Abstract
Chemical studies usually consist of measurements made on large ensembles of molecules with data representing average values for the population. It has been shown that individual molecules of a given enzyme have different properties. Large-scale averaging has in the past masked these differences. Alkaline phosphatase has been used as a model to study this enzyme heterogeneity. The catalytic rates of the individual molecules have been found to differ by over 10-fold, and the activation energy of catalysis by more than two-fold. Differences in properties indicate that differences in structure must exist between the molecules. For alkaline phosphatase, the structural differences have been suggested to be differences in glycosylation, differences due to partial proteolysis, and due to some molecules containing mixtures of active and inactive subunits. The determination of the distribution of activities of populations of this enzyme within a sample has also been shown to be a useful tool in diagnostics. This review discusses the advent of single-molecule enzymology and summarizes its use in the study of alkaline phosphatase using capillary electrophoresis, microscopic well assays, and single-molecule tracking.
1. Introduction
Classical chemical studies involve measurements made on ensembles containing very large numbers of individual molecules. The data obtained represent average values for these large ensembles. Any differences between individual molecules are not observed. With respect to enzyme molecules, it has been shown that individual molecules of a given molecule have different properties. Single-enzyme molecule studies have been performed on a variety of enzymes, including lactate dehydrogenase [1], alkaline phosphatase [2,3,4], β-galactosidase [5], peroxidase [6], β-glucuronidase [7], and cholesterol oxidase [8,9], while using different experimental approaches [1,10,11]. Enzyme molecules have consistently been found to be heterogeneous. Such heterogeneity appears to be a general property of enzymes. Heterogeneous properties include catalytic rate [1], activation energy of catalysis [2], electrophoretic mobility [12], Km [13], Ki [14], and ion binding [15]. Chemical dogma states that differences in structure must underlie differences in properties. Single-molecule enzymology studies have been used as a means to understand these differences and have suggested that differences in conformation may be an important component [16]. Furthermore, single-molecule enzyme analysis has been used as a means to determine the distribution of properties of a population for the diagnosis [17] and elucidation of the differences between fully and half-active subpopulations [18], as well as an analytical tool. In this article, the initial single-molecule enzyme assays and later single-molecule studies involving alkaline phosphatase are reviewed. This is the first such review to summarize the single-molecule work conducted with alkaline phosphatase over the past 3 decades.
2. First Single-Enzyme Molecule Assay
The first report on the measurement of the activity of an individual enzyme molecule was by Rotman in 1961 [19]. A solution of E. coli β-galactosidase containing the fluorogenic substrate, which was also developed in this study, 6-hydroxyfluran-β-D-galactopyranoside was dispersed into 14–15 μm diameter droplets which were coated in silicone oil. The droplets were incubated for 15 h and imaged using a fluorescence microscope.
The average number of molecules in a given droplet, n, is determined from the enzyme concentration and the droplet volume. However, due to random variation, not all droplets are expected to contain the same number of enzyme molecules. The number of enzyme molecules per droplet (r) is given by the Poisson distribution where the probability p(r) of finding a given number of molecules in a particular droplet is:
p(r) = (nre−n/r!)
If n = 1, the probability of a droplet containing no enzyme molecules is 0.37, 1 enzyme molecule is 0.37, 2 enzyme molecules is 0.18, and 3 enzyme molecules is 0.06. From the proportion of the number of droplets where no product is detected, the concentration of the enzyme can be calculated. When no enzyme was present, no signal was detected in the 1700 droplets imaged over 60 experiments. When the enzyme was present, the droplets were found to fall into distinct categories: droplets without fluorescence, droplets with a given amount of fluorescence, droplets with approximately twice the given amount of fluorescence, and droplets with three or more times the given amount of fluorescence. This suggested that the droplets contained zero, one, two, and three enzyme molecules. The concentration of enzyme was determined from this distribution, and from the mass concentration, the molecular weight of the enzyme was determined.
In a different set of experiments, Rotman compared β-galactosidase from extracts of wild-type E. coli to that from a genetically altered strain. Extract from the altered strain showed 50–60-fold less activity per mg of protein in the extract. The objective was to determine if the less active extract contained less active enzyme molecules or molecules of the same activity but present at a lower concentration. He determined that the altered enzyme contained molecules with lower activity. In a third set of experiments, Rotman compared native and partially heat-denatured enzymes. He determined that partial thermal denaturation resulted in a fraction of the molecules being completely inactivated and the remaining being unaffected.
