
A Novel Flavonoid Kushenol Z from Sophora flavescens Mediates mTOR Pathway by Inhibiting Phosphodiesterase and Akt Activity to Induce Apoptosis in Non-Small-Cell Lung Cancer Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Structure of KZ
2.2. KZ Inhibits NSCLC Cell Proliferation
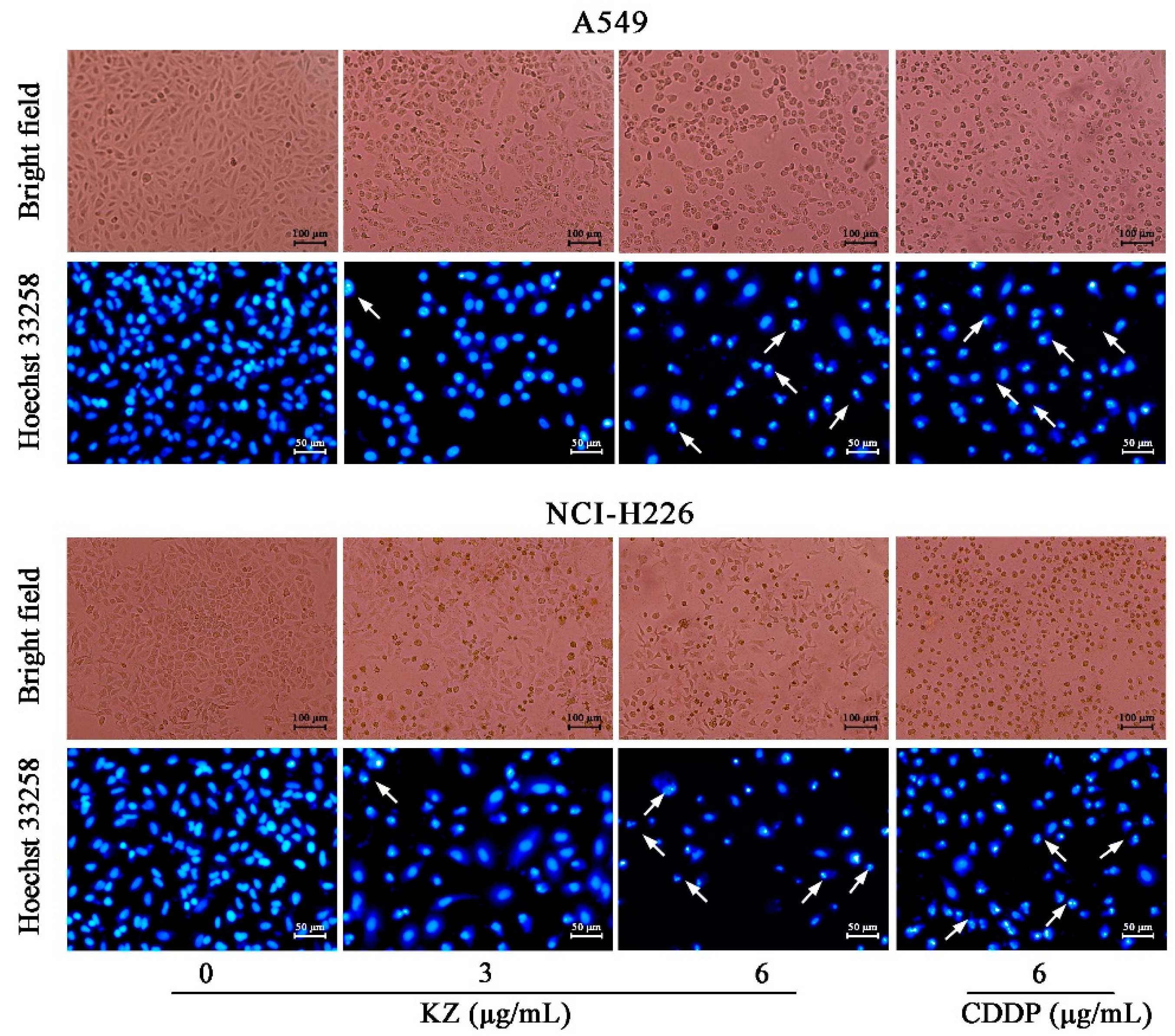
2.3. KZ Induces NSCLC Cell Apoptosis
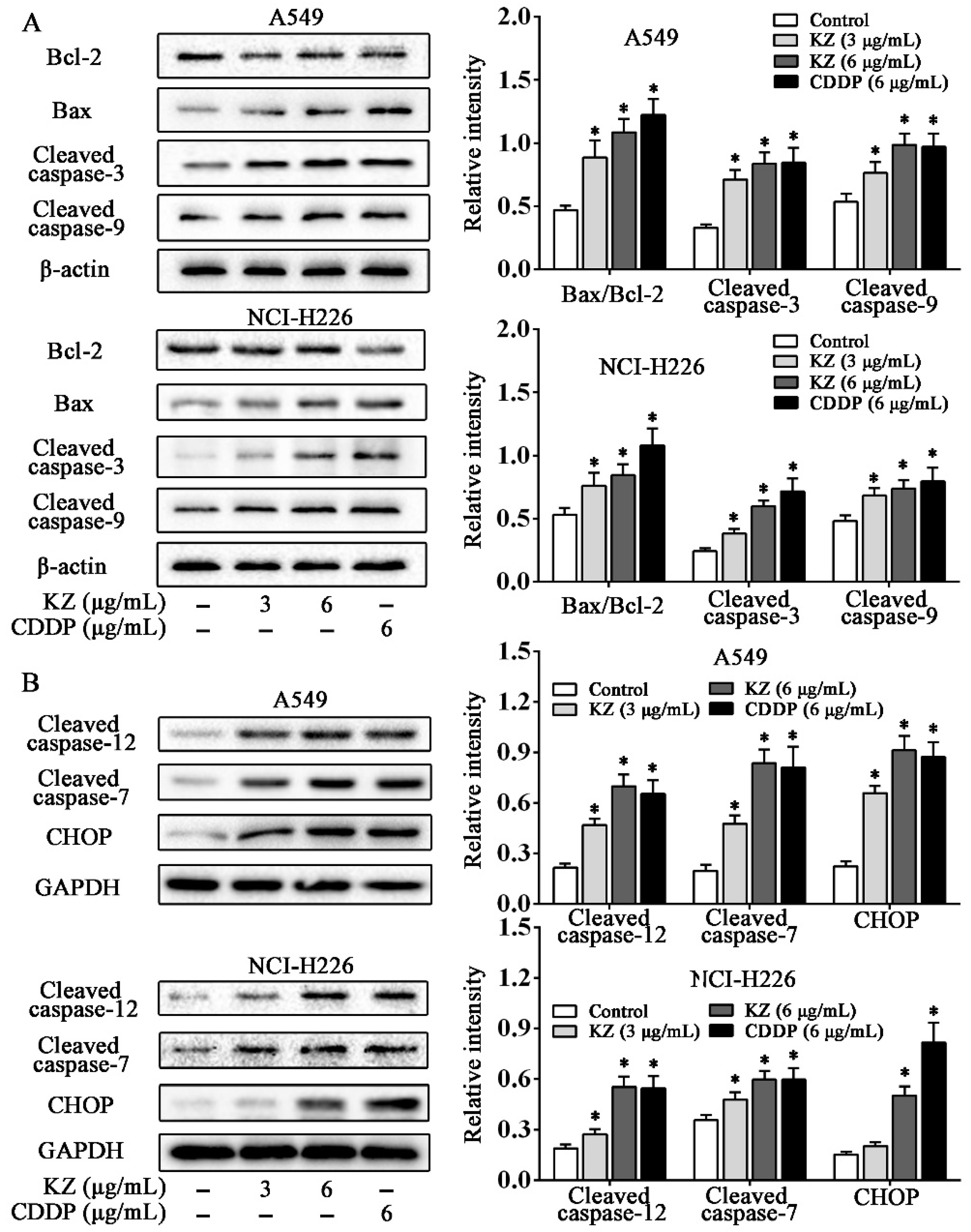
2.4. KZ Promotes NSCLC Cell Apoptosis by Mitochondrial and Endoplasmic Reticulum Stress Pathways
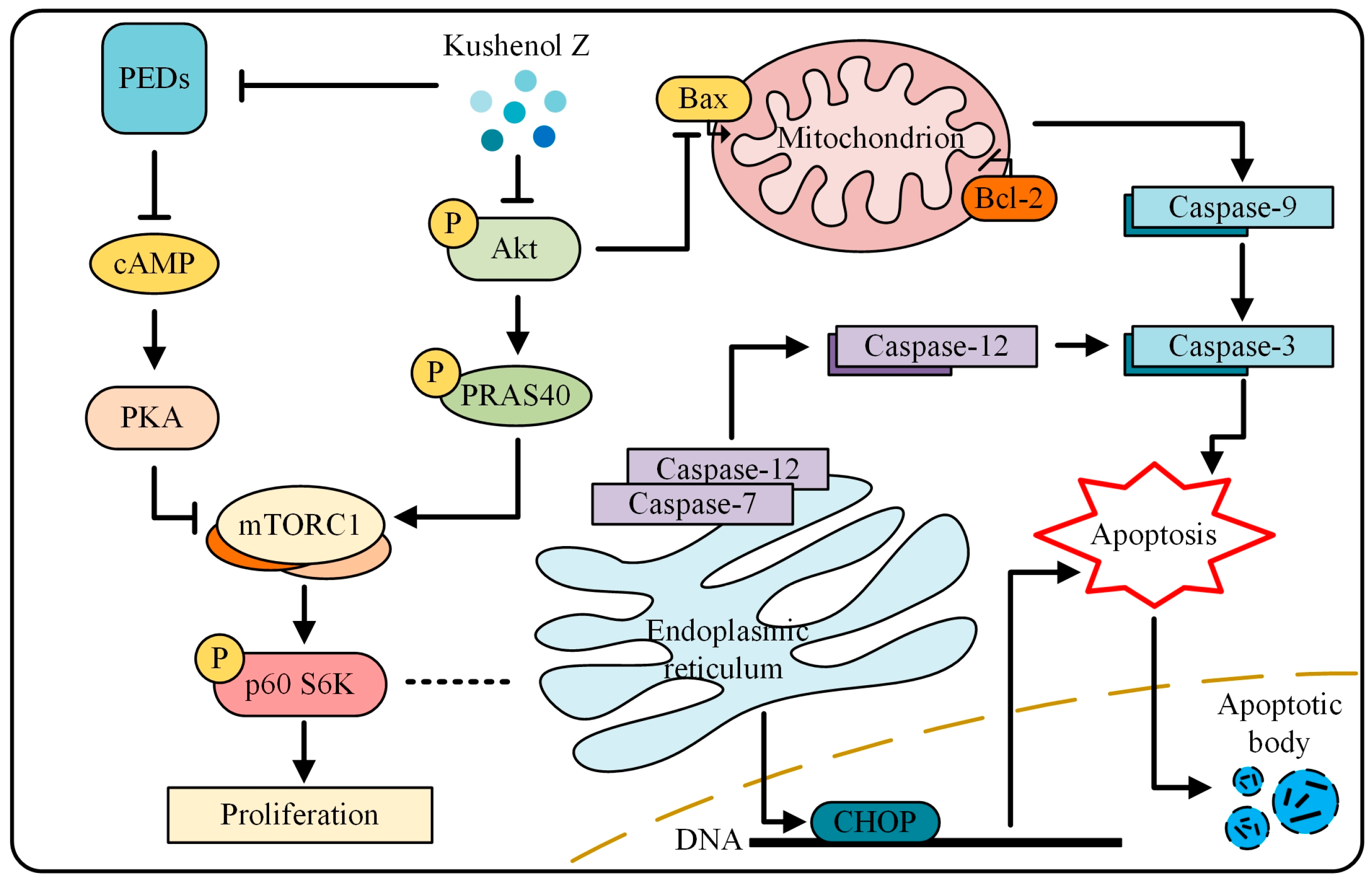
2.5. KZ Mediates Anti-NSCLC Effects by Inhibiting the mTOR Pathway
2.5.1. KZ Upregulates cAMP Levels to Increase PKA Activity That Causes Inhibition of mTORC1
2.5.2. KZ-Induced Antiproliferative Effect Involves the Inhibition of Akt
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Extraction and Isolation
4.3. Kushenol Z (1, KZ)
4.4. Cell Culture and Reagents
4.5. CCK-8 Assay
4.6. Trypan-Blue Exclusion Assay
4.7. Observation of Morphological Changes
4.8. Hoechst 33258 Staining Assay
4.9. Flow Cytometry Analysis (FACS)
4.10. Western Blot Analysis and Immunoassay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kadara, H.; Scheet, P.; Wistuba, I.I.; Spira, A.E. Early events in the molecular pathogenesis of lung cancer. Cancer Prev. Res. 2016, 9, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Niyazi, M.; Maihoefer, C.; Krause, M.; Rödel, C.; Budach, W.; Belka, C. Radiotherapy and “new” drugs-new side effects? Radiat. Oncol. 2011, 6, 177. [Google Scholar] [CrossRef] [PubMed]
- Abu-Surrah, A.S.; Kettunen, M. Platinum group antitumor chemistry: Design and development of new anticancer drugs complementary to cisplatin. Curr. Med. Chem. 2006, 13, 1337–1357. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.M.; Earle, K.A.; Fetter, J.; Xu, S.; Hartman, T.; Chan, D.C.; Zhao, T.L.; Piazza, G.; Klein-Szanto, A.J.; Pamukcu, R. Exisulind-induced apoptosis in a non-small cell lung cancer orthotopic lung tumor model augments docetaxel treatment and contributes to increased survival. Mol. Cancer Ther. 2003, 2, 479–488. [Google Scholar] [PubMed]
- Goldhoff, P.; Warrington, N.M.; Limbrick, D.D.; Hope, A.; Woerner, B.M.; Jackson, E.; Perry, A.; Piwnica-Worms, D.; Rubin, J.B. Targeted inhibition of cyclic AMP phosphodiesterase-4 promotes brain tumor regression. Clin. Cancer Res. 2008, 14, 7717–7725. [Google Scholar] [CrossRef]
- Burvall, K.M.; Palmberg, L.; Larsson, K. The tyrosine kinase inhibitor genistein increases basal cAMP and potentiates forskolin-induced cAMP accumulation in A549 human airway epithelial cells. Mol. Cell. Biochem. 2002, 240, 131–133. [Google Scholar] [CrossRef]
- Ramezani, S.; Vousooghi, N.; Kapourchali, F.R.; Hadjighasem, M.; Hayat, P.; Amini, N.; Joghataei, M.T. Rolipram potentiates bevacizumab-induced cell death in human glioblastoma stem-like cells. Life Sci. 2017, 173, 11–19. [Google Scholar] [CrossRef]
- Smith, P.G.; Wang, F.; Wilkinson, K.N.; Savage, K.J.; Klein, U.; Neuberg, D.S.; Bollag, G.; Shipp, M.A.; Aguiar, R.C. The phosphodiesterase PDE4B limits cAMP-associated PI3K/AKT–dependent apoptosis in diffuse large B-cell lymphoma. Blood 2005, 105, 308–316. [Google Scholar] [CrossRef]
- Xie, J.; Ponuwei, G.A.; Moore, C.E.; Willars, G.B.; Tee, A.R.; Herbert, T.P. cAMP inhibits mammalian target of rapamycin complex-1 and-2 (mTORC1 and 2) by promoting complex dissociation and inhibiting mTOR kinase activity. Cell. Signal. 2011, 23, 1927–1935. [Google Scholar] [CrossRef]
