Epinephrine Infiltration of Adipose Tissue Impacts MCF7 Breast Cancer Cells and Total Lipid Content

 , ,
, ,
Abstract
:
1. Introduction
2. Results
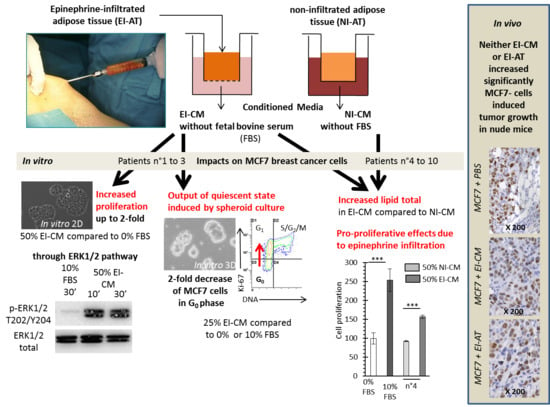
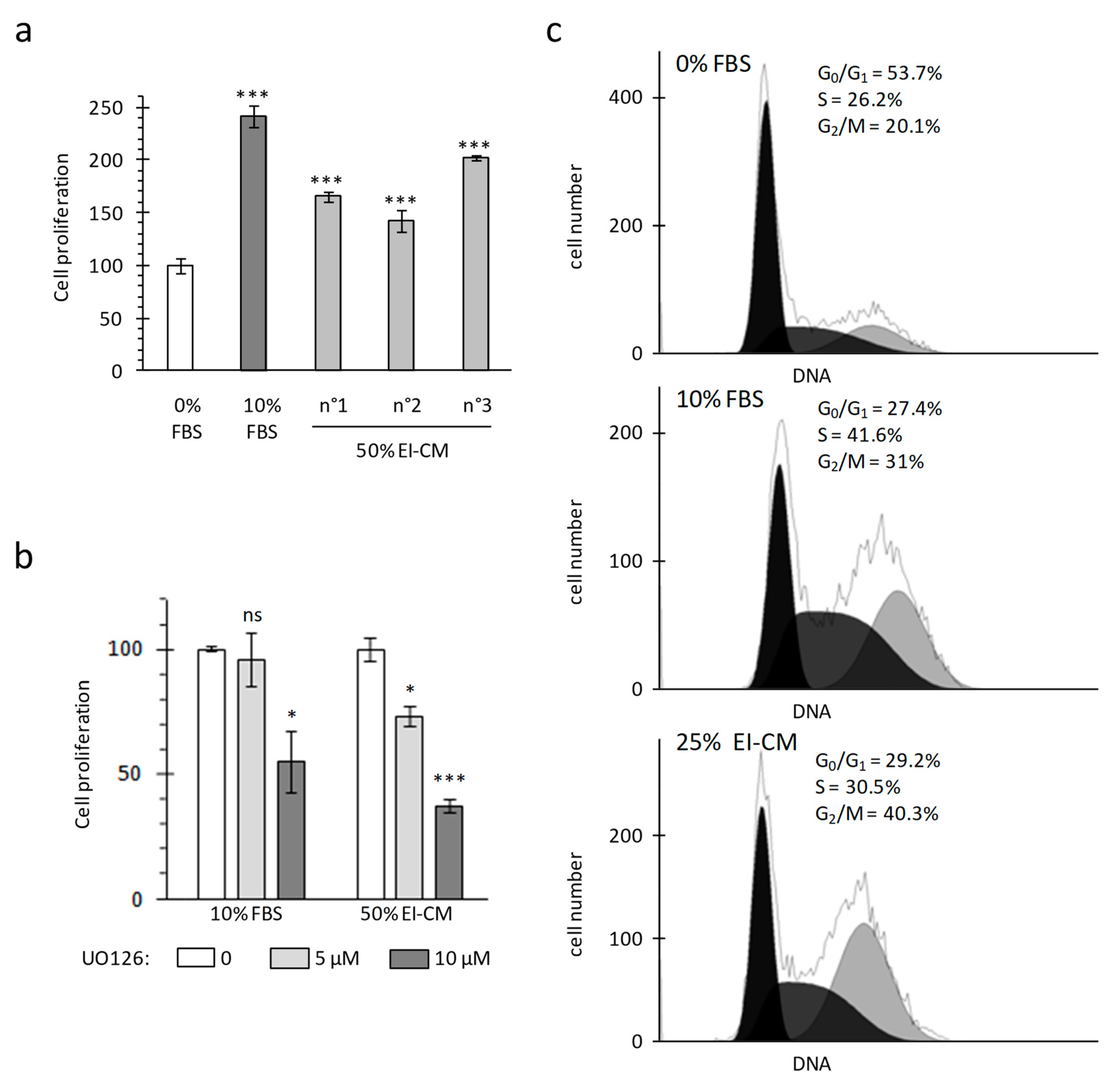
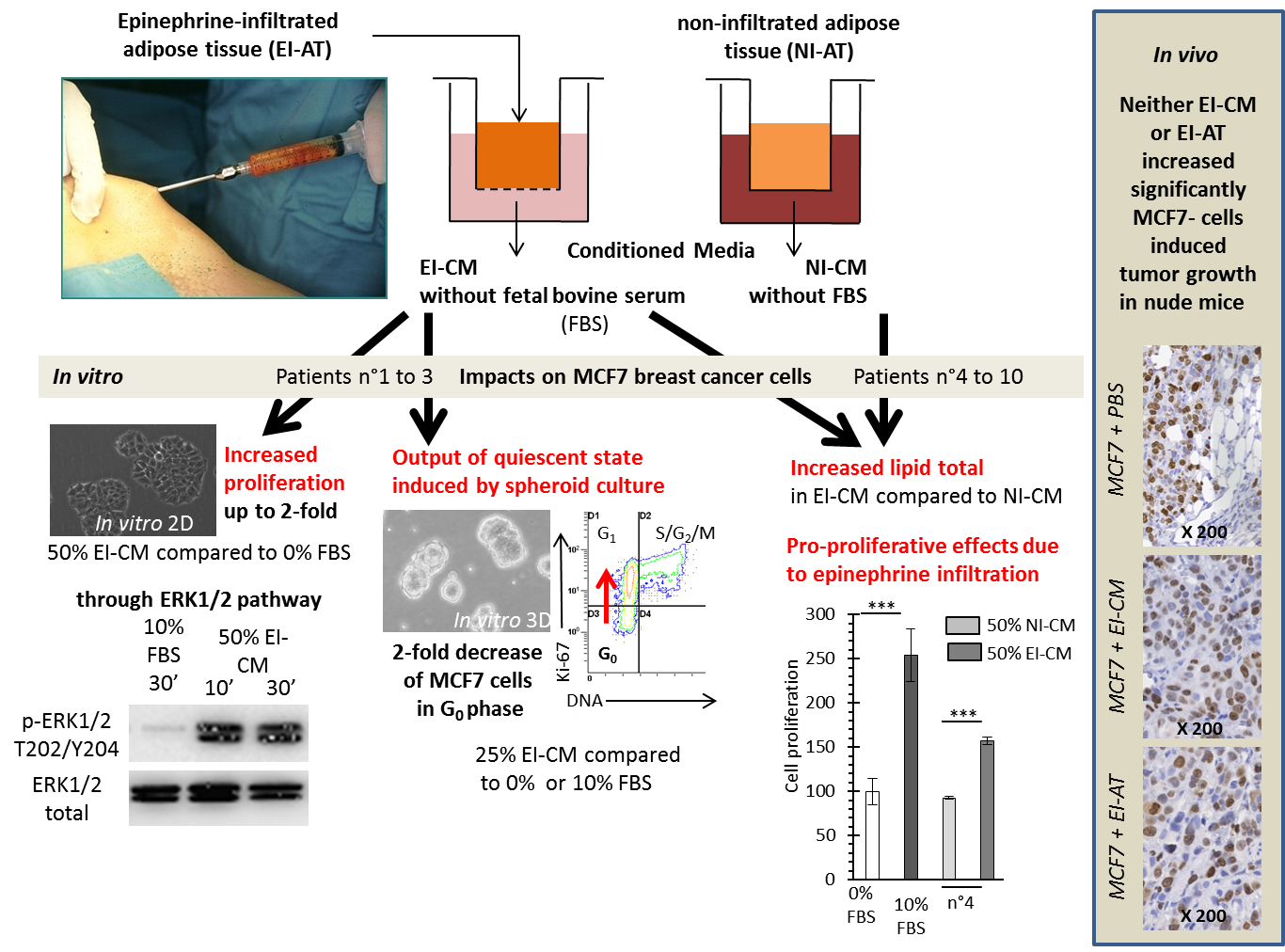
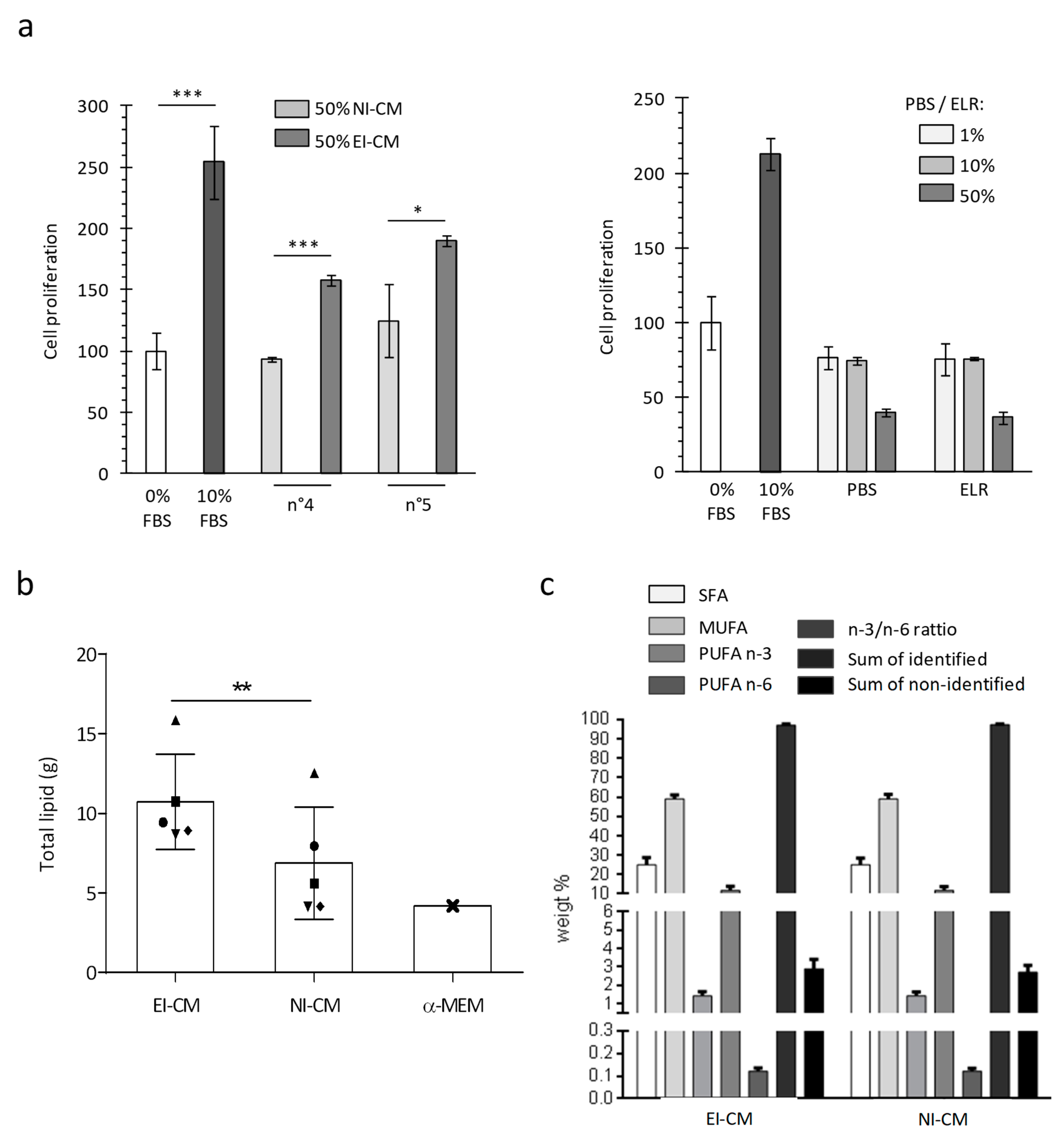
2.1. MCF7 Cell Proliferation was Enhanced by Epinephrine-Infiltrated Adipose Tissue Conditioned Medium
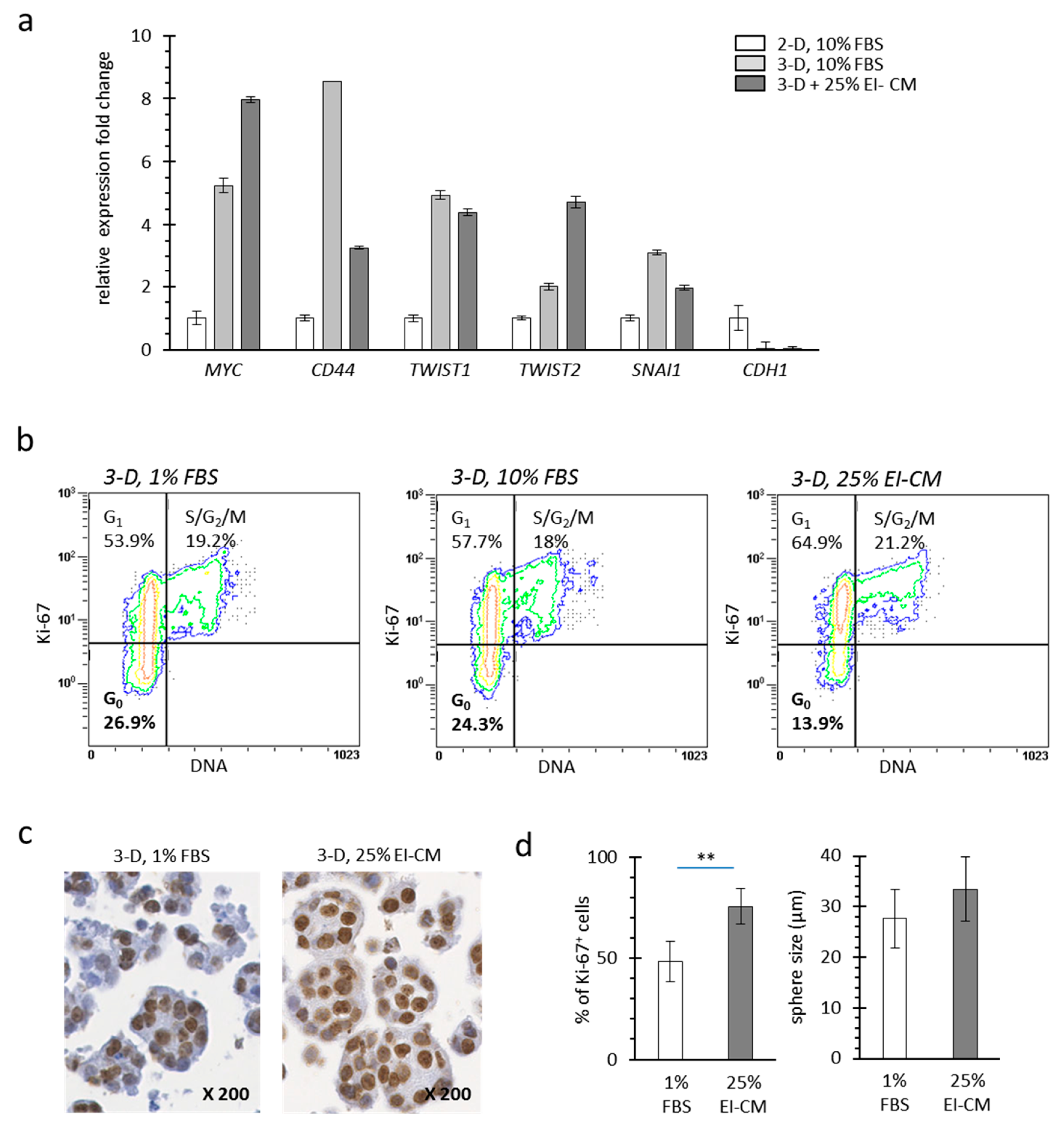
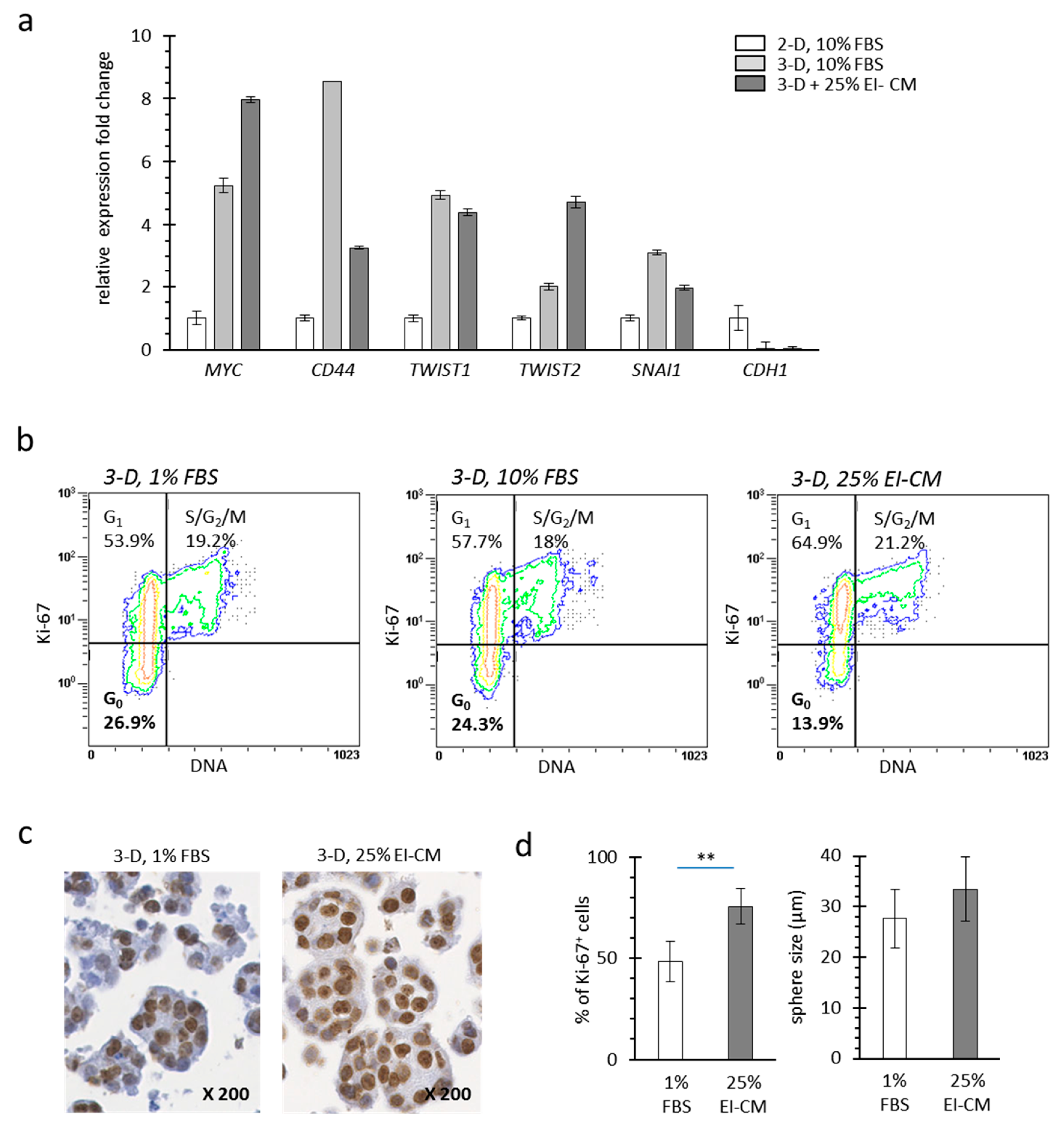
2.2. MCF7 Cell Quiescence was Increased by Sphereoid Culture and Reduced by Epinephrine-Infiltrated Adipose Tissue Conditioned Medium
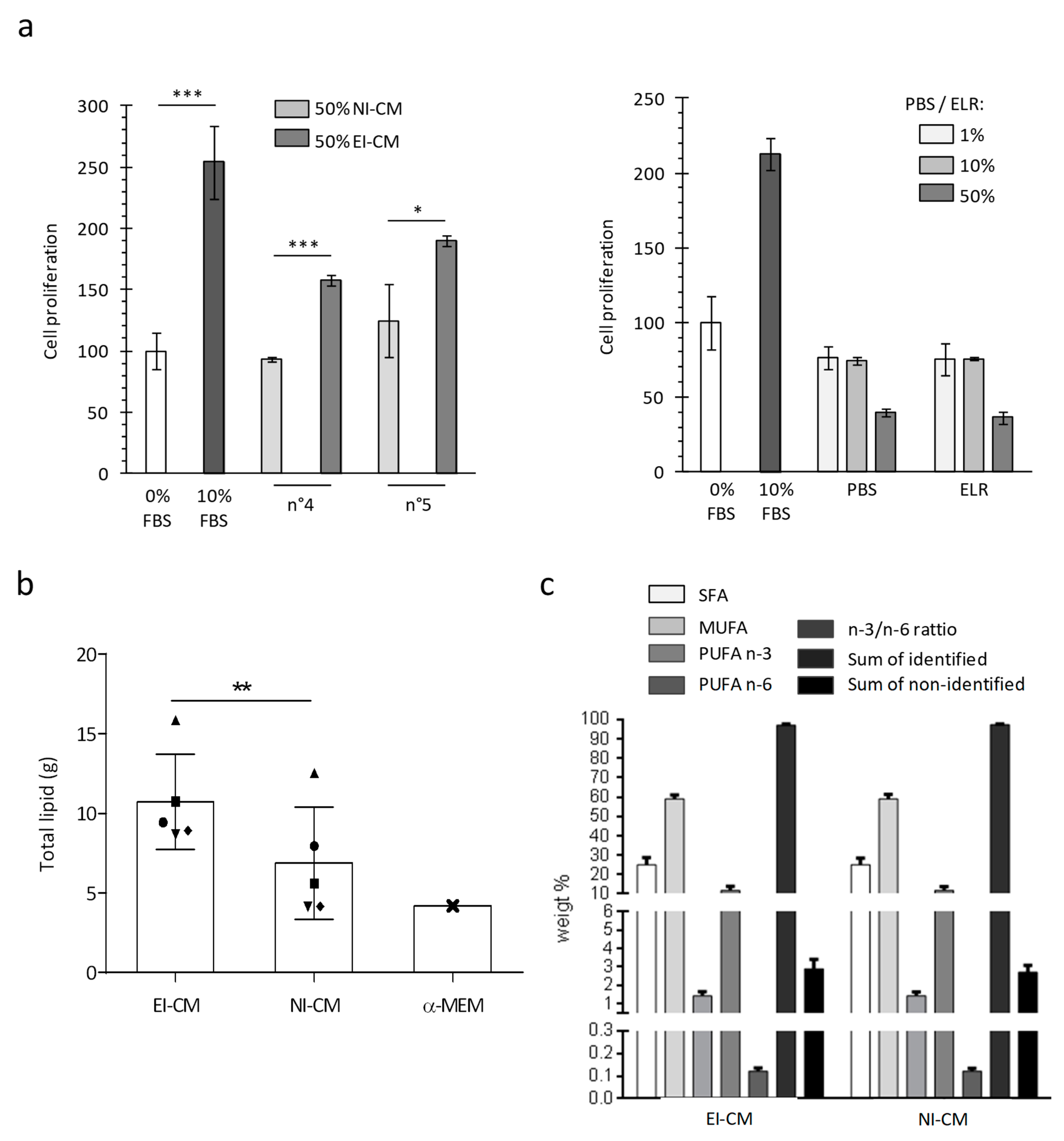
2.3. Epinephrine Infiltration Changed Lipid Content and Proliferative Effect of Adipose Tissue Conditioned Medium
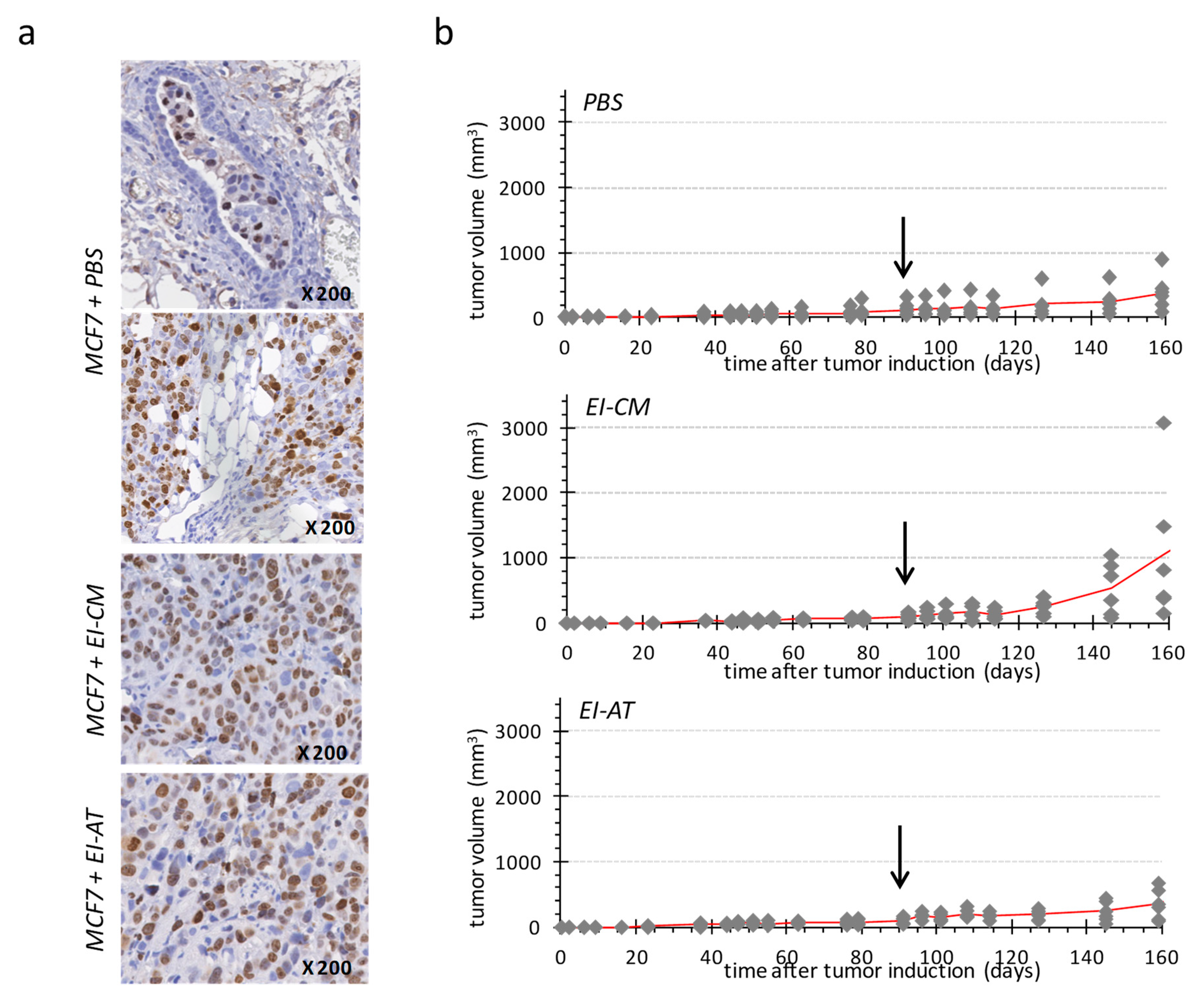
2.4. Injection of Epinephrine-Infiltrated Adipose Tissue or Corresponding Conditioned Medium into MCF7 Tumor in Mice
3. Discussion
4. Materials and Methods
4.1. Adipose Tissue (AT)
4.2. Culture Conditions
4.3. Cell Viability
4.4. Cell Cycle Analysis
4.5. Reverse Transcription and Quantitative PCR
4.6. Breast Carcinoma Model
4.7. Histology Analysis
4.8. Lipid Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wolfson, B.; Eades, G.; Zhou, Q. Adipocyte activation of cancer stem cell signaling in breast cancer. World J. Biol. Chem. 2015, 6, 39–47. [Google Scholar] [CrossRef]
- D’Esposito, V.; Liguoro, D.; Ambrosio, M.R.; Collina, F.; Cantile, M.; Spinelli, R.; Raciti, G.A.; Miele, C.; Valentino, R.; Campiglia, P.; et al. Adipose microenvironment promotes triple negative breast cancer cell invasiveness and dissemination by producing CCL5. Oncotarget 2016, 7, 24495–24509. [Google Scholar]
- Nieman, K.M.; Romero, I.L.; Van Houten, B.; Lengyel, E. Adipose tissue and adipocytes supports tumorigenesis and metastasis. Biochim. Biophys. Acta 2013, 1831, 1533–1541. [Google Scholar] [CrossRef]
