

Composition, Distribution, and Biodiversity of Zooplanktons in Tropical Lentic Ecosystems with Different Environmental Conditions


 , and
, and
Abstract
1. Introduction
2. Material and Methods
2.1. Study Sites
2.2. Field Sampling and Laboratory Analyses
2.3. Data Analyses
3. Results
3.1. Zooplankton Species Composition, Abundance, and Diversity
3.2. Physicochemical Parameters of Studied Sites
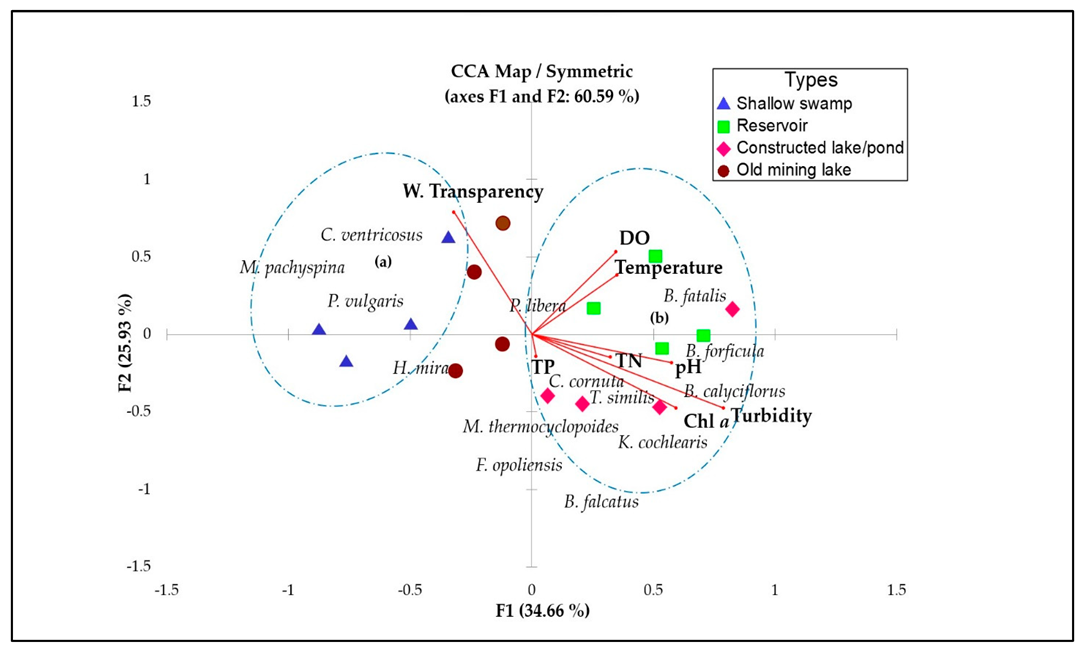
3.3. Relationship between Zooplankton and Physicochemical Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bakhtiyar, Y.; Arafat, M.Y.; Andrabi, S.; Tak, H.I. Zooplankton: The significant ecosystem service provider in aquatic environment. In Bioremediation and Biotechnology, Vol 3: Persistent and Recalcitrant Toxic Substances, 1st ed.; Bhat, R.A., Hakeem, K.R., Al-Saud, N.B.S., Eds.; Springer: Cham, Switzerland, 2020; Volume 3, pp. 227–244. [Google Scholar]
- Lomartire, S.; Marques, J.C.; Gonçalves, A.M. The key role of zooplankton in ecosystem services: A perspective of interaction between zooplankton and fish recruitment. Ecol. Indic. 2021, 129, 107867. [Google Scholar] [CrossRef]
- Almeida, R.; Formigo, N.E.; Sousa-Pinto, I.; Antunes, S.C. Contribution of zooplankton as a biological element in the assessment of reservoir water quality. Limnetica 2020, 39, 245–261. [Google Scholar] [CrossRef]
- Muñoz-Colmenares, M.E.; Soria, J.M.; Vicente, E. Can zooplankton species be used as indicators of trophic status and ecological potential of reservoirs? Aquat. Ecol. 2021, 55, 1143–1156. [Google Scholar] [CrossRef]
- Rashid, H.; Prakash, M.M. Zooplanktons as bioindicators of water pollution from Vikram Tearth Sarovar Ujjain (MP). J. Pharm. Negat. 2022, 13, 888–895. [Google Scholar]
- Pinto, I.; Nogueira, S.; Rodrigues, S.; Formigo, N.; Antunes, S.C. Can zooplankton add value to monitoring water quality? A case study of a meso/eutrophic Portuguese Reservoir. Water 2023, 15, 1678. [Google Scholar] [CrossRef]
- Umi, W.A.D.; Yusoff, F.M.; Aris, A.Z.; Sharip, Z. Rotifer community structure in tropical lakes with different environmental characteristics related to ecosystem health. J. Environ. Biol. 2018, 39, 795–807. [Google Scholar] [CrossRef]
- Umi, W.A.D.; Yusoff, F.M.; Aris, A.Z.; Sharip, Z.; Sinev, A.Y. Planktonic microcrustacean community structure varies with trophic status and environmental variables in tropical shallow lakes in Malaysia. Diversity 2020, 12, 322. [Google Scholar] [CrossRef]
- Qin, S.; Yang, T.; Yu, B.; Zhang, L.; Gu, L.; Sun, Y.; Yang, Z. The stress effect of atrazine on the inducible defense traits of Daphnia pulex in response to fish predation risk: Evidences from morphology, life history traits, and expression of the defense-related genes. Environ. Pollut. 2022, 311, 119965. [Google Scholar] [CrossRef]
- Mao, Z.; Cao, Y.; Gu, X.; Zeng, Q.; Chen, H.; Jeppesen, E. Response of zooplankton to nutrient reduction and enhanced fish predation in a shallow eutrophic lake. Ecol. Appl. 2023, 33, e2750. [Google Scholar] [CrossRef]
- Bomfim, F.F.; Melao, M.G.; Gebara, R.C.; Lansac-Toha, F.A. Warming alters the metabolic rates and life-history parameters of Ceriodaphnia silvestrii (Cladocera). An. Acad. Bras. Cienc. 2022, 94, e20200604. [Google Scholar] [CrossRef]
- Khalifa, U.; Ebenezer, V.; Pierson, J.J. Elevated temperature and low pH affect the development, reproduction, and feeding preference of the tropical cyclopoid copepod Oithona rigida. Int. J. Environ. Stud. 2022, 80, 1704–1720. [Google Scholar] [CrossRef]
- Xue, W.; Jin, J.; Zhang, F.; Chen, H.; Yang, D.; Zhang, Y.; Wei, W. Bosmina fatalis adapting to ammonia through oxidative stress and ribosome increase. J. Freshw. Ecol. 2022, 37, 117–129. [Google Scholar] [CrossRef]
- Le Quesne, K.S.; Özkundakci, D.; Duggan, I.C. Life on the farm: Are zooplankton communities in natural ponds and constructed dams the same? Mar. Freshw. Res. 2020, 72, 330–340. [Google Scholar] [CrossRef]
- Cabral, C.R.; Diniz, L.P.; da Silva, A.J.; Fonseca, G.; Carneiro, L.S.; de Melo Junior, M.; Caliman, A. Zooplankton species distribution, richness and composition across tropical shallow lakes: A large scale assessment by biome, lake origin, and lake habitat. Ann. Limnol. 2020, 56, 25. [Google Scholar] [CrossRef]
- Eskinazi-Sant’Anna, E.M.; Santos, G.D.S.; Alves, N.J.D.S.; Brito, L.A.F.; Leite, M.G.P. The relative importance of regional and local factors in shaping zooplankton diversity in high-altitude tropical shallow lakes. J. Freshw. Ecol. 2020, 35, 203–221. [Google Scholar] [CrossRef]
- Geng, Y.; Li, M.; Yu, R.; Sun, H.; Zhang, L.; Sun, L.; Xu, J. Response of planktonic diversity and stability to environmental drivers in a shallow eutrophic lake. Ecol. Indic. 2022, 144, 109560. [Google Scholar] [CrossRef]
