Abstract
Extracellular Vesicle-associated microRNAs (EV-miRNAs) are emerging as pivotal regulators of corneal health and disease, holding exceptional promise for transforming both diagnostics and therapeutics. These vesicles carry distinct miRNA signatures in biofluids such as tears, offering a powerful, non-invasive approach for early detection, risk stratification, and dynamic monitoring of corneal disorders. In addition, EV-miRNAs act as key mediators of critical biological processes, including inflammation, fibrosis, and tissue repair. Consequently, they represent attractive therapeutic targets; for example, engineered EVs loaded with miRNA mimics or inhibitors can precisely modulate these pathways to promote regeneration and suppress disease progression. Yet, despite this considerable promise, the translation of EV-miRNA research into clinical practice remains constrained by several challenges. Topmost among these are the lack of standardized EV isolation methods, variability in miRNA quantification, and the pressing need for regulatory frameworks tailored to the complexity of these biological therapeutics. Addressing these barriers is essential to ensure reproducibility, scalability, and safety in clinical applications. Accordingly, this review synthesizes current knowledge on EV-miRNA profiles in corneal diseases, critically evaluates their diagnostic and therapeutic potential, and highlights strategies to overcome existing technical and regulatory limitations. Ultimately, the successful integration of EV-miRNA-based approaches into personalized medicine frameworks could revolutionize the management of corneal diseases and substantially improve patient outcomes.
1. Introduction
Corneal disorders impose a substantial global health burden, ranking among the leading causes of visual impairment and blindness, particularly in low- and middle-income countries [1]. In some rural communities, the prevalence of corneal disease approaches 3.7%, with corneal blindness affecting roughly 0.12% of individuals studied [2]. Common etiological factors include infectious keratitis, keratoconus, ocular trauma, corneal dystrophies and degenerations, as well as postsurgical complications such as bullous keratopathy [3]. These conditions frequently necessitate corneal transplantation and drive the development of novel diagnostic and therapeutic strategies to mitigate their impact [4]. This need is especially urgent in regions where surgical interventions are scarce, elevating the importance of preventive and regenerative approaches in maintaining corneal health [5]. Moreover, vision loss from both unilateral and bilateral corneal disorders significantly diminishes quality of life across multiple functional domains [1]. Diseases such as Fuchs endothelial corneal dystrophy (FECD) carry a heavy economic footprint; in the United States, direct medical expenditures for corneal transplant surgery and postoperative care often exceed $20,000 within the first year, with costs rising for bilateral or repeat procedures [6]. Indirect repercussions, including short-term disability and reduced productivity, further compound the financial strain on patients and healthcare systems [6]. However, parallel exosomes, a specialized subset of extracellular vesicles (EVs), originate from the endosomal pathway through the fusion of multivesicular bodies with the plasma membrane and are characterized by their nanoscale dimensions [7,8,9]. Enclosed by a lipid bilayer, they are secreted by diverse cell types, such as dendritic cells, T cells, epithelial cells, and mesenchymal stem cells, among others [10], and can be detected in body fluids such as blood, urine, and cerebrospinal fluid, underscoring their pivotal role in intercellular communication [10,11]. These vesicles ferry a complex cargo of proteins, lipids, nucleic acids, and metabolites, thereby modulating the physiological or pathological states of recipient cells [8,12,13]. Within the cornea, exosomes orchestrate processes including wound healing, angiogenesis, lymphangiogenesis, and nerve repair by activating signaling pathways such as HSP27, STAT, and β-catenin to accelerate epithelial closure and promote cell migration and proliferation [14,15]. Their intrinsic low immunogenicity and high stability further position them as attractive candidates for therapeutic applications, particularly as targeted drug delivery vehicles in ocular pathologies [16,17,18]. Nonetheless, challenges in reliably purifying and characterizing exosomes must be overcome to translate these findings into clinical practice [19]. The unique capacity of exosomes to deliver molecular payloads with precision underscores their promise in advancing treatment strategies for corneal and other ocular diseases [17,20].
Beyond their protein and lipid components, EVs are instrumental in corneal regeneration through the transport of microRNAs (miRNAs) that regulate inflammation, cell proliferation, and other essential cellular functions [4,21]. Mesenchymal stem cell-derived miRNAs encapsulated within EVs have been shown to bolster corneal endothelial cell survival under stress conditions, exemplifying their therapeutic potential in corneal disorders via EV-mediated delivery [21,22]. Distinct expression profiles of EV-associated miRNAs, such as miR-320 and miR-423-5p, also present promising diagnostic biomarkers for corneal diseases [23]. Profiling these miRNAs can uncover novel therapeutic targets and enable personalized treatment strategies in corneal care [4]. The use of plasma-derived EV-miRNAs offers a minimally invasive, stable alternative to conventional markers, facilitating earlier detection and longitudinal monitoring of corneal conditions [21]. In tear fluid, specific exosomal miRNA signatures have been correlated with clinical phenotypes and postoperative fibrosis risk, for example, distinguishing glaucoma patients with a higher likelihood of scarring after surgery, making tear-based EV-miRNA analysis a rapid, high-quality method for risk stratification and early diagnosis [24]. Building on these insights, engineered exosomes can be loaded with targeted miRNA mimics or inhibitors and administered locally or systemically to modulate pathogenic pathways in corneal tissues, positioning EV-associated miRNAs as both therapeutic agents and diagnostic tools [25]. The integration of synthetic biology approaches and advanced nanoprobe technologies further enhances the precision of EV-miRNA detection and delivery systems [5,26]. Ongoing efforts to optimize EV isolation and analytical methodologies aim to improve the sensitivity and specificity of miRNA-based assays, which could fundamentally transform the diagnosis and treatment of corneal disorders and alleviate their clinical burden [27]. However, this review seeks to consolidate current research on extracellular vesicle-associated miRNAs in corneal diseases, emphasizing their diagnostic applications and therapeutic prospects while addressing the technical and regulatory challenges that must be surmounted for successful clinical translation.
2. Method
Recognizing the narrative rather than systematic nature of this review, we deliberately avoided rigid design filters and instead pursued a broad, inclusive search. We queried PubMed, EMBASE, Web of Science, Scopus, and Google Scholar for studies published between January 2000 and August 2025, using both controlled-vocabulary terms and free-text keywords “extracellular vesicle” or “exosome” combined with “microRNA” or “miRNA” and “cornea” or “corneal,” plus specific disorders (keratoconus; Fuchs endothelial corneal dystrophy; dry eye; limbal stem cell deficiency; alkali burn) and supplemented these with methodological and translational concepts (microfluidics; machine learning; artificial intelligence; Good Manufacturing Practices; regulatory frameworks). After deduplication, the authors screened titles and abstracts to identify original, peer-reviewed articles reporting EV isolation or characterization, miRNA profiling, functional assays, or preclinical/clinical corneal applications. We included English-language primary research on EV-associated miRNAs in corneal health or disease, excluding reviews, editorials, conference abstracts, non-ocular studies, and papers lacking relevant miRNA data. Full texts of all eligible studies were then examined, with additional hand-searching of pivotal journals and reference to the MISEV and EV-TRACK guidelines to ensure exhaustive coverage. This comprehensive methodology provided a robust and reproducible basis for integrating diagnostic, mechanistic, and therapeutic perspectives on EV-miRNAs in corneal disorders.
3. Corneal Architecture and Pathological Spectrum
The cornea serves as the eye’s primary refractive interface and protective barrier, comprising anatomically distinct layers: the stratified squamous epithelium, Bowman’s layer, the collagen-rich stroma, Dua’s Layer, Descemet’s membrane, and the monolayer endothelium (Figure 1) [28,29]. The epithelium offers the first line of defense against environmental insults, whereas Bowman’s layer confers tensile strength. Meanwhile, the stroma accounting for approximately 90 percent of corneal thickness ensures transparency and biomechanical stability through its intricately organized collagen matrix and resident keratocytes. Underpinning the endothelium, Descemet’s membrane supports a monolayer of endothelial cells that actively regulate stromal hydration via ion and fluid transport to maintain clarity. Owing to its avascularity, the cornea relies on the aqueous humor for nutrient supply and features one of the highest densities of sensory innervation in the body to elicit protective reflexes upon injury [30,31]. Moreover, limbal epithelial stem cells located at the corneal periphery are essential for continuous epithelial renewal and wound repair [32,33]. Disruption of these finely tuned structures and functions underlies a spectrum of corneal disorders with often profound visual consequences. Keratoconus, for instance, is defined by progressive stromal thinning and conical protrusion that result in irregular astigmatism and distorted vision; its multifactorial etiology encompasses genetic predisposition, altered biomechanics, and dysregulated extracellular matrix turnover mediated by matrix metalloproteinases [34,35,36], while concurrent nerve alterations may elicit neuropathic pain [30,37]. Another significant condition is FECD, a genetic disorder typified by endothelial cell degeneration and Descemet’s membrane guttae formation driven by pathogenic protein aggregation and genetic mutations, which culminate in stromal edema and frequently necessitate corneal transplantation [38,39,40,41]. In addition, corneal neovascularization, provoked by hypoxia or chronic inflammation, compromises transparency through pathological vessel ingrowth and often arises in autoimmune or inflammatory contexts, sometimes requiring surgical intervention [42,43,44]. Similarly, limbal stem cell deficiency (LSCD) resulting from chemical injury or surgical trauma undermines epithelial regeneration and leads to persistent corneal opacification [33,45]. Timely diagnosis using advanced modalities such as in vivo confocal microscopy, coupled with tailored medical and surgical strategies, is critical for preserving vision and optimizing patient quality of life.
