
Molecular Epidemiology of mcr (1–5) and Other Critical Resistance Genes in Bacteria Isolated from Canine Otitis Externa in Ankara, Türkiye



Abstract
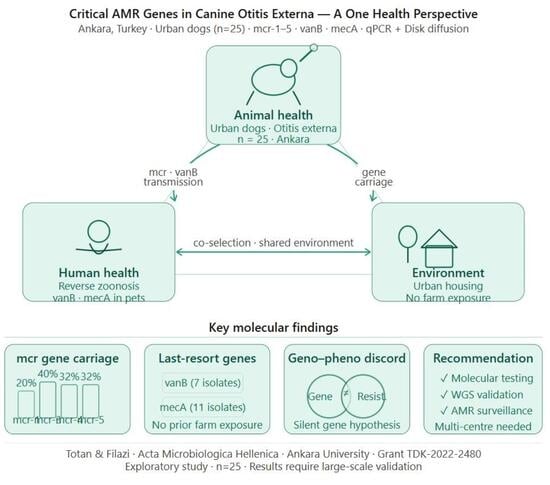
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Bacteriological Examination
2.3. Antibiotic Susceptibility Tests
2.4. Identification of Resistance Genes
2.5. Statistical Analysis
3. Results
3.1. Isolates Detected and Resistance Genes of the Isolates
3.2. Antibiogram Results (40 Antibiotics)
3.3. Antimicrobial Resistance Distribution
3.4. Concordance Between Resistance Genes and Phenotypic Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tesin, N.; Stojanovic, D.; Stancic, L.; Kladar, N.; Ružíc, Z. Prevalence of the microbiological causes of canine otitis externa and the antibiotic susceptibility of the isolated bacterial strains. Pol. J. Vet. Sci. 2023, 26, 449–459. [Google Scholar] [CrossRef]
- Awad, A.; Hassoon, S.; Ali, H.; Mohammad, S. Cultural and biochemical identification of antibi-otic-resistant bacteria in the ears of dogs and patients with otitis externa. Arch. Vet. Med. 2025, 18, 51–70. [Google Scholar] [CrossRef]
- De Martino, L.; Nocera, F.P.; Mallardo, K.; Nizza, S.; Masturzo, E.; Fiorito, F.; Iovane, G.; Catalanotti, P. An update on microbiological causes of canine otitis externa in Campania Region, Italy. Asian Pac. J. Trop. Biomed. 2016, 6, 384–389. [Google Scholar] [CrossRef]
- Li, Y.; Fernández, R.; Durán, I.; Molina-López, R.A.; Darwich, L. Antimicrobial resistance in bacteria isolated from cats and dogs from the Iberian Peninsula. Front. Microbiol. 2021, 11, 3628. [Google Scholar] [CrossRef] [PubMed]
- Parasana, D.K.; Makwana, P.M.; Shobha, K.; Prakruti, J.P.; Irshadullakhan, H.K. Isolation and Identification of Corynebacterium Species from cases of skin and ear infection of dogs. Int. J. Vet. Sci. Anim. Husb. 2023, 8, 31–33. [Google Scholar]
- Beaudoin, A.L.; Bollig, E.R.; Burgess, B.A.; Cohn, L.A.; Cole, S.D.; Dear, J.D.; Fellman, C.L.; Frey, E.; Goggs, R.; Johnston, A.; et al. Prevalence of antibiotic use for dogs and cats in United States veterinary teaching hospitals. J. Vet. Intern. Med. 2023, 37, 1864–1875. [Google Scholar] [CrossRef]
- Chan, W.Y.; Hobi, S.; Ferguson, A.; Elsohaby, I. Canine Pyoderma and Otitis Externa: A Retrospective Analysis of Multidrug-Resistant Bacterial Carriage in Hong Kong. Antibiotics 2025, 14, 685. [Google Scholar] [CrossRef]
- Schwarz, S.; Johnson, A.P. Transferable resistance to colistin: A new but old threat. J. Antimicrob. Chemother. 2016, 71, 2066–2070. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a novel mcr-6 to mcr-9 multiplex PCR and assessment of mcr-1 to mcr-9 occurrence in colistin-resistant Salmonella enterica isolates from environment, feed, animals and food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef]
- Wang, G.; Liu, H.; Feng, Y.; Zhang, Z.; Hu, H.; Liu, J.; Qiu, L.; Guo, Z.; Huang, J.; Qiu, J.; et al. Colistin-resistance mcr genes in Klebsiella pneumoniae from companion animals. J. Glob. Antimicrob. Resist. 2021, 25, 35–36. [Google Scholar] [CrossRef] [PubMed]