3. Individual Molecules of a Given Enzyme Have Different Properties
Rotman’s work on the analysis of single molecules of enzymes was followed by that of Yeung’s group [1] more than 3 decades later. This study used a capillary electrophoresis-based approach. In this approach, a 65 cm long, 50 cm separation length, 20 μm internal diameter capillary was filled with buffer containing lactate, NAD+, and very dilute lactate dehydrogenase-1 (LDH-1) from red blood cells [20]. Enzyme concentration was such that approximately ten molecules were randomly distributed within the internal 290 nL volume of the capillary. Capillary contents were then incubated without mobilization for 1 h. During this time, the LDH-1 molecules converted the NAD+ into the fluorescent product NADH. Since the incubation period was relatively short, and the distance between the individual LDH-1 molecules was relatively large, there was insufficient time for the NADH formed by one enzyme molecule to diffuse to and mix with that of another. Rather, product pools of NADH accumulated in the immediate vicinity of the particular enzyme molecules that produced them. Post-incubation, electrophoretic mobilization of these product pools past a highly sensitive laser-induced fluorescent detector yielded distinct peaks in the resultant electropherogram, each representing the product made by an individual LDH-1 molecule.
The number of peaks was similar to that predicted from the nominal enzyme concentration and the capillary volume. The number of peaks was proportional to enzyme concentration. Peak area was found to differ by a factor of up to 4. In a follow-up experiment, the capillary was filled with enzyme and substrate and subjected to 2 sequential 1 h incubation periods with an intervening 3 min period of mobilization. This period of mobilization separated the LDH-1 molecules from their respective pools of NADH and into the fresh substrate. During the following incubation period, a second product pool was formed by each enzyme molecule. The resultant electropherogram yielded sets of two peaks of indistinguishable areas, one for each incubation. Each set was formed by a particular individual molecule. This demonstrated that the individual molecules have reproducible but distinct activities. This was the first demonstration that individual molecules of a given enzyme are heterogeneous. Yeung suggested that the differences in activity may result from long-lived conformational differences between the individual molecules.
The capillary electrophoresis method has also been used to study the properties of individual molecules of other enzymes, including alkaline phosphatase [2], β-galactosidase [5], and β-glucuronidase [21].
4. Single-Molecule Alkaline Phosphatase Assays Using Capillary Electrophoresis
Dovichi’s group also used the capillary electrophoresis approach and confirmed the finding that enzyme molecules are heterogeneous using calf-intestinal alkaline phosphatase (cIAP) [2,3,4]. A capillary was filled with buffer, and a wide plug of fluorogenic substrate containing very dilute enzyme was injected into a 70 cm long, 10 μm internal diameter capillary. The sample was incubated, and the product formed accumulated in the immediate vicinity of the individual cIAP molecules. After a 20–40 min incubation period, the capillary contents were mobilized past a laser-induced fluorescence detector. The resultant electropherogram contained peaks, each due to the product pool formed by a given individual enzyme molecule. The average number of peaks per run was near-identical to the number of enzyme molecules expected based on the nominal enzyme concentration and the injection volume. The average number of peaks was proportional to enzyme concentration. The number of peaks per run was dominated by Poisson statistics. This was deemed sufficient to demonstrate that individual enzyme molecules were being assayed. As was the case with LDH-1, peak areas varied, in this case, by over 10-fold [2]. Figure 1 shows a representative electropherogram of the capillary electrophoresis-based assay of individual molecules of cIAP [2].