- Simões, N.R.; Dias, J.D.; Meerhoff, M.; Lansac-Toha, F.A.; Bini, L.M.; Bonecker, C.C. Drivers of zooplankton beta diversity in natural shallow lakes and artificial reservoirs in the Neotropics. Hydrobiologia 2022, 849, 3705–3717. [Google Scholar] [CrossRef]
- Shen, J.; Qin, G.; Yu, R.; Zhao, Y.; Yang, J.; An, S.; Wan, Y. Urbanization has changed the distribution pattern of zooplankton species diversity and the structure of functional groups. Ecol. Indic. 2021, 120, 106944. [Google Scholar] [CrossRef]
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Zahoor, I.; Mushtaq, A. Water pollution from agricultural activities: A critical global review. Int. J. Chem. Biochem. Sci. 2023, 23, 164–176. [Google Scholar]
- Razak, S.B.A.; Sharip, Z. Spatio-temporal variation of zooplankton community structure in tropical urban waterbodies along trophic and urban gradients. Ecol. Process. 2019, 8, 44. [Google Scholar] [CrossRef]
- Herath, S.S.; Satoh, S. Environmental impacts of nitrogen and phosphorus from aquaculture. In Feed and Feeding Practices in Aquaculture, 2nd ed.; Davis, D.A., Ed.; Woodhead Publishing: Sawsto, UK, 2022; pp. 427–444. [Google Scholar]
- Wenzel, A.; Vrede, T.; Jansson, M.; Bergström, A.K. Daphnia performance on diets containing different combinations of high-quality algae, heterotrophic bacteria, and allochthonous particulate organic matter. Freshw. Biol. 2021, 66, 157–168. [Google Scholar] [CrossRef]
- Gilbert, J.J. Food niches of planktonic rotifers: Diversification and implications. Limnol. Oceanogr. 2022, 67, 2218–2251. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, H.; Liu, X.; Li, Y.; Boersma, M.; Zhou, L.; Chen, F. The comprehensive effect of natural food quality and quantity on growth rate of herbivore consumers. Ecol. Indic. 2023, 156, 111129. [Google Scholar] [CrossRef]
- Derevenskaia, O.I.; Borisova, N.I.; Unkovskaia, E.N. Zooplankton indices in the evaluation of the ecological state of the eutrophic lake (Case study: Karasikha Lake, Russia). Casp. J. Environ. Sci. 2021, 19, 701–708. [Google Scholar]
- Alprol, A.E.; Heneash, A.M.; Soliman, A.M.; Ashour, M.; Alsanie, W.F.; Gaber, A.; Mansour, A.T. Assessment of water quality, eutrophication, and zooplankton community in Lake Burullus, Egypt. Diversity 2021, 13, 268. [Google Scholar] [CrossRef]
- Amorim, C.A.; do Nascimento Moura, A. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef]
- Benayache, N.Y.; Nguyen-Quang, T.; Hushchyna, K.; McLellan, K.; Afri-Mehennaoui, F.Z.; Bouaïcha, N. An overview of cyanobacteria harmful algal bloom (CyanoHAB) issues in freshwater ecosystems. In Limnology: Some New Aspects of Inland Water Ecology; Gokce, D., Ed.; IntechOpen: London, UK, 2019; pp. 13–37. [Google Scholar]
- Han, T.; Kong, M.; Tang, C.; Xu, X.; Zhu, Y.; Gao, Y.; Li, W. Influence of algal blooms on the efficacy of La/Al-based phoslock in the control of phosphorus release from sediment in shallow lakes: A microcosm study. J. Soils Sediments 2021, 21, 3405–3414. [Google Scholar] [CrossRef]
- Bizani, M.; Bornman, T.G.; Campbell, E.E.; Perissinotto, R.; Deyzel, S.H.P. Mesozooplankton community responses to a large-scale harmful algal bloom induced by the non-indigenous dinoflagellate Lingulodinium polyedra. Sci. Total Environ. 2023, 860, 161030. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Bownik, A. Cyanobacterial anabaenopeptin-B, microcystins and their mixture cause toxic effects on the behavior of the freshwater crustacean Daphnia magna (Cladocera). Toxicon 2021, 198, 1–18. [Google Scholar] [CrossRef]
- Pontin, R. A Key to the Freshwater Planktonic and Semi-Planktonic Rotifera of the British Isles; Freshwater Biological Association: Ambleside, UK, 1978; Volume 38, p. 178. [Google Scholar]
- Idris, B.A.G. Freshwater Zooplankton of Malaysia (Crustacea: Cladocera); University Pertanian Malaysia Press: Serdang, Malaysia, 1983; p. 153. [Google Scholar]
- Shiel, R.J. A Guide to Identification of Rotifers, Cladocerans and Copepods from Australian Inland Waters; Identification Guide Series No. 3; Shiel, J., Albury, N.S.W., Eds.; Co-Operative Research Centre for Freshwater Ecology: Canberra, ACT, Australia, 1995; p. 144. [Google Scholar]
- Segers, H. Global diversity of rotifers (Rotifera) in freshwater. Hydrobiologia 2008, 595, 49–59. [Google Scholar] [CrossRef]
- Sa-Ardrit, P.; Pholpunthin, P.; Segers, H. A checklist of the freshwater rotifer fauna of Thailand (Rotifera, Monogononta, Bdelloidea). J. Limnol. 2013, 72, 361–375. [Google Scholar] [CrossRef][Green Version]
- Phan, D.D.; Nguyen, V.K.; Le, T.N.; Ngoc, T.D.; Ho, T.H. Identification Handbook of Freshwater Zooplankton of the Mekong River and Its Tributaries; Mekong River Commission: Vientiane, Laos, 2015; p. 207. [Google Scholar]
- Ismail, A.H.; Lim, C.C.; Wan Omar, W.M. Evaluation of spatial and temporal variations in zooplankton community structure with reference to water quality in Teluk Bahang Reservoir, Malaysia. Trop. Ecol. 2019, 60, 186–198. [Google Scholar] [CrossRef]
