
Early Detection of Invasive Species on Roadside Slopes in An-Dean Patagonian Forests of Austrocedrus chilensis (Argentina) †

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
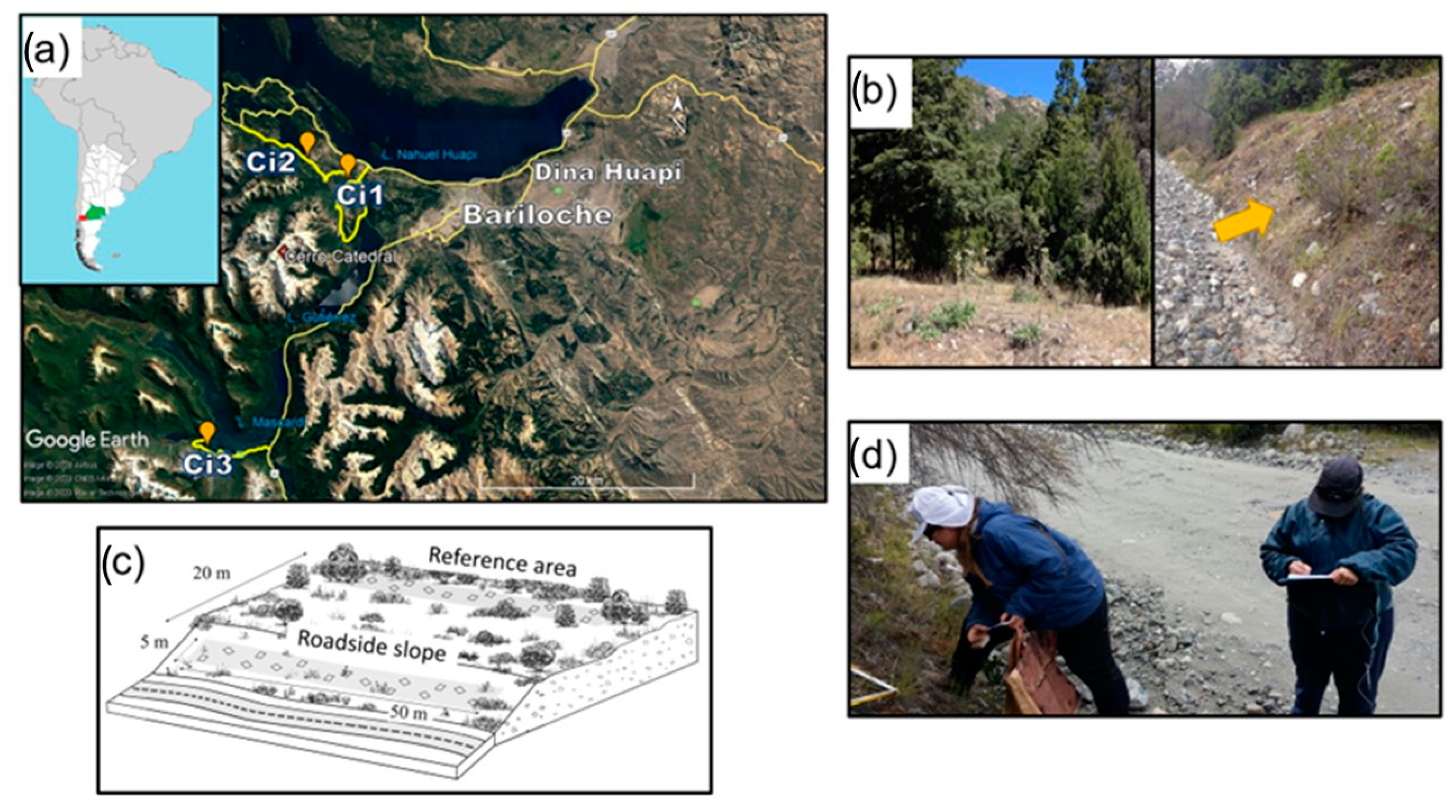
2. Materials and Methods
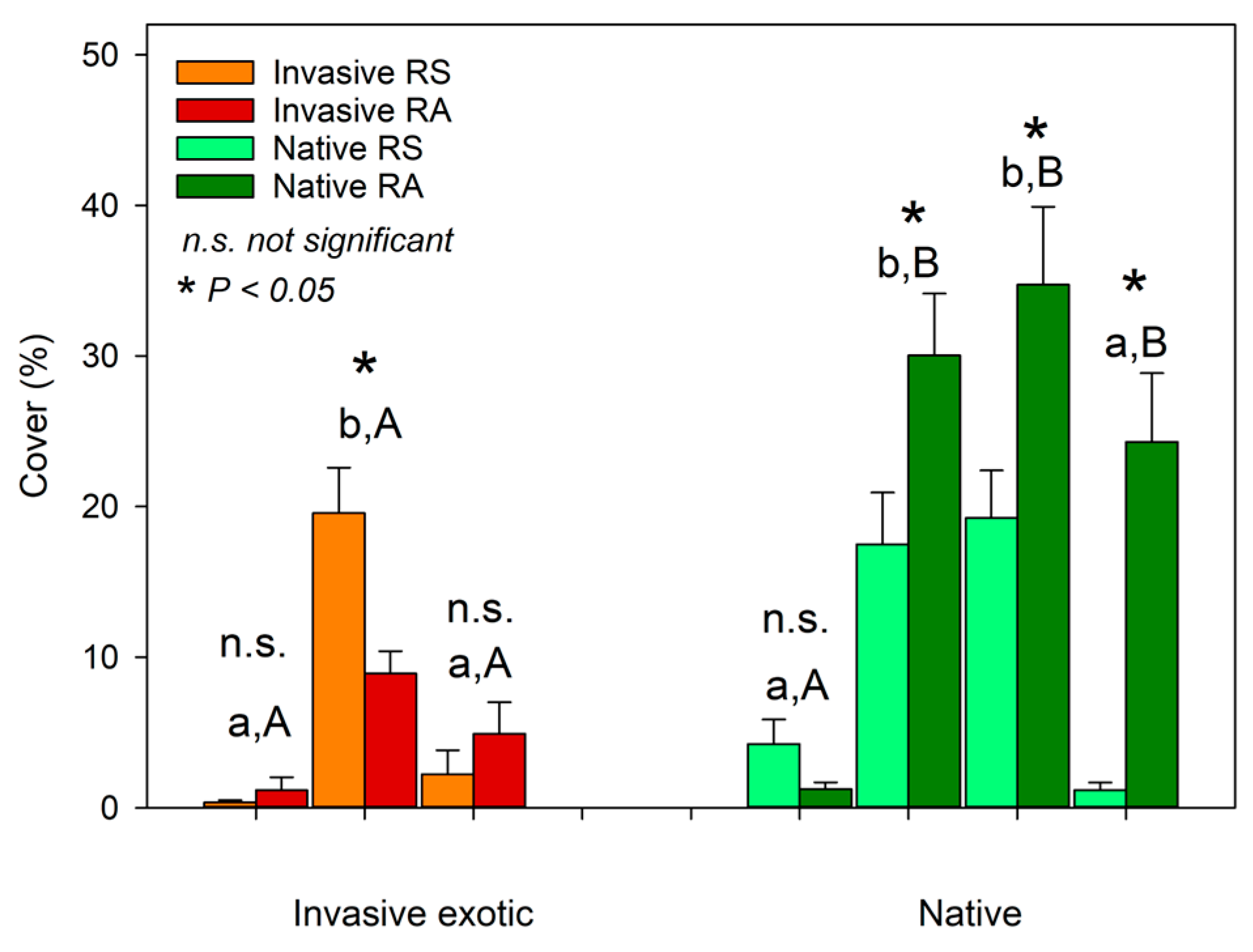
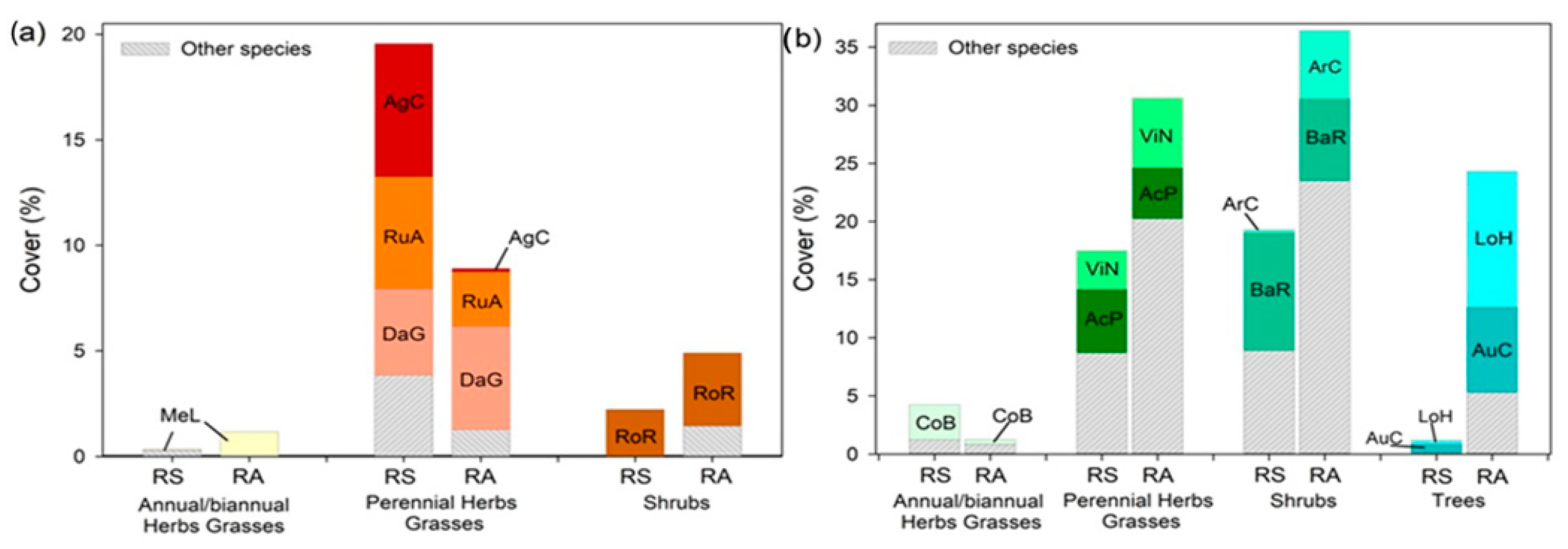
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDougall, K.L.; Lembrechts, J.; Rew, L.J.; Haider, S.; Cavieres, L.A.; Kueffer, C.; Milbau, A.; Naylor, B.J.; Nuñez, M.A.; Pauchard, A.; et al. Running off the Road: Roadside Non-Native Plants Invading Mountain Vegetation. Biol. Invasions 2018, 20, 3461–3473. [Google Scholar] [CrossRef]
- Kotowska, D.; Skórka, P.; Pärt, T.; Auffret, A.G.; Żmihorski, M. Spatial Scale Matters for Predicting Plant Invasions Along Roads. J. Ecol. 2024, 112, 305–318. [Google Scholar] [CrossRef]
- Jauni, M.; Huusela, E.; Urho, L.; Hyvönen, T. Prioritizing Management of Invasive Alien Species of EU Concern—A Northern Perspective. Diversity 2023, 15, 585. [Google Scholar] [CrossRef]
- Girmay, M.; Gebrehiwot, K.; Atinafe, E.; Tareke, Y.; Belay, B. The Study of Exotic and Invasive Plant Species in Gullele Botanic Garden, Addis Ababa, Ethiopia. J. Zool. Bot. Gard. 2024, 5, 36–50. [Google Scholar] [CrossRef]