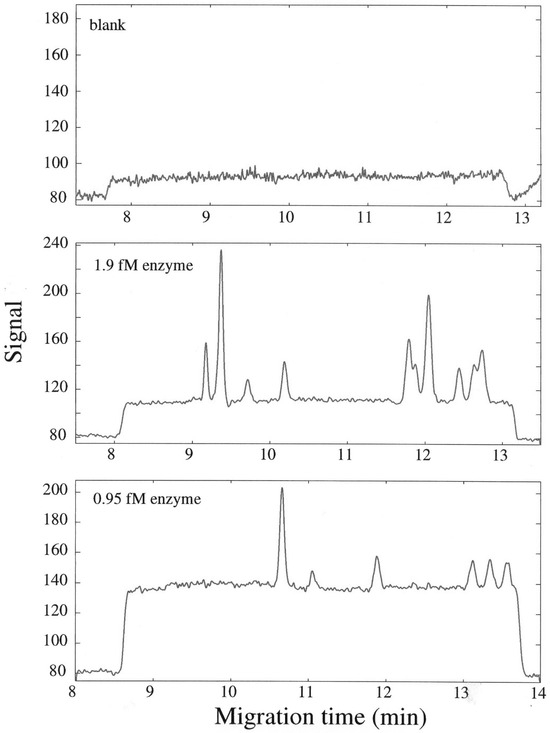
Figure 1.
Assay of individual molecules of alkaline phosphatase. A wide plug of buffer containing substrate (2’-[2-benzthiazoyl]-6’-hydroxy-benzthiazole phosphate bis-[2-amino-2-methyl-1,3-propanesiol salt) was injected into a capillary and incubated statically for 30 min. Post-incubation, the capillary contents were mobilized past the detector at 400 Vcm−1. A plateau due to the presence of product (2’-[2-benzthiazoyl]-6’-hydroxy-benzthiazole), formed from the non-enzymatic hydrolysis of substrate, was observed (upper trace). In the middle trace, a plug of buffer containing substrate and very dilute calf-intestinal alkaline phosphatase was injected into the capillary. The enzyme concentration was such that ten molecules were present in the capillary. The capillary contents were again incubated and then mobilized past the detector. Peaks were observed sitting atop the plateau of the product formed by background hydrolysis. Each peak is due to a pool of product that formed by an individual enzyme molecule. When the concentration of enzyme was reduced by half, the average number of peaks observed was also reduced by half (lower trace) [2].
Individual cIAP molecules were subjected to replicate incubations as performed with LDH-1. Quadruple incubations with incubation times of 8, 4, 2, and 1 min, with intervening 15 s periods of electrophoresis, were performed. In the resulting electropherogram, sets of four evenly spaced peaks were observed. The average linear correlation between peak area and incubation time was 0.996 [2]. This demonstrated that the individual molecules showed consistent catalytic rates despite each incubation occurring in a different position along the capillary. This indicates that differences in rate between the molecules could not be attributed to an artifact caused by differences in the micro-environment, such as adsorption to the capillary wall. This study confirmed Yeung’s finding [1] that enzyme molecules have heterogeneous activity.
In a different experiment, triple incubations were performed, and the capillary temperature was held at a different temperature for each incubation. The resultant electropherograms contained sets of three evenly spaced peaks. Arrhenius plots were constructed for each molecule, and the activation energy of catalysis was determined. Activation energy was also found to be a heterogeneous property, ranging from 39 to 91 kJmol−1 [2].
Thermal denaturation was also studied at the single-molecule level. It was determined that an incubation of 5 min at 64 °C resulted in a loss of 50% of the activity of a bulk solution of alkaline phosphatase. Control and heated enzymes were assayed at the single-molecule level, and it was found that heating decreased the number of active molecules by 50%, and the remaining active molecules had an activity indistinguishable from the control enzyme. This was consistent with Rotman’s [19] finding that partial thermal denaturation results in total inactivation of a portion of the enzyme molecules, with those that were not denatured remaining unaffected.
Eukaryotic alkaline phosphatase is glycosylated, and the glycosylation pattern has been found to be heterogeneous [22,23]. Dovichi [2] proposed that differential glycosylation may be a source of the heterogeneity of cIAP.
