
How Urban-Tolerant Are They? Testing Prey–Capture Behavior of Introduced Jorō Spiders (Trichonephila clavata) Next to Busy Roads


Abstract
1. Introduction
2. Methods
2.1. Road Selection
2.2. Stimulus Procedure
2.3. Data Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradley, C.A.; Altizer, S. Urbanization and the ecology of wildlife diseases. Trends Ecol. Evol. 2007, 22, 95–102. [Google Scholar] [CrossRef]
- Proppe, D.S.; Sturdy, C.B.; St Clair, C.C. Anthropogenic noise decreases urban songbird diversity and may contribute to homogenization. Glob. Chang. Biol. 2013, 19, 1075–1084. [Google Scholar] [CrossRef]
- Dahirel, M.; De Cock, M.; Vantieghem, P.; Bonte, D. Urbanization-driven changes in web building and body size in an orb web spider. J. Anim. Ecol. 2019, 88, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Lowry, H.; Lill, A.; Wong, B.B.M. Behavioural responses of wildlife to urban environments. Biol. Rev. 2013, 88, 537–549. [Google Scholar] [CrossRef]
- Duquette, C.A.; Loss, S.R.; Hovick, T.J. A meta-analysis of the influence of anthropogenic noise on terrestrial wildlife communication strategies. J. Appl. Ecol. 2021, 58, 1112–1121. [Google Scholar] [CrossRef]
- Kleist, N.J.; Guralnick, R.P.; Cruz, A.; Lowry, C.A.; Francis, C.D. Chronic anthropogenic noise disrupts glucocorticoid signaling and has multiple effects on fitness in an avian community. Proc. Natl. Acad. Sci. USA 2018, 115, E648–E657. [Google Scholar] [CrossRef]
- Shannon, G.; McKenna, M.F.; Angeloni, L.M.; Crooks, K.R.; Fristrup, K.M.; Brown, E.; Warner, K.A.; Nelson, M.D.; White, C.; Briggs, J.; et al. A synthesis of two decades of research documenting the effects of noise on wildlife. Biol. Rev. 2016, 91, 982–1005. [Google Scholar] [CrossRef]
- Alvarez, B.X.M.; Carrera-Trevino, R.; Hobson, K.A. Mortality of monarch butterflies (Danaus plexippus) at two highway crossing “hotspots” during autumn migration in northeast Mexico. Front. Ecol. Evol. 2019, 7, 7. [Google Scholar] [CrossRef]
- Keilsohn, W.; Narango, D.L.; Tallamy, D.W. Roadside habitat impacts insect traffic mortality. J. Insect Conserv. 2018, 22, 183–188. [Google Scholar] [CrossRef]