- Hamame, A.; Davoust, B.; Hasnaoui, B.; Mwenebitu, D.L.; Rolain, J.M.; Diene, S.M. Screening of colistin-resistant bacteria in livestock animals from France. Vet. Res. 2022, 53, 96. [Google Scholar] [CrossRef]
- Kwon, J.; Ko, H.J.; Yang, M.H.; Park, C.; Park, S.C. Antibiotic Resistance and Species Profile of Enterococcus Species in Dogs with Chronic Otitis Externa. Vet. Sci. 2022, 9, 592. [Google Scholar] [CrossRef]
- Oh, J.Y.; Kwak, S.M.; Kim, J.Y.; Ro, W.B.; Lee, K.J.; Chae, J.C. Case Report: Transferable IncX4 plasmid carrying mcr-1 in colistin-resistant Escherichia coli from a healthy pet dog in South Korea. Front. Vet. Sci. 2026, 12, 1746399. [Google Scholar] [CrossRef]
- Agnihotri, D.; Charaya, G.; Chabbra, R.; Kumar, T.; Jain, V.K. Antibiogram of bacteria isolated from dogs suffering from otitis externa. Indian J. Comp. Microbiol. Immunol. Infect. Dis. 2019, 40, 15–20. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Test for Bacteria Isolated from Animals. In Approved Standard Document, 5th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- European Medicines Agency. Categorisation of Antibiotics Used in Animals Promotes Responsible Use to Protect Public and Animal Health. 2020. Available online: https://www.ema.europa.eu/en/news/categorisation-antibiotics-used-animals-promotes-responsible-use-protect-public-animal-health (accessed on 20 July 2025).
- DiaRex. Genomic DNA Extraction Kit User Manual; Diagen Biotechnological Systems: Ankara, Turkey, 2024. [Google Scholar]
- Monteiro, J.; Widen, R.H.; Pignatari, A.C.; Kubasek, C.; Silbert, S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J. Antimicrob. Chemother. 2012, 67, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Davies, R.H.; Threlfall, E.J. Mechanisms of quinolone resistance in Escherichia coli and Salmonella: Recent developments. Int. J. Antimicrob. Agents 2005, 25, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef]
- World Health Organisation. Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. 2017. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 20 July 2025).
- World Organisation for Animal Health. WOAH List of Antimicrobials of Veterinary Importance. 2021. Available online: https://www.woah.org/app/uploads/2021/06/a-oie-list-antimicrobials-june2021.pdf (accessed on 20 July 2025).
- Kempf, I.; Fleury, M.A.; Drider, D.; Bruneau, M.; Sanders, P.; Chauvin, C.; Haenni, M.; Jouy, E. What do we know about resistance to colistin in Enterobacteriaceae in avian and pig production in Europe? Int. J. Antimicrob. Agents 2013, 42, 379–383. [Google Scholar] [CrossRef]
- Lei, L.; Wang, Y.; Schwarz, S.; Walsh, T.R.; Ou, Y.; Wu, Y.; Li, M.; Shen, Z. mcr-1 in Enterobacteriaceae from Companion Animals, Beijing, China. Emerg. Infect. Dis. 2017, 23, 710. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.C.; Kim, S.J.; Mechesso, A.F.; Kang, H.Y.; Song, H.J.; Choi, J.-H.; Yoon, S.-S.; Lim, S.-K. Mobile colistin resistance gene mcr-1 detected on an IncI2 plasmid in Salmonella typhimurium sequence type 19 from a healthy pig in South Korea. Microorganisms 2021, 9, 398. [Google Scholar] [CrossRef]
