The Interaction of Wildfire with Post-Fire Herbivory on Arid and Semi-Arid U.S. Rangelands: A Review

 ,
, {kind=link}
{kind=link}
Abstract
1. Introduction
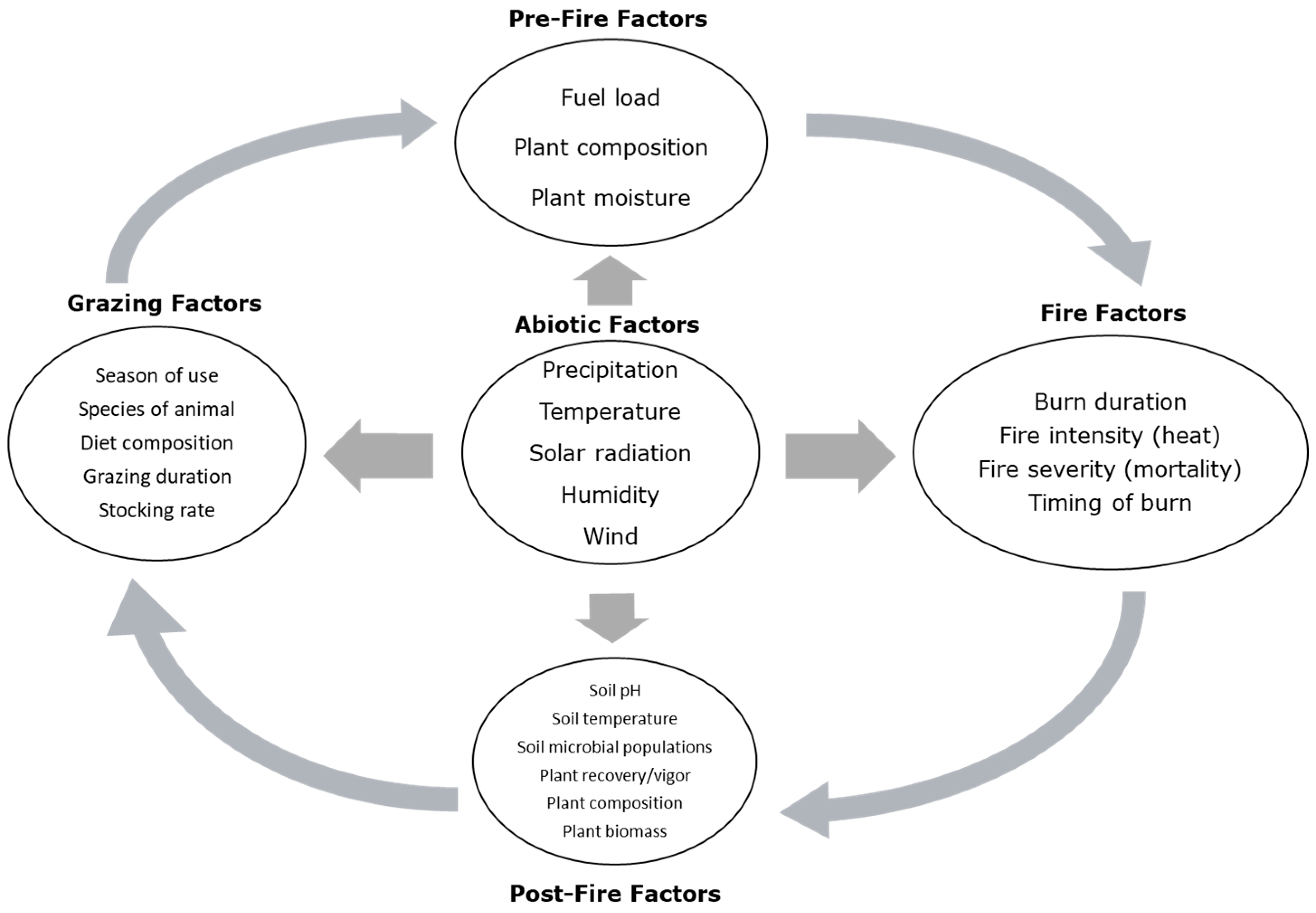
2. Knowledge Gap
3. History of Fire and Rangelands
4. Climate and Fire
5. Current Recommendations
6. Ecological Impacts and Management of Post-Fire Rangelands
7. Herbivores on Post-Fire Rangelands
8. Statement of the Problem
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- The Editors of Encyclopaedia Britannica. “Rangeland”. Encyclopedia Britannica, 11 February 2024. Available online: https://www.britannica.com/science/rangeland (accessed on 8 July 2024).