- McKenna, D.D.; McKenna, K.M.; Malcolm, S.B.; Berenbaum, M.R. Mortality of lepidoptera along roadways in central Illinois. J. Lepid. Soc. 2001, 55, 63–68. [Google Scholar]
- Daniel-Ferreira, J.; Berggren, A.; Bommarco, R.; Wissman, J.; Ockinger, E. Bumblebee queen mortality along roads increase with traffic. Biol. Conserv. 2022, 272, 109643. [Google Scholar] [CrossRef]
- Classen-Rodríguez, L.; Tinghitella, R.; Fowler-Finn, K. Anthropogenic noise affects insect and arachnid behavior, thus changing interactions within and between species. Curr. Opin. Insect Sci. 2021, 47, 142–153. [Google Scholar] [CrossRef]
- Davis, A.K.; Schroeder, H.; Yeager, I.; Pearce, J. Effects of simulated highway noise on heart rates of larval monarch butterflies, Danaus plexippus: Implications for roadside habitat suitability. Biol. Lett. 2018, 14, 20180018. [Google Scholar] [CrossRef]
- Mitchell, T.S.; Agnew, L.; Meyer, R.; Sikkink, K.L.; Oberhauser, K.S.; Borer, E.T.; Snell-Rood, E.C. Traffic influences nutritional quality of roadside plants for monarch caterpillars. Sci. Total Environ. 2020, 724, 138045. [Google Scholar] [CrossRef]
- Orci, K.M.; Petróczki, K.; Barta, Z. Instantaneous song modification in response to fluctuating traffic noise in the tree cricket Oecanthus Pellucens. Anim. Behav. 2016, 112, 187–194. [Google Scholar] [CrossRef]
- Raboin, M.; Elias, D.O. Anthropogenic noise and the bioacoustics of terrestrial invertebrates. J. Exp. Biol. 2019, 222, jeb178749. [Google Scholar] [CrossRef]
- Hoebeke, E.R.; Huffmaster, W.; Freeman, B.J. Nephila clavata L Koch, the Jorō Spider of East Asia, newly recorded from North America (Araneae: Nephilidae). PeerJ 2015, 3, e763. [Google Scholar] [CrossRef] [PubMed]
- Chuang, A.; Deitsch, J.F.; Nelsen, D.R.; Sitvarin, M.I.; Coyle, D.R. The Jorō spider (Trichonephila clavata) in the southeastern U.S.: An opportunity for research and a call for reasonable journalism. Biol. Invasions 2023, 25, 17–26. [Google Scholar]
- Davis, A.K.; Frick, B.L. Physiological evaluation of newly invasive jorō spiders (Trichonephila clavata) in the southeastern USA compared to their naturalized cousin, Trichonephila Clavipes. Physiol. Entomol. 2022, 2022, 170–175. [Google Scholar] [CrossRef]