E. coli alkaline phosphatase is not glycosylated and exists in three forms: Isoform 1 contains an N-terminal arg residue on each of its two subunits, whereas isoform 3 does not. Isoform 2 is a heterodimer of isoforms 1 and 3 [24,25]. Single-molecule assays were originally performed on E. coli alkaline phosphatase by Dovichi’s group [26] in order to determine if non-glycosylated enzymes are heterogeneous. Since the E. coli enzyme has a lower activity, the assay was a modification of that performed using the bovine enzyme. Commercial preparations of the enzyme were found to have heterogeneous activity. The E. coli enzyme was then separated into its different isoforms using iso-electric focusing. Single-molecule assays demonstrated that the individual molecules of the purified isoforms had indistinguishable activities. Furthermore, it was found that if the enzyme was stored in the absence of protease inhibitors the enzyme activity became heterogeneous. Based on the previously reported heterogeneity of the activity of glycosylated cIAP [2], the homogeneity in activity of the non-glycosylated highly purified bacterial enzyme, and that the bacterial enzyme becomes heterogeneous when stored in the absence of protease inhibitor, Dovichi concluded that differences in post-translation modification may account for the heterogeneity of alkaline phosphatase without invoking differences in conformation. However, single-molecule studies on E. coli β-galactosidase have shown that differences in conformation result in differences in both catalytic rate and electrophoretic mobility [16,27]. This suggests that different enzymes may have a different mix of structural causes for their observed heterogeneity.
Measurement of the electrophoretic mobility of individual enzyme molecules allows for insight into what differences in charge, size, and shape may exist between the molecules in order to account for the observed differences in mobility [28,29]. Differences in mobility could be accounted for by small differences in charge and size [21,30,31]. The distribution of electrophoretic mobilities of individual bIAP molecules was measured in the presence of CHES and borate buffers but in an otherwise identical manner in order to assess if small differences in glycosylation can result in an increase in mobility heterogeneity. Borate forms negatively charged complexes with diols, including carbohydrates, whereas CHES does not [32]. Electrophoretic mobility was found to be heterogeneous in both buffers; however, in the presence of the borate, it was significantly more negative and had a significantly higher variance. In the presence of CHES, the variance in mobility was attributed to small differences in charge or size of the individual molecules. The additional variance in the presence of borate was attributed to differences in the carbohydrate moieties attached to the enzyme [31], supporting Dovichi’s [2] proposal that differences in glycosylation may be a source of enzyme heterogeneity for cIAP.
Single-molecule assays were also performed on alkaline phosphatase from Thermus thermophilus grown at 70 °C to determine if multiple active forms of the enzyme exist when grown under stringent conditions [33]. The enzyme was found to be heterogeneous with respect to its activity but to a lesser extent than cIAP.
The premise of the production of recombinant proteins is that they are identical to those produced in the natural source. This was tested by comparing bIAP with the commercially available recombinant enzyme expressed in P. pastoris. The average catalytic rate, its variance, and distribution were found to be indistinguishable between the two sources of the enzyme [34]. This is, however, not necessarily true for all proteins as the activity distribution of in vivo and cell-free expressed E. coli β-galactosidase have been found to differ [35].
5. Single-Molecule Alkaline Phosphatase Assays Using Microscopic Wells
In this methodology, a bundle of thousands of optical fibers was etched on the ends, forming femtoliter-sized wells. A very dilute solution of enzyme-containing fluorogenic substrate was trapped in wells upon sealing the end of the bundle with a glass slide using oil. The rate of increase in fluorescence signal, due to the activity of the trapped enzyme, in each well was monitored over time. Similar to the approach used by Rotman [19], when there is a very low concentration of enzyme, the majority of the wells contain no enzyme, a reasonable proportion contain a single-enzyme molecule, and the probability of a given well containing two or more enzyme molecules is very low. This allows for the assumption that in the very large majority of the time, in any well that product is detected it was formed by a single-enzyme molecule.
This method has been used to study the properties of individual molecules of alkaline phosphatase [36], β-galactosidase [37], β-glucuronidase [7], and peroxidase [6]. Air-sealing the femtoliter wells has also been reported to be an improvement over sealing using oil [38].
Since this method monitors thousands of microscopic wells at a time, it is ideal for the determination of the distribution of single-molecule activities of a given population. Serum contains more than one isozyme of alkaline phosphatase. The ratio of the different isozymes is a marker for some diseases. Walt’s group used this methodology to determine the distribution of activities of standards of 2 different alkaline phosphatase isozymes as well as the distribution of the isozyme mixture found in serum. Comparison of these distributions allowed for the determination of the amount of each of the different isoenzymes present in the mixture [36,39]. Changes in the distribution of activities upon changes in pH and incubation at elevated temperatures have also been used to study changes in the distribution of lower- and higher-activity isoforms [40].
