Ischemia Reperfusion Injury: Mechanisms of Damage/Protection and Novel Strategies for Cardiac Recovery/Regeneration


Abstract
:1. Introduction
2. Reperfusion Injury
2.1. Lethal Reperfusion Injury
2.2. Microvasculature Damage
2.3. Mitochondrial Permeability Transition Pore (mPTP)
3. Cardioprotective Pathways
3.1. The Anti-Apoptotic Pro-Surviving Pathway: The “Reperfusion Injury Salvage Kinase” (RISK) Pathway
3.2. The Survivor Activating Factor Enhancement (SAFE) Pathway
3.3. Nitric Oxide and cGKI Pathway
3.4. Autophagy
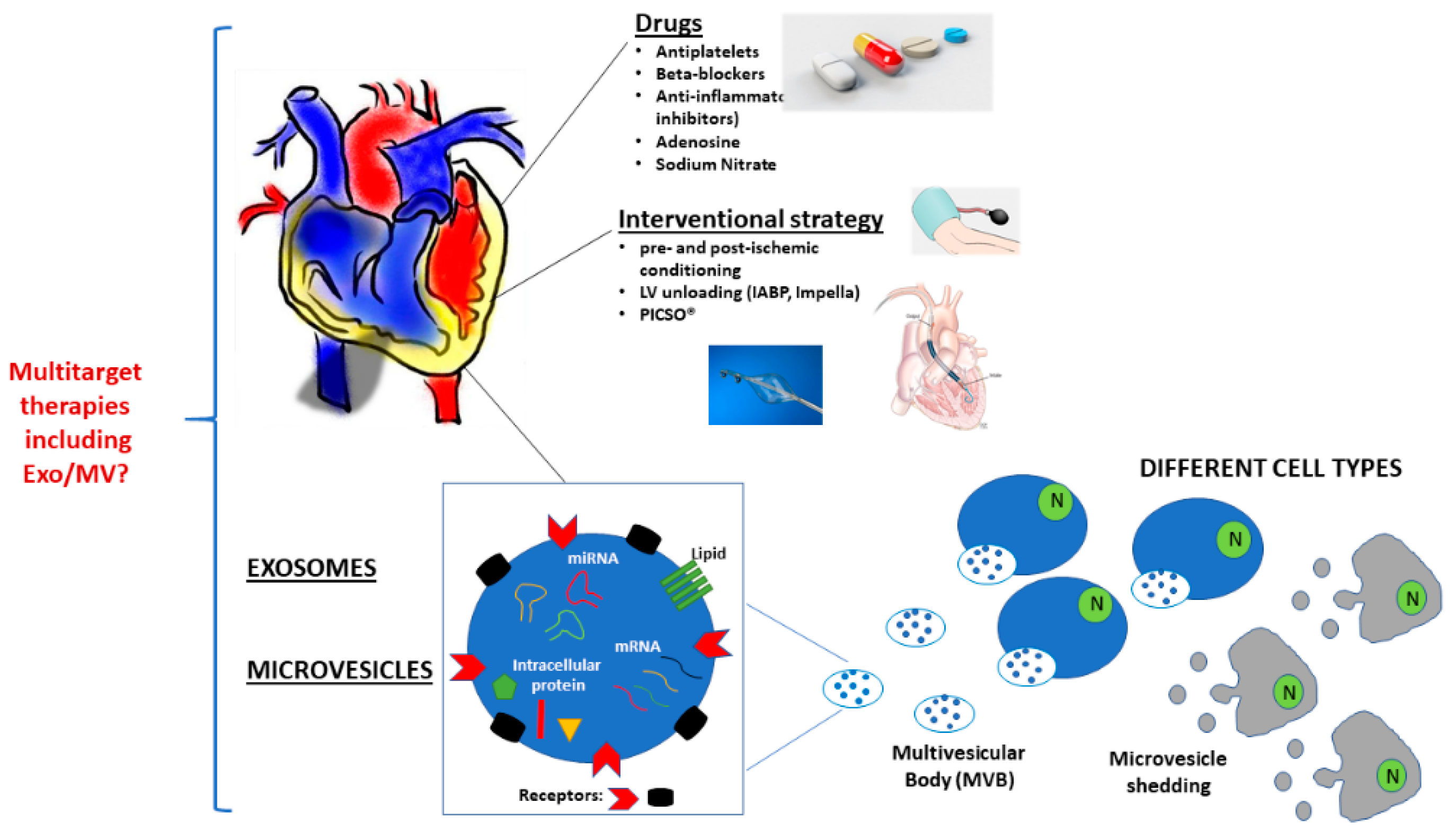
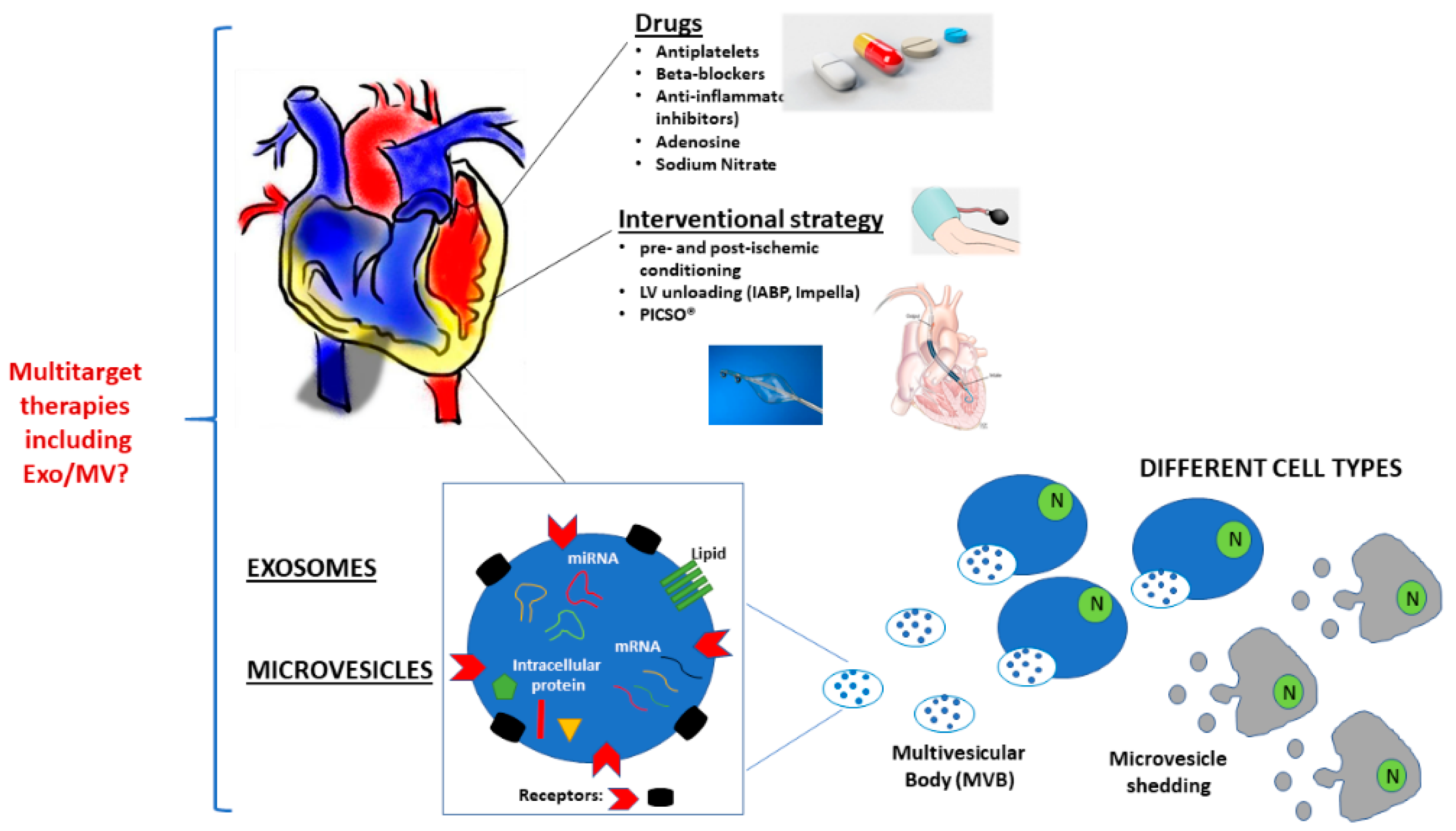
4. Current Strategies to Reduce Ischemic Damage and Reperfusion Injury
4.1. Pharmacological Approaches
4.1.1. Current Multitarget Therapy: Antiplatelet Drugs and Beta-Blockers
4.1.2. Anti-Inflammatory Drugs
4.1.3. Adenosine and Sodium Nitrite
4.2. Non-Pharmacological Approaches
4.2.1. Ischemic Pre-Conditioning
4.2.2. Ischemic Post-Conditioning
4.3. Interventional Strategies
4.3.1. Left Ventricle Unloading
4.3.2. Pressure-Controlled Intermittent Coronary Sinus Occlusion (PICSO®)
5. EV and Cardioprotection
6. MV and Cardioprotection
6.1. Platelet-Derived MV (PMV)
6.2. Endothelial-Derived MV (EMV)
6.3. Other MV
7. Exo and Cardioprotection
7.1. Cardioprotection
7.1.1. MSC-Exo
7.1.2. CPC-Exo
7.2. Autophagy
7.3. Modulation of Inflammation
7.4. Cardiac Telocytes (CTs)-Exo
8. Exo and Cardiac Regeneration
8.1. Angiogenesis
8.1.1. CPC-Exo
8.1.2. MSC-Exo
8.1.3. ADSC-Exo
8.1.4. Plasma-Exo
8.2. Cardioregeneration
9. Therapeutic Device by Manipulating Exo
9.1. Exo Isolation and Production
9.2. Exo Targeting to Increase Cardiac Homing
9.3. “Drug” Loaded Exo
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nesto, R.W.; Kowalchuk, G.J. The ischemic cascade: Temporal sequence of hemodynamic, electrocardiographic and symptomatic expressions of ischemia. Am. J. Cardiol. 1987, 59, 23C–30C. [Google Scholar] [CrossRef]
- Ponikowski, A.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.F.; Coats, A.J.S.; Falk, V.; González-Juanatey, J.R.; Harjola, V.P.; Jankowska, E.A.; et al. 2016 ESC guidelines for the diagnosis and treatment of acute and chronic heart failure. Rev. Esp. Cardiol. 2016, 69, 1167. [Google Scholar] [PubMed]
- Michler, R.E. The current status of stem cell therapy in ischemic heart disease. J. Card. Surg. 2018, 33, 520–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, S.M.; Ferdinandy, P.; Andreadou, I.; Bøtker, H.E.; Heusch, G.; Ibáñez, B.; Ovize, M.; Schulz, R.; Yellon, D.M.; Hausenloy, D.J.; et al. Multitarget Strategies to Reduce Myocardial Ischemia/Reperfusion Injury: JACC Review Topic of the Week. J. Am. Coll. Cardiol. 2019, 73, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating extracellular vesicles in human disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Vicencio, J.M.; Yellon, D.M.; Davidson, S.M. Microvesicles and exosomes: New players in metabolic and cardiovascular disease. J. Endocrinol. 2016, 228, R57–R71. [Google Scholar] [CrossRef]
- Deregibus, M.C.; Cantaluppi, V.; Calogero, R.; Lo Iacono, M.; Tetta, C.; Biancone, L.; Bruno, S.; Bussolati, B.; Camussi, G. Endothelial progenitor cell - Derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood 2007, 110, 2440–2448. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, W.; Liu, G.; Cai, W.; Millard, R.W.; Wang, Y.; Chang, J.; Peng, T.; Fan, G.C. Cardiomyocytes mediate anti-angiogenesis in type 2 diabetic rats through the exosomal transfer of miR-320 into endothelial cells. J. Mol. Cell. Cardiol. 2014, 74, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Yan, I.K.; Haga, H.; Patel, T. Modulation of hypoxia-signaling pathways by extracellular linc-RoR. J. Cell Sci. 2014, 127, 1585–1594. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yan, I.K.; Kogure, T.; Haga, H.; Patel, T. Extracellular vesicle-mediated transfer of long non-coding RNA ROR modulates chemosensitivity in human hepatocellular cancer. FEBS Open Bio 2014, 4, 458–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An integrated database of high-throughput data for systemic analyses of extracellular vesicles. J. Extracell. Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef] [PubMed]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Sluijter, J.P.G.; Davidson, S.M.; Boulanger, C.M.; Buzás, E.I.; De Kleijn, D.P.V.; Engel, F.B.; Giricz, Z.; Hausenloy, D.J.; Kishore, R.; Lecour, S.; et al. Extracellular vesicles in diagnostics and therapy of the ischaemic heart: Position Paper from the Working Group on Cellular Biology of the Heart of the European Society of Cardiology. Cardiovasc. Res. 2018, 114, 19–34. [Google Scholar] [CrossRef]
- Zhang, M.; Schekman, R. Unconventional Secretion, Unconventional Solutions. Science 2013, 340, 559–561. [Google Scholar] [CrossRef]
- Glembotski, C.C. Expanding the Paracrine Hypothesis of Stem Cell–Mediated Repair in the Heart. Circ. Res. 2017, 120, 772–774. [Google Scholar] [CrossRef]
- Gartz, M.; Strande, J.L. Examining the paracrine effects of exosomes in cardiovascular disease and repair. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef]
- Grange, C.; Tritta, S.; Tapparo, M.; Cedrino, M.; Tetta, C.; Camussi, G.; Brizzi, M.F. Stem cell-derived extracellular vesicles inhibit and revert fibrosis progression in a mouse model of diabetic nephropathy. Sci. Rep. 2019, 9, 4468. [Google Scholar] [CrossRef]
- Cavallari, C.; Ranghino, A.; Tapparo, M.; Cedrino, M.; Figliolini, F.; Grange, C.; Giannachi, V.; Garneri, P.; Deregibus, M.C.; Collino, F.; et al. Serum-derived extracellular vesicles (EVs) impact on vascular remodeling and prevent muscle damage in acute hind limb ischemia. Sci. Rep. 2017, 7, 8180. [Google Scholar] [CrossRef]
- Barani, B.; Rajasingh, S.; Rajasingh, J. Exosomes: Outlook for Future Cell-Free Cardiovascular Disease Therapy. In Exosomes in Cardiovascular Diseases: Biomarkers, Pathological and Therapeutic Effects; Xiao, J., Cretoiu, S., Eds.; Springer: Singapore, 2017; pp. 285–307. ISBN 978-981-10-4397-0. [Google Scholar]
- Chen, G.-H.; Xu, J.; Yang, Y.-J. Exosomes: Promising sacks for treating ischemic heart disease? Am. J. Physiol. Circ. Physiol. 2017, 313, H508–H523. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, X.; Cao, W.; Ma, J.; Sun, L.; Qian, H.; Zhu, W.; Xu, W. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Relieve Acute Myocardial Ischemic Injury. Stem Cells Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, C.M.; Loyer, X.; Rautou, P.E.; Amabile, N. Extracellular vesicles in coronary artery disease. Nat. Rev. Cardiol. 2017, 14, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Chen, G.H.; Yang, Y.J. Exosomes: A rising star in falling hearts. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.M.; Yellon, D.M. Exosomes and cardioprotection—A critical analysis. Mol. Aspects Med. 2018, 60, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Rochette, L.; Wu, Y.; Rosenblatt-Velin, N. New Insights into the Role of Exosomes in the Heart After Myocardial Infarction. J. Cardiovasc. Transl. Res. 2019, 12, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Maroko, P.R.; Libby, P.; Ginks, W.R.; Bloor, C.M.; Shell, W.E.; Sobel, B.E.; Ross, J. Coronary artery reperfusion. I. Early effects on local myocardial function and the extent of myocardial necrosis. J. Clin. Invest. 1972, 51, 2710–2716. [Google Scholar] [CrossRef] [PubMed]
- Boersma, E.; Maas, A.C.P.; Deckers, J.W.; Simoons, M.L. Early thrombolytic treatment in acute myocardial infarction: Reappraisal of the golden hour. Lancet 1996, 348, 771–775. [Google Scholar] [CrossRef]
- Boersma, E. Does time matter? A pooled analysis of randomized clinical trials comparing primary percutaneous coronary intervention and in-hospital fibrinolysis in acute myocardial infarction patients. Eur. Heart J. 2006, 27, 779–788. [Google Scholar] [CrossRef] [Green Version]
- Ibanez, B.; James, S.; Agewall, S.; Antunes, M.J.; Bucciarelli-Ducci, C.; Bueno, H.; Caforio, A.L.P.; Crea, F.; Goudevenos, J.A.; Halvorsen, S.; et al. 2017 ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST-segment elevation. Eur. Heart J. 2018, 39, 119–177. [Google Scholar] [CrossRef]
- Roffi, M.; Patrono, C.; Collet, J.P.; Mueller, C.; Valgimigli, M.; Andreotti, F.; Bax, J.J.; Borger, M.A.; Brotons, C.; Chew, D.P.; et al. 2015 ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent st-segment elevation: Task force for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation of. Eur. Heart J. 2016, 37, 267–315. [Google Scholar] [CrossRef]
- Rathore, S.S.; Curtis, J.P.; Chen, J.; Wang, Y.; Nallamothu, B.K.; Epstein, A.J.; Krumholz, H.M.; Hines, H.H. Association of door-to-balloon time and mortality in patients admitted to hospital with ST elevation myocardial infarction: National cohort study. BMJ 2009, 338, 1312–1315. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-Q.; Corvera, J.S.; Halkos, M.E.; Kerendi, F.; Wang, N.-P.; Guyton, R.A.; Vinten-Johansen, J. Inhibition of myocardial injury by ischemic postconditioning during reperfusion: Comparison with ischemic preconditioning. Am. J. Physiol. Circ. Physiol. 2015, 285, H579–H588. [Google Scholar] [CrossRef] [PubMed]
- Kloner, R.A.; Ganote, C.E.; Jennings, R.B. The “no reflow” phenomenon after temporary coronary occlusion in the dog. J. Clin. Invest. 1974, 54, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Bekkers, S.C.A.M.; Yazdani, S.K.; Virmani, R.; Waltenberger, J. Microvascular Obstruction. Underlying Pathophysiology and Clinical Diagnosis. J. Am. Coll. Cardiol. 2010, 55, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Niccoli, G.; Burzotta, F.; Galiuto, L.; Crea, F. Myocardial No-Reflow in Humans. J. Am. Coll. Cardiol. 2009, 54, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Eitel, I.; De Waha, S.; Wöhrle, J.; Fuernau, G.; Lurz, P.; Pauschinger, M.; Desch, S.; Schuler, G.; Thiele, H. Comprehensive prognosis assessment by CMR imaging after ST-segment elevation myocardial infarction. J. Am. Coll. Cardiol. 2014, 64, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Kleinbongard, P.; Baars, T.; Möhlenkamp, S.; Kahlert, P.; Erbel, R.; Heusch, G. Aspirate from human stented native coronary arteries vs. saphenous vein grafts: More endothelin but less particulate debris. Am. J. Physiol. Circ. Physiol. 2013, 305, H1222–H1229. [Google Scholar] [CrossRef] [PubMed]
- Kleinbongard, P.; Böse, D.; Baars, T.; Möhlenkamp, S.; Konorza, T.; Schöner, S.; Elter-Schulz, M.; Eggebrecht, H.; Degen, H.; Haude, M.; et al. Vasoconstrictor potential of coronary aspirate from patients undergoing stenting of saphenous vein aortocoronary bypass grafts and its pharmacological attenuation. Circ. Res. 2011, 108, 344–352. [Google Scholar] [CrossRef]
- Manciet, L.H.; Poole, D.C.; McDonagh, P.F.; Copeland, J.G.; Mathieu-Costello, O. Microvascular compression during myocardial ischemia: Mechanistic basis for no-reflow phenomenon. Am. J. Physiol. Circ. Physiol. 2017, 266, H1541–H1550. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Clarke, S.J.; Javadov, S.A. Mitochondrial permeability transition pore opening during myocardial reperfusion—A target for cardioprotection. Cardiovasc. Res. 2004, 61, 372–385. [Google Scholar] [CrossRef]