- Holechek, J.L.; Pieper, R.; Herbel, C. Rangeland Management Principles and Practices, 6th ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 2011; p. 07632. [Google Scholar]
- NRCS. Range Resources. Available online: https://www.nrcs.usda.gov/conservation-basics/natural-resource-concerns/land/range-pasture/range-resources#:~:text=Rangelands%20include%20grasslands%2C%20savannas%2C%20shrublands,totaling%20about%20770%20million%20acres (accessed on 1 October 2023).
- United States Department of Agriculture. Grazing Statistical Summary FY2020; USFS: Washington, DC, USA; USDA: Washington, DC, USA, 2021.
- Bureau of Land Management (BLM). Public Land Statistics 2021; Bureau of Land Management (BLM): Washington, DC, USA, 2022.
- U.S. Department of Agriculture (USDA). 2017 Census of Agriculture; U.S. Department of Agriculture (USDA): Washington, DC, USA, 2019.
- Maher, A.T.; Quintana Ashwell, N.E.; Maczko, K.A.; Taylor, D.T.; Tanaka, J.A.; Reeves, M.C. An economic valuation of federal and private grazing land ecosystem services supported by beef cattle ranching in the United States. Transl. Anim. Sci. 2021, 5, txab054. [Google Scholar] [CrossRef]
- Krausman, P.R. Rangeland Wildlife; Society for Range Management: Denver, CO, USA, 1996. [Google Scholar]
- DelCurto, T.; Wyffels, S.A.; Vavra, M.; Wisdom, M.J.; Posbergh, C.J. Western Rangeland Livestock Production Systems and Grazing Management. In Rangeland Wildlife Ecology and Conservation; Springer International Publishing: Cham, Switzerland, 2023; pp. 75–106. [Google Scholar]
- Bowers, W. Wildlife Habitats in Managed Rangelands: The Great Basin of Southeastern Oregon: Native Trout; Pacific Northwest Forest and Range Experiment Station, US Department of Agriculture, Forest Service: Fort Valley, AZ, USA, 1979; Volume 84.
- Bixby, R.J.; Cooper, S.D.; Gresswell, R.E.; Brown, L.E.; Dahm, C.N.; Dwire, K.A. Fire effects on aquatic ecosystems: An assessment of the current state of the science. Freshw. Sci. 2015, 34, 1340–1350. [Google Scholar] [CrossRef]
- Williams, R.E.; Allred, H. Conservation, Development, and Use of the World’s Rangelands. Rangel. Ecol. Manag. J. Range Manag. Arch. 1968, 21, 355–360. [Google Scholar] [CrossRef]
- Pyne, S.J. Vestal fires and virgin lands: A reburn. In General Technical Report INT; United States Department of Agriculture Forest Service: Washington, DC, USA, 1995; pp. 15–21. [Google Scholar]
- McAdoo, J.K.; Schultz, B.W.; Swanson, S.R. Aboriginal Precedent for Active Management of Sagebrush-Perennial Grass Communities in the Great Basin. Rangel. Ecol. Manag. 2013, 66, 241–253. [Google Scholar] [CrossRef]
- Williams, G.W. Introduction to aboriginal fire use in North America. Fire Manag. Today 2000, 60, 8–12. [Google Scholar]
- Stewart, O.C. Forgotten Fires: Native Americans and the Transient Wilderness; University of Oklahoma Press: Norman, OK, USA, 2002. [Google Scholar]
- Egan, T.; Dean, R. The Big Burn: Teddy Roosevelt and the Fire that Saved America; Houghton Mifflin Harcourt: Boston, MA, USA, 2009. [Google Scholar]
- Lueck, D.; Yoder, J. The economic foundations of firefighting organizations and institutions. J. For. 2015, 113, 291–297. [Google Scholar] [CrossRef]
- Omi, P.N. Theory and practice of wildland fuels management. Curr. For. Rep. 2015, 1, 100–117. [Google Scholar] [CrossRef]
- Li, Z.; Angerer, J.; Wu, B. The impacts of wildfires of different burn severities on vegetation structure across the western United States rangelands. Sci. Total Environ. 2022, 845, 157214. [Google Scholar] [CrossRef]
- Wildfires and Acres. 2023. Available online: https://www.nifc.gov/fire-information/statistics/wildfires (accessed on 8 July 2024).
- Lindsey, R.; Dahlman, L. Climate Change: Global Temperature; NOAA Climate.gov: Silver Spring, MD, USA, 2020; p. 16. Available online: https://www.climate.gov/ (accessed on 8 July 2024).