- Khosravi, Y.; Tay, S.T.; Vadivelu, J. Metallo-β-lactamase-producing imipenem-resistant Pseudomonas aeruginosa clinical isolates in a university teaching hospital in Malaysia: Detection of IMP-7 and first identification of IMP-4, VIM-2, and VIM-11. Diagn. Microbiol. Infect. Dis. 2010, 67, 294–296. [Google Scholar] [CrossRef]
- Ocak, M.; Özer, B.; İnci, M.; Duran, N. Antibiotic Resistance and Investigation of IMP-1, IMP-2, VIM-1 and VIM-2 Metallo-β-Lactamases in Acinetobacter Strains Isolated From Clinical Samples. Klimik J. 2015, 28, 23. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Schwarz, S.; Zhang, R.; Lei, L.; Liu, X.; Shen, J. IMP-45-producing multidrug-resistant Pseudomonas aeruginosa of canine origin. J. Antimicrob. Chemother. 2014, 69, 2579–2581. [Google Scholar] [CrossRef]
- KuKanich, K.S.; Bagladi-Swanson, M.; KuKanich, B. Pseudomonas aeruginosa susceptibility, antibiogram and clinical interpretation, and antimicrobial prescribing behaviors for dogs with otitis in the Midwestern United States. J. Vet. Pharmacol. Ther. 2022, 45, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Zamankhan Malayeri, H.; Jamshidi, S.; Zahraei Salehi, T. Identification and antimicrobial susceptibility patterns of bacteria causing otitis externa in dogs. Vet. Res. Commun. 2010, 34, 435–444. [Google Scholar] [CrossRef]
- Park, Y.; Oh, J.; Park, S.; Sum, S.; Song, W.; Chae, J.; Park, H. Antimicrobial resistance and novel mutations detected in the gyrA and parC genes of Pseudomonas aeruginosa strains isolated from companion dogs. BMC Vet. Res. 2020, 16, 111. [Google Scholar] [CrossRef]
- Cho, J.K.; Kim, J.M.; Kim, K.H.; Lim, H.S.; Yang, C.R. Antimicrobial resistance of Pseudomonas aeruginosa isolated from dogs and cats. Korean J. Vet. Serv. 2021, 44, 21–26. [Google Scholar]
- Livermore, D.M.; Day, M.; Cleary, P.; Hopkins, K.L.; Toleman, M.A.; Wareham, D.W.; Wiuff, C.; Doumith, M.; Woodford, N. OXA-1 β-lactamase and non-susceptibility to penicillin/β-lactamase inhibitor combinations among ESBL-producing Escherichia coli. J. Antimicrob. Chemother. 2019, 74, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Oteo, J.; González-López, J.J.; Ortega, A.; Quintero-Zárate, J.N.; Bou, G.; Cercenado, E.; Conejo, M.C.; Martínez-Martínez, L.; Navarro, F.; Oliver, A.; et al. Inhibitor-resistant TEM-and OXA-1-producing Escherichia coli isolates resistant to amoxicillin-clavulanate are more clonal and possess lower virulence gene content than susceptible clinical isolates. Antimicrob. Agents Chemother. 2014, 58, 3874–3881. [Google Scholar] [CrossRef] [PubMed][Green Version]



{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Symbol/Concentration | Antibiotic | Symbol/Concentration |
|---|---|---|---|
| Aminosidin | AN-60 µg | Florfenicol | FFC-30 µg |
| Amoxicillin | AML-20 µg | Fosfomycin | FF-30 µg |
| Amoxicillin/clavulanic acid | AMC-30 µg | Imipenem | IMP-10 µg |
| Ampicillin/cloxacillin | APX 30 µg | Lincomycin | L-15 µg |
| Ampicillin/sulbactam | SAM-20 µg | Marbofloxacin | MAR-5 µg |