- Sun, M.; Cao, H.; Sun, L.; Dong, S.; Bian, Y.; Han, J.; Zhang, L.; Ren, S.; Hu, Y.; Liu, C. Antitumor activities of kushen: Literature review. Evid. -Based Complementary Altern. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Ryu, S.Y.; Choi, S.U.; Kim, S.K.; No, Z.; Lee, C.O.; Ahn, J.W.; Kim, S.H. In vitro antitumour activity of flavonoids from Sophora flavescens. Phytother. Res. 1997, 11, 51–53. [Google Scholar] [CrossRef]
- Kim, Y.K.; Min, B.S.; Bae, K.H. A cytotoxic constituent fromSophora flavescens. Arch. Pharmacal Res. 1997, 20, 342. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; Yang, J.; Zhao, W.; Xu, C.; Hong, Z.; Mei, Z.; Yang, X. Kurarinol induces hepatocellular carcinoma cell apoptosis through suppressing cellular signal transducer and activator of transcription 3 signaling. Toxicol. Appl. Pharmacol. 2014, 281, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.-C.; Kim, S.-C.; Song, E.-Y.; Um, Y.-C. Isolation and identification of antibacterial and anticancer substances from roots of Sophora Flavescens Aiton. In Proceedings of the XXVII International Horticultural Congress-IHC2006: International Symposium on Plants as Food and Medicine: The Utilization and Development of Horticultural Plants for Human Health, Seoul, Korea, 1 January 2008; pp. 43–48. [Google Scholar] [CrossRef]
- Zhang, X.-L.; Cao, M.-A.; Pu, L.-P.; Huang, S.-S.; Gao, Q.-X.; Yuan, C.-S.; Wang, C.-M. A novel flavonoid isolated from Sophora flavescens exhibited anti-angiogenesis activity, decreased VEGF expression and caused G0/G1 cell cycle arrest in vitro. Die Pharm. -Int. J. Pharm. Sci. 2013, 68, 369–375. [Google Scholar] [CrossRef]
- Yang, X.; Baburin, I.; Plitzko, I.; Hering, S.; Hamburger, M. HPLC-based activity profiling for GABA A receptor modulators from the traditional Chinese herbal drug Kushen (Sophora flavescens root). Mol. Divers. 2011, 15, 361–372. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Curtis-Long, M.J.; Kim, J.H.; Jeong, S.H.; Yang, M.S.; Lee, K.W.; Lee, W.S.; Park, K.H. Pterocarpans and flavanones from Sophora flavescens displaying potent neuraminidase inhibition. Bioorganic Med. Chem. Lett. 2008, 18, 6046–6049. [Google Scholar] [CrossRef]