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef]
- Li, Z.; Angerer, J.; Wu, X.B. Prefire Vegetation Structure of High Severity Wildfires in Nonherbaceous-Dominated Rangelands in the Western United States. Earth’s Future 2022, 10, e2021EF002624. [Google Scholar] [CrossRef]
- Dennison, P.E.; Brewer, S.C.; Arnold, J.D.; Moritz, M.A. Large wildfire trends in the western United States, 1984–2011. Geophys. Res. Lett. 2014, 41, 2928–2933. [Google Scholar] [CrossRef]
- Wuebbles, D.J.; Fahey, D.W.; Hibbard, K.A. Climate Science Special Report: Fourth National Climate Assessment; US Global Change Research Program: Washington, DC, USA, 2017; Volume 1.
- Office, C.B. Wildfires. 2022. Available online: https://www.cbo.gov/publication/58212 (accessed on 8 July 2024).
- National Centers for Environmental Information. Billion-Dollar Weather and Climate Disasters. 2023. Available online: https://www.ncei.noaa.gov/access/billions/ (accessed on 8 July 2024).
- Blaisdell, J.P. Managing Intermountain Rangelands: Sagebrush-Grass Ranges; US Department of Agriculture, Forest Service, Intermountain Forest and Range: Washington, DC, USA, 1982; Volume 134.
- Seefeldt, S.S.; Germino, M.; DiCristina, K. Prescribed fires in Artemisia tridentata ssp. vaseyana steppe have minor and transient effects on vegetation cover and composition. Appl. Veg. Sci. 2007, 10, 249–256. [Google Scholar] [CrossRef]
- Bates, J.D.; Rhodes, E.C.; Davies, K.W.; Sharp, R. Postfire Succession in Big Sagebrush Steppe with Livestock Grazing. Rangel. Ecol. Manag. 2009, 62, 98–110. [Google Scholar] [CrossRef]
- Bates, J.D.; Davies, K.W. Cattle Grazing and Vegetation Succession on Burned Sagebrush Steppe. Rangel. Ecol. Manag. 2014, 67, 412–422. [Google Scholar] [CrossRef]
- Li, Z.; Angerer, J.; Wu, X.B. Temporal Patterns of Large Wildfires and Their Burn Severity in Rangelands of Western United States. Geophys. Res. Lett. 2021, 48, e2020GL091636. [Google Scholar] [CrossRef]
- Jirik, S.; Bunting, S.C. Postfire defoliation response of Agropyron spicatum and Sitanion hystrix. Int. J. Wildland Fire 1994, 4, 77–82. [Google Scholar] [CrossRef]
- West, N.E.; Yorks, T.P. Vegetation Responses following Wildfire on Grazed and Ungrazed Sagebrush Semi-Desert. J. Range Manag. 2002, 55, 171–181. [Google Scholar] [CrossRef]
- Blaisdell, J.P.; Pechanec, J.F. Effects of Herbage Removal at Various Dates on Vigor of Bluebunch Wheatgrass and Arrowleaf Balsamroot. Ecology 1949, 30, 298–305. [Google Scholar] [CrossRef]
- Gates, E.A.; Vermeire, L.T.; Marlow, C.B.; Waterman, R.C. Reconsidering rest following fire: Northern mixed-grass prairie is resilient to grazing following spring wildfire. Agric. Ecosyst. Environ. 2017, 237, 258–264. [Google Scholar] [CrossRef]
- Davies, K.W.; Bates, J.D.; Svejcar, T.J.; Boyd, C.S. Effects of Long-Term Livestock Grazing on Fuel Characteristics in Rangelands: An Example From the Sagebrush Steppe. Rangel. Ecol. Manag. 2010, 63, 662–669. [Google Scholar] [CrossRef]
- Vermeire, L.T.; Crowder, J.L.; Wester, D.B. Semiarid Rangeland Is Resilient to Summer Fire and Postfire Grazing Utilization. Rangel. Ecol. Manag. 2014, 67, 52–60. [Google Scholar] [CrossRef]
- Vermeire, L.T.; Crowder, J.L.; Wester, D.B. Plant Community and Soil Environment Response to Summer Fire in the Northern Great Plains. Rangel. Ecol. Manag. 2011, 64, 37–46. [Google Scholar] [CrossRef]
- Donovan, V.M.; Twidwell, D.; Uden, D.R.; Tadesse, T.; Wardlow, B.D.; Bielski, C.H.; Jones, M.O.; Allred, B.W.; Naugle, D.E.; Allen, C.R. Resilience to Large, “Catastrophic” Wildfires in North America’s Grassland Biome. Earths Future 2020, 8, e2020EF001487. [Google Scholar] [CrossRef]