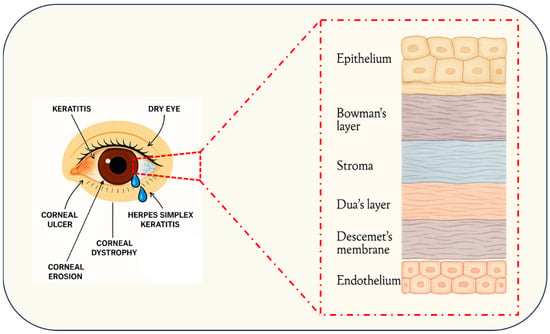
Figure 1.
Schematic cross-section of the human cornea depicting its principal layers epithelium, Bowman’s layer, stroma, Dua’s layer, Descemet’s membrane, and endothelium and the anatomical loci of common corneal pathologies, including keratitis, dry eye, corneal ulceration, epithelial erosions, hereditary dystrophies, and herpes simplex keratitis.
Moreover, corneal pathologies which often correlate with specific cellular layers, Corneal dystrophies, which are genetically determined, non-inflammatory diseases, are accordingly classified based on the affected layer: anterior dystrophies such as epithelial basement membrane dystrophy and Meesmann corneal dystrophy principally compromise the epithelium and its basement membrane, leading to recurrent erosions and visual disturbances [46,47,48]; stromal dystrophies, including lattice and granular variants, manifest as stromal deposits that impair transparency and visual acuity [47,48]; and posterior dystrophies, exemplified by Fuchs’ endothelial corneal dystrophy, target Descemet’s membrane and the endothelium, resulting in corneal edema due to endothelial dysfunction [47,48]. Mustard gas keratopathy further illustrates how exogenous insults can induce layer-specific pathology ranging from epithelial erosion to stromal keratocyte activation, culminating in persistent edema and reduced endothelial cell density [49]. In contrast, keratoconus involves structural and biochemical alterations across all corneal layers, with significant perturbations of stromal collagen architecture and nerve morphology contributing to its progressive nature [50]. Recent advances in genetic and molecular research are therefore redefining corneal dystrophy classification along a molecular basis to facilitate the development of targeted therapies [51,52] (Figure 1).
Limbal stem cell deficiency underscores the critical role of peripheral stem cells in corneal homeostasis; etiologies span congenital aniridia, chemical burns, autoimmune disorders such as Stevens-Johnson syndrome, and iatrogenic factors including contact lens wear [53,54,55] and manifest clinically as conjunctivalization, neovascularization, and persistent epithelial defects that severely compromise vision and quality of life [53,54]. Moreover, diabetic keratopathy demonstrates how systemic hyperglycemia impairs limbal epithelial regeneration, implicating stem cell dysfunction in its pathogenesis [56]. Although LSCD diagnosis can be challenging due to nonspecific clinical features, advanced imaging modalities such as optical coherence tomography and in vivo confocal microscopy have enhanced diagnostic precision [57]. Therapeutic strategies for LSCD include both autologous and allogeneic limbal stem cell transplantation [58], while novel pharmacotherapies aimed at mitigating inflammation and neovascularization hold promise for restoring corneal integrity [59]. Ultimately, deepening our understanding of the cellular and molecular mechanisms underlying LSCD, alongside the refinement of diagnostic and treatment modalities, remains a critical research imperative [60,61], underscoring the need for continued clinical innovation in corneal therapeutics.
4. EV, Cargo Carriers in Action
EVs comprise a heterogeneous family of membrane-bound particles that mediate intercellular communication by transporting proteins, lipids, RNAs, and metabolites across diverse biological systems. Based on their size, origin, and biogenesis pathways, EVs subdivide into three main categories: exosomes (30–150 nm), microvesicles (100–1000 nm), and apoptotic bodies (500–2000 nm) [62,63,64,65] (Figure 2). Exosomes originate within the endosomal pathway when intraluminal vesicles form inside multivesicular bodies (MVBs) and are secreted upon MVB–plasma membrane fusion, a process regulated by the ESCRT machinery, lipids, and proteins such as ALIX and syntenin [66,67]. By contrast, macrovesicles bud directly from the plasma membrane under the influence of signals like calcium flux [68], whereas apoptotic bodies arise during programmed cell death [64,65]. Moreover, EV biogenesis is dynamically modulated by the cell’s metabolic state and external stressors [69].
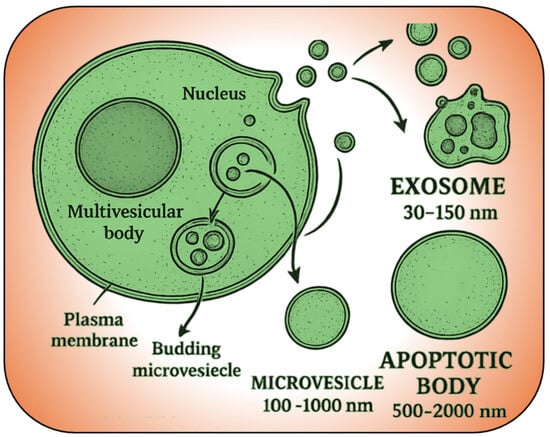
Figure 2.
Overview of extracellular vesicle formation and subtypes. Exosomes (30–150 nm) are released from multivesicular bodies, microvesicles (100–1000 nm) bud directly from the plasma membrane, and apoptotic bodies (500–2000 nm) form during cell apoptosis.
Functionally, EVs ferry bioactive cargos most notably microRNAs that profoundly influence recipient cells, thereby regulating immune responses, cell proliferation, and even cancer progression [70,71,72,73]. In oncology, tumor-derived EVs carry antigens that facilitate immune evasion while disseminating oncogenic factors to promote malignancy in surrounding cells [74,75]. To dissect these roles and harness EVs therapeutically, precise isolation and characterization are essential. Conventional isolation methods, such as ultracentrifugation, are considered the gold standard due to their ability to yield high-purity exosomes, but they are labor-intensive and time-consuming [76,77]. Ultrafiltration, size-exclusion chromatography, and precipitation methods are also traditional techniques, each with specific advantages and limitations. For instance, ultrafiltration is faster but may result in lower purity, while size-exclusion chromatography offers high purity but at a lower yield [78]. Recent advancements have introduced novel methods like microfluidic technologies, which offer rapid, portable, and integrated solutions with improved yield and purity, making them suitable for clinical applications [79,80]. These emerging techniques, including nanolithography and electro-deposition, have significantly enhanced the efficiency of exosome isolation by reducing time and loss [80] (Table 1). In terms of characterization, traditional methods such as dynamic light scattering (DLS), electron microscopy, and nanoparticle tracking analysis (NTA) are widely used to determine size and morphology [81]. However, these methods have limitations in functional tracing and protein quantification. Super-resolution imaging technologies have emerged as a powerful tool, providing real-time, high-resolution analysis that surpasses conventional techniques [81] (Table 2). Comparative studies have shown that while ultracentrifugation provides higher RNA content, commercial kits offer cost-effectiveness and time efficiency, albeit with potential impurities [76,77]. Techniques like qEV Size Exclusion Chromatography have been noted for their purity and concentration of tumor DNA, although they may yield lower overall exosome concentrations [82]. Overall, the choice of isolation and characterization methods depends on the specific requirements of the study, balancing between purity, yield, and operational efficiency.

Table 1.
Overview of conventional and emerging methods for exosome isolation, highlighting their principles, yield, purity, throughput, advantages, and limitations.
Table 2.
Comparative overview of conventional and emerging exosome characterization methods, summarizing each technique’s underlying principle, key advantages, methodological limitations, and typical applications in exosome research and diagnostics.
In sum, EVs serve as both critical mediators of physiological communication and promising therapeutic targets. Advances in our understanding of their classification, biogenesis, and cargo-mediated functions will undoubtedly broaden their application in diagnostics and medicine.
5. EV-Associated miRNAs in Corneal Function
MicroRNAs (miRNAs) are 21–24-nucleotide, non-coding RNAs that bind the 3′ untranslated region of target mRNAs to induce translational repression or degradation, and when encapsulated within EVs, they are protected from RNase-mediated degradation and can be shuttled between cells over variable distances [21,98]. The cornea, a transparent, avascular tissue comprising the eye’s anterior segment, harbors roughly 25% of all ocular miRNAs, many of which are selectively loaded into EVs for precise delivery within the corneal microenvironment (Table 3) [21,25,99]. As critical mediators of cellular homeostasis, EV-associated miRNAs govern wound healing, inflammation, and tissue regeneration. For example, miR-21 promotes epithelial wound closure via PTEN/PI3K/Akt signaling, whereas miR-138 modulates fibrotic cell behavior as well as inflammatory and angiogenic responses following injury or surgery [5,100,101,102]. Moreover, EVs from corneal stromal stem cells contain unique miRNA repertoires distinct from their parent cells, reflecting dynamic cargo remodeling driven by physiological cues such as stress or inflammation [103,104].