- Iyengar, P.; Combs, T.P.; Shah, S.J.; Gouon-Evans, V.; Pollard, J.W.; Albanese, C.; Flanagan, L.; Tenniswood, M.P.; Guha, C.; Lisanti, M.P.; et al. Adipocyte-secreted factors synergistically promote mammary tumorigenesis through induction of anti-apoptotic transcriptional programs and proto-oncogene stabilization. Oncogene 2003, 22, 6408–6423. [Google Scholar] [CrossRef]
- Dirat, B.; Bochet, L.; Dabek, M.; Daviaud, D.; Dauvillier, S.; Majed, B.; Wang, Y.Y.; Meulle, B.; Salles, B.; Gonidec, S.L.; et al. Cancer-associated adipocytes exhibit an activated phenotype and contribute to breast cancer invasion. Cancer Res. 2011, 71, 2455–2465. [Google Scholar] [CrossRef] [PubMed]
- Kornfeld, S.; Goupille, C.; Vibet, S.; Chevalier, S.; Pinet, A.; Lebeau, J.; Tranquart, F.; Bougnoux, P.; Martel, E.; Maurin, A.; et al. Reducing endothelial NOS activation and interstitial fluid pressure with n-3 PUFA offset tumor chemoresistance. Carcinogenesis 2012, 33, 260–267. [Google Scholar] [CrossRef]
- Ouldamer, L.; Goupille, C.; Vildé, A.; Arbion, F.; Body, G.; Chevalier, S.; Cottier, J.P.; Bougnoux, P. N-3 Polyunsaturated Fatty Acids of Marine Origin and Multifocality in Human Breast Cancer. PLoS ONE 2016, 11, e0147148. [Google Scholar] [CrossRef]
- Chauvin, L.; Goupille, C.; Blanc, C.; Pinault, M.; Domingo, I.; Guimaraes, C.; Bougnoux, P.; Chevalier, S.; Maheo, K. Long chain n-3 polyunsaturated fatty acids increase the efficacy of docetaxel in mammary cancer cells by downregulating Akt and PKCε/δ-induced ERK pathways. Biochim. Biophys. Acta 2016, 1861, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Biondo, P.D.; Brindley, D.N.; Sawyer, M.B.; Field, C.J. The potential for treatment with dietary long-chain polyunsaturated n-3 fatty acids during chemotherapy. J. Nutr. Biochem. 2008, 19, 787–796. [Google Scholar] [CrossRef]