- Jones, S.A.; DeKeyser, E.S.; Dixon, C.; Kobiela, B. Invasive Species Change Plant Community Composition of Preserved Prairie Pothole Wetlands. Plants 2023, 12, 1281. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Balaguer, L.; Mola, I.; Escudero, A.; Alfaya, V. Restauración Ecológica de Áreas Afectadas por Infraestructuras de Transporte. Base Científica para Soluciones Técnicas; Fundación Biodiversidad: Madrid, Spain, 2011; pp. 14–41. [Google Scholar]
- Bochet, E.; García-Fayos, P. Non-hazardous Seed Transport by Runoff on Slopes Determines Composition and Ecological Strategies of Plants in Patches and Interpatches in Drylands: A Trait-Based Approach. Catena 2024, 246, 108364. [Google Scholar] [CrossRef]
- Mola, I.; Jiménez, M.D.; López-Jiménez, N.; Casado, M.A.; Balaguer, L. Roadside Reclamation Outside the Revegetation Season: Management Options under Schedule Pressure. Restor. Ecol. 2011, 19, 83–92. [Google Scholar] [CrossRef]
- Chichizola, G.A.; Gonzalez, S.L.; Rovere, A.E. Alien Plant Species on Roadsides of the Northwestern Patagonian Steppe (Argentina). PLoS ONE 2021, 1, e0246657. [Google Scholar] [CrossRef]
- Chichizola, G.A.; Gonzalez, S.L.; Rovere, A.E. Efectos de los Bordes de Caminos y las Plantas Invasoras Sobre la Vegetación en Pie y el Banco de Semillas en Matorrales Norpatagónicos. Ecol. Austral 2023, 33, 923–937. [Google Scholar] [CrossRef]