In a variation of the same assay, a slide was etched to form an array of microscopic wells, and the array was monitored using fluorescence microscopy. In this study, two different enzymes, alkaline phosphatase and β-galactosidase, and substrates for both, were present in the wells. This was conducted to test the feasibility of simultaneously performing two different immunoassays at the single-molecule level. The array was imaged for the products of both enzymes simultaneously, and wells were found to contain either no enzyme, a molecule of β-galactosidase, a molecule of alkaline phosphatase, or a molecule of each [41]. Noji’s group studied E. coli alkaline phosphatase using this method and proposed that the E. coli enzyme contains two distinct sub-populations: half-active dimers containing an active and an inactive monomer, and fully active dimers containing two active monomers [18].
As a means to detect different phosphatase enzymes in the same sample, an array of wells was filled with blood samples. Any given well contained no enzyme molecules or an individual molecule of either alkaline phosphatase, tyrosine phosphatase, or ectonucleotide phosphatase. The wells were assayed for activity with different substrates, and the relative activity for each substrate was used to determine the identity of the particular phosphatase present. From this, the amount of each phosphatase in a sample can be determined. The concentrations of several different phosphatases have been shown to differ in serum samples of healthy and diabetic patients. Methodology for the determination of such distributions has the potential to be a useful tool in diagnosis [17].
6. In-Solution Single-Particle Tracking
Unlike the others, in this method, the product formed by an individual alkaline phosphatase is not what was measured. Rather, the enzyme was labeled with a fluorescent dye, and it was the attached label that produced the signal that was monitored. The labeled enzyme was situated in a free solution between a slide and a cover slip, and the emitted photons were used to map the position of the molecule over time using fluorescence microscopy.
At the nanoscale, Brownian diffusion is the dominant factor in the mobility of a molecule. Studies have suggested that this mobility may be increased in the presence of substrates through various mechanisms. In this study, the diffusion mobilities of individual labeled alkaline phosphatase molecules were measured. It was found that for alkaline phosphatase, no such enhancement in mobility can be attributed to the presence of substrate [11].
7. Significance of Enzyme Heterogeneity
Studies on large ensembles of individual enzyme molecules provide data that represent average values. This obfuscates the diversity of the molecules. Single-molecule enzymology studies have demonstrated the previously unappreciated concept that individual molecules of a given enzyme have different properties. Several different enzymes have been studied at the single-molecule level using a variety of different techniques. In every case, heterogeneity has been observed. Heterogeneity, therefore, appears to be a general characteristic of enzymes.
Dogma states that nature puts considerable effort to ensure the fidelity of protein transcription and translation to ensure that all the molecules of a given protein are made to be the same. One of the research goals of single-molecule enzymology has been to assess these differences and to determine their structural basis. Three different sources of these structural differences can be envisaged. Firstly, there are errors in translation and transcription. Such error rates, leading to differences in protein sequence, have been suggested not to be a significant source of the observed heterogeneity [42]. A second possibility is differences in conformation. This has been suggested to be a major source of enzyme heterogeneity [16]. A third possibility is differential post-translational modifications. Initial work suggests this may be significant [31].
Whether there is a biological role associated with enzyme heterogeneity is not yet clear. The distribution of catalytic rates for E. coli β-galactosidase has been shown to change over time since induction [43]. Differences in Km and KI have been found between individual molecules. However, it has also been shown mathematically that in a large population, the effects of one molecule having a lower value for either of these parameters is largely canceled by other molecules having a different molecule having a larger value for either of these parameters [13,14]. With respect to enzymes where there is a very low load in a given cell, such canceling may not occur.
Heterogeneity in electrophoretic mobility represents a fundamental limit to the resolution of proteins using capillary zone electrophoresis separations [30]. Differences in mobility have been shown to result in a wider peak. Electrophoretic theory allows one to quantify the degree of differences in size, shape, and charge between the molecules that are required to result in the observed range in mobility. Theory suggests that the observed heterogeneity can be attributed to relatively small differences in structure [12,21,31].