- Alavian, K.N.; Beutner, G.; Lazrove, E.; Sacchetti, S.; Park, H.-A.; Licznerski, P.; Li, H.; Nabili, P.; Hockensmith, K.; Graham, M.; et al. An uncoupling channel within the c-subunit ring of the F1FO ATP synthase is the mitochondrial permeability transition pore. Proc. Natl. Acad. Sci. USA 2014, 111, 10580–10585. [Google Scholar] [CrossRef] [PubMed]
- Petronilli, V.; Cola, C.; Bernardi, P. Modulation of the mitochondrial cyclosporin A-sensitive permeability transition pore. II. The minimal requirements for pore induction underscore a key role for transmembrane electrical potential, matrix pH, and matrix Ca2+. J. Biol. Chem. 1993, 268, 1011–1016. [Google Scholar] [PubMed]
- Griffiths, E.J.; Halestrap, A.P. Mitochondrial non-specific pores remain closed during cardiac ischaemia, but open upon reperfusion. Biochem. J. 2015, 307, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Steenbergen, C. What makes the mitochondria a killer? Can we condition them to be less destructive? Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1302–1308. [Google Scholar] [CrossRef] [Green Version]
- Giorgio, V.; Bisetto, E.; Soriano, M.E.; Dabbeni-Sala, F.; Basso, E.; Petronilli, V.; Forte, M.A.; Bernardi, P.; Lippe, G. Cyclophilin D modulates mitochondrial F0F1-ATP synthase by interacting with the lateral stalk of the complex. J. Biol. Chem. 2009, 284, 33982–33988. [Google Scholar] [CrossRef] [PubMed]
- Piot, C.; Croisille, P.; Staat, P.; Thibault, H.; Rioufol, G.; Mewton, N.; Elbelghiti, R.; Cung, T.T.; Bonnefoy, E.; Angoulvant, D.; et al. Effect of Cyclosporine on Reperfusion Injury in Acute Myocardial Infarction. N. Engl. J. Med. 2008, 359, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cung, T.-T.; Morel, O.; Cayla, G.; Rioufol, G.; Garcia-Dorado, D.; Angoulvant, D.; Bonnefoy-Cudraz, E.; Guérin, P.; Elbaz, M.; Delarche, N.; et al. Cyclosporine before PCI in Patients with Acute Myocardial Infarction. N. Engl. J. Med. 2015, 373, 1021–1031. [Google Scholar] [CrossRef]
- Atar, D.; Arheden, H.; Berdeaux, A.; Bonnet, J.L.; Carlsson, M.; Clemmensen, P.; Cuvier, V.; Danchin, N.; Dubois-Randé, J.L.; Engblom, H.; et al. Effect of intravenous TRO40303 as an adjunct to primary percutaneous coronary intervention for acute ST-elevation myocardial infarction: MITOCARE study results. Eur. Heart J. 2015, 36, 112–119. [Google Scholar] [CrossRef]
- Fliss, H.; Gattinger, D. Apoptosis in Ischemic and Reperfused Rat Myocardium. Circ. Res. 1996, 79, 949–956. [Google Scholar] [CrossRef]
- Schulman, D.; Latchman, D.S.; Yellon, D.M. Urocortin protects the heart from reperfusion injury via upregulation of p42/p44 MAPK signaling pathway. Am. J. Physiol. Circ. Physiol. 2002, 283, H1481–H1488. [Google Scholar] [CrossRef] [Green Version]
- Simpkin, J.C.; Yellon, D.M.; Davidson, S.M.; Lim, S.Y.; Wynne, A.M.; Smith, C.C.T. Apelin-13 and apelin-36 exhibit direct cardioprotective activity against ischemiareperfusioninjury. Basic Res. Cardiol. 2007, 102, 518. [Google Scholar] [CrossRef] [PubMed]
- Folino, A.; Accomasso, L.; Giachino, C.; Montarolo, P.G.; Losano, G.; Pagliaro, P.; Rastaldo, R. Apelin-induced cardioprotection against ischaemia/reperfusion injury: Roles of epidermal growth factor and Src. Acta Physiol. 2018, 222, e12924. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, T.; Matsui, T.; Aoyama, T.; Leri, A.; Anversa, P.; Li, L.; Ogawa, W.; del Monte, F.; Gwathmey, J.K.; Grazette, L.; et al. PI3K rescues the detrimental effects of chronic Akt activation in the heart during ischemia/reperfusion injury. J. Clin. Invest. 2005, 115, 2128–2138. [Google Scholar] [CrossRef] [PubMed]
- Yellon, D.M.; Hausenloy, D.J. Myocardial Reperfusion Injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Rossello, X.; Yellon, D.M. The RISK pathway and beyond. Basic Res. Cardiol. 2017, 113, 2. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, A. Cardioprotective signalling: Past, present and future. Eur. J. Pharmacol. 2018, 833, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, S.; Alidadiani, N.; Dilaver, N.; Heidari, H.R.; Parvizi, R.; Rahbarghazi, R.; Soleimani-Rad, J.; Baradaran, B. Role of glycogen synthase kinase following myocardial infarction and ischemia–reperfusion. Apoptosis 2017, 22, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Mocanu, M.M.; Field, D.C.; Yellon, D.M. A Potential Role for PTEN in the Diabetic Heart. Cardiovasc. Drugs Ther. 2006, 20, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Przyklenk, K.; Maynard, M.; Greiner, D.L.; Whittaker, P. Cardioprotection with Postconditioning: Loss of Efficacy in Murine Models of Type-2 and Type-1 Diabetes. Antioxid. Redox Signal. 2010, 14, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Lei, S.; Xia, Z.; Wu, Y.; Meng, Q.; Zhan, L.; Su, W.; Liu, H.; Xu, J.; Liu, Z.; et al. Selective inhibition of PTEN preserves ischaemic post-conditioning cardioprotection in STZ-induced Type 1 diabetic rats: Role of the PI3K/Akt and JAK2/STAT3 pathways. Clin. Sci. 2016, 130, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Mocanu, M.M.; Yellon, D.M. PTEN, the Achilles’ heel of myocardial ischaemia/reperfusion injury? Br. J. Pharmacol. 2007, 150, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Keyes, K.T.; Xu, J.; Long, B.; Zhang, C.; Hu, Z.; Ye, Y. Pharmacological inhibition of PTEN limits myocardial infarct size and improves left ventricular function postinfarction. Am. J. Physiol. Circ. Physiol. 2010, 298, H1198–H1208. [Google Scholar] [CrossRef] [PubMed]
- Boengler, K.; Hilfiker-Kleiner, D.; Heusch, G.; Schulz, R. Inhibition of permeability transition pore opening by mitochondrial STAT3 and its role in myocardial ischemia/reperfusion. Basic Res. Cardiol. 2010, 105, 771–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinliang, N.; Hengxun, H.; Yong, S.; Lianlian, Z.; Yingchao, W.; Zhiwei, Z.; Jing, Z.; Na, Z.; Ya, W.; Yaping, W.; et al. TNFR2 Stimulation Promotes Mitochondrial Fusion via Stat3- and NF-kB–Dependent Activation of OPA1 Expression. Circ. Res. 2017, 121, 392–410. [Google Scholar]
- Hadebe, N.; Cour, M.; Lecour, S. The SAFE pathway for cardioprotection: Is this a promising target? Basic Res. Cardiol. 2018, 113, 9. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, K.E.; Breen, E.P.; Gallagher, H.C.; Buggy, D.J.; Hurley, J.P. Understanding STAT3 signaling in cardiac ischemia. Basic Res. Cardiol. 2016, 111, 27. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S.; Suleman, N.; Deuchar, G.A.; Somers, S.; Lacerda, L.; Huisamen, B.; Opie, L.H. Pharmacological Preconditioning With Tumor Necrosis Factor-α Activates Signal Transducer and Activator of Transcription-3 at Reperfusion Without Involving Classic Prosurvival Kinases (Akt and Extracellular Signal–Regulated Kinase). Circulation 2005, 112, 3911–3918. [Google Scholar] [CrossRef] [PubMed]
- Somers, S.J.; Frias, M.; Lacerda, L.; Opie, L.H.; Lecour, S. Interplay Between SAFE and RISK Pathways in Sphingosine-1-Phosphate--Induced Cardioprotection. Cardiovasc. Drugs Ther. 2012, 26, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Lamont, K.T.; Somers, S.; Lacerda, L.; Opie, L.H.; Lecour, S. Is red wine a SAFE sip away from cardioprotection? Mechanisms involved in resveratrol- and melatonin-induced cardioprotection. J. Pineal Res. 2011, 50, 374–380. [Google Scholar] [CrossRef]
- Brulhart-Meynet, M.C.; Braunersreuther, V.; Brinck, J.; Montecucco, F.; Prost, J.C.; Thomas, A.; Galan, K.; Pelli, G.; Pedretti, S.; Vuilleumier, N.; et al. Improving reconstituted HDL composition for efficient post-ischemic reduction of ischemia reperfusion injury. PLoS ONE 2015, 10, e0119664. [Google Scholar] [CrossRef]
- Watson, A.J.; Gao, L.; Sun, L.; Tsun, J.; Jabbour, A.; Ru Qiu, M.; Jansz, P.C.; Hicks, M.; Macdonald, P.S. Enhanced preservation of the rat heart after prolonged hypothermic ischemia with erythropoietin-supplemented Celsior solution. J. Hear. Lung Transplant. 2013, 32, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, B.; Zhang, M.; Jin, Z.; Duan, W.; Zhao, G.; Yang, Y.; Liu, Z.; Chen, W.; Wang, S.; et al. Melatonin reduces PERK-eIF2$α$-ATF4-mediated endoplasmic reticulum stress during myocardial ischemia--reperfusion injury: Role of RISK and SAFE pathways interaction. Apoptosis 2016, 21, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Gough, D.J.; Koetz, L.; Levy, D.E. The MEK-ERK pathway is necessary for serine phosphorylation of mitochondrial STAT3 and ras-mediated transformation. PLoS ONE 2013, 8, e83395. [Google Scholar] [CrossRef]
- Heusch, G.; Musiolik, J.; Kottenberg, E.; Peters, J.; Jakob, H.; Thielmann, M. STAT5 activation and cardioprotection by remote ischemic preconditioning in humans. Circ. Res. 2012, 110, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Sharkovska, Y.; Kalk, P.; Lawrenz, B.; Godes, M.; Hoffmann, L.S.; Wellkisch, K.; Geschka, S.; Relle, K.; Hocher, B.; Stasch, J.-P. Nitric oxide-independent stimulation of soluble guanylate cyclase reduces organ damage in experimental low-renin and high-renin models. J. Hypertens. 2010, 28. [Google Scholar] [CrossRef] [PubMed]
- Salloum, F.N.; Das, A.; Samidurai, A.; Hoke, N.N.; Chau, V.Q.; Ockaili, R.A.; Stasch, J.-P.; Kukreja, R.C. Cinaciguat, a novel activator of soluble guanylate cyclase, protects against ischemia/reperfusion injury: Role of hydrogen sulfide. Am. J. Physiol. Circ. Physiol. 2012, 302, H1347–H1354. [Google Scholar] [CrossRef] [PubMed]
- Methner, C.; Buonincontri, G.; Hu, C.I.; Vujic, A.; Kretschmer, A.; Sawiak, S.; Carpenter, A.; Stasch, J.P.; Krieg, T. Riociguat reduces infarct size and post-infarct heart failure in mouse hearts: Insights from MRI/PET imaging. PLoS ONE 2013, 8, e83910. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Salloum, F.N.; Xi, L.; Rao, Y.J.; Kukreja, R.C. ERK phosphorylation mediates sildenafil-induced myocardial protection against ischemia-reperfusion injury in mice. Am. J. Physiol. Circ. Physiol. 2009, 296, H1236–H1243. [Google Scholar] [CrossRef]
- Salloum, F.N.; Takenoshita, Y.; Ockaili, R.A.; Daoud, V.P.; Chou, E.; Yoshida, K.; Kukreja, R.C. Sildenafil and vardenafil but not nitroglycerin limit myocardial infarction through opening of mitochondrial KATP channels when administered at reperfusion following ischemia in rabbits. J. Mol. Cell. Cardiol. 2007, 42, 453–458. [Google Scholar] [CrossRef]
- Costa, A.D.T.; Garlid, K.D.; West, I.C.; Lincoln, T.M.; Downey, J.M.; Cohen, M.V.; Critz, S.D. Protein kinase G transmits the cardioprotective signal from cytosol to mitochondria. Circ. Res. 2005, 97, 329–336. [Google Scholar] [CrossRef]