EV-miRNAs also orchestrate intercellular signaling in the cornea. EVs secreted by myofibroblasts enhance epithelial cell migration to support re-epithelialization [100], and cross-talk via miRNA-rich vesicles regulates differentiation, proliferation, and apoptosis of recipient cells [105,106]. Functionally, EV cargo, including miR-145 regulation of ITGB8, shapes progenitor renewal, epithelial integrity, glycogen metabolism, and extracellular matrix organization [25,99]. Following corneal insult, tear-film EVs exhibit altered levels of over 120 miRNAs, many linked to inflammation or repair [107]. Key immunomodulatory miRNAs such as miR-146a and miR-155 tune toll-like receptor pathways to limit fibrosis and pathological inflammation after surgery or infection [24,108,109]. Conversely, aberrant EV-miRNA signatures underlie disorders like keratoconus, where they foster extracellular matrix degradation and biomechanical weakening and neovascularization or dystrophies, highlighting their dual role as both biomarkers and drivers of disease [25,99]. Mechanistically, the dysregulated miRNA landscape in keratoconus converges on three interlinked pathogenic axes: extracellular matrix (ECM) degradation, oxidative stress, and biomechanical dysfunction. Specifically, upregulated miR-143-3p, miR-182-5p, and miR-92a-3p enhance matrix metalloproteinase expression and directly repress structural collagens (e.g., COL1A1, COL4A4) and integrins (ITGA1/ITGA2), thereby accelerating stromal thinning and loss of corneal rigidity [110]. Concurrently, downregulation of miR-29b-3p and miR-205-5p lifts inhibition on pro-fibrotic TGF-β signaling through derepression of SMAD2/3 targets and weakens collagen cross-linking, compounding ECM breakdown [111]. Moreover, oxidative insults in KC epithelial and stromal cells provoke shifts in miR-151a-3p and miR-194-5p levels, which normally modulate antioxidant defense pathways via Nrf2 and superoxide dismutase 2. Their dysregulation tips the redox balance toward lipid peroxidation and mitochondrial dysfunction, further destabilizing the corneal microenvironment [110,112]. Bioinformatic investigation has also uncovered a competitive endogenous RNA network featuring miR-4257 and miR-4494, which may govern posttranslational modifications and cellular stress adaptations in KC [113]. Finally, aberrant biomechanical forces, whether from chronic eye rubbing or elevated intraocular pressure activate mechanotransducers YAP1/TEAD1 and TGF-β1, triggering a feedback loop in which cyclic stretch itself upregulates miR-21 and miR-34a. These miRNAs inhibit SMAD7 and other anti-fibrotic repressors, amplifying myofibroblast differentiation and protease release that propagate ECM loss and ectatic deformation [111,114]. Together, these miRNA-driven processes form an integrated network: mechanical stress and oxidative damage reshape miRNA expression, which in turn dismantles ECM integrity and promotes fibrotic remodeling, offering precise mechanistic entry points for novel diagnostic biomarkers and targeted therapies in keratoconus. Notably, exosomal miR-184 from limbal niche cells preserves corneal avascularity by inhibiting lymphangiogenesis and angiogenesis [25]. In disease-specific contexts, EVs from human corneal endothelial cells enriched in miR-184 and miR-24-3p protect against oxidative stress-induced degeneration by repressing pro-inflammatory cytokines, VEGF, and MMPs [115], whereas mesenchymal stromal cell-derived EVs carrying miR-125b and let-7b attenuate dry eye inflammation via the IRAK1/TAB2/NF-κB axis [116]. Corneal stromal stem cell EVs loaded with miR-663b bolster limbal epithelial stem cell stemness through Notch pathway modulation [27], and normal limbal stromal cell exosomes activate Akt signaling to promote epithelial repair capacity diminished in diabetic counterparts [105]. In keratoconus, stromal cell-derived exosomes display distinct miR-184, miR-328-3p, miR-532-5p, miR-345-5p, and miR-424-5p profiles that affect corneal cell migration and proliferation [117,118]. while tear-film EVs from diabetic patients contain dysregulated miRNAs involved in glycosphingolipid biosynthesis and AMPK signaling, correlating with delayed healing and dry-eye features [119]. Such differential expression confirms EV-miRNAs as both diagnostic biomarkers and therapeutic targets, as evidenced by healthy exosomes normalizing gene expression in keratoconus cells [117]. Finally, mesenchymal stem cell-derived EVs have been harnessed to modulate inflammation and promote scarless healing in corneal tissue engineering [21,101]. They deliver anti-fibrotic miRNAs that impede fibroblast-to-myofibroblast transition [109,120,121] and support epithelial repair by enhancing survival, proliferation, and caspase-3-mediated apoptosis inhibition [122]. Equine MSC secretomes similarly accelerate re-epithelialization through growth-factor delivery [123,124], underscoring the broad therapeutic potential of EV-associated miRNAs in corneal health and disease. Mesenchymal stem cell (MSC)-derived exosomes, in particular, have been highlighted for their immunomodulatory, anti-inflammatory, and anti-angiogenic properties, which are beneficial in treating corneal diseases such as scarring and alkali burns [124,125,126]. Studies demonstrate that MSC-derived exosomes enhance corneal re-epithelialization, reduce inflammation, and inhibit neovascularization, thereby promoting repair and minimizing scar formation [126,127]. Additionally, these vesicles, particularly exosomes, facilitate intercellular communication within the limbal niche, influencing both epithelial and stromal cells. For instance, exosomes derived from normal limbal epithelial cells (N-LEC) have been shown to enhance wound healing and proliferation in limbal stromal cells (LSCs) more effectively than those from diabetic sources, suggesting that the miRNA and protein cargo of these vesicles are critical in modulating cellular responses and maintaining corneal function [128]. Similarly, exosomes from normal limbal stromal cells (N-Exos) significantly improve the migration, proliferation, and marker expression of limbal epithelial cells (LECs) by activating pathways such as Akt signaling, which is less pronounced in diabetic-derived exosomes [105]. Furthermore, corneal stromal stem cell-derived EVs (CSSC-EVs) have been found to enhance the stemness of limbal epithelial stem cells (LESCs) by targeting the Notch pathway, thereby promoting proliferation and maintaining stem cell characteristics [27]. The proteomic profiling of small EVs from limbal mesenchymal stromal cells (LMSC) also indicates their role in extracellular matrix deposition and cell matrix adhesion, which are vital for the structural integrity and function of the limbal niche [129]. Additionally, mesenchymal stem cell-derived EVs (MSC-EVs) have demonstrated potential in promoting corneal epithelial repair and modulating inflammation and apoptosis, further underscoring their therapeutic potential in corneal regeneration [101,121,130]. These findings collectively highlight the significant role of EVs in regulating the limbal stem cell niche, offering insights into potential therapeutic strategies for conditions like limbal stem cell deficiency (LSCD) and other corneal disorders [131,132].
In summary, EV-associated miRNAs are vital regulators of corneal biology, governing critical processes such as wound healing, inflammation, immune modulation, and intercellular communication. Their selective packaging and functional diversity position them as promising biomarkers and cell-free therapeutics for corneal disorders. Future research should continue to elucidate the specific roles of individual miRNAs within EVs (Table 3) and optimize delivery platforms to translate these insights into clinical interventions.
Table 3.
Summary of Extracellular Vesicle (EV)-associated microRNAs identified in corneal-associated disorders, indicating their cellular origin, pathological context, validated molecular targets or pathways, and key literature references.
Table 3.
Summary of Extracellular Vesicle (EV)-associated microRNAs identified in corneal-associated disorders, indicating their cellular origin, pathological context, validated molecular targets or pathways, and key literature references.
| miRNAs | Source Cell | Translational Status | Disease | Target/Pathway | References |
|---|---|---|---|---|---|
| miR-328-3p, miR-532-5p, miR-424-5p. let-7c-5p, miR-665 | Human Corneal stromal cell exosomes | Preclinical | Keratoconus | (unknown) | [118] |
| miR-4466 | Human Corneal stromal exosomes (in vitro); human tears (clinical) | Preclinical | Keratoconus | (unknown; linked to epithelial integrity) | [133] |
| let-7b-5p | Human Corneal Epithelial cell exosomes → macrophages (in vitro) | Preclinical | Fungal keratitis (Aspergillus infection) | Targets SOCS-1; promotes macrophage M1 activation | [134] |
| miR-24-3p | Rabbit ADSC exosomes | Preclinical | Corneal alkali burn/epithelial injury | Upregulates CDC42, EGFR, MMP9; promotes migration | [135] |
| miR-21-5p | Human UMSC exosomes | Preclinical | Corneal epithelial wounding (mechanical) | Downregulates PTEN; activates PI3K/Akt | [102] |
| miR-29b-3p | Mouse BMSC exosomes | Preclinical | General corneal injury (inflammation) | Activates autophagy (↓ PI3K/Akt/mTOR); inhibits NF-κB | [136] |
| miR-19a-3p | Rabbit ADSC exosomes | Preclinical | Corneal stromal fibrosis | Targets HIPK2; anti-fibrotic (↓ collagens, α-SMA) | [137] |
| miR-204-5p | Human (UC-MSC)/Mouse (BMSC) exosomes | Preclinical + Clinical | GVHD-associated dry eye (autoimmune DED) | Targets IL-6/IL-6R/Stat3; anti-inflammatory | [138] |
| miR-223-3p | Mouse adipose MSC exosomes | Preclinical | Dry eye (BAC/scopolamine model) | Targets Fbxw7; anti-inflammatory | [139] |
| miR-127-5p, miR-1273h-3p, miR-1288-5p, miR-130b-5p, miR-139-3p, miR-1910-5p, miR-203b-5p, miR-22-5p, miR-4632-3p | Human tear EVs (clinical study) | Clinical | Non-SS dry eye (tear EVs) | Inflammation-associated (ingenuity analysis) | [107] |
6. Tailored Extracellular Vesicles (TeVs) in Cornea
Engineered EVs have emerged as a promising tool for the treatment of corneal diseases and drug delivery, leveraging their unique properties such as biocompatibility, low immunogenicity, and ability to traverse biological barriers. These vesicles can carry a diverse array of bioactive cargo, including proteins, RNAs, and lipids, facilitating intercellular communication and therapeutic delivery [140]. In the context of corneal diseases, EVs have been particularly effective due to their ability to modulate angiogenesis, lymphangiogenesis, and nerve repair, which are critical processes in corneal healing and regeneration [15]. For instance, platelet-derived extracellular vesicles (PEVs) have been engineered to deliver anti-angiogenic agents like kaempferol, demonstrating significant efficacy in reducing corneal neovascularization in preclinical models [141]. This is achieved through the enhanced internalization of PEVs by corneal cells, leading to the downregulation of angiogenesis-related gene expression and reduced inflammatory cytokine levels [141]. Moreover, the use of EVs has been explored for their potential in corneal repair and regeneration, given their ability to carry therapeutic molecules and their minimal immunogenicity when derived from the patient’s own cells [15,16]. The engineering of EVs to include intelligent cores that enable targeted delivery and controlled release further enhances their therapeutic potential, making them a versatile platform for next-generation nanomedicine [142]. Despite these advancements, challenges remain in optimizing the large-scale production and precise targeting of engineered EVs, which are crucial for their successful clinical translation [143]. Nonetheless, the ongoing research and development in this field hold promise for revolutionizing the treatment of corneal diseases and improving drug delivery systems across various medical applications [144,145]. Challenges with the rapid clearance of EVs from the corneal surface necessitate innovative delivery methods. For instance, Lybecker et al. proposed a sustained release platform utilizing biocompatible collagen-based hydrogels to enhance the therapeutic efficacy of corneal epithelial-derived EVs by allowing localized and prolonged delivery [146]. The integration of hydrogels with EVs not only improves drug delivery efficiency but also provides a supportive scaffold that mimics the native extracellular matrix, enhancing cellular interactions during healing [147,148]. For instance, MSC-exos have been shown to effectively deliver active β-glucuronidase to corneal cells, facilitating the catabolism of accumulated glycosaminoglycans, which is beneficial in treating Mucopolysaccharidosis VII (Sly Syndrome) [125]. In the context of dry eye disease, exosomes coupled with ascorbic acid have been developed into a multifunctional eyedrop that improves corneal epithelium recovery and reduces oxidative stress and inflammation, showcasing their potential in treating complex ocular surface diseases [149]. Furthermore, exosomes have been employed to deliver siRNA targeting NF-κB c-Rel, effectively accelerating corneal wound healing by reducing inflammation [150]. The use of exosomes as carriers for microRNA-29b has also been explored, where they activate autophagy and inhibit inflammation and fibrosis in corneal injuries, highlighting their role in cellular repair mechanisms [136]. Moreover, engineered exosomes loaded with anti-TNF-α antibodies have shown superior efficacy in mitigating corneal injuries by enhancing wound healing and reducing inflammation, particularly when delivered via microneedles for sustained release [151].