- Calviello, G.; Serini, S.; Piccioni, E.; Pessina, G. Antineoplastic effects of n-3 polyunsaturated fatty acids in combination with drugs and radiotherapy: Preventive and therapeutic strategies. Nutr. Cancer 2009, 61, 287–301. [Google Scholar] [CrossRef]
- Bougnoux, P.; Hajjaji, N.; Maheo, K.; Couet, C.; Chevalier, S. Fatty acids and breast cancer: Sensitization to treatments and prevention of metastatic re-growth. Prog. Lipid. Res. 2010, 49, 76–86. [Google Scholar] [CrossRef]
- Kronowitz, S.J.; Mandujano, C.C.; Liu, J.; Kuerer, H.M.; Smith, B.; Garvey, P.; Jagsi, R.; Hsu, L.; Hanson, S.; Valero, V. Lipofilling of the breast does not increase the risk of recurrence of breast cancer: A matched controlled study. Plast. Reconstr. Surg. 2016, 137, 385–393. [Google Scholar] [CrossRef]
- Charvet, H.J.; Orbay, H.; Wong, M.S.; Sahar, D.E. The oncologic safety of breast fat grafting and contradictions between basic science and clinical studies: A systematic review of the recent literature. Ann. Plast. Surg. 2015, 75, 471–479. [Google Scholar] [CrossRef]
- Gentile, P.; Casella, D.; Palma, E.; Calabrese, C. Engineered fat graft enhanced with adipose-derived stromal vascular fraction cells for regenerative medicine: Clinical, histological and instrumental evaluation in breast reconstruction. J. Clin. Med. 2019, 8, 504. [Google Scholar] [CrossRef]
- Petit, J.Y.; Rietjens, M.; Botteri, E.; Rotmensz, N.; Bertolini, F.; Curigliano, G.; Rey, P.; Garusi, F.; De Lorenzi, S.; Martella, S.; et al. Evaluation of fat grafting safety in patients with intraepithelial neoplasia: A matched-cohort study. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2013, 24, 1479–1484. [Google Scholar] [CrossRef]