8. Summary
Alkaline phosphatase was one of the first enzymes to be studied at the single-molecule level. Calf-intestinal alkaline phosphatase, as well as the enzyme from the bacteria Thermus thermophilis, has been found to be heterogeneous with respect to catalytic rate. Furthermore, the activation energy of catalysis and electrophoretic mobility have also been found to be heterogeneous for the calf-intestinal enzyme. Studies have suggested that these heterogeneous properties may be due to differences in glycosylation, limited proteolysis, or a mixture of active and inactive subunits within a given molecule. Changes in the distribution of single-molecule activities of a population have also been used as a means of studying the effect of heat treatment and changes in pH in changing the subpopulation distribution. Single-molecule analysis of alkaline phosphatase has also been used to determine if catalysis has an effect on the rate of enzyme diffusion. A second goal has been the development of single-molecule assays as an analytical tool. The distribution of activities has been used to determine the makeup of mixtures of different isozymes of a given phosphatase or of different phosphatase enzymes within a sample as a diagnostic tool.
Funding
This work was supported by a grant from the Natural Sciences and Engineering Council (Canada).
Conflicts of Interest
The author declares no conflict of interest.
References
- Xue, Q.; Yeung, E.S. Differences in the Chemical Reactivity of Individual Molecules of an Enzyme. Nature 1995, 373, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Arriaga, E.A.; Wong, J.C.Y.; Lu, H.; Dovichi, N.J. Studies on Single Alkaline phosphatase Molecules: Reaction Rate and Activation Energy of a Reaction Catalyzed by a Single Molecule and the Effect of Thermal Denaturation—The Death of an Enzyme. J. Am. Chem. Soc. 1996, 118, 5245–5253. [Google Scholar] [CrossRef]
- Craig, D.B.; Arriaga, E.A.; Wong, J.C.Y.; Lu, H.; Dovichi, N.J. Single Alkaline Phosphatase Molecule Assay by Capillary Electrophoresis Laser-Induced Fluorescence Detection. Tech. Protein Chem. 1997, 8, 121–131. [Google Scholar]
- Craig, D.B.; Arriaga, E.A.; Wong, J.C.Y.; Lu, H.; Dovichi, N.J. Life and Death of a Single Enzyme Molecule. Anal. Chem. 1998, 70, 39A–43A. [Google Scholar] [CrossRef]
- Shoemaker, G.K.; Juers, D.H.; Coombs, J.M.L.; Matthews, B.W.; Craig, D.B. Crystallization of β-Galactosidase Does Not Reduce the Range of Activity of Individual Molecules. Biochemistry 2003, 42, 1707–1710. [Google Scholar] [CrossRef]
- Gorris, H.-H.; Walt, D.R. Mechanistic Aspects of Horseradish Peroxidase Elucidated Through Single-Molecule Studies. J. Am. Chem. Soc. 2009, 131, 6277–6282. [Google Scholar] [CrossRef]
- Liebherr, R.B.; Renner, M.; Gorris, H.-H. A Single molecule Perspective on the Functional Diversity of In Vitro Evolved β-Glucuronidase. J. Am. Chem. Soc. 2014, 136, 5949–5955. [Google Scholar] [CrossRef]
- Bai, C.; Wang, C.; Xie, X.S.; Wolynes, P.G. Single Molecule Physics and Chemistry. Proc. Natl. Acad. Sci. USA 1999, 96, 11075–11076. [Google Scholar] [CrossRef]
- Lu, H.P.; Xun, L.; Xie, X.S. Single-Molecule Enzymatic Dynamics. Science 1998, 282, 1877–1882. [Google Scholar] [CrossRef]
- Gorris, H.-H.; Rissin, D.M.; Walt, D.R. Stochastic Inhibitor Release and binding from Single-Enzyme Molecules. Proc. Natl. Acad. Sci. USA 2007, 104, 17680–17685. [Google Scholar] [CrossRef]
- Chen, Z.; Shaw, A.; Wilson, H.; Woringer, M.; Darzacq, X.; Marqusee, S.; Wang, Q.; Bustamante, C. Single-Molecule Diffusometry Reveals No Catalysis-Induced Diffusion Enhancement of Alkaline Phosphatase as Proposed by FCS Experiments. Proc. Natl. Acad. Sci. USA 2020, 117, 21328–21335. [Google Scholar] [CrossRef] [PubMed]