Moreover, authors propose a modular, stimuli-responsive EV platform that is engineered, validated, and delivered in three integrated steps. First, decorate MSC-derived EVs with RGD peptides for αvβ3-mediated keratocyte targeting and embed pH-sensitive polymer cores for on-demand anti-angiogenic miRNA release in inflamed stroma. Second, load these EVs into hyaluronic-acid contact lenses featuring micro-reservoirs that dispense therapeutic cargo over days, synchronizing with wound-healing phases and eliminating repeated dosing. Third, manufacture EVs in a microfluidic bioreactor under defined shear and oxygenation, coupled to inline Raman spectroscopy for real-time quality control, and refine our approach with fully synthetic exosome-mimetics to guarantee batch consistency. This active, end-to-end strategy maximizes corneal targeting, sustains release, and restores transparency in a single, patient-friendly device.
7. Corneal Disease Detection via EV-miRNA Signatures
EV-associated microRNAs (miRNAs) represent a promising, noninvasive diagnostic modality for corneal disorders. Indeed, mesenchymal stem cell-derived EVs deliver specific miRNAs that attenuate fibrosis and inflammation in preclinical models, thereby underscoring their diagnostic and therapeutic relevance in corneal disease [21]. Furthermore, altering the miRNA cargo within EVs modulates corneal repair and inflammatory pathways, which has important implications for monitoring disease progression. High-throughput resources such as EV-miRNA curate expression profiles from diverse biofluids, facilitating the identification of candidate biomarkers in ocular pathologies [152]. For instance, Fuchs’ endothelial corneal dystrophy (FECD) is characterized by distinct aqueous humor signatures, including pronounced overexpression of hsa-miR-4532 and hsa-miR-6800-5p, indicating their potential as disease-specific indicators [153]. In addition, profiling EV miRNAs in tears and aqueous humor using next-generation sequencing and qRT-PCR enables noninvasive detection of pathological states and elucidates underlying mechanisms [21,154]. Common workflows leverage real-time PCR arrays and microarray systems to quantify miRNAs from biofluids, achieving high classification accuracy in distinguishing normal and disease conditions [153,155]. Clinically, EV-associated miRNAs are being explored in liquid biopsy formats for early diagnosis and personalized management of ocular diseases, capitalizing on their stability and characteristic expression patterns [144]. Moreover, emerging diagnostics for keratoconus and other corneal dystrophies integrate machine-learning algorithms with imaging modalities such as anterior segment OCT and corneal topography to correlate miRNA profiles with morphological and biomechanical changes, thereby enhancing early detection [154,156]. Correlative studies have further linked specific EV miRNA levels to keratoconus severity, supporting their suitability as staging biomarkers [21,154], and in vivo confocal microscopy offers complementary structural mapping of miRNA-associated tissue alterations [157]. Additionally, tear-derived EV miRNAs, such as miR-151-5p and miR-422a, demonstrate diagnostic relevance beyond the cornea, notably in retinal diseases [155]. Despite these advances, standardization of miRNA isolation and quantification protocols, alongside rigorous validation of signature panels, remains essential for clinical translation [144,158] (Figure 3).
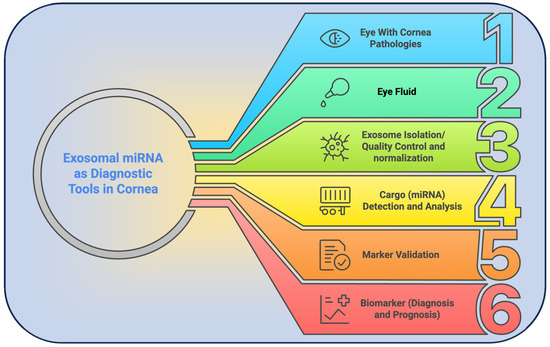
Figure 3.
Schematic overview of the exosomal miRNA–based diagnostic pipeline for corneal disorders. (1) Clinical identification of patients with corneal pathology; (2) tear fluid or tissue sampling; (3) extracellular vesicle isolation; (4) extraction and profiling of miRNA cargo; (5) validation of disease-specific miRNA markers; and (6) translation of validated miRNA signatures into diagnostic and prognostic assays.
Integration of microfluidic tear-chip technology with AI/ML algorithms substantially enhances detection and analysis of extracellular vesicle microRNA (EV-miRNA) profiles by combining the precision and throughput of microfluidics with the analytical power of artificial intelligence. Microfluidic systems provide a streamlined, rapid, and sensitive approach for EV extraction, lysis, and miRNA quantification, reducing the time and labor demands of conventional methods [159]. These platforms can reach detection limits in the femtomolar range, which is necessary for physiological miRNA concentrations, and can process samples in as little as 30 min [160]. The integration with AI/ML further expands capabilities by enabling analysis of complex multiomics datasets, improving molecular classification, and facilitating personalized therapeutic strategies [161]. AI models can optimize the miRNA feature space to construct robust predictive models for disease staging and progression [162]. Machine learning-based detection schemes support multiplexed classification of miRNA targets, delivering fast and cost-effective results without extensive sample processing [163]. This synergy accelerates diagnostics while improving the accuracy and reliability of miRNA profiling [164,165], and it supports the development of high-performance, point-of-care solutions that are accessible and informative for clinical decision-making [162]. Additionally, applied to corneal disease diagnosis and therapy, microfluidic platforms offer rapid, sensitive, and low-sample-requirement quantification of EV-miRNAs relevant to ocular pathology; for example, platforms such as that developed by Ramshani et al. enable absolute miRNA quantification with high sensitivity and minimal sample volumes, reducing the complexity associated with RT-qPCR [160]. Coupling these systems with AI automates data analysis and improves decision-making by processing the large datasets generated by high-throughput microfluidics, thereby enabling rapid identification and characterization of disease-relevant miRNA signatures [166]. In ophthalmology, AI has already demonstrated strong performance in diagnosing and managing corneal conditions from imaging data, in some cases matching or exceeding expert accuracy [167,168]. Furthermore, AI-powered microfluidic workflows can enable point-of-care miRNA detection and monitoring of therapeutic responses in corneal disease settings [169]. To ensure clinical translation, these approaches must address population heterogeneity and prioritize explainable AI models that support clinical applicability and trust [170,171].
To operationalize these discoveries, we propose a comprehensive platform wherein EVs are isolated from tear fluid and aqueous humor for rapid multiplex digital PCR quantification of a targeted miRNA panel, including miR-4532, miR-6800-5p, miR-151-5p, and miR-422a, and integrated with anterior segment OCT and in vivo confocal microscopy data. Thereafter, machine-learning models correlate molecular signatures with structural and biomechanical parameters to facilitate early detection of keratoconus, FECD, and postoperative fibrosis. To ensure reproducibility across centers, a microfluidic “tear chip” design could perform on-chip EV capture, lysis, and miRNA analysis within 30 min, backed by a centralized reference database of healthy and diseased profiles. By shifting corneal care from reactive treatment to predictive monitoring, this end-to-end diagnostic solution promises personalized interventions before irreversible tissue damage occurs.
8. Therapeutic Implications of EV-Associated miRNAs
EVs act as carriers of microRNAs (miRNAs), mediating intercellular communication and signaling pathways that regulate both physiological and pathological processes in the cornea. By binding to the 3′ untranslated regions of target mRNAs, EV-derived miRNAs induce translational repression or mRNA degradation [172]. Furthermore, their encapsulation within EVs shields miRNAs from extracellular RNases, thereby enhancing stability and bioavailability [173,174]. Specifically, miRNAs from corneal stromal stem cells (CSSCs) have demonstrated anti-inflammatory and regenerative effects, promoting scarless wound healing through modulation of fibrosis-related gene expression [4,21].