- Geissler, P.J.; Davis, K.; Roostaeian, J.; Unger, J.; Huang, J.; Rohrich, R.J. Improving fat transfer viability: The role of aging, body mass index, and harvest site. Plast. Reconstr. Surg. 2014, 134, 227–232. [Google Scholar] [CrossRef]
- Gentile, P.; De Angelis, B.; Di Pietro, V.; Amorosi, V.; Scioli, M.G.; Orlandi, A.; Cervelli, V. Gentle Is Better: The original “Gentle Technique” for fat placement in breast lipofilling. J. Cutan. Aesthet. Surg. 2018, 11, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Orlandi, A.; Scioli, M.G.; Di Pasquali, C.; Bocchini, I.; Curcio, C.B.; Floris, M.; Fiaschetti, V.; Floris, R.; Cervell, V. A comparative translational study: The combined use of enhanced stromal vascular fraction and platelet-rich plasma improves fat grafting maintenance in breast reconstruction. Stem Cells Transl. Med. 2012, 1, 341–351. [Google Scholar] [CrossRef]
- Gentile, P.; Scioli, M.G.; Orlandi, A.; Cervelli, V. Breast reconstruction with enhanced stromal vascular fraction fat grafting: What is the best method? Plast. Reconstr. Surg. Glob. Open 2015, 3, e406. [Google Scholar] [CrossRef]
- Hamza, A.; Lohsiriwat, V.; Rietjens, M. Lipofilling in breast cancer surgery. Gland Surg. 2013, 2, 7–14. [Google Scholar] [PubMed]
- Large, V.; Hellström, L.; Reynisdottir, S.; Lönnqvist, F.; Eriksson, P.; Lannfelt, L.; Arner, P. Human beta-2 adrenoceptor gene polymorphisms are highly frequent in obesity and associate with altered adipocyte beta-2 adrenoceptor function. J. Clin. Investig. 1997, 100, 3005–3013. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.K.; Wang, C.; Ravi, S.; Howell, M.C.; Mallela, J.; Alibrahim, M.; Green, R.; Hellemann, G.; Mohapatra, S.S.; Mohapatra, S. A 3D fibrous scaffold inducing tumoroids: A platform for anticancer drug development. PLoS ONE 2013, 8, e75345. [Google Scholar] [CrossRef]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef]