- Frankenreiter, S.; Bednarczyk, P.; Kniess, A.; Bork, N.I.; Straubinger, J.; Koprowski, P.; Wrzosek, A.; Mohr, E.; Logan, A.; Murphy, M.P.; et al. cGMP-Elevating Compounds and Ischemic Conditioning Provide Cardioprotection Against Ischemia and Reperfusion Injury via Cardiomyocyte-Specific BK Channels. Circulation 2017, 136, 2337–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methner, C.; Lukowski, R.; Grube, K.; Loga, F.; Smith, R.A.J.; Murphy, M.P.; Hofmann, F.; Krieg, T. Protection through postconditioning or a mitochondria-targeted S-nitrosothiol is unaffected by cardiomyocyte-selective ablation of protein kinase G. Basic Res. Cardiol. 2013, 108, 337. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [PubMed]
- Yutaka, M.; Hiromitsu, T.; Xueping, Q.; Maha, A.; Hideyuki, S.; Tomoichiro, A.; Beth, L.; Junichi, S. Distinct Roles of Autophagy in the Heart During Ischemia and Reperfusion. Circ. Res. 2007, 100, 914–922. [Google Scholar]
- Han, Z.; Cao, J.; Song, D.; Tian, L.; Chen, K.; Wang, Y.; Gao, L.; Yin, Z.; Fan, Y.; Wang, C. Autophagy is involved in the cardioprotection effect of remote limb ischemic postconditioning on myocardial ischemia/reperfusion injury in normal mice, but not diabetic mice. PLoS ONE 2014, 9, e86838. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Panda, P.K.; Sinha, N.; Das, D.N.; Bhutia, S.K. Autophagy and apoptosis: Where do they meet? Apoptosis 2014, 19, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Forte, M.; Frati, G.; Sadoshima, J. New insights into the role of mtor signaling in the cardiovascular system. Circ. Res. 2018, 122, 489–505. [Google Scholar] [CrossRef]
- Ma, X.; Liu, H.; Foyil, S.R.; Godar, R.J.; Weinheimer, C.J.; Diwan, A. Autophagy is impaired in cardiac ischemia-reperfusion injury. Autophagy 2012, 8, 1394–1396. [Google Scholar] [CrossRef] [Green Version]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741. [Google Scholar] [CrossRef]
- Randomised trial of intravenous streptokinase, oral aspirin, both, or neither among 17 187 cases of suspected acute myocardial infarction: ISIS-2. Lancet 1988, 332, 349–360. [CrossRef]
- Yusuf, S.; Mehta, S.; Anand, S.; Avezum, A.; Awan, N.; Bertrand, M.; Blumenthal, M.; Bouthier, J.; Budaj, A.; Ceremuzynski, L.; et al. The Clopidogrel in Unstable angina to prevent Recurrent Events (CURE) trial programme: Rationale, design and baseline characteristics including a meta-analysis of the effects of thienopyridines in vascular disease. Eur. Heart J. 2000, 21, 2033–2041. [Google Scholar]
- Chin, C.T.; Roe, M.T.; Fox, K.A.A.; Prabhakaran, D.; Marshall, D.A.; Petitjean, H.; Lokhnygina, Y.; Brown, E.; Armstrong, P.W.; White, H.D.; et al. Evaluation of prasugrel compared with clopidogrel in patients with acute coronary syndromes: Design and rationale for the TRial to assess Improvement in Therapeutic Outcomes by optimizing platelet InhibitioN with prasugrel Thrombolysis In Myocardial Infar. Am. Heart J. 2006, 152, 16–22. [Google Scholar]
- James, S.; Åkerblom, A.; Cannon, C.P.; Emanuelsson, H.; Husted, S.; Katus, H.; Skene, A.; Steg, P.G.; Storey, R.F.; Harrington, R.; et al. Comparison of ticagrelor, the first reversible oral P2Y12 receptor antagonist, with clopidogrel in patients with acute coronary syndromes: Rationale, design, and baseline characteristics of the PLATelet inhibition and patient Outcomes (PLATO) trial. Am. Heart J. 2009, 157, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, G.; Fernández-Friera, L.; Fuster, V.; Fernández-Jiménez, R.; García-Ruiz, J.M.; García-Álvarez, A.; Mateos, A.; Barreiro, M.V.; Escalera, N.; Rodriguez, M.D.; et al. Long-term benefit of early pre-reperfusion metoprolol administration in patients with acute myocardial infarction: Results from the Metocard-CNIC trial (Effect of Metoprolol in Cardioprotection during an Acute Myocardial Infarction). J. Am. Coll. Cardiol. 2014, 63, 2356–2362. [Google Scholar] [CrossRef] [PubMed]
- Abbate, A.; Kontos, M.C.; Grizzard, J.D.; Biondi-Zoccai, G.G.L.; Van Tassell, B.W.; Robati, R.; Roach, L.M.; Arena, R.A.; Roberts, C.S.; Varma, A.; et al. Interleukin-1 Blockade With Anakinra to Prevent Adverse Cardiac Remodeling After Acute Myocardial Infarction (Virginia Commonwealth University Anakinra Remodeling Trial [VCU-ART] Pilot Study). Am. J. Cardiol. 2010, 105, 1371–1377.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, A.C.; Rothman, A.M.K.; Greenwood, J.P.; Gunn, J.; Chase, A.; Clarke, B.; Hall, A.S.; Fox, K.; Foley, C.; Banya, W.; et al. The effect of interleukin-1 receptor antagonist therapy on markers of inflammation in non-ST elevation acute coronary syndromes: The MRC-ILA Heart Study. Eur. Heart J. 2015, 36, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef]
- Ross, A.M.; Gibbons, R.J.; Stone, G.W.; Kloner, R.A.; Alexander, R.W. A randomized, double-blinded, placebo-controlled multicenter trial of adenosine as an adjunct to reperfusion in the treatment of acute myocardial infarction (AMISTAD-II). J. Am. Coll. Cardiol. 2005, 45, 1775–1780. [Google Scholar] [CrossRef]
- Kloner, R.A.; Forman, M.B.; Gibbons, R.J.; Ross, A.M.; Alexander, R.W.; Stone, G.W. Impact of time to therapy and reperfusion modality on the efficacy of adenosine in acute myocardial infarction: The AMISTAD-2 trial. Eur. Heart J. 2006, 27, 2400–2405. [Google Scholar] [CrossRef]
- Garcia-Dorado, D.; García-Del-Blanco, B.; Otaegui, I.; Rodríguez-Palomares, J.; Pineda, V.; Gimeno, F.; Ruiz-Salmerón, R.; Elizaga, J.; Evangelista, A.; Fernandez-Avilés, F.; et al. Intracoronary injection of adenosine before reperfusion in patients with ST-segment elevation myocardial infarction: A randomized controlled clinical trial. Int. J. Cardiol. 2014, 177, 935–941. [Google Scholar] [CrossRef]
- Desmet, W.; Bogaert, J.; Dubois, C.; Sinnaeve, P.; Adriaenssens, T.; Pappas, C.; Ganame, J.; Dymarkowski, S.; Janssens, S.; Belmans, A.; et al. High-dose intracoronary adenosine for myocardial salvage in patients with acute ST-segment elevation myocardial infarction. Eur. Heart J. 2011, 32, 867–877. [Google Scholar] [CrossRef]
- Siddiqi, N.; Neil, C.; Bruce, M.; MacLennan, G.; Cotton, S.; Papadopoulou, S.; Feelisch, M.; Bunce, N.; Lim, P.O.; Hildick-Smith, D.; et al. Intravenous sodium nitrite in acute ST-elevation myocardial infarction: A randomized controlled trial (NIAMI). Eur. Heart J. 2014, 35, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Bøtker, H.E.; Kharbanda, R.; Schmidt, M.R.; Bøttcher, M.; Kaltoft, A.K.; Terkelsen, C.J.; Munk, K.; Andersen, N.H.; Hansen, T.M.; Trautner, S.; et al. Remote ischaemic conditioning before hospital admission, as a complement to angioplasty, and effect on myocardial salvage in patients with acute myocardial infarction: A randomised trial. Lancet 2010, 375, 727–734. [Google Scholar] [CrossRef]
- Staat, P.; Rioufol, G.; Piot, C.; Cottin, Y.; Cung, T.T.; L’Huillier, I.; Aupetit, J.F.; Bonnefoy, E.; Finet, G.; André-Fouët, X.; et al. Postconditioning the human heart. Circulation 2005, 112, 2143–2148. [Google Scholar] [CrossRef] [PubMed]
- Thibault, H.; Piot, C.; Staat, P.; Bontemps, L.; Sportouch, C.; Rioufol, G.; Cung, T.T.; Bonnefoy, E.; Angoulvant, D.; Aupetit, J.F.; et al. Long-term benefit of postconditioning. Circulation 2008, 117, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.Y.; Song, Y.B.; Kim, E.K.; Yu, C.W.; Bae, J.W.; Chung, W.Y.; Choi, S.H.; Choi, J.H.; Bae, J.H.; An, K.J.; et al. Ischemic postconditioning during primary percutaneous coronary intervention: The effects of postconditioning on myocardial reperfusion in patients with st-segment elevation myocardial infarction (POST) randomized trial. Circulation 2013, 128, 1889–1896. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Hahn, J.Y.; Song, Y.B.; Lee, S.C.; Choi, J.H.; Choi, S.H.; Lee, S.H.; Choe, Y.H.; Gwon, H.C. Effect of ischemic postconditioning on myocardial salvage in patients undergoing primary percutaneous coronary intervention for ST-segment elevation myocardial infarction: Cardiac magnetic resonance substudy of the POST randomized trial. Int. J. Cardiovasc. Imaging 2015, 31, 629–637. [Google Scholar] [CrossRef]
- Engstrøm, T.; Kelbæk, H.; Helqvist, S.; Høfsten, D.E.; Kløvgaard, L.; Clemmensen, P.; Holmvang, L.; Jørgensen, E.; Pedersen, F.; Saunamaki, K.; et al. Effect of ischemic postconditioning during primary percutaneous coronary intervention for patients with ST-segment elevation myocardial infarction: A randomized clinical trial. JAMA Cardiol. 2017, 2, 490–497. [Google Scholar] [CrossRef]
- Patel, M.R.; Smalling, R.W.; Thiele, H.; Barnhart, H.X.; Zhou, Y.; Chandra, P.; Chew, D.; Cohen, M.; French, J.; Perera, D.; et al. Intra-aortic balloon counterpulsation and infarct size in patients with acute anterior myocardial infarction without shock: The CRISP AMI randomized trial. JAMA J. Am. Med. Assoc. 2011, 306, 1329–1337. [Google Scholar] [CrossRef]
- Van Nunen, L.X.; Van ’TVeer, M.; Schampaert, S.; Rutten, M.C.M.; Van De Vosse, F.N.; Patel, M.R.; Pijls, N.H.J. Intra-aortic balloon counterpulsation reduces mortality in large anterior myocardial infarction complicated by persistent ischaemia: A CRISP-AMI substudy. EuroIntervention 2015, 11, 286–292. [Google Scholar] [CrossRef]
- De Maria, G.L.; Alkhalil, M.; Borlotti, A.; Wolfrum, M.; Gaughran, L.; Dall’Armellina, E.; Langrish, J.P.; Lucking, A.J.; Choudhury, R.P.; Kharbanda, R.K.; et al. Index of microcirculatory resistance-guided therapy with pressure-controlled intermittent coronary sinus occlusion improves coronary microvascular function and reduces infarct size in patients with ST-elevation myocardial infarction: The Oxford Acute Myoc. EuroIntervention 2018, 14, e352–e359. [Google Scholar] [CrossRef] [PubMed]
- Dewood, M.A.; Spores, J.; Notske, R.; Mouser, L.T.; Burroughs, R.; Golden, M.S.; Lang, H.T.; Dewood, M.A. Prevalence of Total Coronary Occlusion during the Early Hours of Transmural Myocardial Infarction. N. Engl. J. Med. 1980, 303, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Chazov, E.I.; Matveeva, L.S.; Mazaev, A.V.; Sargin, K.E.; Sadovskaia, G.V.; Ruda, M.I. [Intracoronary administration of fibrinolysin in acute myocardial infarct]. Ter. Arkh. 1976, 48, 8–19. [Google Scholar] [PubMed]
- Hartzler, G.O.; Rumerford, B.D.; McConahay, D.R.; Johnson, W.L.; McCallister, B.D.; Gura, G.M.; Conn, R.C.; Crockett, J.E. Percutaneous transluminal coronary angioplasty with and without thrombolytic therapy for treatment of acute myocardial infarction. Am. Heart J. 1983, 106, 965–973. [Google Scholar] [CrossRef]
- Chen, Z.; Jiang, L. Addition of clopidogrel to aspirin in 45 852 patients with acute myocardial infarction: Randomised placebo-controlled trial. Lancet 2005, 366, 1607–1621. [Google Scholar]
- Ibanez, B.; Fuster, V.; MacAya, C.; Sánchez-Brunete, V.; Pizarro, G.; López-Romero, P.; Mateos, A.; Jiménez-Borreguero, J.; Fernández-Ortiz, A.; Sanz, G.; et al. Study design for the effect of METOprolol in CARDioproteCtioN during an acute myocardial InfarCtion (METOCARD-CNIC): A randomized, controlled parallel-group, observer-blinded clinical trial of early pre-reperfusion metoprolol administration in ST-segment. Am. Heart J. 2012, 164, 473–480.e5. [Google Scholar] [CrossRef]