- Du, X.; Bai, Y.; Liang, H.; Wang, Z.; Zhao, Y.; Zhang, Q.; Huang, L. Solvent effect in 1H NMR spectra of 3′-hydroxy-4′-methoxy isoflavonoids from Astragalus membranaceus var. mongholicus. Magn. Reson. Chem. 2006, 44, 708–712. [Google Scholar] [CrossRef]
- Jung, H.A.; Moon, H.E.; Oh, S.H.; Kim, B.-W.; Sohn, H.S.; Choi, J.S. Kinetics and molecular docking studies of kaempferol and its prenylated derivatives as aldose reductase inhibitors. Chem. -Biol. Interact. 2012, 197, 110–118. [Google Scholar] [CrossRef]
- Quang, T.H.; Ngan, N.T.T.; Minh, C.V.; Kiem, P.V.; Tai, B.H.; Nhiem, N.X.; Thao, N.P.; Luyen, B.T.T.; Yang, S.Y.; Kim, Y.H. Anti-inflammatory and PPAR transactivational properties of flavonoids from the roots of Sophora flavescens. Phytother. Res. 2013, 27, 1300–1307. [Google Scholar] [CrossRef]
- Chen, R.; Liu, X.; Zou, J.; Yin, Y.; Ou, B.; Li, J.; Wang, R.; Xie, D.; Zhang, P.; Dai, J. Regio-and stereospecific prenylation of flavonoids by Sophora flavescens prenyltransferase. Adv. Synth. Catal. 2013, 355, 1817–1828. [Google Scholar] [CrossRef]
- Sato, S.; Takeo, J.; Aoyama, C.; Kawahara, H. Na+-glucose cotransporter (SGLT) inhibitory flavonoids from the roots of Sophora flavescens. Bioorganic Med. Chem. 2007, 15, 3445–3449. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Jeong, D.-M.; Chung, H.Y.; Lim, H.A.; Kim, J.Y.; Yoon, N.Y.; Choi, J.S. Re-evaluation of the antioxidant prenylated flavonoids from the roots of Sophora flavescens. Biol. Pharm. Bull. 2008, 31, 908–915. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kang, T.-H.; Jeong, S.-J.; Ko, W.-G.; Kim, N.-Y.; Lee, B.