- Toruan, R.L. Zooplankton diversity in Lake Tondano, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2021, 744, 012092. [Google Scholar] [CrossRef]
- Tang, C.; Yi, Y.; Yang, Z.; Zhou, Y.; Zerizghi, T.; Wang, X.; Duan, P. Planktonic indicators of trophic states for a shallow lake (Baiyangdian Lake, China). Limnologica 2019, 78, 125712. [Google Scholar] [CrossRef]
- Makwinja, R.; Mengistou, S.; Kaunda, E.; Alamirew, T. Spatial distribution of zooplankton in response to ecological dynamics in tropical shallow lake: Insight from Lake Malombe, Malawi. J. Freshw. Ecol. 2021, 36, 127–148. [Google Scholar] [CrossRef]
- Beyene, G.; Kifle, D.; Fetahi, T. Spatial distribution of zooplankton in relation to some selected physicochemical water quality parameters of Lake Hawassa, Ethiopia. Afr. J. Aquat. Sci. 2022, 47, 163–172. [Google Scholar] [CrossRef]
- Macedo, R.L.; Franco, A.C.S.; Klippel, G.; Oliveira, E.F.; Silva, L.H.S.; dos Santos, L.N.; Branco, C.W. Small in size but rather pervasive: The spread of the North American rotifer Kellicottia bostoniensis (Rousselet, 1908) through Neotropical basins. BioInvasions Rec. 2020, 9, 287–302. [Google Scholar] [CrossRef]
- Bonecker, C.C.; Diniz, L.P.; Braghin, L.D.S.M.; Mantovano, T.; da Silva, J.V.F.; de Fátima Bomfim, F.; Lansac-Tôha, F.A. Synergistic effects of natural and anthropogenic impacts on zooplankton diversity in a subtropical floodplain: A long-term study. Oecol. Aust. 2020, 24, 524–537. [Google Scholar] [CrossRef]
- Espinosa-Rodríguez, C.A.; Sarma, S.S.S.; Nandini, S. Zooplankton community changes in relation to different macrophyte species: Effects of Egeria densa removal. Ecohydrol. Hydrobiol. 2021, 21, 153–163. [Google Scholar] [CrossRef]
- Brito, M.T.D.S.; Heino, J.; Pozzobom, U.M.; Landeiro, V.L. Ecological uniqueness and species richness of zooplankton in subtropical floodplain lakes. Aquat. Sci. 2020, 82, 43. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. Responses of rotifer community to microhabitat changes caused by summer-concentrated rainfall in a shallow reservoir, South Korea. Diversity 2020, 12, 113. [Google Scholar] [CrossRef]
- Coelho, P.N.; Henry, R. Functional groups of microcrustaceans along a horizontal gradient in a neotropical lake colonized by macrophytes. Aquat. Sci. 2021, 83, 3. [Google Scholar] [CrossRef]
- Quirino, B.A.; Teixeira de Mello, F.; Deosti, S.; Bonecker, C.C.; Cardozo, A.L.P.; Yofukuji, K.Y.; Fugi, R. Interactions between a planktivorous fish and planktonic microcrustaceans mediated by the biomass of aquatic macrophytes. J. Plankton Res. 2021, 43, 46–60. [Google Scholar] [CrossRef]
- Celewicz-Gołdyn, S.; Kuczyńska-Kippen, N. Ecological value of macrophyte cover in creating habitat for microalgae (diatoms) and zooplankton (rotifers and crustaceans) in small field and forest water bodies. PLoS ONE 2017, 12, e0177317. [Google Scholar] [CrossRef] [PubMed]
- Moody, E.K.; Wilkinson, G.M. Functional shifts in lake zooplankton communities with hypereutrophication. Freshw. Biol. 2019, 64, 608–616. [Google Scholar] [CrossRef]
- Howarth, R.W.; Chan, F.; Swaney, D.P.; Marino, R.M.; Hayn, M. Role of external inputs of nutrients to aquatic ecosystems in determining prevalence of nitrogen vs. phosphorus limitation of net primary productivity. Biogeochemistry 2021, 154, 293–306. [Google Scholar] [CrossRef]
- Xiong, W.; Ni, P.; Chen, Y.; Gao, Y.; Li, S.; Zhan, A. Biological consequences of environmental pollution in running water ecosystems: A case study in zooplankton. Environ. Pollut. 2019, 252, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Cao, X.; Cui, L.; Lv, Q.; Chen, T. The influence of human interference on zooplankton and fungal diversity in Poyang Lake watershed in China. Diversity 2020, 12, 296. [Google Scholar] [CrossRef]
- Li, Y.; Chen, F. Are zooplankton useful indicators of water quality in subtropical lakes with high human impacts? Ecol. Indic. 2020, 113, 106167. [Google Scholar] [CrossRef]
- Doubek, J.P.; Campbell, K.L.; Doubek, K.M.; Hamre, K.D.; Lofton, M.E.; McClure, R.P.; Carey, C.C. The effects of hypolimnetic anoxia on the diel vertical migration of freshwater crustacean zooplankton. Ecosphere 2018, 9, e02332. [Google Scholar] [CrossRef]
- Keister, J.E.; Winans, A.K.; Herrmann, B. Zooplankton community response to seasonal hypoxia: A test of three hypotheses. Diversity 2020, 12, 21. [Google Scholar] [CrossRef]
- Walczyńska, A.; Sobczyk, M. Experimental evolution shows body size decrease in response to hypoxia, with a complex effect on plastic size response to temperature. Biol. Bull. 2022, 243, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.K.; Wei, J.H.; Welander, P.V. De novo cholesterol biosynthesis in the bacterial domain. bioRxiv 2022. [Google Scholar] [CrossRef]