- Lenhart, C.; Smiley, P.C. Ecological Restoration in the Midwest: Past, Present, and Future; University of Iowa Press: Iowa, IA, USA, 2018; pp. 11–207. [Google Scholar]
- Gonzalez, S.L.; Ghermandi, L. Overgrazing Causes a Reduction in the Vegetation Cover and Seed Bank of Patagonian Grasslands. Plant Soil 2021, 464, 75–87. [Google Scholar] [CrossRef]
- Šerá, B. Road-side Herbaceous Vegetation: Life History Groups and habitat preferences. Pol. J. Ecol. 2010, 58, 69–79. [Google Scholar]
- Wang, Y.J.; Liu, Y.Y.; Chen, D.; Du, D.L.; Müller-Schärer, H.; Yu, F.H. Clonal Functional Traits Favor the Invasive Success of Alien Plants into Native Comunities. Ecol. Appl. 2022, 34, e2756. [Google Scholar] [CrossRef] [PubMed]
- Iseli, E.; Chisholm, C.; Lenoir, J.; Haider, S.; Seipel, T.; Barros, A.; Hargreaves, A.L.; Kardol, P.; Lembrechts, J.J.; McDougall, K.; et al. Rapid Upwards Spread of Non-Native Plants in Mountains Across Continents. Nat. Ecol. Evol. 2023, 7, 405–413. [Google Scholar] [CrossRef]
- García-Mozo, H. Assessment of the Diversity, Abundance and Range of Invasive Alien Plant Species in Córdoba, a Mediterranean Urban Area. Diversity 2024, 16, 777. [Google Scholar] [CrossRef]
- Guo, Q.; Brockway, D.G.; Larson, D.L.; Wang, D.; Ren, H. Improving ecological restoration to curb biotic invasion—A practical guide. Invasive Plant Sci. Manag. 2018, 11, 163–174. [Google Scholar] [CrossRef]