- Garcia-Prieto, J.; Fernandez-Jimenez, R.; Sreeramkumar, V.; Lunar, I.G.; Sanz-Rosa, D.; Zorita, V.; Pizarro, G.; Hidalgo, A.; Fuster, V.; Ibanez, B. 272Pre-reperfusion metoprolol administration reduces ischemia/reperfusion injury (IRI) through beta1-adrenergic receptor (b1AR) blockade in the circulating cells. Cardiovasc. Res. 2014, 103, S49. [Google Scholar] [CrossRef]
- Harouki, N.; Nicol, L.; Remy-Jouet, I.; Henry, J.P.; Dumesnil, A.; Lejeune, A.; Renet, S.; Golding, F.; Djerada, Z.; Wecker, D.; et al. The IL-1β Antibody Gevokizumab Limits Cardiac Remodeling and Coronary Dysfunction in Rats With Heart Failure. JACC Basic Transl. Sci. 2017, 2, 418–430. [Google Scholar] [CrossRef]
- Abbate, A.; Van Tassell, B.W.; Biondi-Zoccai, G.; Kontos, M.C.; Grizzard, J.D.; Spillman, D.W.; Oddi, C.; Roberts, C.S.; Melchior, R.D.; Mueller, G.H.; et al. Effects of interleukin-1 blockade with anakinra on adverse cardiac remodeling and heart failure after acute myocardial infarction [from the virginia commonwealth university-anakinra remodeling trial (2) (vcu-art2) pilot study]. Am. J. Cardiol. 2013, 111, 1394–1400. [Google Scholar] [CrossRef]
- Garratt, K.N.; Holmes, D.R.J.; Molina-Viamonte, V.; Reeder, G.S.; Hodge, D.O.; Bailey, K.R.; Lobl, J.K.; Laudon, D.A.; Gibbons, R.J. Intravenous adenosine and lidocaine in patients with acute myocardial infarction. Am. Heart J. 1998, 136, 196–204. [Google Scholar] [CrossRef]
- Marzilli, M.; Orsini, E.; Marraccini, P.; Testa, R. Beneficial effects of intracoronary adenosine as an adjunct to primary angioplasty in acute myocardial infarction. Circulation 2000, 101, 2154–2159. [Google Scholar] [CrossRef] [PubMed]
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D.J.; Kharbanda, R.K.; Møller, U.K.; Ramlall, M.; Aarøe, J.; Butler, R.; Bulluck, H.; Clayton, T.; Dana, A.; Dodd, M.; et al. Effect of remote ischaemic conditioning on clinical outcomes in patients with acute myocardial infarction (CONDI-2/ERIC-PPCI): A single-blind randomised controlled trial. Lancet 2019. [Google Scholar] [CrossRef]
- LeDoux, J.F.; Tamareille, S.; Fellis, P.R.; Amirian, J.; Smalling, R.W. Left ventricular unloading with intra-aortic counter pulsation prior to reperfusion reduces myocardial release of endothelin-1 and decreases infarction size in a porcine ischemia-reperfusion model. Catheter. Cardiovasc. Interv. 2008, 72, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Pierrakos, C.N.; Bonios, M.J.; Drakos, S.G.; Charitos, E.I.; Tsolakis, E.J.; Ntalianis, A.; Nanas, S.N.; Charitos, C.E.; Nanas, J.N.; Terrovitis, J. V Mechanical assistance by intra-aortic balloon pump counterpulsation during reperfusion increases coronary blood flow and mitigates the no-reflow phenomenon: An experimental study. Artif. Organs 2011, 35, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Esposito, M.L.; Zhang, Y.; Qiao, X.; Reyelt, L.; Paruchuri, V.; Schnitzler, G.R.; Morine, K.J.; Annamalai, S.K.; Bogins, C.; Natov, P.S.; et al. Left Ventricular Unloading Before Reperfusion Promotes Functional Recovery After Acute Myocardial Infarction. J. Am. Coll. Cardiol. 2018, 72, 501–514. [Google Scholar] [CrossRef]
- Kapur, N.K.; Alkhouli, M.A.; DeMartini, T.J.; Faraz, H.; George, Z.H.; Goodwin, M.J.; Hernandez-Montfort, J.A.; Iyer, V.S.; Josephy, N.; Kalra, S.; et al. Unloading the Left Ventricle Before Reperfusion in Patients With Anterior ST-Segment-Elevation Myocardial Infarction: A Pilot Study Using the Impella CP. Circulation 2019, 139, 337–346. [Google Scholar] [CrossRef]
- Massey, H.T.; Choi, J.H.; Maynes, E.J.; Tchantchaleishvili, V. Temporary support strategies for cardiogenic shock: Extracorporeal membrane oxygenation, percutaneous ventricular assist devices and surgically placed extracorporeal ventricular assist devices. Ann. Cardiothorac. Surg. 2019, 8, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Weigel, G.; Kajgana, I.; Bergmeister, H.; Riedl, G.; Glogar, H.D.; Gyöngyösi, M.; Blasnig, S.; Heinze, G.; Mohl, W. Beck and back: A paradigm change in coronary sinus interventions-pulsatile stretch on intact coronary venous endothelium. J. Thorac. Cardiovasc. Surg. 2007, 133, 1581–1587. [Google Scholar] [CrossRef]
- Mohl, W.; Mina, S.; Milasinovic, D.; Kasahara, H.; Wei, S.; Maurer, G. Is activation of coronary venous cells the key to cardiac regeneration? Nat. Clin. Pract. Cardiovasc. Med. 2008, 5, 528–530. [Google Scholar] [CrossRef]
- Wang, Z.; Su, X.; Ashraf, M.; Kim, I.M.; Weintraub, N.L.; Jiang, M.; Tang, Y. Regenerative therapy for cardiomyopathies. J. Cardiovasc. Transl. Res. 2018, 11, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Terashvili, M.; Bosnjak, Z.J. Stem Cell Therapies in Cardiovascular Disease. J. Cardiothorac. Vasc. Anesth. 2019, 33, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt-Velin, N.; Badoux, S.; Liaudet, L. Pharmacological therapy in the heart as an alternative to cellular therapy: A place for the brain natriuretic peptide. Stem Cells Int. 2016, 2016, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bielmann, C.; Rignault-Clerc, S.; Liaudet, L.; Li, F.; Kunieda, T.; Sogawa, C.; Zehnder, T.; Waeber, B.; Feihl, F.; Rosenblatt-Velin, N. Brain natriuretic peptide is able to stimulate cardiac progenitor cell proliferation and differentiation in murine hearts after birth. Basic Res. Cardiol. 2015, 110, 455. [Google Scholar] [CrossRef]
- Rignault-Clerc, S.; Bielmann, C.; Liaudet, L.; Waeber, B.; Feihl, F.; Rosenblatt-Velin, N. Natriuretic Peptide Receptor B modulates the proliferation of the cardiac cells expressing the Stem Cell Antigen-1. Sci. Rep. 2017, 7, 41936. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhu, W.; Cheng, M.; Chen, L.; Zhang, J.; Sun, T.; Kishore, R.; Phillips, M.I.; Losordo, D.W.; Qin, G. Hypoxic preconditioning enhances the benefit of cardiac progenitor cell therapy for treatment of myocardial infarction by inducing CXCR4 expression. Circ. Res. 2009, 104, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, I.; Smith, R.R.; Li, T.S.; Gerstenblith, G.; Messina, E.; Giacomello, A.; Marbán, E. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ. Res. 2010, 106, 971–980. [Google Scholar] [CrossRef]
- Vicencio, J.M.; Yellon, D.M.; Sivaraman, V.; Das, D.; Boi-Doku, C.; Arjun, S.; Zheng, Y.; Riquelme, J.A.; Kearney, J.; Sharma, V.; et al. Plasma exosomes protect the myocardium from ischemia-reperfusion injury. J. Am. Coll. Cardiol. 2015, 65, 1525–1536. [Google Scholar] [CrossRef]
- Kishore, R.; Khan, M. Cardiac cell-derived exosomes: Changing face of regenerative biology. Eur. Heart J. 2017, 38, 212–215. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7. [Google Scholar] [CrossRef]
- Pan, B.T.; Teng, K.; Wu, C.; Adam, M.; Johnstone, R.M. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol. 1985, 101, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [PubMed]
- Denzer, K.; Kleijmeer, M.J.; Heijnen, H.F.; Stoorvogel, W.; Geuze, H.J. Exosome: From internal vesicle of the multivesicular body to intercellular signaling device. J. Cell Sci. 2000, 113, 3365–3374. [Google Scholar] [PubMed]
- Hanson, P.I.; Cashikar, A. Multivesicular Body Morphogenesis. Annu. Rev. Cell Dev. Biol. 2012, 28, 337–362. [Google Scholar] [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Szatanek, R.; Baj-Krzyworzeka, M.; Zimoch, J.; Lekka, M.; Siedlar, M.; Baran, J. The methods of choice for extracellular vesicles (EVs) characterization. Int. J. Mol. Sci. 2017, 18, 1153. [Google Scholar] [CrossRef]
- Wendt, S.; Goetzenich, A.; Goettsch, C.; Stoppe, C.; Bleilevens, C.; Kraemer, S.; Benstoem, C. Evaluation of the cardioprotective potential of extracellular vesicles—A systematic review and meta-analysis. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Nomura, S. Microparticle and atherothrombotic diseases. J. Atheroscler. Thromb. 2016, 23, 1–9. [Google Scholar] [CrossRef]
- Mooberry, M.J.; Key, N.S. Microparticle analysis in disorders of hemostasis and thrombosis. Cytom. Part A 2016, 89, 111–122. [Google Scholar] [CrossRef]
- Brill, A.; Dashevsky, O.; Rivo, J.; Gozal, Y.; Varon, D. Platelet-derived microparticles induce angiogenesis and stimulate post-ischemic revascularization. Cardiovasc. Res. 2005, 67, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Leroyer, A.S.; Isobe, H.; Lesèche, G.; Castier, Y.; Wassef, M.; Mallat, Z.; Binder, B.R.; Tedgui, A.; Boulanger, C.M. Cellular Origins and Thrombogenic Activity of Microparticles Isolated From Human Atherosclerotic Plaques. J. Am. Coll. Cardiol. 2007, 49, 772–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, M.; Grainger, D.; Mayr, U.; Leroyer, A.S.; Leseche, G.; Sidibe, A.; Herbin, O.; Yin, X.; Gomes, A.; Madhu, B.; et al. Proteomics, metabolomics, and immunomics on microparticles derived from human atherosclerotic plaques. Circ. Cardiovasc. Genet. 2009, 2, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Tandon, N.N.; Nakamura, T.; Cone, J.; Fukuhara, S.; Kambayashi, J. High-shear-stress-induced activation of platelets and microparticles enhances expression of cell adhesion molecules in THP-1 and endothelial cells. Atherosclerosis 2001, 158, 277–287. [Google Scholar] [CrossRef]
- Ma, F.; Liu, H.; Shen, Y.; Zhang, Y.; Pan, S. Platelet-derived microvesicles are involved in cardio-protective effects of remote preconditioning. Int. J. Clin. Exp. Pathol. 2015, 8, 10832–10839. [Google Scholar] [PubMed]
- Giricz, Z.; Varga, Z.V.; Baranyai, T.; Sipos, P.; Pálóczi, K.; Kittel, Á.; Buzás, E.I.; Ferdinandy, P. Cardioprotection by remote ischemic preconditioning of the rat heart is mediated by extracellular vesicles. J. Mol. Cell. Cardiol. 2014, 68, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Wang, Y.; Zhu, Q.; Zhao, J.; Wang, Y.; Shang, M.; Liu, M.; Wu, Y.; Song, J.; Liu, Y. Protective effects of circulating microvesicles derived from ischemic preconditioning on myocardial ischemia/reperfusion injury in rats by inhibiting endoplasmic reticulum stress. Apoptosis 2018, 23, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Jeanneteau, J.; Hibert, P.; Martinez, M.C.; Tual-Chalot, S.; Tamareille, S.; Furber, A.; Andriantsitohaina, R.; Prunier, F. Microparticle release in remote ischemic conditioning mechanism. Am. J. Physiol. Hear. Circ. Physiol. 2012, 303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shang, M.; Zhang, M.; Wang, Y.; Chen, Y.; Wu, Y.; Liu, M.; Song, J.; Liu, Y. Microvesicles derived from hypoxia/reoxygenation-treated human umbilical vein endothelial cells promote apoptosis and oxidative stress in H9c2 cardiomyocytes. BMC Cell Biol. 2016, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Loyer, X.; Zlatanova, I.; Devue, C.; Yin, M.; Howangyin, K.Y.; Klaihmon, P.; Guerin, C.L.; Khelouf, M.; Vilar, J.; Zannis, K.; et al. Intra-cardiac release of extracellular vesicles shapes inflammation following myocardial infarction short communication. Circ. Res. 2018, 123, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Gong, M.; Wang, Y.; Millard, R.W.; Pasha, Z.; Yang, Y.; Ashraf, M.; Xu, M. Cardiomyocyte Protection by GATA-4 Gene Engineered Mesenchymal Stem Cells Is Partially Mediated by Translocation of miR-221 in Microvesicles. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, Y.; Shang, M.; Wang, Y.; Zhang, Q.; Wang, S.; Wei, S.; Zhang, K.; Liu, C.; Wu, Y.; et al. Flow cytometric analysis of circulating microvesicles derived from myocardial Ischemic preconditioning and cardioprotection of Ischemia/reperfusion Injury in rats. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2015, 31, 524–531. [Google Scholar] [PubMed]