- Mohd-Asharuddin, S.; Zayadi, N.; Rasit, W.; Othman, N. Water quality characteristics of Sembrong Dam Reservoir, Johor, Malaysia. IOP Conf. Ser. Mater. Sci. Eng. 2016, 136, 012058. [Google Scholar] [CrossRef]
- Hashim, S.I.N.S.; Talib, S.H.A.; Abustan, M.S. Nutrient correlation analysis between sediment and water at Sembrong Dam, Johor. Int. J. Adv. Eng. Technol. 2020, 11, 986–993. [Google Scholar]
- Zhang, S.; Lu, W.; Zhou, Z.; Chen, W. Spatial differences in zooplankton community structure between two fluvial lakes in the middle and lower reaches of the Yangtze River: Effects of land use patterns and physicochemical factors. Diversity 2022, 14, 908. [Google Scholar] [CrossRef]
- Sharip, Z.; Mohamad, M.F. Microbial contamination in urban tropical lentic waterbodies and ponds along urbanisation gradient. J. Trop. Agric. 2019, 42, 165–184. [Google Scholar]
- Krupa, E.; Romanova, S.; Berkinbaev, G.; Yakovleva, N.; Sadvakasov, E. Zooplankton as indicator of the ecological state of protected aquatic ecosystems (Lake Borovoe, Burabay National Nature Park, Northern Kazakhstan). Water 2020, 12, 2580. [Google Scholar] [CrossRef]
- Hamil, S.; Bouchelouche, D.; Arab, S.; Alili, M.; Baha, M.; Arab, A. The relationship between zooplankton community and environmental factors of Ghrib Dam in Algeria. Environ. Sci. Pollut. Res. 2021, 28, 46592–46602. [Google Scholar] [CrossRef]
- Bera, D.B. Using zooplankton community to assess water quality and trophic condition of Lake Saheb Bandh, Purulia, West Bengal, India. Int. J. Aquatic Sci. 2021, 12, 4471–4498. [Google Scholar]
- Apaydın Yağcı, M.; Yılmaz, M.; Akkan, T.; Yazıcıoğlu, O.; Yazıcı, R.; Yağcı, A.; Gülsoy, S. Structure of the zooplankton community in Sıddıklı Küçükboğaz Dam Lake, Türkiye: The importance of environmental factors. Iran. J. Sci. 2023, 47, 719–732. [Google Scholar] [CrossRef]
- Guermazi, W.; El-khateeb, M.; Abu-Dalo, M.; Sallemi, I.; Al-Rahahleh, B.; Rekik, A.; Annabi-Trabelsi, N. Assessment of the zooplankton community and water quality in an artificial freshwater lake from a semi-arid area (Irbid, Jordan). Water 2023, 15, 2796. [Google Scholar] [CrossRef]
- Meremo, W.T.; Reuben, O.; Wamalwa, Y.A.; Ndegwa, D.M. Changes in water quality parameters and their effect on zooplankton distribution in a shallow bay of Lake Victoria, Kenya. Int. J. Fish. Aquat. 2022, 10, 206–212. [Google Scholar] [CrossRef]
- Nieman, C.L.; Gray, S.M. Visual performance impaired by elevated sedimentary and algal turbidity in walleye Sander vitreus and emerald shiner Notropis atherinoides. J. Fish Biol. 2019, 95, 186–199. [Google Scholar] [CrossRef] [PubMed]
- García-Chicote, J.; Armengol, X.; Rojo, C. Zooplankton species as indicators of trophic state in reservoirs from Mediterranean river basins. Inland Waters 2019, 9, 113–123. [Google Scholar] [CrossRef]
- Rosińska, J.; Romanowicz-Brzozowska, W.; Kozak, A.; Gołdyn, R. Zooplankton changes during bottom-up and top-down control due to sustainable restoration in a shallow urban lake. Environ. Sci. Pollut. Res. 2019, 26, 19575–19587. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Q.; Wei, N.; Tang, C.; Sun, X.; Yang, Y. Biological indicators of ecological quality in typical urban river-lake ecosystems: The planktonic rotifer community and its response to environmental factors. Ecol. Indic. 2020, 112, 106127. [Google Scholar] [CrossRef]
- Kim, S.K.; Yun, J.H.; Joo, G.J.; Choi, J.Y. Hydrological characteristics and trophic status as dominant drivers of rotifer community composition in artificially created riverine wetlands. Animals 2022, 12, 461. [Google Scholar] [CrossRef]
- Kumari, S.; Lianthuamluaia, L.; Sarkar, U.K.; Puthiyottil, M.; Karnatak, G.; Meena, D.K.; Das, B.K. Environmental characterization of two ecologically distinct Gangetic oxbow lakes using zooplankton taxonomic indices through comparative approach for wetland monitoring. Wetlands 2023, 43, 14. [Google Scholar] [CrossRef]
- Zhang, H.; Hollander, J.; Hansson, L.A. Bi-directional plasticity: Rotifer prey adjust spine length to different predator regimes. Sci. Rep. 2017, 7, 10254. [Google Scholar] [CrossRef] [PubMed]
- Nandini, S.; Sarma, S.S.S. Experimental studies on zooplankton-toxic cyanobacteria interactions: A Review. Toxics 2023, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Qin, S.; Zhu, S.; Lu, N.; Sun, Y.; Zhang, L.; Yang, Z. Microcystis aeruginosa affects the inducible anti-predator responses of Ceriodaphnia cornuta. Environ. Pollut. 2020, 259, 113952. [Google Scholar] [CrossRef] [PubMed]