- Nichols, E.R.; Shadabi, E.; Craig, D.B. Effect of Alteration of Translational Error Rate on Enzyme Microheterogeneity as Assessed by Variation in Single Molecule Electrophoretic Mobility and catalytic Activity. Biochem. Cell Biol. 2009, 87, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Haslam, A.M.; Coombs, J.M.L.; Nichols, E.R. Kinetic Studies on Unmodified Individual Escherichia coli β-Galactosidase Molecules in Free Solution. Biochem. Cell Biol. 2009, 88, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.J.; Hollett, J.W.; Craig, D.B. Determination of the Inhibitor Dissociation Constant of an Individual Unmodified Enzyme Molecule in Free Solution. Electrophoresis 2016, 37, 2217–2225. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Hall, T.; Goltz, D.M. E. coli β-Galactosidase is Heterogeneous with Respect to a Requirement for Magnesium. BioMetals 2000, 13, 223–229. [Google Scholar] [CrossRef]
- Craig, D.B.; Nichols, E.R. Continuous Flow Assay for the Simultaneous Measurement of the electrophoretic Mobility, Catalytic Activity and its Variation Over time of Individual Molecules of Escherichia coli β-Galactosidase. Electrophoresis 2008, 29, 4296–4303. [Google Scholar] [CrossRef]
- Sakamoto, S.; Komatsu, T.; Watanabe, R.; Zhang, Y.; Inoue, T.; Kawaguchi, M.; Nakagawa, H.; Ueno, T.; Okusaka, T.; Honda, K.; et al. Multiplexed Single-Molecule Enzyme Activity Analysis for Counting Disease-Related proteins in Biological Samples. Sci. Adv. 2020, 6, eaay0888. [Google Scholar] [CrossRef]
- Ueno, H.; Kato, M.; Minagawa, Y.; Hirose, Y.; Noji, H. Elucidation and Control of low and High Active Populations of Alkaline Phosphatase Molecules for Quantitative Digital Bioassay. Protein Sci. 2021, 30, 1628–1639. [Google Scholar] [CrossRef]
- Rotman, B. Measurement of activity of single molecules of β-D-galactosidase. Proc. Natl. Acad. Sci. USA 1961, 47, 1981–1991. [Google Scholar] [CrossRef]
- Xue, Q.; Yeung, E.S. Variability of Intracellular Lactate Dehydrogenase Isoenzymes in Single Human Erythrocytes. Anal. Chem. 1994, 66, 1175–1178. [Google Scholar] [CrossRef]
- Craig, D.B.; King, S.D.; Reinfelds, G.; Henderson, A.R.P.; Wood, T.E.H. Electrophoretic Mobility, Catalytic Rate, and Activation Energy of Catalysis of Single molecules of the Enzyme β-Glucuronidase from Escherichia coli. Int. J. Biol. Macromol. 2017, 96, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, L. Studies on Calf-Intestinal Alkaline phosphatase 1. Chromatographic Purification, Microheterogeneity and Some Other Properties of the Purified Enzyme. Biochim. Biophys. Acta 1961, 52, 36–48. [Google Scholar] [CrossRef]
- Saini, P.K.; Done, J. The diversity of Alkaline Phosphatase from Rat Intestine: Isolation and Purification of the Enzyme(s). Biochim. Biophys. Acta 1972, 258, 147–153. [Google Scholar] [CrossRef]
- Bradshaw, R.A.; Cancedda, F.; Ericcson, I.H.; Neumann, P.A.; Piccoli, S.P.; Schlesinger, M.J.; Shriefer, K.; Walsh, K.A. Amino Acid sequence of Escherichia coli Alkaline Phosphatase. Proc. Natl. Acad. Sci. USA 1981, 78, 3473–3477. [Google Scholar] [CrossRef]
- Schlesinger, M.J.; Andersen, L. Multiple Molecular Forms of the Alkaline Phosphatase of Escherichia coli. Ann. N Y Acad. Sci. 1968, 151, 159–170. [Google Scholar] [CrossRef]
- Polakowski, R.; Craig, D.B.; Skelley, A.; Dovichi, N.J. Single molecules of Highly Purified Bacterial Alkaline Phosphatase Have Identical Activity. J. Am. Chem. Soc. 2000, 122, 4853–4855. [Google Scholar] [CrossRef]