Preclinical models have substantiated the therapeutic potential of EV-associated miRNAs in corneal disorders. For instance, mesenchymal stem cell-derived EVs enriched with select miRNAs markedly reduced corneal fibrosis and inflammation in murine injury models [21]. Critically, knockdown of Alix, an ESCRT component essential for miRNA incorporation, diminished EV miRNA content and abolished their regenerative effects [21]. Similarly, human amniotic epithelial cell-derived EVs accelerated corneal repair by enhancing epithelial cell proliferation and migration via miRNA-dependent mechanisms [4]. These outcomes have been elucidated across in vitro, ex vivo, and in vivo systems [175]. In vitro scratch assays using HCE-T cells confirm EV uptake and accelerated epithelial gap closure [175]. In vivo, mouse models of alkali burn, or LPS-induced inflammation, reveal that topically or locally administered fluorescently labeled EVs reach corneal tissues, improving clarity, reducing opacity, attenuating inflammation, and inhibiting neovascularization [21,175]. Moreover, omics-based approaches, including RNA sequencing and microarrays, enable comprehensive profiling of EV miRNA cargo and downstream transcriptomic changes [175]. Beyond CSSC- and bone marrow MSC-derived EVs, which exhibit antifibrotic, anti-inflammatory, and regenerative properties yet require further mechanistic insight [101,176], specific miRNAs such as miR-145 have emerged as key regulators of myofibroblast differentiation, thereby preventing scarring [177]. However, challenges include an incomplete understanding of miRNA-mediated pathways [176] and rapid ocular clearance necessitates sustained delivery systems; promising solutions involve the incorporation of EVs into collagen-based hydrogels to prolong retention and release [146].
Advances in miRNA-based therapeutics encompass administration of synthetic mimics or inhibitors targeting fibrosis and inflammatory pathways to restore corneal homeostasis [172]. In parallel, genetic and bioengineering strategies facilitate exogenous loading of therapeutic miRNAs into isolated EVs using methods with representative efficiency, such as passive incubation, co-incubation (1–5%), electroporation (10–60%), sonication (10–40%), extrusion (10–50%), and freeze–thaw cycles (5–25%) [178,179] or manipulation of parent stem cells to enrich desired miRNAs naturally [175,180]. Targeted delivery is further enhanced by engineering EV surface proteins for improved tissue specificity [181] and embedding EVs within decellularized extracellular matrix hydrogels, which sustain and localize miRNA release at injury sites [182]. Candidate miRNAs with demonstrated relevance include miR-21 (wound healing), miR-204 (angiogenesis inhibition), miR-145 (fibrosis suppression), miR-155-5p (inflammation control), and miR-29b-3p (neuroprotection and anti-fibrosis) [21,25,183]. Engineering EVs to deliver these miRNAs or their inhibitors enables precise modulation of pathological processes (Figure 4). In summary, EV-associated miRNAs constitute a modular, cell-free platform for corneal repair and homeostasis restoration. In our hands, EVs isolated from mesenchymal or corneal stromal stem cells and loaded with fibrosis-suppressing miRNAs (for example, miR-145 and miR-29b-3p) or anti-inflammatory miR-155-5p reliably accelerate epithelial gap closure in scratch assays and reduce stromal opacity in alkali-burned mouse corneas.
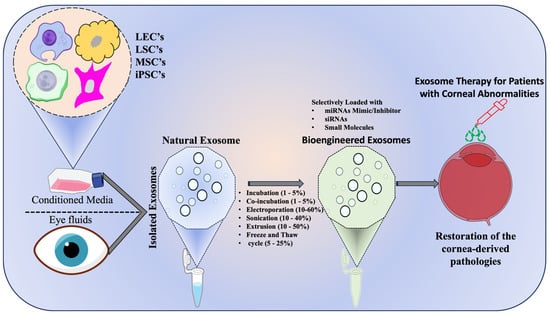
Figure 4.
Schematic representation of exosome mediated therapy for corneal disorders- Conditioned media from ocular cell sources—limbal epithelial cells (LECs), limbal stromal cells (LSCs), mesenchymal stem cells (MSCs), and induced pluripotent stem cells (iPSCs) is processed to isolate native exosomes. These vesicles are subsequently loaded with therapeutic cargo (miRNA mimics or inhibitors, siRNAs, and small molecules) using bioengineering methods and representative loading efficiencies are indicated such as passive incubation (1–5%), co-incubation (1–5%), electroporation (10–60%), sonication (10–40%), extrusion (10–50%), and freeze–thaw cycles (5–25%). The engineered exosomes are then administered to the diseased cornea to modulate cellular pathways, attenuate inflammation, and promote tissue repair, highlighting their translational potential in ocular regenerative medicine.
To translate these findings into scalable therapies, we propose three innovations. First, we will integrate EV capture, on-chip lysis, and droplet-digital PCR into a contact-lens-like microfluidic device that continuously releases miRNA-loaded EVs at the ocular surface while monitoring local miRNA levels. Second, engineer parent MSCs with light-responsive RNA switches that trigger miRNA enrichment on demand, enabling spatiotemporal control of cargo loading. Third, we will embed EVs into an in situ-forming, decellularized extracellular-matrix hydrogel tailored for corneal curvature, which prolongs miRNA release and overcomes rapid tear clearance. Finally, we will standardize potency assays by quantifying target gene knockdown in ex vivo human corneal lenticules and by correlating functional endpoints, wound closure rate, α-SMA expression, and vascular ingrowth with specific miRNA signatures. Establishing GMP-compliant bioreactors for EV production, coupled with quantitative release assays and scalable hydrogel molds, will streamline our path to Phase I trials. Collectively, these approaches promise a versatile, predictive, and patient-adaptable strategy to treat fibrosis, inflammation, and angiogenesis in human corneal disease.
9. Roadblocks to Breakthroughs
Regulatory frameworks originally formulated for small molecules and biologics improperly capture the complexity and variability of extracellular vesicle-associated miRNAs (EV-miRNAs), thereby impeding their clinical translation [184,185]. Moreover, the absence of clear guidelines on characterization, isolation, and analytical protocols further obstructs reproducibility and safety assessments in clinical applications [185,186] and underscores an urgent need for standardized methodologies [184,185,187]. A critical bottleneck lies in defining and controlling critical quality attributes such as purity, cellular origin, and consistent manufacturing processes, which must accommodate the inherent heterogeneity of EV populations and their variable miRNA cargo [188,189,190]. Consequently, regulatory policies must evolve to balance innovation with patient safety [188], particularly given the rigorous approval pathway established by advanced cell therapies in ophthalmology, like expanded human corneal epithelial cells [188]. Despite compelling evidence that EV can mitigate inflammation and foster corneal regeneration, the lack of harmonized, ophthalmology-specific guidelines continues to pose a significant translational barrier [176,187].
In parallel, technical and standardization challenges further widen this regulatory gap. Isolation techniques including ultracentrifugation (UC), size-exclusion chromatography (SEC), ultrafiltration (UF), polymer-based precipitation, and microfluidic immunoaffinity capture yield inconsistent vesicle recovery, purity, and cargo integrity, complicating both research interpretation and clinical outcomes [84,191,192]. Quality control assays such as Nanoparticle Tracking Analysis, Resistive Pulse Sensing, and flow cytometry vary in accuracy and throughput, while RNA quantification methods qRT-PCR, next-generation sequencing (NGS), and digital droplet PCR (ddPCR) face distinct limitations in normalization, input requirements, and contamination control [84,192,193,194,195,196]. For example, UC can damage vesicles and reduce yield, precipitation methods often co-precipitate protein and polymer contaminants, SEC dilutes samples necessitating reconcentration, UF risks non-specific membrane binding, and microfluidic platforms may alter cargo through antibody interactions [84,197,198]. Furthermore, variability in instrument settings impacts NTA sizing, RPS demands multiple pore sizes to avoid clogging, and flow cytometry requires stringent controls for reliable marker detection [84]. Although qRT-PCR offers low-input sensitivity, it lacks universal endogenous controls [84,196], NGS delivers comprehensive profiling at high cost and with rRNA contamination challenges [84,199], and ddPCR enables absolute quantification but depends on precise partitioning [84]. Additionally, disparate RNA purification kits yield inconsistent RNA size distributions and may co-purify surface-bound nucleic acids unless pretreated with proteinase and RNase [84]. Together, these methodological disparities generate pronounced variability and impede reproducibility across studies, underscoring the imperative for standardization [84,189,199,200,201]. Accordingly, initiatives such as the Minimal Information for Studies of Extracellular Vesicles (MISEV) guidelines and the EV-TRACK platform seek to harmonize reporting standards and data sharing [189,202].
Furthermore, ensuring the safety of EV-miRNA therapies demands thorough evaluation of immunogenicity and off-target risks [203,204,205,206,207]. Although EVs typically exhibit low immunogenicity and toxicity compared to whole-cell treatments, a vesicle’s cellular origin can modulate immune responses; for instance, antigen-presenting cell-derived vesicles may activate T cells, warranting detailed immunological assessment [190,208,209]. Off-target effects represent a critical safety challenge, since miRNAs demonstrate partial complementarity to multiple mRNAs and can inadvertently repress non-target genes [176,203,210]. While engineering targeting motifs onto EV surfaces may enhance delivery specificity, such modifications risk altering vesicle cargo and function, necessitating additional validation [190,211]. Moreover, rigorous screening is required to exclude immunogenic, oncogenic, or infectious contaminants, and the long-term impacts of EV-miRNA administration remain to be fully characterized [203,207]. Indeed, emerging data suggest that only a small subset of EVs carry functional miRNAs, raising questions about cargo protection from extracellular nucleases, particularly for surface-associated miRNAs [212,213]. Therefore, despite their biodegradability and generally low baseline immunogenicity, continuous and stringent safety assessments are essential to advance EV-miRNA therapeutics toward clinical reality [208].