-H.; Inagaki, M.; Miyamoto, T.; Higuchi, R.; Kim, Y.-C. Cytotoxic lavandulyl flavanones from Sophora f lavescens. J. Nat. Prod. 2000, 63, 680–681. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Taylor, A.W.; Nickerson, G.B.; Ivancic, M.; Henning, J.; Haunold, A.; Deinzer, M.L. Prenylflavonoid variation in Humulus lupulus: Distribution and taxonomic significance of xanthogalenol and 4′-O-methylxanthohumol. Phytochemistry 2000, 53, 759–775. [Google Scholar] [CrossRef]
- Hwang, E.M.; Ryu, Y.B.; Kim, H.Y.; Kim, D.-G.; Hong, S.-G.; Lee, J.H.; Curtis-Long, M.J.; Jeong, S.H.; Park, J.-Y.; Park, K.H. BACE1 inhibitory effects of lavandulyl flavanones from Sophora flavescens. Bioorganic Med. Chem. 2008, 16, 6669–6674. [Google Scholar] [CrossRef]
- Zhao, N.; Tian, K.-T.; Cheng, K.-G.; Han, T.; Hu, X.; Li, D.-H.; Li, Z.-L.; Hua, H.-M. Antiproliferative activity and apoptosis inducing effects of nitric oxide donating derivatives of evodiamine. Bioorg. Med. Chem. 2016, 24, 2971–2978. [Google Scholar] [CrossRef]
- Ohmoto, T.; AikawA, R.; Nikaido, T.; Sankawa, U.; Wu, L.; Ueno, A.; Hukushima, S. Inhibition of adenosine 3’, 5’-cyclic monophosphate phosphodiesterase by components of Sophora flavescens Aiton. Chem. Pharm. Bull. 1986, 34, 2094–2099. [Google Scholar] [CrossRef]
- Peng, T.; Gong, J.; Jin, Y.; Zhou, Y.; Tong, R.; Wei, X.; Bai, L.; Shi, J. Inhibitors of phosphodiesterase as cancer therapeutics. Eur. J. Med. Chem. 2018, 150, 742–756. [Google Scholar] [CrossRef]
- Ardizzoni, A.; Boni, L.; Tiseo, M.; Fossella, F.V.; Schiller, J.H.; Paesmans, M.; Radosavljevic, D.; Paccagnella, A.; Zatloukal, P.; Mazzanti, P. Cisplatin-versus carboplatin-based chemotherapy in first-line treatment of advanced non–small-cell lung cancer: An individual patient data meta-analysis. J. Natl. Cancer Inst. 2007, 99, 847–857. [Google Scholar] [CrossRef]