- Ezcurra, C.; Brion, C. Plantas del Nahuel Huapi: Catálogo de la Flora Vascular del Parque Nacional Nahuel Huapi, Argentina, 1st ed.; Universidad Nacional del Comahue y Red Latinoamericana de Botánica: Bariloche, Argentina, 2005; pp. 1–64. [Google Scholar]
- Speziale, K.L.; Ezcurra, C. Patterns of alien plant invasions in northwestern Patagonia, Argentina. J. Arid. Environ. 2011, 75, 890–897. [Google Scholar] [CrossRef]
- Blackhall, M.; Raffaele, E. Flammability of Patagonian Invaders and Natives: When Exotic Plant Species Affect Live Fine Fuel Ignitability in Wildland-Urban Interfaces. Landsc. Urban. Plan. 2019, 189, 1–10. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Altamirano, A.; Pauchard, A.; Meli, P. Proximity to Forest Plantations is Associated with Presence and Abundance of Invasive Plants in landscapes of South-Central Chile. NeoBiota 2024, 92, 129–153. [Google Scholar] [CrossRef]
- Amoroso, M.M.; Peri, P.L.; Lencinas, M.V.; Soler, E.; Rovere, A.E.; González Peñalba, M.; Chauchard, L.; Urretavizcaya, M.F.; Loguercio, G.; Mundo, I.A.; et al. Región patagónica (Bosques Andino Patagónicos). In Uso Sostenible del Bosque: Aportes Desde la Silvicultura Argentina, 1st ed.; Peri, P.L., Martínez-Pastur, G., Schlichter, T., Eds.; Ministro de Ambiente y Desarrollo Sostenible de la Nación (MAyDS): Buenos Aires, Argentina, 2021; pp. 46–78. [Google Scholar]