Concurrently, robust quality control (QC) measures are indispensable for the consistent and reliable application of EV-derived miRNAs in research and clinical settings [190,202]. A key challenge lies in ensuring batch-to-batch consistency and rigorous purity assessment of EV preparations [190,214,215], since contamination by non-EV components such as proteins, lipoproteins, and extracellular nucleic acids can significantly confound miRNA profiling and functional assays [84,198]. Moreover, although the particle-to-protein ratio (particles per microgram of protein) serves as a common purity metric, this parameter varies substantially with source and extraction method [84]. Consequently, integrating QC steps throughout the experimental workflow is essential to mitigate variability [214]. In the absence of universal endogenous controls for EV-derived RNAs, accurate miRNA quantification relies on spike-in controls [84,216] or the use of stable reference genes like miR-22-5p and miR-29a-5p in specific cell types [85,217]. Moreover, the choice of isolation technique profoundly influences vesicle recovery, purity, and downstream miRNA analysis [198,218]; for example, SEC can enhance EV purity and miRNA yield, although non-EV particles may still affect results [198]. Batch effects arising from divergent RNA isolation protocols or sequencing platforms further underscore the need for careful monitoring of quality parameters and bias-reduction methods [200,219]. Although emerging approaches such as rolling circle amplification within encoded hydrogel microparticles offer heightened sensitivity and absolute quantification of EV-miRNAs, the field continues to lack comprehensive reference materials and standardized analytical techniques for assessing particle size distribution, concentration, and cargo homogeneity [190].
Finally, these regulatory, technical, and safety hurdles have direct implications for clinical trial design in EV-derived miRNA therapies, as inherent disease heterogeneity and variability in EV sources and their miRNA cargo [187,190,200] complicate patient selection, endpoint definition, and regulatory approval processes [189,220]. Specifically, factors such as patient age, gender, ethnicity, disease stage, medication use, lifestyle, and dietary habits can alter both the abundance and composition of biofluid-derived EVs, necessitating rigorous normalization across study cohorts [84]. For example, distinct EV-miRNA expression profiles can distinguish lung adenocarcinoma from granuloma, demonstrating both the biomarker potential of EVs and the imperative for precise patient stratification [221]. Moreover, defining sensitive and specific clinical endpoints ranging from quantification of target miRNAs within EVs to relevant functional outcomes and composite clinical measures remains challenging given the novelty and complexity of EV-miRNA biology [220]. Therefore, well-designed multicenter trials are essential to validate the therapeutic efficacy of EV-based interventions and to confirm the utility of associated biomarkers [222].
Fundamentally, we propose that the field unite around an integrated roadmap to accelerate EV translation. First, partner with regulatory agencies and key stakeholders to draft harmonized, EV-specific guidelines that define critical quality attributes, acceptable variability thresholds, and streamlined approval pathways. Second, spearhead international consortia to validate and disseminate standardized protocols for EV isolation, characterization, and miRNA quantification, complete with universally accepted reference materials and normalization controls. Third, build a modular safety-assessment platform that combines high-throughput immunogenicity assays, off-target profiling, and long-term biodistribution studies. Finally, design adaptive, biomarker-driven clinical trials with well-stratified patient cohorts and clear functional endpoints, thereby ensuring both scientific rigor and clinical relevance. Collectively, these efforts will bridge current gaps, safeguard patient outcomes, and unlock the therapeutic promise of EV-miRNAs.
10. Conclusions
Extracellular vesicle-associated microRNAs have emerged as critical regulators of corneal physiology and disease, offering both mechanistic insights and practical applications. Their selective packaging and remarkable stability enable minimally invasive biomarker discovery in tears and aqueous humor, facilitating early detection, risk stratification, and dynamic monitoring of corneal disorders, while engineered EVs loaded with miRNA mimics or inhibitors demonstrate potent anti-inflammatory, antifibrotic, and regenerative effects in preclinical models. Moreover, the integration of microfluidic isolation platforms, artificial intelligence-driven biomarker profiling, and precision engineering of vesicle cargo affords unprecedented sensitivity and specificity in both diagnosis and targeted modulation of pathological pathways. However, translation to the clinic is impeded by technical variability in EV isolation, miRNA quantification, and functional assays, as well as by gaps in regulatory frameworks tailored to complex biological nanotherapeutics. To overcome these obstacles, harmonized standards for EV characterization, cargo loading, and safety testing must be established alongside the development of sustained-release delivery systems such as hydrogel scaffolds or contact-lens-integrated reservoirs to overcome rapid ocular clearance and ensure scalable manufacturing. Advances in targeted surface modification and patient-specific EV sourcing will further enhance tissue tropism and minimize off-target effects. Looking forward, the formation of an international consortium comprising academic investigators, industry partners, and regulatory bodies will be essential to co-develop standardized reporting guidelines, harmonized quality criteria, and adaptive clinical trial designs that integrate molecular and imaging biomarkers. By addressing these technical, regulatory, and biological challenges in a unified translational pipeline, EV-mediated miRNA communication can fulfill its promise as a versatile paradigm for personalized diagnostics and therapeutics, ultimately revolutionizing early diagnosis, management, and restoration of vision in patients with corneal disease.
Author Contributions
Conceptualization, N.V.; investigation, N.V.; resources, S.A.; data curation, A.K.; writing—original draft preparation, N.V.; writing—review and editing, S.A. and A.K.S.; visualization, A.K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data were used for the research described in the article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| EV | Extracellular Vesicles |
| FECD | Fuchs endothelial corneal dystrophy |
| LSCD | Limbal Stem Cell Deficiency |
| DLS | Dynamic Light Scattering |
| NTA | Nanoparticle Tracking Analysis |
| LECs | Limbal Epithelial Cells |
| LSCs | Limbal Stromal Cells |
| iPSCs | Induced Pluripotent Stem Cells |
| CSSCs | Corneal Stromal Stem Cells |
| LESCs | Limbal Epithelial Stem Cells |
| LMSCs | Limbal Mesenchymal Stromal Cells |
| MSCs | Mesenchymal Stem Cells |
| AI | Artificial Intelligence |
| ML | Machine Learning |
| OCT | Optical Coherence Tomography |
| UCMSCs | Umbilical Cord-derived Mesenchymal Stem Cells |
| BMSCs | Bone Marrow Stem Cells |
| ADSCs | Adipose-Derived Stem Cells |
References
- Raj, N.; Gupta, N.; Kumar, D.; Vashist, P.; Tandon, R. Population-based study on the prevalence, clinical characteristics and vision-related quality of life in patients with corneal opacity resulting from infectious keratitis: Results from the Corneal Opacity Rural Epidemiological study. Br. J. Ophthalmol. 2023, 107, 476–482. [Google Scholar] [CrossRef]
- Gupta, N.; Vashist, P.; Tandon, R.; Gupta, S.K.; Dwivedi, S.; Mani, K. Prevalence of corneal diseases in the rural Indian population: The Corneal Opacity Rural Epidemiological (CORE) study. Br. J. Ophthalmol. 2015, 99, 147–152. [Google Scholar] [CrossRef]
- Tidke, S.C.; Tidake, P. A Review of Corneal Blindness: Causes and Management. Cureus 2022, 14, e30097. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, Z.; Jin, C.; Chen, Q.; Fang, Y.; Jin, J.; Chen, J.; Lu, L.; Tian, H.; Xu, J.; et al. Human amniotic epithelial cell-derived extracellular vesicles provide an extracellular matrix-based microenvironment for corneal injury repair. J. Tissue Eng. 2022, 13, 20417314221122123. [Google Scholar] [CrossRef] [PubMed]
- McKay, T.B.; Yeung, V.; Hutcheon, A.E.K.; Guo, X.; Zieske, J.D.; Ciolino, J.B. Extracellular Vesicles in the Cornea: Insights from Other Tissues. Anal. Cell. Pathol. 2021, 2021, 9983900. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, D.K.; Chirikov, V.; Schmier, J.; Rege, S.; Newton, S. Cost Burden of Endothelial Keratoplasty in Fuchs Endothelial Dystrophy: Real-World Analysis of a Commercially Insured US Population (2014–2019). Clin. Ophthalmol. 2022, 16, 1055–1067. [Google Scholar] [CrossRef]