- Vilar, M.C.P.; da Costa Pena Rodrigues, T.F.; da Silva Ferrão-Filho, A.; de Oliveira e Azevedo, S.M.F. Grazer-induced chemical defense in a microcystin-producing Microcystis aeruginosa (Cyanobacteria) exposed to Daphnia gessneri infochemicals. J. Chem. Ecol. 2021, 47, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Gerasimova, T.N.; Pogozhev, P.I.; Sadchikov, A.P. Suppression of cyanobacterial blooms by zooplankton: Experiments in natural water bodies with the use of flow-through ecosystems. Russ. J. Gen. Chem. 2019, 89, 2840–2844. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.D.S.; da Silva, D.A.C. Saxitoxin-producing Raphidiopsis raciborskii (cyanobacteria) inhibits swimming and physiological parameters in Daphnia similis. Sci. Total Environ. 2020, 706, 135751. [Google Scholar] [CrossRef] [PubMed]
- Krztoń, W.; Kosiba, J.; Wilk-Woźniak, E. Features that matter: Studying how phytoplankton drives zooplankton community functional traits. Hydrobiologia 2022, 849, 2647–2662. [Google Scholar] [CrossRef]
- Das Candeias, D.A.; Moi, D.A.; Simões, N.R.; Azevedo, F.; Meerhoff, M.; Bonecker, C.C. High temperature, predation, nutrient, and food quality drive dominance of small-sized zooplankton in Neotropical lakes. Aquat. Sci. 2022, 84, 49. [Google Scholar] [CrossRef]
- Picapedra, P.H.S.; Fernandes, C.; Baumgartner, G.; Sanches, P.V. Zooplankton communities and their relationship with water quality in eight reservoirs from the midwestern and southeastern regions of Brazil. Braz. J. Biol. 2020, 81, 701–713. [Google Scholar] [CrossRef]
- Valencia-Vargas, M.A.; Nandini, S.; Sarma, S.S.S. Demographic characteristics of two freshwater cyclopoid copepods in Mexico, fed a plankton diet: The native Mesocyclops longisetus Thiébaud and the invasive Mesocyclops pehpeinsis Hu. Inland Waters 2020, 10, 128–136. [Google Scholar] [CrossRef]
- Ismail, A.H.; Abd Rahman, A.; Chin, L.C. The use of GIS to visualize spatial distribution of zooplankton in Teluk Bahang Reservoir, Penang, Malaysia. J. Environ. Sci. Manag. 2020, 23, 60–71. [Google Scholar] [CrossRef]
- Fajardo, L.J.; Lebeng, R.S.; Morales, M.L.; Reyes, A.T. Plankton abundance and diversity in Pantabangan Reservoir, Pantabangan, Nueva Ecija, Philippines. AACL Bioflux 2022, 15, 1541–1552. [Google Scholar]
- Kottage, C.D.; Patrick, A.E.S. Hydro-climatic consequences on zooplankton diversity and abundance in perennial Vavuniya reservoir and seasonal Thandikulam reservoir in Vavuniya, Sri Lanka. J. Basic Appl. Zool. 2023, 84, 7. [Google Scholar] [CrossRef]
- Ebesi, E.J.; Mohammed, Y.M.; Iloba, K.I.; Adamu, K.M.; Adama, S.B. Zooplankton community structure of a tropical lake in a Northcentral State, Nigeria. Ruhuna J. Sci. 2022, 13, 217–230. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Sites | Coordinate | Site Descriptions | Surface Area and Depth | Catchment Area | Role |
|---|---|---|---|---|---|---|
| Natural ecosystem | ||||||
| Natural lakes/swamps | ||||||
| S1. | Bera Lake, Pahang | 3.1325N, 102.6056E |
| Surface area: 5400 ha Depth: 3.5 m | Agricultural sites, logging, iron ore mining, rubber plantations, palm oil plantations, and villages. | Flood mitigation (natural wetland ecosystem due to its presence that can decrease the velocity, frequency, and level of floods) and recreational site (fishing, camping, boating). |
| S2. | Chini Lake, Pahang | 3.4351N, 102.9185E |
| Surface area: 200 ha Depth: 2.7 m | Agricultural sites, logging, iron ore mining, rubber plantations, and palm oil plantations. | Flood mitigation (natural wetland ecosystem due to its presence that can decrease the velocity, frequency, and level of floods) and recreational site (fishing, camping, boating). |
| S3. & S4. | Swamps in Selangor | 2.8599N 101.6214E 2.8591N 101.6262E |
| Surface area: 10 ha Depth: 0.5 m | Forest and tourist activities. | Natural wetland ecosystem due to its presence that can decrease the velocity, frequency, and level of floods. Recreational site (camping, jungle tracking, cycling, picnic). |
| Artificial ecosystem | ||||||
| Reservoirs | ||||||
| S5. | Chenderoh Lake, Perak | 4.9696N 100.9578E |
| Surface area: 2500 ha Depth: 4.8 m | Agricultural sites. | Hydroelectric power plant, flood mitigation, water supply, tourism. |
| S6. | Timah Tasoh Lake, Perlis | 6.5875N 100.2286E |
| Surface area: 1333 ha Depth: 3.5 m | Agricultural activities, recreational site, villages, and mountains. | Flood mitigation, water supply, irrigation, and tourism. |
| S7. | Kenyir Lake, Terengganu | 5.1394N 102.7904E |
| Surface area: 36,900 ha Depth: 75 m | Forest and recreational sites. | Hydroelectric power plant, flood mitigation, water supply, and tourism. |
| S8. | Sembrong Lake, Johor | 1.9845N 103.1919E |
| Surface area: 850 ha Depth: 5.1 m | Agricultural sites, husbandry areas, and villages. | Flood mitigation and water supply. |
| Constructed lakes\ponds | ||||||
| S9. | Putrajaya Lake, W.P. Putrajaya | 2.9494N 101.6936E |
| Surface area: 400 ha Depth: 6.6 m | Administrative and residential area. | Tourism and recreational water sports. |