- Newton, A.C. Forest Ecology and Conservation. A Handbook of Techniques; Oxford University Press: Oxford, UK, 2007; pp. 1–235. [Google Scholar]
- Zuloaga, F.O.; Morrone, O.; Belgrano, M.J. Catálogo de las Plantas Vasculares del Cono Sur: (Argentina, Sur de Brasil, Chile, Paraguay y Uruguay); Missouri Botanical Garden Press: St. Louis, MO, USA, 2008; Volume 107. [Google Scholar]
- MAyDS. Resolución 109/21 Lista de Especies Exóticas Invasoras, Potencialmente Invasoras y Criptogénicas; Anexos I y II; Ministerio de Ambiente y Desarrollo Sostenible: Buenos Aires, Argentina, 2021. Available online: https://www.boletinoficial.gob.ar/detalleAviso/primera/242964/20210414 (accessed on 24 September 2024).
- Gonzalez, S.L.; Ghermandi, L. Postfire Seed Bank Dynamics in Semiarid Grasslands. Plant Ecol. 2008, 199, 175–185. [Google Scholar] [CrossRef]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Bolker, B. Getting Started with the glmmTMB Package; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Stroup, W.W. Generalized Linear Mixed Models: Modern Concepts, Methods and Applications; CRC Press: Boca Raton, FL, USA, 2012; pp. 203–493. [Google Scholar]
- Deen, M.; de Rooij, M. ClusterBootstrap: An R package for the analysis of hierarchical data Using Generalized Linear Models with The Cluster Bootstrap. Behav. Res. 2020, 52, 572–590. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 30 May 2024).