- Lin, Y.; Anderson, J.D.; Rahnama, L.M.A.; Gu, S.V.; Knowlton, A.A. Exosomes in disease and regeneration: Biological functions, diagnostics, and beneficial effects. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H1162–H1180. [Google Scholar] [CrossRef]
- Di Bella, M.A. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Z.; Gu, X.; Sheng, X.; Xiao, L.; Wang, X. Exosomes: New regulators of reproductive development. Mater. Today Bio 2023, 19, 100608. [Google Scholar] [CrossRef]
- Sun, B.; Peng, J.; Wang, S.; Liu, X.; Zhang, K.; Zhang, Z.; Wang, C.; Jing, X.; Zhou, C.; Wang, Y. Applications of stem cell-derived exosomes in tissue engineering and neurological diseases. Rev. Neurosci. 2018, 29, 531–546. [Google Scholar] [CrossRef]
- Lasser, C. Exosomes in diagnostic and therapeutic applications: Biomarker, vaccine and RNA interference delivery vehicle. Expert Opin. Biol. Ther. 2015, 15, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Nazarenko, I.; Rupp, A.-K.; Altevogt, P. Exosomes as a Potential Tool for a Specific Delivery of Functional Molecules. In Ovarian Cancer: Methods and Protocols; Malek, A., Tchernitsa, O., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 495–511. [Google Scholar]
- Kushch, A.A.; Ivanov, A.V. Exosomes in the life cycle of viruses and the pathogenesis of viral infections. Vopr. Virusol. 2023, 68, 181–197. [Google Scholar] [CrossRef]
- Desjardins, P.; Berthiaume, R.; Couture, C.; Le-Bel, G.; Roy, V.; Gros-Louis, F.; Moulin, V.J.; Proulx, S.; Chemtob, S.; Germain, L.; et al. Impact of Exosomes Released by Different Corneal Cell Types on the Wound Healing Properties of Human Corneal Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 12201. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.; Chen, V.T.; Herbst, P.; Zhang, R.; Elfert, A.; Krishan, A.; Azar, D.T.; Chang, J.H.; Hu, W.Y.; Kremsmayer, T.P.; et al. Target specification and therapeutic potential of extracellular vesicles for regulating corneal angiogenesis, lymphangiogenesis, and nerve repair. Ocul. Surf. 2024, 34, 459–476. [Google Scholar] [CrossRef]
- Robbins, B.T.; Montreuil, K.A.; Kundu, N.; Kumar, P.; Agrahari, V. Corneal Treatment, Repair, and Regeneration: Exosomes at Rescue. Pharmaceutics 2024, 16, 1424. [Google Scholar] [CrossRef]
- Yu, H.; Wu, J.; Pan, G. Targeting the Ophthalmic Diseases Using Extracellular Vesicles ‘Exosomes’: Current Insights on Their Clinical Approval and Present Trials. Aging Dis. 2024, 16, 1225–1241. [Google Scholar] [CrossRef]
- Verma, N.; Arora, S.; Singh, A.K.; Ahmed, J. Unlocking the potential of exosomes ‘extracellular vesicles’: Drug delivery advancements and therapeutics in ocular diseases. RSC Pharm. 2025. [Google Scholar] [CrossRef]
- Tiwari, A.; Singh, A.; Verma, S.; Stephenson, S.; Bhowmick, T.; Sangwan, V.S. Mini Review: Current Trends and Understanding of Exosome Therapeutic Potential in Corneal Diseases. Front. Pharmacol. 2021, 12, 684712. [Google Scholar] [CrossRef]
- Blanco-Agudín, N.; Ye, S.; González-Fernández, S.; Alcalde, I.; Merayo-Lloves, J.; Quirós, L.M. Exosomes in Ocular Health: Recent Insights into Pathology, Diagnostic Applications and Therapeutic Functions. Biomedicines 2025, 13, 233. [Google Scholar] [CrossRef] [PubMed]
- Shojaati, G.; Khandaker, I.; Funderburgh, M.L.; Mann, M.M.; Basu, R.; Stolz, D.B.; Geary, M.L.; Dos Santos, A.; Deng, S.X.; Funderburgh, J.L. Mesenchymal Stem Cells Reduce Corneal Fibrosis and Inflammation via Extracellular Vesicle-Mediated Delivery of miRNA. Stem Cells Transl. Med. 2019, 8, 1192–1201. [Google Scholar] [CrossRef]
- Buono, L.; Scalabrin, S.; De Iuliis, M.; Tanzi, A.; Grange, C.; Tapparo, M.; Nuzzi, R.; Bussolati, B. Mesenchymal Stem Cell-Derived Extracellular Vesicles Protect Human Corneal Endothelial Cells from Endoplasmic Reticulum Stress-Mediated Apoptosis. Int. J. Mol. Sci. 2021, 22, 4930. [Google Scholar] [CrossRef]
- Roszkowska, A.M.; Aguennouz, M.; Aragona, E.; Gargano, R.; Oliverio, G.W.; Inferrera, L.; Aragona, P. Extensive Contact Lens Wear Modulates Expression of miRNA-320 and miRNA-423-5p in the Human Corneal Epithelium: Possible Biomarkers of Corneal Health and Environmental Impact. Genes 2024, 15, 816. [Google Scholar] [CrossRef]
- Sanroque-Muñoz, M.; Garcia, S.G.; Clos-Sansalvador, M.; Font-Morón, M.; Botella-Garcia, J.; Franquesa, M.; Loscos-Arenas, J.; Borràs, F.E. miRNAs from tear fluid-derived extracellular vescicles to identify patients at high risk of fibrotic lesions after glaucoma surgery. Acta Ophthalmol. 2025, 103. [Google Scholar] [CrossRef]
- Arora, S.; Verma, N. Exosomal microRNAs as potential biomarkers and therapeutic targets in corneal diseases. Mol. Vis. 2024, 30, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, Y.; Luo, S.; Yang, C.; Zhong, G.; Huang, G.; Zhang, X.; Li, B.; Liu, C.; Li, L.; et al. DNA Nanowire Guided-Catalyzed Hairpin Assembly Nanoprobe for In Situ Profiling of Circulating Extracellular Vesicle-Associated MicroRNAs. ACS Sens. 2022, 7, 1075–1085. [Google Scholar] [CrossRef]
- Wang, L.; Xu, X.; Chen, Q.; Wei, Y.; Wei, Z.; Jin, Z.B.; Liang, Q. Extracellular Vesicle MicroRNAs From Corneal Stromal Stem Cell Enhance Stemness of Limbal Epithelial Stem Cells by Targeting the Notch Pathway. Investig. Ophthalmol. Vis. Sci. 2023, 64, 42. [Google Scholar] [CrossRef]
- Ahmed, H.M.; Salem, N.M.; Al-Atabany, W. Impact of Post–Refractive Surgeries on Corneal Biomechanics—A Review. J. Clin. Eng. 2021, 46, 177–188. [Google Scholar] [CrossRef]
- Willoughby, C.E.; Ponzin, D.; Ferrari, S.; Lobo, A.; Landau, K.; Omidi, Y. Anatomy and physiology of the human eye: Effects of mucopolysaccharidoses disease on structure and function—A review. Clin. Exp. Ophthalmol. 2010, 38, 2–11. [Google Scholar] [CrossRef]
- McKay, T.B.; Seyed-Razavi, Y.; Ghezzi, C.E.; Dieckmann, G.; Nieland, T.J.F.; Cairns, D.M.; Pollard, R.E.; Hamrah, P.; Kaplan, D.L. Corneal pain and experimental model development. Prog. Retin. Eye Res. 2019, 71, 88–113. [Google Scholar] [CrossRef]
- Guerrero-Moreno, A.; Baudouin, C.; Melik Parsadaniantz, S.; Reaux-Le Goazigo, A. Morphological and Functional Changes of Corneal Nerves and Their Contribution to Peripheral and Central Sensory Abnormalities. Front. Cell. Neurosci. 2020, 14, 610342. [Google Scholar] [CrossRef]
- Ruan, Y.; Jiang, S.; Musayeva, A.; Pfeiffer, N.; Gericke, A. Corneal Epithelial Stem Cells-Physiology, Pathophysiology and Therapeutic Options. Cells 2021, 10, 2302. [Google Scholar] [CrossRef]
- Sprogyte, L.; Park, M.; Girolamo, N.D. Pathogenesis of Alkali Injury-Induced Limbal Stem Cell Deficiency: A Literature Survey of Animal Models. Cells 2023, 12, 1294. [Google Scholar] [CrossRef]
- Loukovitis, E.; Kozeis, N.; Gatzioufas, Z.; Kozei, A.; Tsotridou, E.; Stoila, M.; Koronis, S.; Sfakianakis, K.; Tranos, P.; Balidis, M.; et al. The Proteins of Keratoconus: A Literature Review Exploring Their Contribution to the Pathophysiology of the Disease. Adv. Ther. 2019, 36, 2205–2222. [Google Scholar] [CrossRef]
- Wisse, R.P.; Kuiper, J.J.; Gans, R.; Imhof, S.; Radstake, T.R.; Van der Lelij, A. Cytokine Expression in Keratoconus and its Corneal Microenvironment: A Systematic Review. Ocul. Surf. 2015, 13, 272–283. [Google Scholar] [CrossRef]
- Navel, V.; Malecaze, J.; Belville, C.; Choltus, H.; Henrioux, F.; Dutheil, F.; Malecaze, F.; Chiambaretta, F.; Blanchon, L.; Sapin, V. Dysregulation of Receptor for Advanced Glycation End Products (RAGE) Expression as a Biomarker of Keratoconus. Dis. Markers 2022, 2022, 1543742. [Google Scholar] [CrossRef] [PubMed]
- Spadea, L.; Salvatore, S.; Vingolo, E.M. Corneal sensitivity in keratoconus: A review of the literature. Sci. World J. 2013, 2013, 683090. [Google Scholar] [CrossRef]
- Okumura, N.; Hayashi, R.; Nakano, M.; Yoshii, K.; Tashiro, K.; Sato, T.; Blake, D.J.; Aleff, R.; Butz, M.; Highsmith, E.W.; et al. Effect of Trinucleotide Repeat Expansion on the Expression of TCF4 mRNA in Fuchs’ Endothelial Corneal Dystrophy. Investig. Ophthalmol. Vis. Sci. 2019, 60, 779–786. [Google Scholar] [CrossRef]
- Okumura, N.; Hayashi, R.; Koizumi, N. Perspective of Future Potent Therapies for Fuchs Endothelial Corneal Dystrophy. Open Ophthalmol. J. 2018, 12, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Gattey, D.; Zhu, A.Y.; Stagner, A.; Terry, M.A.; Jun, A.S. Fuchs endothelial corneal dystrophy in patients with myotonic dystrophy: A case series. Cornea 2014, 33, 96–98. [Google Scholar] [CrossRef]
- Medeiros, C.S.; Marino, G.K.; Santhiago, M.R.; Wilson, S.E. The Corneal Basement Membranes and Stromal Fibrosis. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4044–4053. [Google Scholar] [CrossRef] [PubMed]