- Gleason, H.A. The Vegetational History of the Middle West. Ann. Assoc. Am. Geogr. 1922, 12, 39–85. [Google Scholar] [CrossRef]
- Anderson, R.C. Evolution and origin of the Central Grassland of North America: Climate, fire, and mammalian grazers. J. Torrey Bot. Soc. 2006, 133, 626–647. [Google Scholar] [CrossRef]
- Stebbins, G.L. Coevolution of grasses and herbivores. Ann. Mo. Bot. Gard. 1981, 68, 75–86. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Engle, D.M.; Kerby, J.A.Y.; Hamilton, R. Pyric Herbivory: Rewilding Landscapes through the Recoupling of Fire and Grazing. Conserv. Biol. 2009, 23, 588–598. [Google Scholar] [CrossRef]
- Bates, J.D.; Boyd, C.S.; Davies, K.W. Davies, Longer-term post-fire succession on Wyoming big sagebrush steppe. Int. J. Wildland Fire 2020, 29, 229–239. [Google Scholar] [CrossRef]
- Conrad, C.E.; Poulton, C.E. Effect of a wildfire on Idaho fescue and bluebunch wheatgrass. Rangel. Ecol. Manag. J. Range Manag. Arch. 1966, 19, 138–141. [Google Scholar] [CrossRef]
- Taylor, M.H.; Rollins, K.; Kobayashi, M.; Tausch, R.J. The economics of fuel management: Wildfire, invasive plants, and the dynamics of sagebrush rangelands in the western United States. J. Environ. Manag. 2013, 126, 157–173. [Google Scholar] [CrossRef]
- Fusco, E.J.; Finn, J.T.; Balch, J.K.; Nagy, R.C.; Bradley, B.A. Invasive grasses increase fire occurrence and frequency across US ecoregions. Proc. Natl. Acad. Sci. USA 2019, 116, 23594–23599. [Google Scholar] [CrossRef] [PubMed]
- McIver, J.; Brunson, M.W.; Bunting, S.; Chambers, J.C.; Devoe, N.; Doescher, P.; Grace, J.; Johnson, D.; Knick, S.; Miller, R.F.; et al. The Sagebrush Steppe Treatment Evaluation Project (SageSTEP): A Test of State-and-Transition Theory; United States Department of Agriculture: Washington, DC, USA, 2010.
- Briggs, J.M.; Knapp, A.K.; Blair, J.M.; Heisler, J.L.; Hoch, G.A.; Lett, M.S.; McCarron, J.K. An ecosystem in transition: Causes and consequences of the conversion of mesic grassland to shrubland. BioScience 2005, 55, 243–254. [Google Scholar] [CrossRef]
- Prendergast-Miller, M.T.; De Menezes, A.B.; Macdonald, L.M.; Toscas, P.; Bissett, A.; Baker, G.; Farrell, M.; Richardson, A.E.; Wark, T.; Thrall, P.H. Wildfire impact: Natural experiment reveals differential short-term changes in soil microbial communities. Soil Biol. Biochem. 2017, 109, 1–13. [Google Scholar] [CrossRef]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- McGinty, A.; Smeins, F.E.; Merrill, L.B. Influence of spring burning on cattle diets and performance on the Edwards Plateau Rangeland. Rangel. Ecol. Manag. J. Range Manag. Arch. 1983, 36, 175–178. [Google Scholar]
- Allred, B.W.; Fuhlendorf, S.D.; Engle, D.M.; Elmore, R.D. Ungulate preference for burned patches reveals strength of fire–grazing interaction. Ecol. Evol. 2011, 1, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.; Martin, B.; Dreitz, V.J.; Allred, B.W. Grazing Preferences and Vegetation Feedbacks of the Fire-Grazing Interaction in the Northern Great Plains. Rangel. Ecol. Manag. 2018, 71, 45–52. [Google Scholar] [CrossRef]
- Vermeire, L.T.; Mitchell, R.B.; Fuhlendorf, S.D.; Gillen, R.L. Patch burning effects on grazing distribution. J. Range Manag. 2004, 57, 248–252. [Google Scholar] [CrossRef]
- Long, R.A.; Rachlow, J.L.; Kie, J.G. Effects of season and scale on response of elk and mule deer to habitat manipulation. J. Wildl. Manag. 2008, 72, 1133–1142. [Google Scholar] [CrossRef]
- Cruz, R.; Ganskopp, D. Seasonal preferences of steers for prominent northern Great Basin grasses. Rangel. Ecol. Manag. J. Range Manag. Arch. 1998, 51, 557–565. [Google Scholar] [CrossRef]