- Xiao, Z.; Wang, C.-Q.; Feng, J.-H.; Zhou, M.-H.; Wang, Y.-Z.; Li, N.-N.; Sun, Y.-P.; Liu, S.-Y.; Yao, X.-S.; Li, C.-W. Effectiveness and safety of chemotherapy with cytokine-induced killer cells in non–small cell lung cancer: A systematic review and meta-analysis of 32 randomized controlled trials. Cytotherapy 2018. [Google Scholar] [CrossRef]
- Levy, I.; Horvath, A.; Azevedo, M.; de Alexandre, R.B.; Stratakis, C.A. Phosphodiesterase function and endocrine cells: Links to human disease and roles in tumor development and treatment. Curr. Opin. Pharmacol. 2011, 11, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Faucz, F.R.; Horvath, A.; Rothenbuhler, A.; Almeida, M.Q.; Libe, R.; Raffin-Sanson, M.-L.; Bertherat, J.; Carraro, D.M.; Soares, F.A.; de Campos Molina, G. Phosphodiesterase 11A (PDE11A) genetic variants may increase susceptibility to prostatic cancer. J. Clin. Endocrinol. Metab. 2011, 96, E135–E140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-D.; Li, Y.; Xu, Y.; Kim, J.; Huang, S. Phosphodiesterase 7B/microRNA-200c relationship regulates triple-negative breast cancer cell growth. Oncogene 2019, 38, 1106. [Google Scholar] [CrossRef] [PubMed]
- Savai, R.; Pullamsetti, S.S.; Banat, G.-A.; Weissmann, N.; Ghofrani, H.A.; Grimminger, F.; Schermuly, R.T. Targeting cancer with phosphodiesterase inhibitors. Expert Opin. Investig. Drugs 2010, 19, 117–131. [Google Scholar] [CrossRef]
- Davies, S.P.; Reddy, H.; Caivano, M.; Cohen, P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem. J. 2000, 351, 95–105. [Google Scholar] [CrossRef]