- Cuylen, S.; Blaukopf, C.; Politi, A.Z.; Müller-Reichert, T.; Neumann, B.; Poser, I.; Ellenberg, J.; Hyman, A.A.; Gerlich, D.W. Ki-67 acts as a biological surfactant to disperse mitotic chromosomes. Nature 2016, 535, 308–312. [Google Scholar] [CrossRef]
- Niddam, J.; Vidal, L.; Hersant, B.; Meningaud, J.P. Primary Fat Grafting to the Pectoralis Muscle during Latissimus Dorsi Breast Reconstruction. Plast. Reconstr. Surg. Glob. Open 2016, 4, 1059. [Google Scholar] [CrossRef]
- Lohsiriwat, V.; Curigliano, G.; Rietjens, M.; Goldhirsch, A.; Petit, J.Y. Autologous fat transplantation in patients with breast cancer: «silencing» or “fueling” cancer recurrence? Breast Edinb. Scotl. 2011, 20, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Charvet, H.J.; Orbay, H.; Harrison, L.; Devi, K.; Sahar, D.E. In vitro effects of adipose-derived stem cells on breast cancer cells harvested from the same patient. Ann. Plast. Surg. 2016, 76, S241–S245. [Google Scholar] [CrossRef]
- Fraser, J.K.; Hedrick, M.H.; Cohen, S.R. Oncologic risks of autologous fat grafting to the breast. Aesthet. Surg. J. 2011, 31, 68–75. [Google Scholar] [CrossRef]
- Silva-Vergara, C.; Fontdevila, J.; Weshahy, O.; Yuste, M.; Descarrega, J.; Grande, L. Breast cancer recurrence is not increased with lipofilling reconstruction: A case-controlled study. Ann. Plast. Surg. 2017, 79, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Largo, R.D.; Tchang, L.A.H.; Mele, V.; Scherberich, A.; Harder, Y.; Wettstein, R.; Schaefer, D. Efficacy, safety and complications of autologous fat grafting to healthy breast tissue: A systematic review. J. Plast. Reconstr. Aesthetic Surg. 2014, 67, 437–448. [Google Scholar] [CrossRef]
- Cohen, O.; Lam, G.; Karp, N.; Choi, M. Determining the oncologic safety of autologous fat grafting as a reconstructive modality: An institutional review of breast cancer recurrence rates and surgical outcomes. Plast. Reconstr. Surg. 2017, 140, 382e–392e. [Google Scholar] [CrossRef]