- Barry, O.P.; Praticò, D.; Lawson, J.A.; FitzGerald, G.A. Transcellular activation of platelets and endothelial cells by bioactive lipids in platelet microparticles. J. Clin. Invest. 1997, 99, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- Sims, P.J.; Wiedmer, T.; Esmon, C.T.; Weiss, H.J.; Shattil, S.J. Assembly of the platelet prothrombinase complex is linked to vesiculation of the platelet plasma membrane. Studies in Scott syndrome: An isolated defect in platelet procoagulant activity. J. Biol. Chem. 1989, 264, 17049–17057. [Google Scholar] [PubMed]
- Jung, C.; Sörensson, P.; Saleh, N.; Arheden, H.; Rydén, L.; Pernow, J. Circulating endothelial and platelet derived microparticles reflect the size of myocardium at risk in patients with ST-elevation myocardial infarction. Atherosclerosis 2012, 221, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Chironi, G.N.; Boulanger, C.M.; Simon, A.; Dignat-George, F.; Freyssinet, J.M.; Tedgui, A. Endothelial microparticles in diseases. Cell Tissue Res. 2009, 335, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.; Yang, N.; Nadithe, V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: Current perspectives and future challenges. Acta Pharm. Sin. B 2016, 6, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.C.; Arslan, F.; Lee, M.M.; Sze, N.S.K.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Huang, W.; Wani, M.; Yu, X.; Ashraf, M. Ischemic Preconditioning Potentiates the Protective Effect of Stem Cells through Secretion of Exosomes by Targeting Mecp2 via miR-22. PLoS ONE 2014, 9, 841–848. [Google Scholar] [CrossRef]
- Yu, B.; Kim, H.W.; Gong, M.; Wang, J.; Millard, R.W.; Wang, Y.; Ashraf, M.; Xu, M. Exosomes secreted from GATA-4 overexpressing mesenchymal stem cells serve as a reservoir of anti-apoptotic microRNAs for cardioprotection. Int. J. Cardiol. 2015, 182, 349–360. [Google Scholar] [CrossRef]
- Wang, K.; Jiang, Z.; Webster, K.A.; Chen, J.; Hu, H.; Zhou, Y.; Zhao, J.; Wang, L.; Wang, Y.; Zhong, Z.; et al. Enhanced Cardioprotection by Human Endometrium Mesenchymal Stem Cells Driven by Exosomal MicroRNA-21. Stem Cells Transl. Med. 2017, 6, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jin, X.; Hu, C.-F.; Li, R.; Zhou, Z.; Shen, C.-X. Exosomes Derived from Mesenchymal Stem Cells Rescue Myocardial Ischaemia/Reperfusion Injury by Inducing Cardiomyocyte Autophagy Via AMPK and Akt Pathways. Cell. Physiol. Biochem. 2017, 43, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Changchen, X.; Kan, W.; Yinchuan, X.; Hengxun, H.; Na, Z.; Yingchao, W.; Zhiwei, Z.; Jing, Z.; Qingju, L.; Dan, Z.; et al. Transplanted Mesenchymal Stem Cells Reduce Autophagic Flux in Infarcted Hearts via the Exosomal Transfer of miR-125b. Circ. Res. 2018, 123, 564–578. [Google Scholar]
- Jiang, X.; Lew, K.-S.; Chen, Q.; Wang, A.M.R.; Human, P. Mesenchymal Stem Cell-derived Exosomes Reduce Ischemia/Reperfusion Injury by the Inhibitions of Apoptosis and Autophagy. Curr. Pharm. Des. 2018, 24, 5334–5341. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; He, Z.; Liang, Z.; Chen, Z.; Wang, H.; Zhang, J. Exosomes from Adipose-derived Mesenchymal Stem Cells Protect the Myocardium Against Ischemia/Reperfusion Injury Through Wnt/b-Catenin Signaling Pathway. J. Cardiovasc. Pharmacol. 2017, 70, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Zilun, W.; Shuaihua, Q.; Jinxuan, Z.; Yihai, L.; Qiaoling, L.; Zhonghai, W.; Qing, D.; Lina, K.; Biao, X. miRNA-181a over-expression in mesenchymal stem cell-derived exosomes influenced inflammatory response after myocardial ischemia-reperfusion injury. Life Sci. 2019, 116632. [Google Scholar]
- Huang, P.; Wang, L.; Li, Q.; Tian, X.; Xu, J.; Xu, J.; Xiong, Y.; Chen, G.; Qian, H.; Jin, C.; et al. Atorvastatin enhances the therapeutic efficacy of mesenchymal stem cells-derived exosomes in acute myocardial infarction via up-regulating long non-coding RNA H19. Cardiovasc. Res. 2019. [Google Scholar] [CrossRef]
- Kang, K.; Ma, R.; Cai, W.; Huang, W.; Paul, C.; Liang, J.; Wang, Y.; Zhao, T.; Kim, H.W.; Xu, M.; et al. Exosomes Secreted from CXCR4 Overexpressing Mesenchymal Stem Cells Promote Cardioprotection via Akt Signaling Pathway following Myocardial Infarction. Stem Cells Int. 2015, 2015. [Google Scholar] [CrossRef]
- Ma, T.; Chen, Y.; Chen, Y.; Meng, Q.; Sun, J.; Shao, L.; Yu, Y.; Huang, H.; Hu, Y.; Yang, Z.; et al. MicroRNA-132, delivered by mesenchymal stem cell-derived exosomes, promote angiogenesis in myocardial infarction. Stem Cells Int. 2018, 2018. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, J.; Yan, W.; Li, Y.; Shen, Z.; Asahara, T. Pretreatment of cardiac stem cells with exosomes derived from mesenchymal stem cells enhances myocardial repair. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.; Pan, Y.; Zhang, L.; Shen, C.; Qin, G.; Ashraf, M.; Weintraub, N.; Ma, G.; Tang, Y. Cardiac progenitor-derived exosomes protect ischemic myocardium from acute ischemia/reperfusion injury. Biochem. Biophys. Res. Commun. 2013, 431, 566–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, A.G.E.; Cheng, K.; Marbán, E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Reports 2014, 2, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular vesicles fromhuman cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function aftermyocardial infarction. Cardiovasc. Res. 2014, 103, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Cervio, E.; Lionetti, V.; Milano, G.; Ciullo, A.; Biemmi, V.; Bolis, S.; Altomare, C.; Matteucci, M.; Di Silvestre, D.; et al. Cardioprotection by cardiac progenitor cell-secreted exosomes: Role of pregnancy-associated plasma protein-A. Cardiovasc. Res. 2018, 114, 992–1005. [Google Scholar] [CrossRef] [PubMed]
- Cambier, L.; de Couto, G.; Ibrahim, A.; Echavez, A.K.; Valle, J.; Liu, W.; Kreke, M.; Smith, R.R.; Marbán, L.; Marbán, E. Y RNA fragment in extracellular vesicles confers cardioprotection via modulation of IL-10 expression and secretion. EMBO Mol. Med. 2017, 9, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Gallet, R.; Dawkins, J.; Valle, J.; Simsolo, E.; De Couto, G.; Middleton, R.; Tseliou, E.; Luthringer, D.; Kreke, M.; Smith, R.R.; et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 2017, 38, 201–211. [Google Scholar] [CrossRef]
- Xiao, J.; Pan, Y.; Li, X.H.; Yang, X.Y.; Feng, Y.L.; Tan, H.H.; Jiang, L.; Feng, J.; Yu, X.Y. Cardiac progenitor cell-derived exosomes prevent cardiomyocytes apoptosis through exosomal miR-21 by targeting PDCD4. Cell Death Dis. 2016, 7, e2277. [Google Scholar]
- Youn, S.-W.; Li, Y.; Kim, Y.-M.; Sudhahar, V.; Abdelsaid, K.; Kim, H.; Liu, Y.; Fulton, D.; Ashraf, M.; Tang, Y.; et al. Modification of Cardiac Progenitor Cell-Derived Exosomes by miR-322 Provides Protection against Myocardial Infarction through Nox2-Dependent Angiogenesis. Antioxidants 2019, 8, 18. [Google Scholar] [CrossRef]
- Ciullo, A.; Biemmi, V.; Milano, G.; Bolis, S.; Cervio, E.; Fertig, E.T.; Gherghiceanu, M.; Moccetti, T.; Camici, G.G.; Vassalli, G.; et al. Exosomal expression of CXCR4 targets cardioprotective vesicles to myocardial infarction and improves outcome after systemic administration. Int. J. Mol. Sci. 2019, 20, 468. [Google Scholar] [CrossRef]
- Khan, M.; Nickoloff, E.; Abramova, T.; Johnson, J.; Verma, S.K.; Krishnamurthy, P.; Mackie, A.R.; Vaughan, E.; Garikipati, V.N.S.; Benedict, C.; et al. Embryonic Stem Cell-Derived Exosomes Promote Endogenous Repair Mechanisms and Enhance Cardiac Function Following Myocardial Infarction. Circ. Res. 2015, 117, 52–64. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, M.; Deng, S.; Lu, J.; Huang, H.; Zhang, Y.; Gong, P.; Shen, X.; Ruan, H.; Jin, M.; et al. miR-93-5p-Containing Exosomes Treatment Attenuates Acute Myocardial Infarction-Induced Myocardial Damage. Mol. Ther. Nucleic Acids 2018, 11, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Alimujiang, M.; Chen, Q.; Shi, H.; Luo, X. Exosomes derived from miR-146a-modified adipose-derived stem cells attenuate acute myocardial infarction−induced myocardial damage via downregulation of early growth response factor 1. J. Cell. Biochem. 2019, 120, 4433–4443. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Guo, D.; Liu, G.; Chen, G.; Hang, M.; Jin, M. Exosomes from MiR-126-Overexpressing Adscs Are Therapeutic in Relieving Acute Myocardial Ischaemic Injury. Cell. Physiol. Biochem. 2018, 44, 2105–2116. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, Y.; Chen, X.; Cheng, X.; Liao, Y.; Yu, X. Exosomal transfer of miR-30a between cardiomyocytes regulates autophagy after hypoxia. J. Mol. Med. 2016, 94, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Liao, Z.; Chen, S.; Yuan, Z.; Yilin, C.; Lee, K.K.; Qi, X.; Shen, X.; Zheng, X.; Quinn, T.; et al. Intramyocardial transplantation of cardiac telocytes decreases myocardial infarction and improves post-infarcted cardiac function in rats. J. Cell. Mol. Med. 2014, 18, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Manole, C.G.; Cismaşiu, V.; Gherghiceanu, M.; Popescu, L.M. Experimental acute myocardial infarction: Telocytes involvement in neo-angiogenesis. J. Cell. Mol. Med. 2011, 15, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gao, W.; Yuan, J.; Wu, C.; Yao, K.; Zhang, L.; Ma, L.; Zhu, J.; Zou, Y.; Ge, J. Exosomes derived from dendritic cells improve cardiac function via activation of CD4 + T lymphocytes after myocardial infarction. J. Mol. Cell. Cardiol. 2016, 91, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liao, Y.; Gao, L.; Zhuang, T.; Huang, Z.; Zhu, H.; Ge, J. Coronary serum exosomes derived from patients with myocardial ischemia regulate angiogenesis through the miR-939-mediated nitric oxide signaling pathway. Theranostics 2018, 8, 2079–2093. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Bradley, M.J. Mesenchymal Stem Cells and Their Potential as Cardiac Therapeutics. Circ. Res. 2004, 95, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi, M.; He, H.; Noiseux, N.; Liang, O.D.; Zhang, L.; Morello, F.; Mu, H.; Melo, L.G.; Pratt, R.E.; Ingwall, J.S.; et al. Evidence supporting paracrine hypothesis for Akt-modified mesenchymal stem cell-mediated cardiac protection and functional improvement. FASEB J. 2006, 20, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Ratajczak, J. Horizontal transfer of RNA and proteins between cells by extracellular microvesicles: 14 years later. Clin. Transl. Med. 2016, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arslan, F.; Lai, R.C.; Smeets, M.B.; Akeroyd, L.; Choo, A.; Aguor, E.N.E.; Timmers, L.; van Rijen, H.V.; Doevendans, P.A.; Pasterkamp, G.; et al. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013, 10, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Xiang, M.; Meng, D.; Sun, N.; Chen, S. Inhibition of Myocardial Ischemia/Reperfusion Injury by Exosomes Secreted from Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Perez-polo, J.R.; Qian, J.; Birnbaum, Y.; Ye, Y.; Perez-polo, J.R.; Qian, J.; Birnbaum, Y. The role of microRNA in modulating myocardial ischemia-reperfusion injury The role of microRNA in modulating myocardial ischemia-reperfusion injury. 2013, 77555, 534–542. [Google Scholar]