- Nelsen, D.R.; Corbit, A.G.; Chuang, A.; Deitsch, J.F.; Sitvarin, M.I.; Coyle, D.R. Veni, vidi, vici? Future spread and ecological impacts of a rapidly expanding invasive predator population. Ecol. Evol. 2023, 13, e10728. [Google Scholar] [CrossRef] [PubMed]
- Lowe, E.C.; Wilder, S.M.; Hochuli, D.F. Life history of an urban-tolerant spider shows resilience to anthropogenic habitat disturbance. J. Urban Ecol. 2017, 3, jux004. [Google Scholar] [CrossRef]
- Ripp, J.; Eldakar, O.T.; Gallup, A.C.; Arena, P.T. The successful exploitation of urban environments by the golden silk spider, Nephila clavipes (Araneae, Nephilidae). J. Urban Ecol. 2018, 4, juy005. [Google Scholar] [CrossRef]
- Sabanal, B.; Achondo, M.J.M.; Alviola, P.; Gamalo, L.E.; Responte, M. Diversity of spiders (Araneae) in the anthropic land covers of Davao City, Philippines. J. Anim. Divers. 2022, 4, 28–39. [Google Scholar] [CrossRef]
- Davis, A.K.; Anerao, A. Startle responses of jorō spiders (Trichonephila clavata) to artificial disturbance. Arthropoda 2023, 2023, 60–67. [Google Scholar] [CrossRef]
- Davies, M.S.; Hesselberg, T. The use of tuning forks for studying behavioural responses in orb web spiders. Insects 2022, 13, 370. [Google Scholar] [CrossRef]
- Stevenson, P.A.; Dyakonova, V.; Rillich, J.; Schildberger, K. Octopamine and experience-dependent modulation of aggression in crickets. J. Neurosci. 2005, 25, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.E.; Pervin, E.; Graham, S.L.; Henry, M.; Fahrig, L. Abundance of aerially-dispersing spiders declines with increasing road traffic. Ecoscience 2019, 26, 383–388. [Google Scholar] [CrossRef]
- Pessman, B.J.; Hays, M.; Agpawa, E.; Hebets, E.A. Urbanization affects web abundance and aggregation of a funnel-weaving spider, Agelenopsis pennsylvanica (Agelenidae). Urban Ecosyst. 2023, 26, 1275–1292. [Google Scholar] [CrossRef]
- Landolfa, M.A.; Barth, F.G. Vibrations in the orb web of the spider Nephila clavipes: Cues for discrimination and orientation. J. Comp. Physiol. A-Sens. Neural Behav. Physiol. 1996, 179, 493–508. [Google Scholar] [CrossRef]