- Chen, H.; Yuan, J.; Hao, J.; Wen, Y.; Lv, Y.; Chen, L.; Yang, X. α-Humulene inhibits hepatocellular carcinoma cell proliferation and induces apoptosis through the inhibition of Akt signaling. Food Chem. Toxicol. 2019, 110830. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, B.; Yang, J.; Ma, X.; Deng, S.; Huang, Y.; Wen, Y.; Yuan, J.; Yang, X. Essential oil derived from eupatorium adenophorum spreng. Mediates anticancer effect by inhibiting stat3 and akt activation to induce apoptosis in hepatocellular carcinoma. Front. Pharmacol. 2018, 9, 483. [Google Scholar] [CrossRef]
Sample Availability: Sample of the compound is available from the authors. |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Yang, J.; Hao, J.; Lv, Y.; Chen, L.; Lin, Q.; Yuan, J.; Yang, X. A Novel Flavonoid Kushenol Z from Sophora flavescens Mediates mTOR Pathway by Inhibiting Phosphodiesterase and Akt Activity to Induce Apoptosis in Non-Small-Cell Lung Cancer Cells. Molecules 2019, 24, 4425. https://doi.org/10.3390/molecules24244425
Chen H, Yang J, Hao J, Lv Y, Chen L, Lin Q, Yuan J, Yang X. A Novel Flavonoid Kushenol Z from Sophora flavescens Mediates mTOR Pathway by Inhibiting Phosphodiesterase and Akt Activity to Induce Apoptosis in Non-Small-Cell Lung Cancer Cells. Molecules. 2019; 24(24):4425. https://doi.org/10.3390/molecules24244425
Chicago/Turabian StyleChen, Hao, Jie Yang, Ji Hao, Yibing Lv, Lu Chen, Qinxiong Lin, Jingquan Yuan, and Xinzhou Yang. 2019. "A Novel Flavonoid Kushenol Z from Sophora flavescens Mediates mTOR Pathway by Inhibiting Phosphodiesterase and Akt Activity to Induce Apoptosis in Non-Small-Cell Lung Cancer Cells" Molecules 24, no. 24: 4425. https://doi.org/10.3390/molecules24244425
APA StyleChen, H., Yang, J., Hao, J., Lv, Y., Chen, L., Lin, Q., Yuan, J., & Yang, X. (2019). A Novel Flavonoid Kushenol Z from Sophora flavescens Mediates mTOR Pathway by Inhibiting Phosphodiesterase and Akt Activity to Induce Apoptosis in Non-Small-Cell Lung Cancer Cells. Molecules, 24(24), 4425. https://doi.org/10.3390/molecules24244425