- Qiao, S.; Olson, J.M.; Paterson, M.; Yan, Y.; Zaja, I.; Liu, Y.; Riess, M.L.; Kersten, J.R.; Liang, M.; Warltier, D.C.; et al. MicroRNA-21 Mediates Isoflurane-induced Cardioprotection against Ischemia–Reperfusion Injury via Akt/Nitric Oxide Synthase/Mitochondrial Permeability Transition Pore Pathway. Anesthesiol. J. Am. Soc. Anesthesiol. 2015, 123, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Liu, X.; Zhang, S.; Lin, Y.; Yang, J.; Zhang, C. MicroRNA-21 protects against the H2O2-induced injury on cardiac myocytes via its target gene PDCD4. J. Mol. Cell. Cardiol. 2009, 47, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, D.; He, M.; Hong, C.; Gao, S.; Rane, S.; Yang, Z.; Abdellatif, M. MicroRNA-21 is a downstream effector of AKT that mediates its antiapoptotic effects via suppression of fas ligand. J. Biol. Chem. 2010, 285, 20281–20290. [Google Scholar] [CrossRef] [PubMed]
- Jazbutyte, V.; Thum, T. MicroRNA-21: From cancer to cardiovascular disease. Curr. Drug Targets 2010, 11, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Zhu, H.; Zhu, C.; Wang, Y.; Pu, W.T.; Jegga, A.G.; Fan, G.C. Synergistic effects of the GATA-4-mediated miR-144/451 cluster in protection against simulated ischemia/reperfusion-induced cardiomyocyte death. J. Mol. Cell. Cardiol. 2010, 49, 841–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Rohailla, S.; Gelber, N.; Rutka, J.; Sabah, N.; Gladstone, R.A.; Wei, C.; Hu, P.; Kharbanda, R.K.; Redington, A.N. MicroRNA-144 is a circulating effector of remote ischemic preconditioning. Basic Res. Cardiol. 2014, 109, 423. [Google Scholar] [CrossRef] [PubMed]
- Lusha, E.; Jiang, H.; Lu, Z. MicroRNA-144 attenuates cardiac ischemia/reperfusion injury by targeting FOXO1. Exp. Ther. Med. 2019, 17, 2152–2160. [Google Scholar]
- Wang, X.; Zhu, H.; Zhang, X.; Liu, Y.; Chen, J.; Medvedovic, M.; Li, H.; Weiss, M.J.; Ren, X.; Fan, G.-C. Loss of the miR-144/451 cluster impairs ischaemic preconditioning-mediated cardioprotection by targeting Rac-1. Cardiovasc. Res. 2012, 94, 379–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anversa, P.; Kajstura, J. Ventricular myocytes are not terminally differentiated in the adult mammalian heart. Circ. Res. 1998, 83, 1–14. [Google Scholar] [CrossRef]
- Libby, W.F.; Berger, R.; Mead, J.F.; Alexander, G.V.; Ross, J.F. Replacement rates for human tissue from atmospheric radiocarbon. Science 1964, 146, 1170–1172. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Messina, E.; Giacomello, A.; Marbán, E. Endogenous Cardiac Stem Cells. Prog. Cardiovasc. Dis. 2007, 50, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Ye, Z.; Song, X.; Huang, R. Differences in the cargos and functions of exosomes derived from six cardiac cell types: A systematic review. Stem Cell Res. Ther. 2019, 10, 194. [Google Scholar] [CrossRef] [PubMed]
- Prathipati, P.; Nandi, S.S.; Mishra, P.K. Stem Cell-Derived Exosomes, Autophagy, Extracellular Matrix Turnover, and miRNAs in Cardiac Regeneration during Stem Cell Therapy. Stem Cell Rev. Reports 2017, 13, 79–91. [Google Scholar] [CrossRef]
- Shao, L.; Zhang, Y.; Pan, X.; Liu, B.; Liang, C.; Zhang, Y.; Wang, Y.; Yan, B.; Xie, W.; Sun, Y.; et al. Knockout of beta-2 microglobulin enhances cardiac repair by modulating exosome imprinting and inhibiting stem cell-induced immune rejection. Cell. Mol. Life Sci. 2019, 1, 3. [Google Scholar] [CrossRef]
- Wang, D.; Hu, X.; Lee, S.H.; Chen, F.; Jiang, K.; Tu, Z.; Liu, Z.; Du, J.; Wang, L.; Yin, C.; et al. Diabetes Exacerbates Myocardial Ischemia/Reperfusion Injury by Down-Regulation of MicroRNA and Up-Regulation of O-GlcNAcylation. JACC Basic Transl. Sci. 2018, 3, 350–362. [Google Scholar] [CrossRef]
- Hofmann, U.; Beyersdorf, N.; Weirather, J.; Podolskaya, A.; Bauersachs, J.; Ertl, G.; Kerkau, T.; Frantz, S. Activation of CD4 + T lymphocytes improves wound healing and survival after experimental myocardial infarction in mice. Circulation 2012, 125, 1652–1663. [Google Scholar] [CrossRef]
- Song, J.; Chen, X.; Wang, M.; Xing, Y.; Zheng, Z.; Hu, S. Cardiac endothelial cell-derived exosomes induce specific regulatory B cells. Sci. Rep. 2014, 4, 7583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, M.; Ibba-Manneschi, L.; Manetti, M. Cardiac telocyte-derived exosomes and their possible implications in cardiovascular pathophysiology. Adv. Exp. Med. Biol. 2017, 998, 237–254. [Google Scholar] [PubMed]
- Wang, J.; Jin, M.; Ma, W.H.; Zhu, Z.; Wang, X. The history of telocyte discovery and understanding. Adv. Exp. Med. Biol. 2016, 913, 1–21. [Google Scholar] [PubMed]
- Tao, L.; Wang, H.; Wang, X.; Kong, X.; Li, X. Cardiac Telocytes. Curr. Stem Cell Res. Ther. 2016, 11, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Zhu, H.; Li, S.; Wang, Y.; Bei, Y.; Xiao, J. Telocytes in Cardiac Protection. Curr. Stem Cell Res. Ther. 2016, 11, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Z.; Li, C.; Chen, Q.; Jing, Y.; Carpenter, R.; Jiang, Y.; Kung, H.F.; Lai, L.; Jiang, B.H. Mir-21 induced angiogenesis through AKT and ERK activation and HIF-1α expression. PLoS ONE 2011, 6, e19139. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Olson, E.N. A neonatal blueprint for cardiac regeneration. Stem Cell Res. 2014, 13, 556–570. [Google Scholar] [CrossRef] [Green Version]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef]
- Kojima, H.; Ieda, M. Discovery and progress of direct cardiac reprogramming. Cell. Mol. Life Sci. 2017, 74, 2203–2215. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Q. Stem/progenitor cells in vascular regeneration. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1114–1119. [Google Scholar] [CrossRef]
- Sherman, L.S.; Shaker, M.; Mariotti, V.; Rameshwar, P. Mesenchymal stromal/stem cells in drug therapy: New perspective. Cytotherapy 2017, 19, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Blin, G.; Nury, D.; Stefanovic, S.; Neri, T.; Guillevic, O.; Brinon, B.; Bellamy, V.; Rücker-Martin, C.; Barbry, P.; Bel, A.; et al. A purified population of multipotent cardiovascular progenitors derived from primate pluripotent stem cells engrafts in postmyocardial infarcted nonhuman primates. J. Clin. Invest. 2010, 120, 1125–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.D.; Johansson, H.J.; Graham, C.S.; Vesterlund, M.; Pham, M.T.; Bramlett, C.S.; Montgomery, E.N.; Mellema, M.S.; Bardini, R.L.; Contreras, Z.; et al. Comprehensive proteomic analysis of mesenchymal stem cell exosomes reveals modulation of angiogenesis via nuclear factor-kappaB signaling. Stem Cells 2016, 34, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Vrijsen, K.R.; Maring, J.A.; Chamuleau, S.A.J.; Verhage, V.; Mol, E.A.; Deddens, J.C.; Metz, C.H.G.; Lodder, K.; van Eeuwijk, E.C.M.; van Dommelen, S.M.; et al. Exosomes from Cardiomyocyte Progenitor Cells and Mesenchymal Stem Cells Stimulate Angiogenesis Via EMMPRIN. Adv. Healthc. Mater. 2016, 5, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.P.; Boon, R.A. Exosomes and non-coding RNA, the healers of the heart? Cardiovasc. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Johnson, T.; Liu, D. Therapeutic angiogenesis of adipose-derived stem cells for ischemic diseases. Stem Cell Res. Ther. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Van Balkom, B.W.M.; de Jong, O.G.; Smits, M.; Brummelman, J.; den Ouden, K.; de Bree, P.M.; van Eijndhoven, M.A.J.; Pegtel, D.M.; Stoorvogel, W.; Würdinger, T.; et al. Endothelial cells require miR-214 to secrete exosomes that suppress senescence and induce angiogenesis in human and mouse endothelial cells. Blood 2013, 121, 3997–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Naranjo, J.C.; Wu, H.-J.; Ugaz, V.M. Microfluidics for exosome isolation and analysis: Enabling liquid biopsy for personalized medicine. Lab Chip 2017, 17, 3558–3577. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.-G.; Kong, M.-Q.; Zhou, S.; Sheng, Y.-F.; Wang, P.; Yu, T.; Inci, F.; Kuo, W.P.; Li, L.-J.; Demirci, U.; et al. An integrated double-filtration microfluidic device for isolation, enrichment and quantification of urinary extracellular vesicles for detection of bladder cancer. Sci. Rep. 2017, 7, 46224. [Google Scholar] [CrossRef]
- Wu, M.; Ouyang, Y.; Wang, Z.; Zhang, R.; Huang, P.-H.; Chen, C.; Li, H.; Li, P.; Quinn, D.; Dao, M.; et al. Isolation of exosomes from whole blood by integrating acoustics and microfluidics. Proc. Natl. Acad. Sci. USA 2017, 114, 10584–10589. [Google Scholar] [CrossRef] [Green Version]
- Dragovic, R.A.; Gardiner, C.; Brooks, A.S.; Tannetta, D.S.; Ferguson, D.J.P.; Hole, P.; Carr, B.; Redman, C.W.G.; Harris, A.L.; Dobson, P.J.; et al. Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, L.C.P.; Janiszewski, M.; Pontieri, V.; Pedro, M.d.A.; Bassi, E.; Tucci, P.J.F.; Laurindo, F.R.M. Platelet-derived exosomes from septic shock patients induce myocardial dysfunction. Crit. Care 2007, 11. [Google Scholar] [CrossRef] [PubMed]
- Andriolo, G.; Provasi, E.; Lo Cicero, V.; Brambilla, A.; Soncin, S.; Torre, T.; Milano, G.; Biemmi, V.; Vassalli, G.; Turchetto, L.; et al. Exosomes from human cardiac progenitor cells for therapeutic applications: Development of a GMP-grade manufacturing method. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Pachler, K.; Lener, T.; Streif, D.; Dunai, Z.A.; Desgeorges, A.; Feichtner, M.; Öller, M.; Schallmoser, K.; Rohde, E.; Gimona, M. A Good Manufacturing Practice–grade standard protocol for exclusively human mesenchymal stromal cell–derived extracellular vesicles. Cytotherapy 2017, 19, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Whitford, W.; Guterstam, P. Exosome manufacturing status. Future Med. Chem. 2019, 11, 1225–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.S.; Arslan, F.; Yin, Y.; Tan, S.S.; Lai, R.C.; Choo, A.B.H.; Padmanabhan, J.; Lee, C.N.; de Kleijn, D.P.V.; Lim, S.K. Enabling a robust scalable manufacturing process for therapeutic exosomes through oncogenic immortalization of human ESC-derived MSCs. J. Transl. Med. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Nishikawa, M.; Shinotsuka, H.; Matsui, Y.; Ohara, S.; Imai, T.; Takakura, Y. Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection. J. Biotechnol. 2013, 165, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Takahashi, Y.; Nishikawa, M.; Kato, K.; Morishita, M.; Yamashita, T.; Matsumoto, A.; Charoenviriyakul, C.; Takakura, Y. Macrophage-dependent clearance of systemically administered B16BL6-derived exosomes from the blood circulation in mice. J. Extracell. Vesicles 2015, 4, 1–8. [Google Scholar] [CrossRef]