- Blackledge, T.A. Prey capture in orb weaving spiders: Are we using the best metric? J. Arachnol. 2011, 39, 205–210. [Google Scholar] [CrossRef]
- Grabarczyk, E.E.; Querejeta, M.; Tillman, P.G.; Wallace, R.D.; Barnes, B.F.; Meinecke, C.D.; Villari, C.; Gandhi, K.J.K.; LaForest, J.; Elliott, M.; et al. DNA metabarcoding analysis of three material types to reveal Jorō spider (Trichonephila clavata) trophic interactions and web capture. Front. Ecol. Evol. 2023, 11, 1177446. [Google Scholar] [CrossRef]


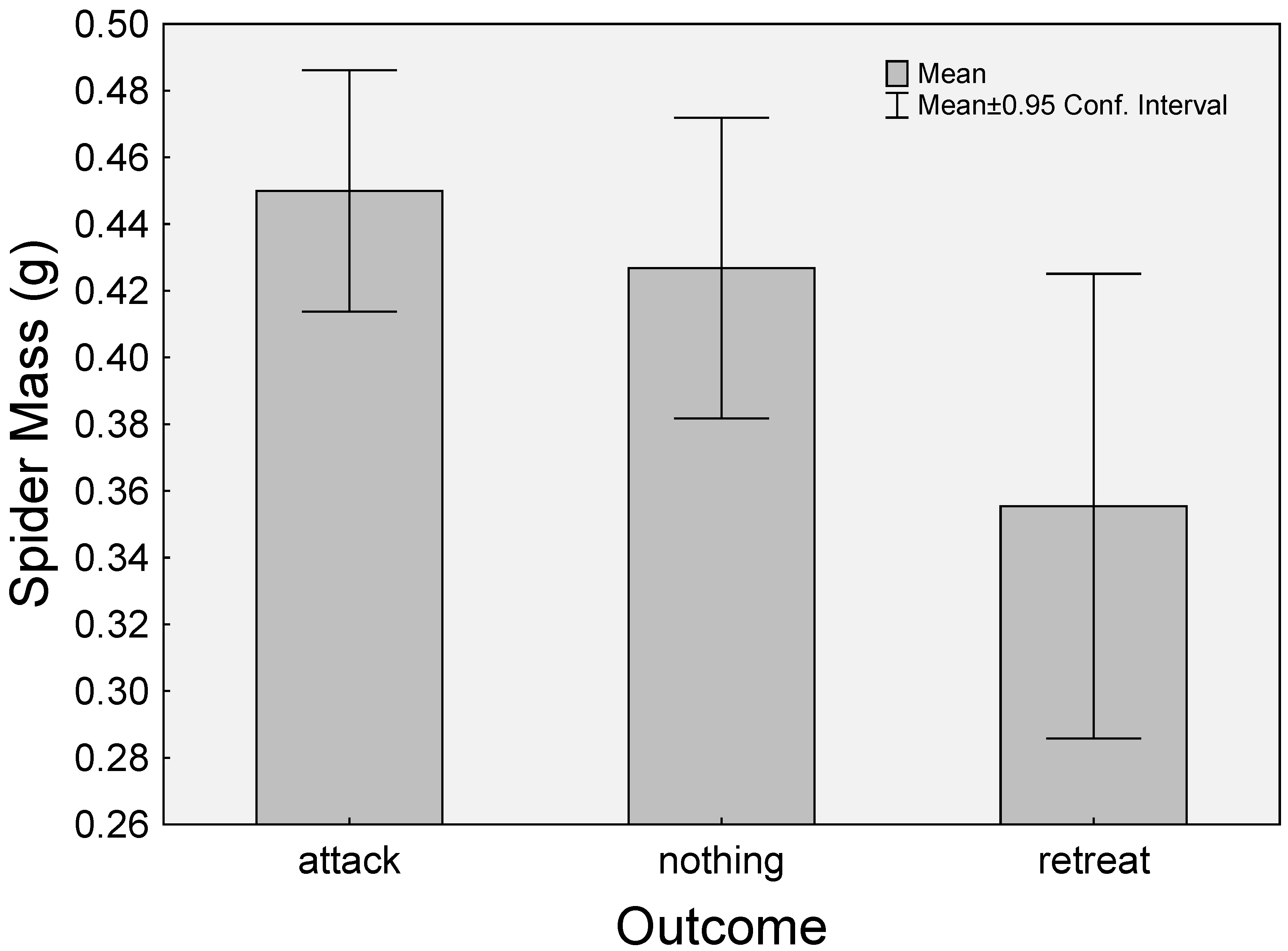


{kind=link}
{kind=link}
{kind=link}
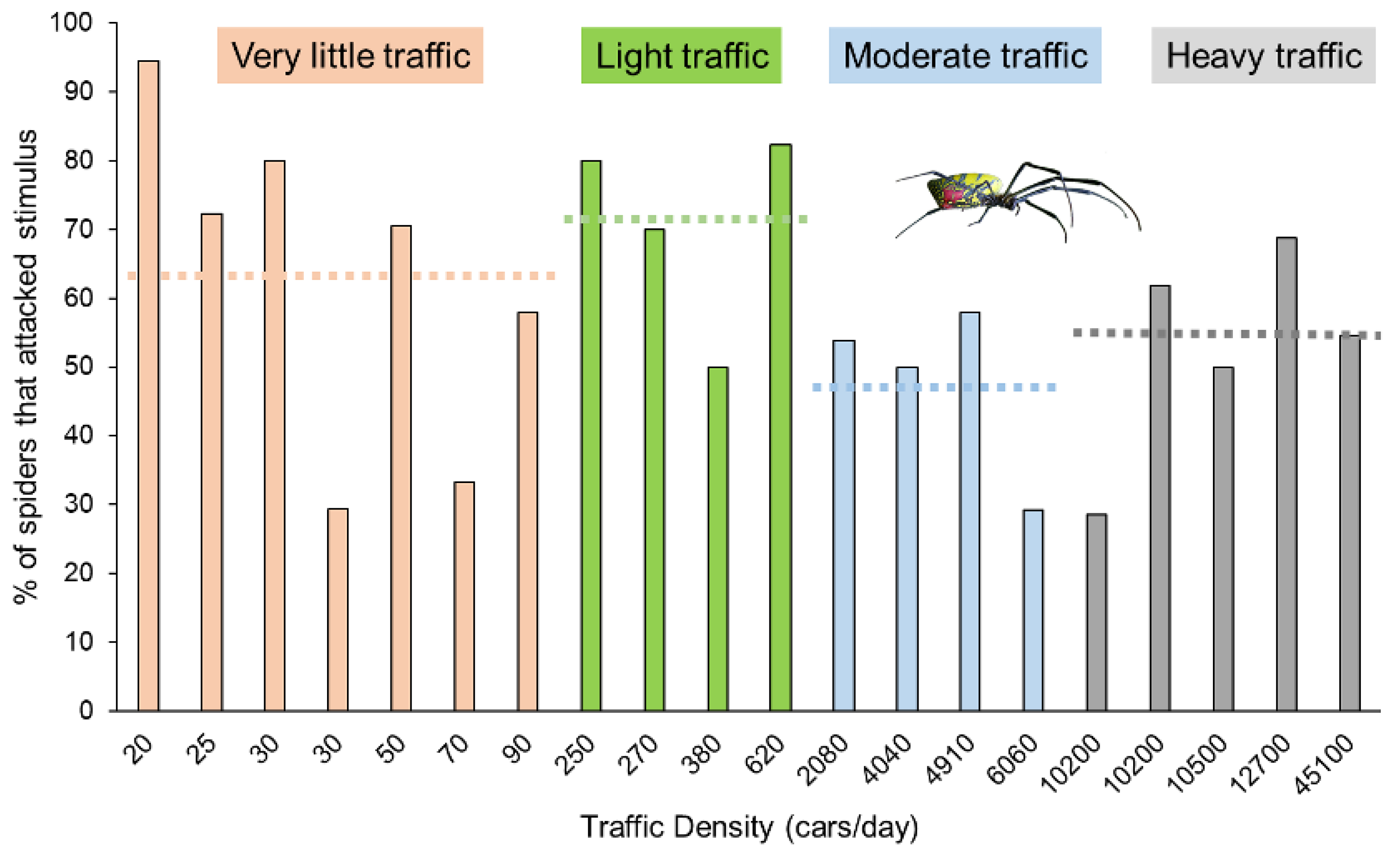{kind=link}
| Road Name | County | Traffic Density | Density Category | # Attack | # Nothing | # Retreat | Total Trials | % Attack | Ave. Spider Mass (g) |
|---|---|---|---|---|---|---|---|---|---|
| Marshall Store Rd. | Oconee | 20 | 1 | 17 | 1 | 0 | 18 | 94.4 | 0.33 |
| Rose Creek Dr. | Oconee | 25 | 1 | 13 | 5 | 0 | 18 | 72.2 | 0.55 |
| Freeman Creek Rd. | Oconee | 30 | 1 | 12 | 1 | 2 | 15 | 80.0 | 0.37 |
| JT Elder Rd. | Oconee | 30 | 1 | 5 | 11 | 1 | 17 | 29.4 | 0.34 |
| Hardigree Bell Rd. | Oconee | 50 | 1 | 12 | 4 | 1 | 17 | 70.6 | 0.23 |
| Old Farmington Rd. | Oconee | 70 | 1 | 5 | 7 | 3 | 15 | 33.3 | 0.66 |
| Old Ila Rd. | Madison | 90 | 1 | 11 | 6 | 2 | 19 | 57.9 | 0.40 |
| Antioch Church Rd. | Oconee | 250 | 2 | 16 | 3 | 1 | 20 | 80.0 | 0.42 |
| Mayne Mill Rd. | Oconee | 270 | 2 | 14 | 4 | 2 | 20 | 70.0 | 0.40 |
| Oliver Bridge Rd. | Oconee | 380 | 2 | 9 | 7 | 2 | 18 | 50.0 | 0.53 |
| Wesley Chapel Rd. | Madison | 620 | 2 | 14 | 1 | 2 | 17 | 82.4 | 0.36 |
| Colham Ferry Rd. | Oconee | 2080 | 3 | 14 | 11 | 1 | 26 | 53.8 | 0.64 |
| South Main St. Watkinsville | Oconee | 4040 | 3 | 9 | 7 | 2 | 18 | 50.0 | 0.35 |
| Greensboro Hwy. | Oconee | 4910 | 3 | 11 | 6 | 2 | 19 | 57.9 | 0.58 |
| E Whitehall Rd. | Clarke | 6060 | 3 | 7 | 11 | 6 | 24 | 29.2 | 0.38 |
| S. Milledge Rd. | Clarke | 10,200 | 4 | 2 | 2 | 3 | 7 | 28.6 | 0.22 |
| US 441 | Oconee | 10,200 | 4 | 13 | 7 | 1 | 21 | 61.9 | 0.52 |