| S10. | Sri Serdang Pond, Selangor | 3.0042N 101.7138E |
| Surface area: 1.8 ha Depth: 2.7 m | Residential area, commercial area, and restaurants. | Recreational sites (jogging, cycling). |
| S11. | UPM Pond, Selangor | 2.9874N 101.7127E |
| Surface area: 1.0 ha Depth: 1.5 m | Institution buildings and forest. | Recreational sites (jogging, picnic). |
| S12. | Cempaka Lake, Selangor | 2.9600N 101.7591E |
| Surface area: 6.4 ha Depth: 3.2 m | Residential area, commercial area, and restaurants. | Recreational activities, jogging, and cycling. |
| Old mining lakes | ||||||
| S13. | Puchong Lake, Selangor | 2.9541N 101.6094E |
| Surface area: 6.4 ha Depth: 3.2 m | Residential area. | Recreational site and fishing. |
| S14. | The Mines Lake, Selangor | 3.0341N 101.7127E |
| Surface area: 62.1 ha Depth: 100 m | Commercial area and residential area. | Tourism and recreational site. |
| S15. | Biru Kundang Lake, Selangor | 3.2516N 101.5251E |
| Surface area: 32 ha Depth: 5.2 m | Residential area. | Recreational site. |
| S16. | Taiping Lake, Perak | 4.8526N 100.7464E |
| Surface area: 64 ha Depth: 4.0 m m | Garden and residential area. | Tourism and recreational site. |
| Family | Species | Natural Ecosystem | Artificial Ecosystem | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Natural Shallow Swamps | Reservoirs | Constructed Lakes | Old Mining Lakes | ||||||
| Mean Densities | % | Mean Densities | % | Mean Densities | % | Mean Densities | % | ||
| Cladocerans | |||||||||
| Bosminidae | Bosmina fatalis | 0.0 ± 0.0 | 0.0 | 6.7 ± 1.3 | 2.2 | 26.1 ± 3.8 | 7.1 | 1.6 ± 0.5 | 0.7 |
| Bosminopsis africanus | 5.5 ± 2.7 | 2.8 | 71.2 ± 28.8 | 23.1 | 0.7 ± 0.2 | 0.2 | 0.0 ± 0.0 | 0.0 | |
| Chydoridae | Anthalona harti harti | 7.8 ± 1.3 | 3.9 | 1.1 ± 0.6 | 0.4 | 0.9 ± 0.4 | 0.2 | 6.8 ± 1.2 | 2.8 |
| Chydorus ventricosus | 30.0 ± 7.9 | 15.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Daphniidae | Ceriodaphnia cornuta | 2.9 ± 1.2 | 1.5 | 32.1 ± 14.2 | 10.4 | 54.9 ± 11.2 | 15.0 | 1.6 ± 0.1 | 0.7 |
| Simocephalus serrulatus | 2.4 ± 0.3 | 1.2 | 1.1 ± 0.1 | 0.4 | 0.8 ± 0.3 | 0.2 | 0.1 ± 0.0 | 0.0 | |
| Ilyocryptidae | Ilyocryptus spinifer | 3.7 ± 0.9 | 1.9 | 1.2 ± 0.9 | 0.4 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 |
| Macrothricidae | Macrothrix spinosa | 6.6 ± 3.2 | 3.4 | 0.6 ± 0.3 | 0.2 | 0.0 ± 0.0 | 0.0 | 1.9 ± 0.3 | 0.8 |
| Macrothrix triserialis | 0.0 ± 0.0 | 0.0 | 3.3 ± 1.2 | 1.1 | 2.5 ± 0.9 | 0.7 | 0.0 ± 0.0 | 0.0 | |
| Moinidae | Moina micrura | 2.5 ± 1.0 | 1.3 | 0.0 ± 0.0 | 0.0 | 0.1 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 |
| Sididae | Diaphanosoma excisum | 4.8 ± 0.9 | 2.4 | 1.2 ± 0.2 | 0.4 | 1.6 ± 2.3 | 0.4 | 0.0 ± 0.0 | 0.0 |
| Diaphanosoma sarsi | 0.2 ± 0.1 | 0.1 | 1.6 ± 0.6 | 0.5 | 3.2 ± 1.2 | 0.9 | 0.0 ± 0.0 | 0.0 | |
| Subtotal | 53.2 ± 12.7 | 27.0 | 120.1 ± 23.2 | 38.9 | 85.1 ± 17.9 | 23.2 | 11.9 ± 3.2 | 4.9 | |
| Copepods | |||||||||
| Copepodites | 3.2 ± 0.2 | 1.6 | 3.7 ± 0.1 | 1.2 | 7.8 ± 0.2 | 2.1 | 2.4 ± 0.0 | 1.0 | |
| Nauplii | 2.4 ± 0.0 | 1.2 | 19.6 ± 5.0 | 6.3 | 28.0 ± 3.7 | 7.7 | 3.7 ± 0.9 | 1.5 | |
| Cyclopidae | Eucyclops taiwanensis | 2.8 ± 1.5 | 1.4 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 |
| Mesocyclop affinis | 1.2 ± 0.6 | 0.6 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Mesocyclops francisci | 1.2 ± 0.6 | 0.6 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Mesocyclos ogunnus | 1.6 ± 0.5 | 0.8 | 1.2 ± 0.3 | 0.4 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Mesocyclos thermocyclopoides | 0.0 ± 0.0 | 0.0 | 16.6 ± 2.3 | 5.4 | 26.6 ± 2.7 | 7.3 | 2.6 ± 0.9 | 1.1 | |
| Microcyclops pachyspina | 12.5 ± 5.2 | 6.2 | 0.9 ± 0.1 | 0.3 | 0.6 ± 0.0 | 0.2 | 0.3 ± 0.0 | 0.1 | |
| Microcyclops sumatranus | 0.6 ± 0.1 | 0.3 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Thermocyclop crassus | 5.5 ± 2.7 | 2.7 | 3.7 ± 1.1 | 1.2 | 12.4 ± 1.7 | 3.4 | 0.0 ± 0.0 | 0.0 | |
| Thermocyclops decipiens | 0.2 ± 0.0 | 0.1 | 0.2 ± 0.0 | 0.1 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Diaptomidae | Mongolodiaptomus malaindosinensis | 3.7 ± 0.8 | 1.8 | 3.8 ± 0.5 | 1.2 | 12.7 ± 3.9 | 3.5 | 8.7 ± 1.0 | 3.6 |
| Subtotal | 34.7 ± 5.7 | 17.3 | 49.5 ± 5.7 | 16.0 | 88.1 ± 16.7 | 24.1 | 14.0 ± 3.9 | 7.3 | |
| Rotifers | |||||||||
| Adinetidae | Adineta ricciae | 1.7 ± 0.7 | 0.8 | 5.5 ± 1.2 | 1.8 | 0.6 ± 0.3 | 0.2 | 0.0 ± 0.0 | 0.0 |
| Asplanchanidae | Asplanchna priodonta | 0.2 ± 0.1 | 0.1 | 0.7 ± 0.1 | 0.2 | 5.7 ± 0.5 | 1.6 | 0.5 ± 0.2 | 0.2 |
| Brachionidae | Anuraeopsis fissa | 2.4 ± 1.1 | 1.2 | 1.3 ± 0.4 | 0.4 | 2.0 ± 0.3 | 0.5 | 0.6 ± 0.1 | 0.2 |
| Brachionus angularis | 0.0 ± 0.0 | 0.0 | 0.7 ± 0.2 | 0.2 | 2.0 ± 0.1 | 0.5 | 2.6 ± 0.5 | 1.1 | |
| Brachionus caudatus | 0.0 ± 0.0 | 0.0 | 0.2 ± 0.1 | 0.1 | 3.2 ± 0.2 | 0.9 | 1.4 ± 0.5 | 0.6 | |