- Davies, K.W.; Bates, J.D.; Miller, R.F. Short-term effects of burning Wyoming big sagebrush steppe in southeast Oregon. Rangel. Ecol. Manag. 2007, 60, 515–522. [Google Scholar] [CrossRef]
- Davies, K.W.; Bates, J.D.; Perryman, B.; Arispe, S. Fall-Winter Grazing After Fire in Annual Grass-Invaded Sagebrush Steppe Reduced Annuals and Increased a Native Bunchgrass. Rangel. Ecol. Manag. 2021, 77, 1–8. [Google Scholar] [CrossRef]
- Launchbaugh, K.; Brammer, B.; Brooks, M.L.; Bunting, S.C.; Clark, P.; Davison, J.; Fleming, M.; Kay, R.; Pellant, M.; Pyke, D.A. Interactions among livestock grazing, vegetation type, and fire behavior in the Murphy Wildland Fire Complex in Idaho and Nevada, July 2007. In US Geological Survey Open-File Report; US Geological Survey: Reston, VA, USA, 2008; 42p. [Google Scholar]
- Roselle, L.; Seefeldt, S.S.; Launchbaugh, K. Delaying sheep grazing after wildfire in sagebrush steppe may not affect vegetation recovery. Int. J. Wildland Fire 2010, 19, 115–122. [Google Scholar] [CrossRef]
- Mueggler, W.F. Rate and pattern of vigor recovery in Idaho fescue and bluebunch wheatgrass. J. Range Manag. 1975, 28, 198–204. [Google Scholar] [CrossRef]
- McCarty, E.; Price, R. Growth and Carbohydrate Content of Important Mountain Forage Plants in Central Utah as Affected by Clipping and Grazing; Technical Bulletin; United States Department of Agriculture, Ed.; United States Department of Agriculture: Washington, DC, USA, 1942.
- Bunting, S.C.; Robberecht, R.; Defosse, G. Length and timing of grazing on postburn productivity of two bunchgrasses in an Idaho experimental range. Int. J. Wildland Fire 1998, 8, 15–20. [Google Scholar] [CrossRef]
- Stavi, I.; Barkai, D.; Knoll, Y.M.; Zaady, E. Livestock grazing impact on soil wettability and erosion risk in post-fire agricultural lands. Sci. Total Environ. 2016, 573, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Damiran, D.; DelCurto, T.; Findholt, S.L.; Johnson, B.K.; Vavra, M. The Effects of Previous Grazing on the Subsequent Nutrient Supply of Ungulates Grazing Late-summer Mixed-Conifer Rangelands. Sustain. Agric. Res. 2019, 8, 13–27. [Google Scholar] [CrossRef]
- Plumb, G.E.; Dodd, J.L. Foraging ecology of bison and cattle on a mixed prairie: Implications for natural area management. Ecol. Appl. 1993, 3, 631–643. [Google Scholar] [CrossRef]
- Towne, E.G.; Hartnett, D.C.; Cochran, R.C. Vegetation trends in tallgrass prairie from bison and cattle grazing. Ecol. Appl. 2005, 15, 1550–1559. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kluth, J.; Wyffels, S.; Eberly, J.; Vermeire, L.; Marlow, C.; DelCurto, T. The Interaction of Wildfire with Post-Fire Herbivory on Arid and Semi-Arid U.S. Rangelands: A Review. Grasses 2024, 3, 143-153. https://doi.org/10.3390/grasses3030010
Kluth J, Wyffels S, Eberly J, Vermeire L, Marlow C, DelCurto T. The Interaction of Wildfire with Post-Fire Herbivory on Arid and Semi-Arid U.S. Rangelands: A Review. Grasses. 2024; 3(3):143-153. https://doi.org/10.3390/grasses3030010
Chicago/Turabian StyleKluth, Janessa, Samuel Wyffels, Jed Eberly, Lance Vermeire, Clayton Marlow, and Timothy DelCurto. 2024. "The Interaction of Wildfire with Post-Fire Herbivory on Arid and Semi-Arid U.S. Rangelands: A Review" Grasses 3, no. 3: 143-153. https://doi.org/10.3390/grasses3030010
APA StyleKluth, J., Wyffels, S., Eberly, J., Vermeire, L., Marlow, C., & DelCurto, T. (2024). The Interaction of Wildfire with Post-Fire Herbivory on Arid and Semi-Arid U.S. Rangelands: A Review. Grasses, 3(3), 143-153. https://doi.org/10.3390/grasses3030010