| Mitchell Bridge Rd. | Clarke | 10,500 | 4 | 6 | 4 | 2 | 12 | 50.0 | 0.45 |
| Experiment Stn Rd. | Oconee | 12,700 | 4 | 11 | 5 | 0 | 16 | 68.8 | 0.50 |
| Athens Perimeter/10 Loop | Clarke | 45,100 | 4 | 12 | 6 | 4 | 22 | 54.5 | 0.28 |
| Grand Total | 213 | 109 | 37 | 359 | 59.3 | 0.43 |
| Traffic Density | Attack | Nothing | Retreat | No Attack | Grand Total | % Attack |
|---|---|---|---|---|---|---|
| 1—Very little a | 75 | 35 | 9 | 44 | 119 | 63.0 |
| 2—Light a | 53 | 15 | 7 | 22 | 75 | 70.7 |
| 3—Moderate b | 41 | 35 | 11 | 46 | 87 | 47.1 |
| 4—Heavy b | 44 | 24 | 10 | 34 | 78 | 56.4 |
| Grand Total | 213 | 109 | 37 | 146 | 359 |
| Predictor | Estimate | SE | Wald | Lower 95% CL | Upper 95% CL | p |
| Spider Mass | 0.688 | 0.442 | 2.429 | −1.177 | 1.554 | 0.1191 |
| Traffic Volume | −0.106 | 0.044 | 5.592 | −0.193 | −0.018 | 0.0180 |
| Predictor | Estimate | SE | Wald | Lower 95% CL | Upper 95% CL | p |
| Spider Mass | −0.828 | 0.450 | 3.391 | −1.709 | 0.053 | 0.0656 |
| Volume Category | −0.498 | 0.212 | 5.534 | −0.914 | −0.083 | 0.0187 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, A.K.; Stewart, K.; Phelan, C.; Schultz, A. How Urban-Tolerant Are They? Testing Prey–Capture Behavior of Introduced Jorō Spiders (Trichonephila clavata) Next to Busy Roads. Arthropoda 2024, 2, 55-65. https://doi.org/10.3390/arthropoda2010004
Davis AK, Stewart K, Phelan C, Schultz A. How Urban-Tolerant Are They? Testing Prey–Capture Behavior of Introduced Jorō Spiders (Trichonephila clavata) Next to Busy Roads. Arthropoda. 2024; 2(1):55-65. https://doi.org/10.3390/arthropoda2010004
Chicago/Turabian StyleDavis, Andrew K., Kade Stewart, Caitlin Phelan, and Alexa Schultz. 2024. "How Urban-Tolerant Are They? Testing Prey–Capture Behavior of Introduced Jorō Spiders (Trichonephila clavata) Next to Busy Roads" Arthropoda 2, no. 1: 55-65. https://doi.org/10.3390/arthropoda2010004
APA StyleDavis, A. K., Stewart, K., Phelan, C., & Schultz, A. (2024). How Urban-Tolerant Are They? Testing Prey–Capture Behavior of Introduced Jorō Spiders (Trichonephila clavata) Next to Busy Roads. Arthropoda, 2(1), 55-65. https://doi.org/10.3390/arthropoda2010004