| Brachionus calyciflorus | 0.6 ± 0.1 | 0.3 | 0.3 ± 0.1 | 0.1 | 24.4 ± 5.2 | 6.7 | 0.1 ± 0.0 | 0.0 | |
| Brachionus donneri | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.7 ± 0.1 | 0.2 | 0.1 ± 0.1 | 0.0 | |
| Brachionus falcatus | 3.1 ± 0.8 | 1.5 | 12.4 ± 0.9 | 4.0 | 14.9 ± 3.5 | 4.1 | 0.0 ± 0.0 | 0.0 | |
| Brachionus forficula | 1.9 ± 1.2 | 0.9 | 9.2 ± 1.5 | 3.0 | 5.1 ± 1.2 | 1.4 | 0.0 ± 0.0 | 0.0 | |
| Brachionus patulus | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.1 ± 0.1 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Brachionus quadridentatus | 0.0 ± 0.0 | 0.0 | 1.1 ± 0.2 | 0.4 | 0.8 ± 0.4 | 0.2 | 0.6 ± 0.3 | 0.2 | |
| Brachionus sericus | 0.0 ± 0.0 | 0.0 | 7.1 ± 0.9 | 2.3 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Keratella cochlearis | 16.8 ± 7.2 | 8.5 | 26.5 ± 5.2 | 8.6 | 32.3 ± 4.2 | 8.8 | 1.9 ± 0.4 | 0.8 | |
| Keratella tecta | 2.5 ± 0.9 | 1.2 | 1.9 ± 0.6 | 0.6 | 0.6 ± 0.2 | 0.2 | 0.9 ± 0.2 | 0.4 | |
| Keratella tropica | 0.7 ± 0.2 | 0.3 | 12.9 ± 1.7 | 4.2 | 0.5 ± 0.3 | 0.1 | 0.5 ± 0.3 | 0.2 | |
| Lepadella ovalis | 0.9 ± 0.1 | 0.4 | 0.0 ± 0.0 | 0.0 | 0.2 ± 0.1 | 0.1 | 0.0 ± 0.0 | 0.0 | |
| Filinidae | Filinia camasecla | 0.0 ± 0.0 | 0.0 | 0.1 ± 0.0 | 0.0 | 0.3 ± 0.1 | 0.1 | 0.0 ± 0.0 | 0.0 |
| Filinia longiseta | 0.0 ± 0.0 | 0.0 | 0.1 ± 0.1 | 0.0 | 0.5 ± 0.2 | 0.1 | 0.0 ± 0.0 | 0.0 | |
| Filinia opoliensis | 1.2 ± 0.5 | 0.6 | 1.6 ± 0.4 | 0.5 | 16.9 ± 2.3 | 4.6 | 1.3 ± 0.1 | 0.5 | |
| Filinia terminalis | 3.7 ± 1.3 | 1.8 | 1.2 ± 0.2 | 0.4 | 0.0 ± 0.0 | 0.0 | 0.5 ± 0.2 | 0.2 | |
| Flosculariidae | Ptygura libera | 3.1 ± 0.4 | 1.5 | 25.1 ± 7.2 | 8.1 | 30.8 ± 5.2 | 8.4 | 96.7 ± 11.2 | 39.7 |
| Gastropodidae | Ascomorpha ecaudis | 5.6 ± 1.9 | 2.8 | 0.8 ± 0.2 | 0.3 | 0.5 ± 0.2 | 0.1 | 0.0 ± 0.0 | 0.0 |
| Gastropus stylifer | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.1 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Hexarthridae | Hexathra mira | 11.4 ± 6.2 | 5.7 | 5.2 ± 1.1 | 1.7 | 21.0 ± 1.0 | 5.7 | 1.3 ± 0.1 | 0.5 |
| Lecanidae | Lecane bulla | 2.5 ± 0.3 | 1.2 | 0.3 ± 0.1 | 0.1 | 0.1 ± 0.0 | 0.0 | 0.3 ± 0.1 | 0.1 |
| Lecane curvicornis | 1.2 ± 0.5 | 0.6 | 0.3 ± 0.1 | 0.1 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Monostyla bulla | 0.0 ± 0.0 | 0.0 | 1.0 ± 0.2 | 0.3 | 0.5 ± 0.3 | 0.1 | 0.3 ± 0.1 | 0.1 | |
| Mytillinidae | Lophocharis curvata | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.4 ± 0.2 | 0.1 | 94.4 ± 8.4 | 38.8 |
| Mytillina | 0.1 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.1 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Synchaetidae | Polyarthra vulgaris | 30.3 ± 6.2 | 15.1 | 6.6 ± 1.6 | 2.1 | 17.6 ± 1.7 | 4.8 | 2.2 ± 0.7 | 0.9 |
| Testudinellidae | Pompholyx complanata | 4.4 ± 1.0 | 2.2 | 5.8 ± 1.2 | 1.9 | 2.5 ± 0.9 | 0.7 | 0.0 ± 0.0 | 0.0 |
| Pompholyx sulcata | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Trichocercidae | Trichocerca similis | 2.5 ± 1.1 | 1.2 | 7.7 ± 0.4 | 2.5 | 7.3 ± 1.2 | 2.0 | 4.6 ± 1.2 | 1.9 |
| Trichocerca elongata | 0.3 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | 0.2 ± 0.1 | 0.1 | 5.6 ± 1.3 | 2.3 | |
| Trichocerca pusilla | 1.8 ± 0.2 | 0.9 | 0.3 ± 0.1 | 0.1 | 0.0 ± 0.0 | 0.0 | 0.0 ± 0.0 | 0.0 | |
| Trichocerca cylindrica | 0.9 ± 0.1 | 0.4 | 3.3 ± 0.3 | 1.1 | 0.7 ± 0.3 | 0.2 | 0.9 ± 0.4 | 0.4 | |
| Subtotal | 112.6 ± 28.3 | 56.2 | 139.2 ± 26.3 | 45.1 | 192.5 ± 37.9 | 52.6 | 217.5 ± 42.9 | 87.8 | |
| Grand total | 200.5 ± 25.5 | 100 | 308.8 ± 34.9 | 100 | 365.7 ± 13.7 | 100 | 243.4 ± 28.5 | 100 | |
| Total number of species | 41 | 43 | 43 | 29 | |||||
| Shannon–Wiener diversity index (H’) | 2.2 ± 0.0 | 1.9 ± 0.1 | 1.7 ± 0.1 | 1.5 ± 0.1 | |||||
| Temperature, °C | pH | DO, mg L−1 | Turbidity, NTU | TP, mg L−1 | TN, mg L−1 | Chl a, µg L−1 | W. Transp., m | |
|---|---|---|---|---|---|---|---|---|
| Natural ecosystem | ||||||||
| Shallow swamps | ||||||||
| Chini Lake | 29.68 ± 0.47 d | 6.46 ± 0.39 d | 4.58 ± 0.46 e | 16.80 ± 7.21 h | 0.04 ± 0.02 e | 0.20 ± 0.01 f | 3.88 ± 0.90 p | 1.06 ± 0.10 de |
| Bera Lake | 29.00 ± 0.53 g | 5.80 ± 0.71 e | 4.70 ± 1.0 e | 15.60 ± 3.8 k | 0.03 ± 0.02 e | 0.30 ±0.01 f | 5.72 ± 1.25 o | 1.10 ± 0.20 b |
| Swamp S3 | 27.68 ± 1.2 k | 4.87 ± 0.5 f | 1.97 ± 0.25 h | 17.29 ± 5.16 g | 0.05 ± 0.01 e | 0.27 ± 0.01 f | 11.15 ± 2.30 j | 0.95 ± 0.05 cd |
| Swamp S4 | 28.60 ± 0.97 i | 5.67 ± 0.25 e | 2.30 ± 0.30 g | 14.82 ± 6.29 l | 0.06 ± 0.02 e | 0.24 ± 0.01 f | 10.63 ± 4.28 k | 0.92 ± 0.01 de |
| Artificial ecosystem | ||||||||
| Reservoirs | ||||||||
| Chenderoh Reservoir | 29.90 ±0.90 c | 7.76 ± 1.30 a | 7.40 ± 1.7 b | 20.10 ± 3.52 f | 0.05 ± 0.01 e | 2.05 ± 0.10 b | 9.70 ± 1.95 l | 0.95 ± 0.01 cd |
| Timah Tasoh Reservoir | 28.53 ± 1.2 i | 6.61 ± 0.95 d | 7.50 ± 0.50 b | 16.16 ± 2.77 i | 0.03 ± 0.01 e | 0.20 ± 0.05 f | 12.50 ± 2.85 i | 0.90 ± 0.00 de |