- Crawford, J.J.; Itzkow, F.; MacLean, J.; Craig, D.B. Conformational Change in Individual Enzyme Molecules. Biochem. Cell. Biol. 2015, 93, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Winzor, D.J.; Jones, S.; Harding, S.E. Determination of Protein Charge by Capillary Zone Electrophoresis. Anal. Biochem. 2004, 333, 225–229. [Google Scholar] [CrossRef]
- Winzor, D.J. Determination of the Net Charge (Valence) of a Protein: A Fundamental but Elusive Parameter. Anal. Biochem. 2004, 325, 1–20. [Google Scholar] [CrossRef]
- Nichols, E.R.; Craig, D.B. Measurement of the Differences in Electrophoretic Mobility of Individual Molecules of E. coli β-Galactosidase Provides Insight Into Structural Differences Which Underlie Enzyme Microheterogeneity. Electrophoresis 2008, 29, 1–13. [Google Scholar] [CrossRef]
- Craig, D.B.; Malhi, S.; Ahmad, B.; Breckman, K.; Patal, A. Electrophoretic Mobility of Individual Molecules of Alkaline Phosphatase. Biochem. Cell Biol. 2022, 100, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Hoffstatter-Kuhn, S.; Paulus, A.; Gassmann, E.; Widmer, H.M. Influence of Borate Complexation on the Electrophoretic Behaviour of Carbohydrates in Capillary Electrophoresis. Anal. Chem. 1991, 63, 1541–1547. [Google Scholar] [CrossRef]
- Dyck, A.C.; Craig, D.B. Individual Molecules of Thermostable Alkaline Phosphatase Support Different Catalytic Rates at Room Temperature. Luminescence 2002, 17, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Hanlon-Dearman, F.; Beaudry, S.; Shek, K.; King, S.D. Comparison of the Single Molecule Activity Distribution of Recombinant and Non-Recombinant Bovine Intestinal Alkaline Phosphatase. Protein Expr. Purif. 2015, 114, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Nichols, E.R.; Craig, D.B. Single Molecule Assays Reveal Differences Between In Vitro and In Vivo Synthesized β-Galactosidase. Protein J. 2008, 27, 376–383. [Google Scholar] [CrossRef]
- Gilboa, T.; Ogata, A.F.; Walt, D.R. Single-Molecule Enzymology for Diagnostics: Profiling Alkaline Phosphatase Activity in Clinical Samples. ChemBioChem 2022, 23, e202100358. [Google Scholar] [CrossRef]
- Rissin, D.M.; Gorris, H.-H.; Walt, D.R. Distinct and Long-Lived Activity States of Single Enzyme Molecules. J. Am. Chem. Soc. 2008, 130, 5349–5353. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Minigawa, Y.; Noji, H.; Tabata, K.V. Multidimensional Digital Bioassay Platform Based on an Air-Sealed Femtoliter Reactor Array Device. Anal. Chem. 2021, 93, 5494–5502. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, X.; Walt, D.R. Single-Molecule Analysis Determines Isozymes of Human Alkaline Phosphatase in Serum. Agnew. Chem. Int. Ed. Engl. 2020, 59, 18010–18015. [Google Scholar] [CrossRef]
- Gilboa, T.; Ogata, A.F.; Reilly, C.B.; Walt, D.R. Single-Molecules Studies Reveal Method for Tuning the heterogeneous Activity of Alkaline Phosphatase. Biophys. J. 2022, 121, 2027–2034. [Google Scholar] [CrossRef]
- Obayashi, Y.; Iino, R.; Noji, H. A Single-Molecule Digital Enzyme assay Using Alkaline Phosphatase with a Cumarin-Based Fluorogenic Substrate. Analyst 2015, 140, 5065–5073. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Schwab, T.; Sterner, R. Random mutagenesis suggests that sequence errors are not a major cause of variation in the activity of individual molecules of β-galactosidase. Biochem. Cell. Biol. 2012, 90, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Hall, T. Newly induced β-galactosidase molecules have a higher activity than the basally expressed enzyme. J. Clin. Laser Med. Surg. 2000, 18, 209–214. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).