- Wazir, U.; El Hage Chehade, H.; Headon, H.; Oteifa, M.; Kasem, A.; Mokbel, K. Oncological safety of lipofilling in patients with breast cancer: A meta-analysis and update on clinical practice. Anticancer Res. 2016, 36, 4521–4528. [Google Scholar] [CrossRef] [Green Version]
- Gigli, S.; Amabile, M.I.; Pastena, F.D.; De Luca, A.; Gulia, C.; Manganaro, L.; Monti, M.; Ballesio, L. Lipofilling outcomes mimicking breast cancer recurrence: Case report and update of the literature. Anticancer. Res. 2017, 37, 5395–5398. [Google Scholar]
- Suszynski, T.M.; Sieber, D.A.; Van Beek, A.L.; Cunningham, B.L. Characterization of adipose tissue for autologous fat grafting. Aesthet. Surg. J. 2015, 35, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Spear, S.L.; Coles, C.N.; Leung, B.K.; Gitlin, M.; Parekh, M.; Macarios, D. The safety, effectiveness, and efficiency of autologous fat grafting in breast surgery. Plast. Reconstr. Surg. Glob. Open 2016, 4, e827. [Google Scholar] [CrossRef]
- Waked, K.; Colle, J.; Doornaert, M.; Cocquyt, V.; Blondeel, P. Systematic review: The oncological safety of adipose fat transfer after breast cancer surgery. Breast Edinb. Scotl. 2017, 31, 128–136. [Google Scholar] [CrossRef]
- Lafontan, M.; Berlan, M. Fat cell adrenergic receptors and the control of white and brown fat cell function. J. Lipid Res. 1993, 34, 1057–1091. [Google Scholar]
- Bougnères, P.; Stunff, C.L.; Pecqueur, C.; Pinglier, E.; Adnot, P.; Ricquier, D. In vivo resistance of lipolysis to epinephrine. A new feature of childhood onset obesity. J. Clin. Investig. 1997, 99, 2568–2573. [Google Scholar] [CrossRef]
- Jocken, J.W.E.; Blaak, E.E. Catecholamine-induced lipolysis in adipose tissue and skeletal muscle in obesity. Physiol. Behav. 2008, 94, 219–230. [Google Scholar] [CrossRef]