| Kenyir Reservoir | 29.56 ± 1.23 e | 7.21 ± 1.1 b | 7.38 ± 0.92 b | 14.40 ± 1.22 m | 0.10 ± 0.01 d | 0.20 ± 0.01 f | 8.70 ± 1.07 n | 1.70 ± 0.01 a |
| Sembrong Reservoir | 29.70 ± 0.31 d | 7.80 ± 0.22 a | 5.27 ± 0.4 c | 28.40 ± 2.90 d | 0.14 ± 0.01 cde | 2.26 ± 0.33 a | 97.18 ± 4.70 a | 0.30 ± 0.01 g |
| Constructed lakes | ||||||||
| Putrajaya Lake | 30.53 ± 0.22 a | 6.81 ± 0.28 c | 7.50 ± 0.37 b | 23.16 ± 0.75 e | 0.03 ± 0.00 e | 0.17 ± 0.28 f | 15.53 ± 0.65 h | 1.06 ± 0.02 bc |
| Sri Serdang Pond | 28.34 ± 0.2 j | 6.84 ± 0.22 c | 0.74 ± 0.30 i | 39.94 ± 5.02 b | 0.69 ± 0.10 a | 0.20 ± 0.09 f | 95.00 ± 17.9 b | 0.25 ± 0.02 g |
| UPM Pond | 29.12 ± 0.95 g | 7.23 ± 1.22 b | 4.93 ± 0.5 d | 29.94 ± 7.35 c | 0.10 ± 0.0 de | 1.00 ± 0.01 e | 44.70 ±7.23 d | 0.85 ± 0.05 de |
| Cempaka Lake | 28.82 ± 0.57 h | 6.60 ± 2.30 d | 4.94 ± 0.92 d | 42.00 ± 8.55 a | 0.52 ± 0.02 b | 1.24 ± 0.01 d | 48.00 ± 8.92 c | 0.35 ± 0.01 g |
| Old mining lakes | ||||||||
| SKVE Lake | 28.48 ± 0.15 i | 7.10 ± 0.25 b | 2.59 ± 0.28 f | 17.30 ± 1.13 g | 0.20 ± 0.03 cd | 1.85 ± 0.13 c | 16.50 ± 0.14 f | 0.65 ± 0.10 f |
| The Mines Lake | 29.30 ± 0.29 f | 7.22 ± 0.28 b | 0.91 ± 0.17 h | 12.74 ± 0.11 o | 0.22 ± 0.07 cd | 1.72 ± 0.22 c | 16.11 ± 2.11 g | 1.58 ± 0.05 a |
| Biru Kundang Lake | 28.53 ± 0.75 i | 5.70 ± 0.57 e | 8.40 ± 1.20 a | 15.84 ± 2.48 j | 0.26 ± 0.01 c | 1.26 ± 0.05 d | 18.00 ± 3.6 e | 0.80 ± 0.02 e |
| Taiping Lake | 30.32 ± 1.20 b | 6.81 ± 1.23 c | 7.50 ± 0.9 b | 13.16 ± 3.02 n | 0.03 ± 0.00 e | 0.20 ± 0.00 f | 9.50 ± 2.08 m | 0.80 ± 0.03 e |
| pH | Temperature | DO | Turbidity | TP | TN | Chlorophyll a | W. Transparency | |
|---|---|---|---|---|---|---|---|---|
| Temperature | 0.360 | |||||||
| Dissolved oxygen (DO) | −0.161 | 0.445 * | ||||||
| Turbidity | 0.171 | −0.090 | 0.113 | |||||
| Total phosphorus (TP) | 0.129 | −0.354 | −0.263 | −0.017 | ||||
| Total nitrogen (TN) | 0.490* | 0.090 | 0.325 | 0.414 | 0.249 | |||
| Chlorophyll a | 0.401 | −0.134 | 0.240 | 0.663 ** | 0.352 | 0.676 ** | ||
| Water transparency | 0.047 | 0.421 | 0.101 | −0.697 ** | −0.129 | −0.200 | −0.512 * | |
| Diversity | −0.115 | −0.362 | −0.281 | 0.244 | −0.107 | −0.121 | 0.217 | −0.262 |
| Density | 0.086 | −0.009 | 0.357 | 0.496 * | 0.371 | 0.457 * | 0.571 ** | −0.487 * |
| Cladocera | 0.063 | 0.157 | 0.207 | 0.490 * | −0.040 | −0.031 | 0.182 | −0.296 |
| Copepoda | −0.165 | 0.028 | 0.193 | 0.083 | 0.068 | −0.244 | 0.045 | −0.172 |
| Rotifera | 0.097 | 0.060 | 0.224 | 0.236 | 0.152 | −0.144 | 0.270 | −0.292 |
| Ceriodaphnia cornuta | 0.181 | 0.127 | 0.272 | 0.597 * | 0.438 * | 0.529 * | 0.543 * | −0.303 |
| Polyarthra vulgaris | −0.363 | 0.026 | −0.143 | −0.189 | −0.522 * | −0.669 ** | −0.666 ** | −0.047 |
| Keratella cochlearis | 0.322 | 0.380 | 0.407 | 0.540 * | 0.075 | 0.506 * | 0.526 * | −0.114 |
| Trichocerca similis | 0.232 | −0.211 | −0.106 | 0.735 ** | 0.426 * | 0.291 | 0.752 ** | −0.643 ** |
| Brachionus calyciflorus | 0.368 | −0.078 | −0.134 | 0.713 ** | 0.276 | 0.250 | 0.714 ** | −0.472 * |
| Brachionus falcatus | 0.308 | −0.340 | −0.057 | 0.708 ** | 0.108 | 0.200 | 0.421 | −0.632 ** |
| Thermocyclops crassus | 0.203 | −0.044 | 0.126 | 0.641 ** | 0.039 | 0.193 | 0.397 | −0.529 * |
| Global Rho (ρ) = 0.508; p = 0.1% | ||
|---|---|---|
| No. Variables | Correlation (ρ) | Selections |
| 1 | 0.334 | Chlorophyll a |
| 2 | 0.456 | Chlorophyll a, total nitrogen |
| 3 | 0.508 | Chlorophyll a, turbidity, total nitrogen |
| 4 | 0.454 | Chlorophyll a, turbidity, total nitrogen, temperature |
| 5 | 0.437 | Chlorophyll a, total nitrogen, temperature, pH, turbidity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umi, W.A.D.; Yusoff, F.M.; Balia Yusof, Z.N.; Ramli, N.M.; Sinev, A.Y.; Toda, T. Composition, Distribution, and Biodiversity of Zooplanktons in Tropical Lentic Ecosystems with Different Environmental Conditions. Arthropoda 2024, 2, 33-54. https://doi.org/10.3390/arthropoda2010003
Umi WAD, Yusoff FM, Balia Yusof ZN, Ramli NM, Sinev AY, Toda T. Composition, Distribution, and Biodiversity of Zooplanktons in Tropical Lentic Ecosystems with Different Environmental Conditions. Arthropoda. 2024; 2(1):33-54. https://doi.org/10.3390/arthropoda2010003
Chicago/Turabian StyleUmi, Wahidah Ahmad Dini, Fatimah M. Yusoff, Zetty Norhana Balia Yusof, Norulhuda Mohamed Ramli, Artem Y. Sinev, and Tatsuki Toda. 2024. "Composition, Distribution, and Biodiversity of Zooplanktons in Tropical Lentic Ecosystems with Different Environmental Conditions" Arthropoda 2, no. 1: 33-54. https://doi.org/10.3390/arthropoda2010003
APA StyleUmi, W. A. D., Yusoff, F. M., Balia Yusof, Z. N., Ramli, N. M., Sinev, A. Y., & Toda, T. (2024). Composition, Distribution, and Biodiversity of Zooplanktons in Tropical Lentic Ecosystems with Different Environmental Conditions. Arthropoda, 2(1), 33-54. https://doi.org/10.3390/arthropoda2010003