| Cefazolin | CZ-30 µg | Methicillin | ME-5 µg |
| Cefepime | FEP-10 µg | Metronidazole | MET-10 µg |
| Cefoperazone | CEP-75 µg | Neomycin | N-30 µg |
| Cefquinome | CEQ-30 µg | Oxytetracycline | T-30 µg |
| Ceftriaxone | CRO-30 µg | Penicillin G | P-10 U |
| Cephalexin | CL-30 µg | Spectinomycin | SPT-25 µg |
| Chloramphenicol | C-30 µg | Spiramycin | SP-30 µg |
| Ciprofloxacin | CIP-10 µg | Streptomycin | S-25 µg |
| Clarithromycin | CLR-15 µg | Sulfadiazine | SD-300 µg |
| Clindamycin | DA-10 µg | Sulphamethoxazole | SMZ-100 µg |
| Cloxacillin | CX-30 µg | Tetracycline | TE-30 µg |
| Colistin | CT-25 µg | Trimethoprim | TMP-30 µg |
| Danofloxacin | DAN-5 µg | Trimethoprim/sulphamethoxazole | SXT-25 µg |
| Doxycycline | DO-30 µg | Tulathromycin | TUL-30 µg |
| Erythromycin | E-15 µg | Vancomycin | VA-10 µg |
| Target Gen | Primer Name | Primer Sequence (5′ → 3′) |
|---|---|---|
| mcr-1 | colistin_mcr_1-F | GTCCGTTTGTTCTTGTGG |
| colistin_mcr_1-R | GTCTGTAGGGCATTTTGG | |
| mcr-2 | colistin_mcr_2-F | GTATTCTGTGCCGTGTATG |
| colistin_mcr_2-R | GTATTGTTGGTTGCTGATTT | |
| mcr-3 | colistin_mcr_3-F | GCCTCATTTTGATTGGTTTC |
| colistin_mcr_3-R | TAAGTTTGGTTTCGCCATTT | |
| mcr-4 | colistin_mcr_4-F | CCCGAACACTAAACCTAAC |
| colistin_mcr_4-R | AAACATACAGGGTAGAGACA | |
| mcr-5 | colistin_mcr_5-F | ACTGATTCTGCTTGCTGT |
| colistin_mcr_5-R | TCATTACCGCTTGTTTCC | |
| blaIMP | Imipenem-F | GAGTGGCTTAATTCTCRATC |
| Imipenem-R | AACTAYCCAATAYRTAAC | |
| gyrA | gyrA-F | TGTCCGAGATGGCCTGAAGC |
| gyrA-R | TACCGTCATAGTTATCCACG | |
| gyrB | gyrB-F | TCGGCGTCGTTGTTGTCATA |
| gyrB-R | GCGGTGGGTTTCAAAATCTG | |
| ampC | Cefepime_ampC-F | TTCTTGTCTACTTTTATCCCC |
| Cefepime_ampC-R | ACTGCTATTTACGGCTTTTT | |
| OXA-1 | Amoxicillin + clavulanic acid_OXA1-F | TTTTCTGTTGTTTGGGTTTC |
| Amoxicillin + clavulanic acid_OXA1-R | CTATGGTGTTTTCTATGGCT |
| No. | Dog Breed | Age-Weight-Sex | Dominant Microorganism | Detected Resistance Genes/Determinants 2 | Phenotypic Resistance Profile |
|---|---|---|---|---|---|
| 1 | Belgian Sheepdog | 4 years old-35 kg-female | Pseudomonas aeruginosa | mcr-4, gyrA | Ciprofloxacin, Sulfonamides |
| 2 | Beagle | 3 years old-17 kg-male | Pseudomonas aeruginosa | mcr-4, ampC, gyrA | Ciprofloxacin, Beta-lactams |
| 3 | Yorkshire Terrier | 5 years old-3 kg-male | Staphylococcus aureus | – | Beta-lactams, Sulfonamides |
| 4 | Basset Hound | 10 years old-36 kg-male | Staphylococcus epidermidis | gyrA, gyrB, OXA-1 | Ciprofloxacin, Amoxicillin-Clav, Beta-lactams |
| 5 | Golden Retriever | 10 years old-33 kg-male | Pseudomonas aeruginosa | mcr-3, ampC, gyrA, gyrB | Ciprofloxacin, Beta-lactams |
| 6 | German Shepherd | 7 years old-30 kg-male | Enterobacter spp. | mcr-4, ampC, gyrA, gyrB | Imipenem, Ciprofloxacin |
| 7 | Pug | 4 years old-8 kg-female | Enterobacter spp. | mcr-1, mcr-5 | Imipenem, Sulfonamides |
| 8 | Akita | 1 year old-36 kg-male | Streptococcus spp. | gyrA, gyrB, OXA-1 | Ciprofloxacin, Beta-lactams |
| 9 | English Cocker Spaniel | 7 years old-10 kg-male | Staphylococcus epidermidis | gyrA, gyrB, OXA-1 | Ciprofloxacin |