- Danilo, C.; Gutierrez-Pajares, J.L.; Mainieri, M.A.; Mercier, I.; Lisanti, M.P.; Frank, P.G. Scavenger receptor class B type I regulates cellular cholesterol metabolism and cell signaling associated with breast cancer development. Breast Cancer Res. 2013, 15, R87. [Google Scholar] [CrossRef]
- Milde-Langosch, K.; Bamberger, A.M.; Rieck, G.; Grund, D.; Hemminger, G.; Müller, V.; Longing, T. Expression and prognostic relevance of activated extracellular-regulated kinases (ERK1/2) in breast cancer. Br. J. Cancer 2005, 92, 2206–2215. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Attané, C.; Milhas, D.; Dirat, B.; Dauvillier, S.; Guerard, A.; Gilhodes, G.; Lazar, I.; Alet, N.; Laurent, V.; et al. Mammary adipocytes stimulate breast cancer invasion through metabolic remodeling of tumor cells. JCI. Insight. 2017, 2, e87489. [Google Scholar] [CrossRef] [Green Version]
- Basen-Engquist, K.; Chang, M. Obesity and cancer risk: Recent review and evidence. Curr. Oncol. Rep. 2011, 13, 71–76. [Google Scholar] [CrossRef]
- Chipoy, C.; Brounais, B.; Trichet, V.; Battaglia, S.; Berreur, M.; Oliver, L.; Juin, P.; Redini, F.; Heymann, D.; Blanchard, F. Sensitization of osteosarcoma cells to apoptosis by oncostatin M depends on STAT5 and p53. Oncogene 2007, 26, 6653–6664. [Google Scholar] [CrossRef] [Green Version]
- Avril, P.; Le Nail, L.R.; Brennan, M.Á.; Rosset, P.; De Pinieux, G.; Layrolle, P.; Layrolle, P.; Heymann, D.; Trichet, V.; Perrot, P. Mesenchymal stem cells increase proliferation but do not change quiescent state of osteosarcoma cells: Potential implications according to the tumor resection status. J. Bone Oncol. 