- Han, C.; Zhou, J.; Liang, C.; Liu, B.; Pan, X.; Zhang, Y.; Wang, Y.; Yan, B.; Xie, W.; Liu, F.; et al. Human umbilical cord mesenchymal stem cell derived exosomes encapsulated in functional peptide hydrogels promote cardiac repair. Biomater. Sci. 2019, 7, 2920–2933. [Google Scholar] [CrossRef]
- Vandergriff, A.; Huang, K.; Shen, D.; Hu, S.; Hensley, M.T.; Caranasos, T.G.; Qian, L.; Cheng, K. Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide. Theranostics 2018, 8, 1869–1878. [Google Scholar] [CrossRef]
- Kim, H.; Yun, N.; Mun, D.; Kang, J.Y.; Lee, S.H.; Park, H.; Park, H.; Joung, B. Cardiac-specific delivery by cardiac tissue-targeting peptide-expressing exosomes. Biochem. Biophys. Res. Commun. 2018, 499, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Y.; Zhao, Z.; Meng, Q.; Yu, Y.; Sun, J.; Yang, Z.; Chen, Y.; Li, J.; Ma, T.; et al. Engineered exosomes with ischemic myocardium-targeting peptide for targeted therapy in myocardial infarction. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.K.A.; Luciani, N.; Gazeau, F.; Aubertin, K.; Bonneau, S.; Chauvierre, C.; Letourneur, D.; Wilhelm, C. Combining magnetic nanoparticles with cell derived microvesicles for drug loading and targeting. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 645–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, J.; Allard-Vannier, E.; Munnier, E.; Soucé, M.; Chourpa, I. Recent advances in theranostic nanocarriers of doxorubicin based on iron oxide and gold nanoparticles. J. Control. Release 2013, 169, 48–61. [Google Scholar] [CrossRef] [PubMed]
- MacKie, A.R.; Klyachko, E.; Thorne, T.; Schultz, K.M.; Millay, M.; Ito, A.; Kamide, C.E.; Liu, T.; Gupta, R.; Sahoo, S.; et al. Sonic hedgehog-modified human CD34+ cells preserve cardiac function after acute myocardial infarction. Circ. Res. 2012, 111, 312–321. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Chen, W.; Xie, L.; Zhao, Z.A.; Yang, J.; Chen, Y.; Lei, W.; Shen, Z. MicroRNA-133 overexpression promotes the therapeutic efficacy of mesenchymal stem cells on acute myocardial infarction. Stem Cell Res. Ther. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobor, F.; Dallmann, A.; Ball, N.J.; Cicchini, C.; Battistelli, C.; Ogrodowicz, R.W.; Christodoulou, E.; Martin, S.R.; Castello, A.; Tripodi, M.; et al. A cryptic RNA-binding domain mediates Syncrip recognition and exosomal partitioning of miRNA targets. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Pomatto, M.A.C.; Bussolati, B.; D’Antico, S.; Ghiotto, S.; Tetta, C.; Brizzi, M.F.; Camussi, G. Improved Loading of Plasma-Derived Extracellular Vesicles to Encapsulate Antitumor miRNAs. Mol. Ther. Methods Clin. Dev. 2019, 13, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Spannbauer, A.; Traxler, D.; Lukovic, D.; Zlabinger, K.; Winkler, J.; Gugerell, A.; Ferdinandy, P.; Hausenloy, D.J.; Pavo, N.; Emmert, M.Y.; et al. Effect of ischemic preconditioning and postconditioning on exosome-rich fraction microrna levels, in relation with electrophysiological parameters and ventricular arrhythmia in experimental closed-chest reperfused myocardial infarction. Int. J. Mol. Sci. 2019, 20, 2140. [Google Scholar] [CrossRef]

{kind=link}
| Study, Year | Population (N) | Design | Treatment | Primary Endpoints | Results | Refs |
|---|---|---|---|---|---|---|
| ISIS 2, 1988 | Suspected acute MI (17187) | Multicenter, double-blinded, two-by-two factorial, placebo-controlled, randomized trial | Streptokinase vs. 1-month ASA vs. both vs. neither | Vascular mortality at 5 weeks, non-fatal reinfarction, bleeds requiring transfusion, non-fatal stroke, and cerebral hemorrhage. | The co-administration of streptokinase and ASA reduced vascular mortality compared to single drug treatment (40% vs. 23% vs. 20%). | [91] |
| Cure, 2001 | UA/NSTEMI (12562) | Multicenter, double-blind, parallel group, placebo-controlled, randomized trial | ASA + clopidogrel vs. ASA + placebo | Composite of CV mortality, non-fatal MI, or stroke. | Dual antiplatelet therapy reduced CV mortality, non-fatal MI, or stroke but increased the rate of major bleeding | [92] |
| TRITON-TIMI 38, 2007 | ACS (13608) | Multicenter, double-blind, randomized trial | ASA + prasugrel vs. ASA + clopidogrel | CV mortality, non-fatal MI, or non-fatal cerebrovascular events. | Prasugrel reduced CV morbidity and mortality but increases bleeding compared to clopidogrel | [93] |
| PLATO, 2009 | ACS (18624) | Multicenter, double-blind, randomized trial | ASA + ticagrelor vs. ASA + clopidogrel | Vascular mortality, MI, or cerebrovascular events, major bleeding. | Ticagrelor reduced the rate of CV death, MI, or stroke without increasing the rate of overall major bleeding | [94] |
| METOCARD-CNIC, 2014 | Anterior STEMI undergoing PCI (270) | Randomized trial | Metoprolol iv | Infarct size at 5–7 days (underpowered). | Beta-blocker was associated with a smaller infarct size compared with control; improved LVEF at 6 months | [95] |
| VCU-ART, 2010 | STEMI (10) | Double-blind, placebo controlled, randomized trial | Anakinra vs. placebo | Change in LVESVi at CMR and echocardiography at 3 months. | Anakinra decreased LVESVi and LVEDVi | [96] |
| MRC-ILA Heart Study, 2014 | NSTEMI (182) | Double-blind placebo-controlled, randomized trial | Anakinra vs. placebo | AUC for CRP over the first 7 days. | Anakinra reduced CRP levels, but increased the incidence of CV events at 12 months | [97] |
| CANTOS, 2012 | Post-MI and elevated CRP (10061) | Double-blind, multi-center, placebo-controlled, randomized trial | Canakinumab (50, 150 or 300 mg) | Composite of nonfatal MI, nonfatal stroke, or cardiovascular death. | Canakinumab 150 mg reduced the composite outcome mainly reducing non-fatal MI; reduction in lung cancer, but associated with higher risk of fatal infections | [98] |
| AMISTAD-II, 2005 | STEMI (2118) | Double-blind, multi-center, placebo-controlled, randomized trial | Adenosine infusion vs placebo for 3 h before PPCI/fibrinolysis. | New congestive heart failure beginning >24 h after randomization, or the first re-hospitalization for CHF, or death from any causes within six months. | No difference between placebo and adenosine. Adenosine dose-response relationship in decreasing median infarct size. | [99] |
| AMISTAD-II post-hoc analysis, 2006 | STEMI (2118) | Double-blind, multi-center, placebo-controlled, randomized trial | Adenosine infusion vs placebo for 3 h before PCI/fibrinolysis. | New congestive HF beginning >24 h, or the first re-hospitalization for CHF, or death from any causes within six months. Endpoint analyzed according to time of reperfusion therapy. | Adenosine (<3.17 h) reduced mortality at both 1 and 6 months as well as the primary clinical endpoint at 6 months, with no distinction between adenosine dose regimens. | [100] |
| David Garcia-Dorado et al., 2014 | STEMI (201) | Double-blind, placebo-controlled, randomized trial | Intracoronary infusion of 4.5 mg Adenosine vs saline immediately prior to reperfusion | Percentage of total myocardial necrotic mass assessed by CMR at 2–7 days post-reperfusion. | Intracoronary Adenosine administration prior to PCI did not limit infarct size. | [101] |
| Desmet et al., 2011 | STEMI (112) | Prospective, double-blind, placebo-controlled clinical study | Intracoronary infusion 4 mg of Adenosine or matching placebo distal to the coronary occlusion site immediately before initial balloon inflation | Myocardial salvage defined as the percentage of the area at risk (AAR), which was not necrotic on CMR at day 2 and 3. | No evidence of changes in myocardial salvage. | [102] |
| NIAMI, 2014 | STEMI (229) | Double-blind, multi-center, placebo-controlled, randomized trial | IV administration of 70 mmol sodium nitrite or matching placebo over 5 min immediately before PPCI | Difference in percentage of LV myocardial mass between active and placebo at 6–8 days post-infarct assessed by CMR. | No reduction in infarct size | [103] |
| Bøtker et al. 2010 | STEMI (251) | Prospective, single-center randomized controlled trial | rIPC (intermittent arm ischemia through four cycles of 5-min. of inflation and deflation of a blood-pressure cuff) vs nothing before PPCI. | Myocardial salvage index at day 30 after primary percutaneous coronary intervention, estimated by G-SPECT. | rIPC before hospital admission increases myocardial salvage. | [104] |
| CONDI-2/ERIC-PPCI 2019 | STEMI (5401) | Single-blind, multi-center randomized controlled trial | rIPC (intermittent arm ischemia through four cycles of 5-min. of inflation and deflation of a blood-pressure cuff) vs. nothing before PPCI. | Cardiac death or hospitalisation for heart failure at 12 months | rIPC does not improve clinical outcomes | [104] |
| Staat et al., 2005 | STEMI (30) | Prospective, multi-center, randomized, open-label, con- trolled study | Post-conditioning after PPCI performed within 1 min of reflow by 4 cycles of 1 min. inflation and deflation of the angioplasty balloon | Infarct assessed by measuring total creatinine kinase release over 72 h. | Post-conditioning reduced infarct size. | [105] |
| Thibault et al., 2008 | STEMI (38) | Prospective randomized controlled trial | Post-conditioning after PPCI performed within 1 min of reflow by 4 cycles of 1 min. inflation and deflation of the angioplasty balloon. | Persistent infarct size reduction, assessed by SPECT imaging with rest-redistribution index at 6 months. | Post-conditioning affords persistent infarct size reduction | [106] |
| POST, 2013 | STEMI (700) | Multi-center, randomized, open-label, blinded trial | Post-conditioning after PPCI performed within 1 min of reflow by 4 cycles of 1 min. inflation and deflation of the angioplasty balloon. | Complete ST-segment resolution (percentage resolution of ST-segment elevation >70%) measured at 30 min after PCI | Post-conditioning did not improve myocardial reperfusion in STEMI patients | [107] |
| POST substudy, 2015 | STEMI (111) | Multi-center, randomized, open-label, blinded trial | Post-conditioning after PPCI performed within 1 min of reflow by 4 cycles of 1 min. inflation and deflation of the angioplasty balloon. | Myocardial salvage measured by CMR at day 3 after the index event. | Myocardial salvage index was not improved. | [108] |
| DANAMI-3–iPOST, 2017 | STEMI (1234) | Multi-center, randomized clinical trial | Conventional PPCI vs post-conditioning performed as 4 cycles of 30-s balloon occlusions and reperfusion after opening of the infarct-related artery and before stent implantation. | A combination of all-causes of death and hospitalization for heart failure at follow-up. | Post-conditioning during PPCI failed to improve clinical outcomes. | [109] |
