Effects. Molecules. 2020; 25(2):263. https://doi.org/10.3390/molecules25020263
Chicago/Turabian StyleEl-Sayed, Ashraf S. A., Maher Fathalla, Marwa A. Yassin, Nabila Zein, Shaima Morsy, Mahmoud Sitohy, and Basel Sitohy. 2020. "Conjugation of Aspergillus flavipes Taxol with Porphyrin Increases the Anticancer Activity of Taxol and Ameliorates Its Cytotoxic Effects" Molecules 25, no. 2: 263. https://doi.org/10.3390/molecules25020263
APA StyleEl-Sayed, A. S. A., Fathalla, M., Yassin, M. A., Zein, N., Morsy, S., Sitohy, M., & Sitohy, B. (2020). Conjugation of Aspergillus flavipes Taxol with Porphyrin Increases the Anticancer Activity of Taxol and Ameliorates Its Cytotoxic Effects. Molecules, 25(2), 263. https://doi.org/10.3390/molecules25020263