2015, 5, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gernapudi, R.; Yao, Y.; Zhang, Y.; Wolfson, B.; Roy, S.; Duru, N.; Eades, G.; Yang, P.; Zhou, Q. Targeting exosomes from preadipocytes inhibits preadipocyte to cancer stem cell signaling in early-stage breast cancer. Breast Cancer Res. Treat. 2015, 150, 685–695. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Symbol | Official Full Name | Reverse Primer |
|---|---|---|
| HPTR1 | Hypoxanthine PhosphoRibosyl Transferase 1 | CGAGCAAGACGTTCAGTCCT |
| CD44 | Cluster of Differentiation 44 | CGGCAGGTTATATTCAAATCG |
| TWIST1 | Twist-related protein 1 | TGCAGAGGTGTGAGGATGGTGC |
| TWIST2 | Twist-related protein 2 | AGAAGGTCTGGCAATGGCAGCA |
| SNAI1 | Snail family transcriptional repressor 1 | CAGCAGGTGGGCCTGGTCGTA |
| CDH1 | Cadherin 1 | CCAGCGGCCCCTTCACAGTC |
| MYC | Myelocytomatosis viral oncogene homolog | GATCCAGACTCTGACCTTTTGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avril, P.; Vidal, L.; Barille-Nion, S.; Le Nail, L.-R.; Redini, F.; Layrolle, P.; Pinault, M.; Chevalier, S.; Perrot, P.; Trichet, V. Epinephrine Infiltration of Adipose Tissue Impacts MCF7 Breast Cancer Cells and Total Lipid Content. Int. J. Mol. Sci. 2019, 20, 5626. https://doi.org/10.3390/ijms20225626
Avril P, Vidal L, Barille-Nion S, Le Nail L-R, Redini F, Layrolle P, Pinault M, Chevalier S, Perrot P, Trichet V. Epinephrine Infiltration of Adipose Tissue Impacts MCF7 Breast Cancer Cells and Total Lipid Content. International Journal of Molecular Sciences. 2019; 20(22):5626. https://doi.org/10.3390/ijms20225626
Chicago/Turabian StyleAvril, Pierre, Luciano Vidal, Sophie Barille-Nion, Louis-Romée Le Nail, Françoise Redini, Pierre Layrolle, Michelle Pinault, Stéphane Chevalier, Pierre Perrot, and Valérie Trichet. 2019. "Epinephrine Infiltration of Adipose Tissue Impacts MCF7 Breast Cancer Cells and Total Lipid Content" International Journal of Molecular Sciences 20, no. 22: 5626. https://doi.org/10.3390/ijms20225626