| 10 | Golden Retriever | 13 years old-28 kg-female | Staphylococcus aureus | gyrA, gyrB | Ciprofloxacin, Beta-lactams |
| 11 | Maltese Terrier | 4 years old-2.5 kg-female | Enterobacter spp. | mcr-3, gyrA, OXA-1 | Imipenem, Ciprofloxacin |
| 12 | English Cocker Spaniel | 11 years old-9 kg-male | Staphylococcus epidermidis | gyrA | Ciprofloxacin, Beta-lactams |
| 13 | Bernese Mountain Dog | 6 years old-50 kg-female | Staphylococcus aureus | OXA-1, gyrA | Ciprofloxacin, Amoxicillin-Clav |
| 14 | Chihuahua | 7 years old-1.5 kg-male | Staphylococcus aureus | gyrA, gyrB | Ciprofloxacin, Beta-lactams, Sulfonamides |
| 15 | Golden Retriever | 3 years old-28 kg-male | Staphylococcus epidermidis | gyrA | Ciprofloxacin |
| 16 | Golden Retriever | 3 years old-33 kg-female | Staphylococcus aureus | gyrA | Ciprofloxacin |
| 17 | Cavalier King Charles Spaniel | 10 years old-15 kg-male | Enterobacter spp. | mcr-1, ampC | Imipenem, Colistin, Beta-lactams |
| 18 | Yorkshire Terrier | 7 years old-2 kg-male | Staphylococcus aureus | gyrA | Ciprofloxacin |
| 19 | Golden Retriever | 3 years old-25 kg-female | Streptococcus spp. | OXA-1 | Beta-lactams |
| 20 | Golden Retriever | 10 years old-33 kg-female | Pseudomonas aeruginosa | mcr-3, mcr-5, gyrA, gyrB | Ciprofloxacin, Beta-lactams |
| 21 | Hybrid Dog | 2 years old-23 kg-female | Staphylococcus epidermidis | – | Beta-lactams |
| 22 | Golden Retriever | 13 years old-40 kg-female | Staphylococcus aureus | gyrA | Ciprofloxacin, Beta-lactams |
| 23 | Kangal Shepherd Dog | 1 year old-45 kg-male | Enterobacter spp. | mcr-3, mcr-5, gyrA | Imipenem, Ciprofloxacin |
| 24 | Golden Retriever | 9 years old-35 kg-female | Streptococcus spp. | gyrA, OXA-1 | Ciprofloxacin |
| 25 | English Cocker Spaniel | 5 years old-11 kg-female | Staphylococcus epidermidis | gyrA | Ciprofloxacin, Beta-lactams |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Published by MDPI on behalf of the Hellenic Society for Microbiology. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Totan, F.E.; Filazi, A. Molecular Epidemiology of mcr (1–5) and Other Critical Resistance Genes in Bacteria Isolated from Canine Otitis Externa in Ankara, Türkiye. Acta Microbiol. Hell. 2026, 71, 14. https://doi.org/10.3390/amh71020014
Totan FE, Filazi A. Molecular Epidemiology of mcr (1–5) and Other Critical Resistance Genes in Bacteria Isolated from Canine Otitis Externa in Ankara, Türkiye. Acta Microbiologica Hellenica. 2026; 71(2):14. https://doi.org/10.3390/amh71020014
Chicago/Turabian StyleTotan, Fatma Esra, and Ayhan Filazi. 2026. "Molecular Epidemiology of mcr (1–5) and Other Critical Resistance Genes in Bacteria Isolated from Canine Otitis Externa in Ankara, Türkiye" Acta Microbiologica Hellenica 71, no. 2: 14. https://doi.org/10.3390/amh71020014
APA StyleTotan, F. E., & Filazi, A. (2026). Molecular Epidemiology of mcr (1–5) and Other Critical Resistance Genes in Bacteria Isolated from Canine Otitis Externa in Ankara, Türkiye. Acta Microbiologica Hellenica, 71(2), 14. https://doi.org/10.3390/amh71020014