- Paiaro, V.; Cabido, M.; Pucheta, E. Altitudinal Distribution of Native and Alien Plant Species in Roadside Communities from Central Argentina. Austral Ecol. 2011, 36, 176–184. [Google Scholar] [CrossRef]
- Karki, D.; Pandeya, B.; Ghimire, B. Diversity and Distribution of Invasive Alien Plant Species Along Elevation Gradient in Makawanpur District, Central Nepal. J. Ecol. Environ. 2023, 47, 8. [Google Scholar] [CrossRef]
- Gioria, M.; Hulme, P.E.; Richardson, D.M.; Pyšek, P. Why Are Invasive Plants Successful? Annu. Rev. Plant Biol. 2023, 74, 635–670. [Google Scholar] [CrossRef] [PubMed]
- Chichizola, G.A.; Gonzalez, S.L.; Rovere, A.E. Rasgos Eco-Morfológicos de Especies Nativas de la Estepa Como Criterio de Selección en Prácticas de Restauración. In Restauración Ecológica en la Diagonal Árida de la Argentina 5, 1st ed.; Del Ponti, O., Galmes, M., Sosa, A., Morici, E., Eds.; Facultad de Ciencias Exactas y Naturales, UNLPam: Santa Rosa, Argentina, 2024; pp. 112–119. [Google Scholar]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Chiuffo, M.C.; Cock, M.C.; Prina, A.O.; Hierro, J.L. Response of Native and Non-Native Ruderals to Natural and Human Disturbance. Biol. Invasions 2018, 20, 2915–2925. [Google Scholar] [CrossRef]
- Franzese, J.; Ghermandi, L. El Grado de Invasión de Rumex acetosella L. (Polygonaceae) y su Relación con los Atributos de la Vegetación de dos Comunidades de Pastizal en el NO de la Patagonia. Ecol. Austral 2012, 22, 101–111. [Google Scholar]
- Zuloaga, F.O.; Rugolo, Z.E.; Anton, A.M. Poaceae. Tomo II. In Flora Vascular de la República Argentina, 1st ed.; Anton, A.M., Zuloaga, F.O., Eds.; Gráficamente Ediciones: Córdoba, Argentina, 2012; Volume 3. [Google Scholar]
- Chichizola, G.A.; Gonzalez, S.L.; Rovere, A.E. Estudios ecológicos para la implementación de prácticas de la restauración en ambientes de estepa. In Restauración Ecológica en la Diagonal Árida de la Argentina 5, 1st ed.; Del Ponti, O., Galmes, M., Sosa, A., Morici, E., Eds.; Facultad de Ciencias Exactas y Naturales, UNLPam: Santa Rosa, Argentina, 2024; pp. 103–111. [Google Scholar]
- Clements, D.R.; Kato-Noguchi, H. Defensive Mechanisms of Mikania micrantha Likely Enhance its Invasiveness as One of the World’s Worst Alien Species. Plants 2025, 14, 269. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Pauchard, A.; Lenoir, J.; Nuñez, M.A.; Geron, C.; Ven, A.; Bravo-Monasterio, P.; Teneb, E.; Nijs, I.; Milbau, A. Disturbance is the Key to Plant Invasions in Cold Environments. Proc. Natl. Acad. Sci. USA 2016, 113, 14061–14066. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chichizola, G.A.; Gonzalez, S.L.; Rovere, A.E. Early Detection of Invasive Species on Roadside Slopes in An-Dean Patagonian Forests of Austrocedrus chilensis (Argentina). Environ. Earth Sci. Proc. 2024, 31, 18. https://doi.org/10.3390/eesp2024031018
Chichizola GA, Gonzalez SL, Rovere AE. Early Detection of Invasive Species on Roadside Slopes in An-Dean Patagonian Forests of Austrocedrus chilensis (Argentina). Environmental and Earth Sciences Proceedings. 2024; 31(1):18. https://doi.org/10.3390/eesp2024031018
Chicago/Turabian StyleChichizola, Giselle Ailin, Sofía Laura Gonzalez, and Adriana Edit Rovere. 2024. "Early Detection of Invasive Species on Roadside Slopes in An-Dean Patagonian Forests of Austrocedrus chilensis (Argentina)" Environmental and Earth Sciences Proceedings 31, no. 1: 18. https://doi.org/10.3390/eesp2024031018
APA StyleChichizola, G. A., Gonzalez, S. L., & Rovere, A. E. (2024). Early Detection of Invasive Species on Roadside Slopes in An-Dean Patagonian Forests of Austrocedrus chilensis (Argentina). Environmental and Earth Sciences Proceedings, 31(1), 18. https://doi.org/10.3390/eesp2024031018