| CRISP-AMI, 2011 | STEMI (337) | Multi-center, randomized clinical trial | Initiation of IABP before PPCI and continuation for at least 12 h (IABP plus PPCI) vs PPCI alone. | Infarct size expressed as a percentage of LV mass measured by CMR 3 to 5 days after PPCI. | IABP plus PPCI compared with PCI alone did not result in reduced infarct size. | [110] |
| CRISP-AMI substudy, 2015 | STEMI (36) | Retrospective analysis | PPCI + IABP vs PPCI alone in large myocardial infarction and poor ST segment resolution | All-causes of mortality at six months, and composite endpoint of death, cardiogenic shock and new or worsening HF at six months. | IABP associated with decreased six-month mortality in large STEMI complicated by persistent ischemia after PPCI | [111] |
| OxAMI-PICSO, 2018 | STEMI (105) | Single-center, investigator-initiated study, prospective study | PICSO in patients with index of microcirculatory resistance >40 compared to historical cohort of controls. | Infarct size assessment within 48 h after PPCI and at six months. | IMR-guided treatment with PICSO may be associated with reduced infarct size | [112] |
| Source | Animal Model | Administration | Effects | Mechanisms | Refs | |
|---|---|---|---|---|---|---|
| In Vitro | In Vivo | |||||
| Platelet | Rat MI | Intramyocardial | Angiogenesis EC proliferation | Angiogenesis | VEGF-PDGF—bFGF RISK pathway | [152] |
| Platelet from rat undergoing rIPC | Rat I/R | Intravenous | - | Improved cardiac function | Increase of MV circulating in periferial blood. Undefined | [156] |
| EV from coronary blood after rIPC | Langendorf—mode isolated heart | Intracoronary | - | Decrease of infarct size | Undefined | [157] |
| MV isolated after IPC in periferial rat blood | Rat LAD ligation | Intravenous | - | Decrease of infarct size Reduced cardiomyocyte apoptosis | Decrease of caspase-3 and -12 activity Reduced endoplasmic reticulum stress | [158] |
| MV isolated after IRC in periferial rat blood | Rat coronary ligation | Intravenous | Failure to decrease infarct size compared to MI alone without MV | [159] | ||
| MV isolated from HUVEC after H/R | H9c2 cardiomyocytes | Incubation in vitro | Increased apoptosis | - | Higher level of ROS and lipid peroxidation Bcl-2 inhibition | [160] |
| Cardiac MV isolated from cardiac ischemic tissue | Rat coronary ligation | Incubation in vitro of MV with Ly6+ monocyte | Modulation of inlammation | . | Increased release of Il6 and CCL2 and CCL7 | [161] |
| MV collected from MSC overxpressing GATA-4 | Cardiomyocyte after H/R | Incubation in vitro | Reduced apoptosis | - | miR-221 overexpression modulation of PUMA | [162] |
| Donor Cells | Animal Model | Administration | Effects | Mechanisms | Refs | |
|---|---|---|---|---|---|---|
| In Vitro | In Vivo | |||||
| Mesenchymal stem cell (MSC) | ||||||
| MSC-conditioned medium (MSC-CM) | Mouse I/R | Intravenous | Undefined | Reduction of infarct size | Undefined | [169] |
| MSCs following ischemic preconditioning (EXOIPC) | C57BL/6J mouse LAD ligation | Intramyocardial | Anti-apoptosis | Reduction of cardiac fibrosis | miR-22 targets methyl CpG binding protein 2 (Mecp2) | [170] |
| MSC overexpressing GATA-4 (ExoGATA-4) | Mouse LAD ligation | Intramyocardial | Increase of CM survival, reduction of CM apoptosis and preservation of mitochondrial membrane potential | Recovery of contractile function reduction of the infarct size | Anti-apoptotic miRs (e.g., miR-19a), by reducing PTEN expression drive the activation of the Akt-ERK signalling pathway | [171] |
| Endometrium-derived mesenchymal stem cells (EnMSCs) | Mouse MI | Intramyocardial | Anti-apoptosis Angiogenesis | Anti-apoptotic effects Angiogenesis myocardial salvage and improvement of cardiac function | mir-21, PTEN, Akt pathway | [172] |
| MSCs | Rat I/R | Intramyocardial | Anti-apoptosis Autophagy | Increase sautophagy, reduction of apoptosis and myocardial infarct size | AMPK/mTOR and Akt/mTOR pathway | [173] |
| Transplanted MSCs | Mouse MI | Transplantation | Autophagy reduction | Autophagy reduction | miR-125b modulates p53-Bnip3 signalling | [174] |
| Human-derived MSCs | Isolated rat heart I/R | Intramyocardial | Autophagy and apoptosis inhibition | Cardiac function recovery | BCL2 up-regulation | [175] |
| Adipose-derived MSCs (ADMSCs) | Mouse I/R | Intramyocardial | Anti-apoptosis | Reduction of infarct size | Wnt/β-catenin signaling pathway | [176] |
| B2M deletion-human Umbilical Cord Mesenchymal Stem Cells (B2M-UMSC) | Rat MI | Intramyocardial | Undefined | Cardiac fibrosis inhibition, cardiac function recovery | mir-24/Bim pathway | [22] |
| MSCs | Mouse I/R | Intramyocardial | Anti-inflammation Treg polarization | Anti-inflammation, delayed ischemic damage | mir-181a (lentiviruses), c-Fos inhibition | [177] |
| Atorvastatine-pretreated MSCs (MSCATV-Exo) | Mouse MI | Intramyocardial | Angiogenesis Anti-apoptosis | Cardiac function improvement, infarct size reduction, anti-apoptotic effects, angiogenesis and anti-inflammation | lncRNA H19 regulation of miR-675, activation of VEGF and ICAM-1 | [178] |
| MSC transduced with lentiviral CXCR4 | Rat MI | Intramyocardial | Anti-apoptosis Angiogenesis | Angiogenesis, infarct size reduction, improvement of cardiac remodelling | IGF-1α and pAkt up-regulation, active caspase 3 downregulation, VEGF enhancement | [179] |
| MSCs | Mouse LAD ligation | Intramyocardial | Angiogenesis | Angiogenesis, heart function preservation | miR-132, RASA1 gene | [180] |
| Cardiac stem cells (CSCs) preconditioned with MSC-EXO | Mouse LAD ligation | Intramyocardial | Proliferation, migration, and tube formation of c-kit+ CSCs | Angiogenesis, reduction of fibrosis, LV function recovery | Upregulation of miR-147, let-7i-3p, miR-503-5p, and miR-362-3p | [181] |
| Cardiac-derived progenitor cell (CPC) | ||||||
| CPCs | Mouse MI/R | Intramyocardial | Anti-H2O2 induced apoptosis | Anti-apoptotic effects | miR-451 | [182] |
| CDC-conditioned medium (CDC-CM) | Mouse MI | Intramyocardial | Angiogenesis, anti-apoptotic effects and proliferation | Reduction of the scar mass, improvement of cardiac function | miR-146a, suppression of Irak1 and Traf6 (TLR pathway), NOX-4 and SMAD4 (TGF-β pathway) | [183] |
| Human derived-CPCs | Mouse LAD ligation | Intramyocardial | Anti-apoptotic effects and angiogenesis | Reduction of the scar mass, angiogenesis, improvement of cardiac function | miR-210 -> down-regulation of ephrin A3 and PTP1b miR-132 -> down-regulation of RasGAP-p120 miR-146a-3p | [184] |
| Human derived CPCs and bone marrow-derived mesenchymal stem/progenitor cells (BMCs) | Rat MI and I/R | Intramyocardial | Anti-apoptotic effects (CPCs > BMCs) | Reduction of the scar size, improvement of LVEF (CPCs> BMCs) in I/R model (CPCs only) Angiogenesis | PAPP-A (Exo-CPC), IGF-1 release, activation of the Akt-ERK signaling pathway | [185] |
| CDCs | Mouse I/R | Intracoronary | Protection against oxidative stress | Reduction of infarct size | Y RNA fragment (EV-YF1) induces IL-10 secretion | [186] |
| Human-derived CDCs | Pig Acute and chronic MI | Intracoronary Intramyocardial | Undefined | Reduction of infarct size (Acute MI) Reduction of the scar size (Chronic MI) | Alteration of pro-inflammatory and pro-fibrotic pathway | [187] |
| Mouse-derived CPCs | Mouse CMs Oxidative stress | Undefined | Anti-apoptotic effects | Undefined | miR-21 downregulates PDCD4, inhibition of caspase 3/7-mediated apoptosis | [188] |
| Mouse-derived CPCs | Mouse LAD ligation | Intravenous | Angiogenesis | Angiogenesis Infarct size reduction | miR-322 (transfection), Nox2-dependent H2O2 production | [189] |
| CXCR4-overexpressing CPC (ExoCXCR4) | Rat I/R | Intravenous | Anti-apoptotic effects | Infarct size reduction, LV function improvement | Increased cardiac homing | [190] |
| Embryonic Stem Cell (ESC) | ||||||
| Mouse-derived ESCs | Mouse MI | Intramyocardial | CPC survival, proliferation, and cardiac commitment | Neovascularization, cardiomyocyte survival, reduction of fibrosis. CPC survival, proliferation, and cardiac commitment | miR-294, induced expression of cyclins (E1, A2, and D1) | [191] |
| Adipose-derived stem cell (ADSC) | ||||||
| ADSCs | Mouse MI | Intramyocardial | Reduction of autophagy, apoptosis and inflammatory response | Reduction of autophagy | miR-93-5p-mediated suppression of hypoxia-induced autophagy and inflammatory cytokine expression by targeting Atg7 and Toll-like receptor 4 (TLR4) | [192] |
| miR-146a-modified ADSCs | Mouse LAD ligation | Intravenous | Anti-apoptotic anti-inflammatory, and anti-fibrotic effects | Anti-apoptotic, anti-inflammatory, and anti-fibrotic effects | Downregulation of EGR1 | [193] |
| miR-126-overexpressing ADSCs | Mouse LAD ligation | Intravenous | Anti-inflammatory, anti-fibrotic, angiogenesis | Reduction of infarct size and cardiac fibrosis, angiogenesis | Spred1, PI3KR2/VEGF signalling pathway | [194] |
| Cardiomyocyte (CM) | ||||||
| AMI patients CMs | H9C2 cardiomyoblasts | Undefined | Autophagy | Undefined | Inhibition of miR-30a or release of Exo increased expression of the core autophagy regulators beclin-1, Atg12, and LC3II/LC3I | [195] |
| Cardiac telocyte (CT) | ||||||
| Mouse-derived CTs | Mouse LAD ligation | Intramyocardial | Undefined | Infarct size reduction, Cardiac function improvement Angiogenesis | CTs and endothelial cell contact VEGF and NOS2 secretion Various miRNA | [196,197] |
| Dendritic cell (DC) | ||||||
| Murine cultured bone marrow derived DCs (BMDCs) | Mouse MI | Intravenous | Activation of CD4(+) T cells | Improvement of cardiac function | Increased expression of chemokines and cytokines (IFN-γ and TNF) | [198] |
| Plasma | ||||||
| Human coronary serum from ischemic patients | Mouse Limb ischemia | Intramuscular | Endothelial cell proliferation, migration and tube formation | Angiogenesis | miR-939-iNOS-NO pathway | [199] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caccioppo, A.; Franchin, L.; Grosso, A.; Angelini, F.; D’Ascenzo, F.; Brizzi, M.F. Ischemia Reperfusion Injury: Mechanisms of Damage/Protection and Novel Strategies for Cardiac Recovery/Regeneration. Int. J. Mol. Sci. 2019, 20, 5024. https://doi.org/10.3390/ijms20205024
Caccioppo A, Franchin L, Grosso A, Angelini F, D’Ascenzo F, Brizzi MF. Ischemia Reperfusion Injury: Mechanisms of Damage/Protection and Novel Strategies for Cardiac Recovery/Regeneration. International Journal of Molecular Sciences. 2019; 20(20):5024. https://doi.org/10.3390/ijms20205024
Chicago/Turabian StyleCaccioppo, Andrea, Luca Franchin, Alberto Grosso, Filippo Angelini, Fabrizio D’Ascenzo, and Maria Felice Brizzi. 2019. "Ischemia Reperfusion Injury: Mechanisms of Damage/Protection and Novel Strategies for Cardiac Recovery/Regeneration" International Journal of Molecular Sciences 20, no. 20: 5024. https://doi.org/10.3390/ijms20205024