- Ang, M.; Cai, Y.; Shahipasand, S.; Sim, D.A.; Keane, P.A.; Sng, C.C.; Egan, C.A.; Tufail, A.; Wilkins, M.R. En face optical coherence tomography angiography for corneal neovascularisation. Br. J. Ophthalmol. 2016, 100, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Barbariga, M.; Vallone, F.; Mosca, E.; Bignami, F.; Magagnotti, C.; Fonteyne, P.; Chiappori, F.; Milanesi, L.; Rama, P.; Andolfo, A.; et al. The role of extracellular matrix in mouse and human corneal neovascularization. Sci. Rep. 2019, 9, 14272. [Google Scholar] [CrossRef]
- Donthula, G.; Daigavane, S. Secondary Glaucoma Following Corneal Transplantation: A Comprehensive Review of Pathophysiology and Therapeutic Approaches. Cureus 2024, 16, e69882. [Google Scholar] [CrossRef]
- Kitamoto, K.; Taketani, Y.; Fujii, W.; Inamochi, A.; Toyono, T.; Miyai, T.; Yamagami, S.; Kuroda, M.; Usui, T.; Ouchi, Y. Generation of mouse model of TGFBI-R124C corneal dystrophy using CRISPR/Cas9-mediated homology-directed repair. Sci. Rep. 2020, 10, 2000. [Google Scholar] [CrossRef] [PubMed]
- Vemuganti, G.K.; Rathi, V.M.; Murthy, S.I. Histological Landmarks in Corneal Dystrophy: Pathology of Corneal Dystrophies. In Corneal Dystrophies; Seitz, B., Lisch, W., Eds.; S.Karger AG: Basel, Switzerland, 2011; Volume 48, pp. 24–50. [Google Scholar]
- Latifi, G. Corneal Dystrophies. In In Vivo Confocal Microscopy in Eye Disease; Latifi, G., Hau, S., Eds.; Springer: London, UK, 2022; pp. 61–89. [Google Scholar] [CrossRef]
- Finis, D.; Stammen, J.; Lisch, W.; Geerling, G. Epithelial Dystrophies of the Cornea. Klin. Monbl. Augenheilkd. 2019, 236, e23–e36. [Google Scholar] [CrossRef]
- Alemi, H.; Dehghani, S.; Forouzanfar, K.; Surico, P.L.; Narimatsu, A.; Musayeva, A.; Sharifi, S.; Wang, S.; Dohlman, T.H.; Yin, J.; et al. Insights into mustard gas keratopathy-characterizing corneal layer-specific changes in mice exposed to nitrogen mustard. Exp. Eye Res. 2023, 236, 109657. [Google Scholar] [CrossRef]
- Dua, S.H.; Darren, S.J.; Mouhamed Al-Aqaba, T.; Said, D.G. Pathophysiology of Keratoconus. In Keratoconus; Izquierdo, L., Henriquez, M., Mannis, M., Eds.; Elsevier: New Delhi, India, 2023; pp. 51–64. [Google Scholar]
- Zhou, H. Deciphering cell fates of the human cornea with a multi-omics meta atlas. Acta Ophthalmol. 2024, 102. [Google Scholar] [CrossRef]
- Vincent, A.L.; Rootman, D.; Munier, F.L.; Héon, E. A Molecular Perspective on Corneal Dystrophies. In Genetics in Ophthalmology; Wissinger, B., Kohl, S., Langenbeck, U., Eds.; S.Karger AG: Basel, Switzerland, 2003; Volume 37, pp. 50–66. [Google Scholar]
- Lee, Y.F.; Yong, D.W.W.; Manotosh, R. A Review of Contact Lens-Induced Limbal Stem Cell Deficiency. Biology 2023, 12, 1490. [Google Scholar] [CrossRef]
- Matai, H.; Agarwal, S.; Srinivasan, B.; Iyer, G. Simple Limbal Epithelial Transplantation. In Current Advances in Ocular Surgery; Tsui, E., Fung, S.S.M., Singh, R.B., Eds.; Current Practices in Ophthalmology; Springer Nature: Singapore, 2023; pp. 189–199. [Google Scholar]
- Rossen, J.; Amram, A.; Milani, B.; Park, D.; Harthan, J.; Joslin, C.; McMahon, T.; Djalilian, A. Contact Lens-induced Limbal Stem Cell Deficiency. Ocul. Surf. 2016, 14, 419–434. [Google Scholar] [CrossRef]
- Nureen, L.; Di Girolamo, N. Limbal Epithelial Stem Cells in the Diabetic Cornea. Cells 2023, 12, 2458. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Liang, Q.F. Research progress on imagine diagnosis of limbal stem cell deficiency. [Zhonghua yan ke za Zhi] Chin. J. Ophthalmol. 2023, 59, 673–676. [Google Scholar] [CrossRef]
- Rama, P. Limbal Stem Cell Deficiency in Inflammatory Disorders. In Complications in Uveitis; Pichi, F., Neri, P., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 13–21. [Google Scholar]
- Lagali, N. Cellular insights and new therapeutic approaches for limbal stem cell deficiency. Acta Ophthalmol. 2025, 103. [Google Scholar] [CrossRef]
- Lagali, N. The limbal stem cell niche in development, insights from mouse models and children with aniridia. Acta Ophthalmol. 2024, 102. [Google Scholar] [CrossRef]
- Hu, J.C.W.; Trief, D. A narrative review of limbal stem cell deficiency & severe ocular surface disease. Ann. Eye Sci. 2023, 8, 13. [Google Scholar] [CrossRef]
- Record, M.; Silvente-Poirot, S.; Poirot, M.; Wakelam, M.J.O. Extracellular vesicles: Lipids as key components of their biogenesis and functions. J. Lipid Res. 2018, 59, 1316–1324. [Google Scholar] [CrossRef]
- Willms, E.; Cabanas, C.; Mager, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef]
- Goncalves, D.; Pinto, S.N.; Fernandes, F. Extracellular Vesicles and Infection: From Hijacked Machinery to Therapeutic Tools. Pharmaceutics 2023, 15, 1738. [Google Scholar] [CrossRef]
- Stahl, A.L.; Johansson, K.; Mossberg, M.; Kahn, R.; Karpman, D. Exosomes and microvesicles in normal physiology, pathophysiology, and renal diseases. Pediatr. Nephrol. 2019, 34, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Stahl, P.D.; Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles, Integrators of Homeostasis. Physiology 2019, 34, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Roucourt, B.; Meeussen, S.; Bao, J.; Zimmermann, P.; David, G. Heparanase activates the syndecan-syntenin-ALIX exosome pathway. Cell Res. 2015, 25, 412–428. [Google Scholar] [CrossRef]
- Taylor, J.; Azimi, I.; Monteith, G.; Bebawy, M. Ca2+ mediates extracellular vesicle biogenesis through alternate pathways in malignancy. J. Extracell. Vesicles 2020, 9, 1734326. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Tkach, M.; Kowal, J.; Thery, C. Why the need and how to approach the functional diversity of extracellular vesicles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20160479. [Google Scholar] [CrossRef]
- Iraci, N.; Leonardi, T.; Gessler, F.; Vega, B.; Pluchino, S. Focus on Extracellular Vesicles: Physiological Role and Signalling Properties of Extracellular Membrane Vesicles. Int. J. Mol. Sci. 2016, 17, 171. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res. 2019, 47, D516–D519. [Google Scholar] [CrossRef]
- Kalra, H.; Drummen, G.P.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [PubMed]
- Mohankumar, S.; Patel, T. Extracellular vesicle long noncoding RNA as potential biomarkers of liver cancer. Brief. Funct. Genom. 2016, 15, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Poggio, M.; Hu, T.; Pai, C.C.; Chu, B.; Belair, C.D.; Chang, A.; Montabana, E.; Lang, U.E.; Fu, Q.; Fong, L.; et al. Suppression of Exosomal PD-L1 Induces Systemic Anti-tumor Immunity and Memory. Cell 2019, 177, 414–427.e13. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.A.; Seo, B.; Hussaini, H.M.; Hibma, M.; Rich, A.M. Comparing Two Methods for the Isolation of Exosomes. J. Nucleic Acids 2022, 2022, 8648373. [Google Scholar] [CrossRef]
- Habibian, A.; Soleimanjahi, H.; Hashemi, S.M.; Babashah, S. Characterization and Comparison of Mesenchymal Stem Cell-Derived Exosome Isolation Methods using Culture Supernatant. Arch. Razi Inst. 2022, 77, 1383–1388. [Google Scholar] [CrossRef]
- Kurian, T.K.; Banik, S.; Gopal, D.; Chakrabarti, S.; Mazumder, N. Elucidating Methods for Isolation and Quantification of Exosomes: A Review. Mol. Biotechnol. 2021, 63, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Nalabotala, R.; Koo, K.M.; Bose, S.; Nayak, R.; Shiddiky, M.J.A. Separation of distinct exosome subpopulations: Isolation and characterization approaches and their associated challenges. Analyst 2021, 146, 3731–3749. [Google Scholar] [CrossRef]
- Gao, J.; Li, A.; Hu, J.; Feng, L.; Liu, L.; Shen, Z. Recent developments in isolating methods for exosomes. Front. Bioeng. Biotechnol. 2022, 10, 1100892. [Google Scholar] [CrossRef]
- Wu, S.; Zhao, Y.; Zhang, Z.; Zuo, C.; Wu, H.; Liu, Y. The Advances and Applications of Characterization Technique for Exosomes: From Dynamic Light Scattering to Super-Resolution Imaging Technology. Photonics 2024, 11, 101. [Google Scholar] [CrossRef]
- Lobb, R.J.; Becker, M.; Wen Wen, S.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Tiwari, S.; Kumar, V.; Randhawa, S.; Verma, S.K. Preparation and characterization of extracellular vesicles. Am. J. Reprod. Immunol. 2021, 85, e13367. [Google Scholar] [CrossRef]
- Ramirez, M.I.; Amorim, M.G.; Gadelha, C.; Milic, I.; Welsh, J.A.; Freitas, V.M.; Nawaz, M.; Akbar, N.; Couch, Y.; Makin, L.; et al. Technical challenges of working with extracellular vesicles. Nanoscale 2018, 10, 881–906. [Google Scholar] [CrossRef]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Thery, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques used for the isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Pariset, E.; Agache, V.; Millet, A. Extracellular Vesicles: Isolation Methods. Adv. Biosyst. 2017, 1, e1700040. [Google Scholar] [CrossRef] [PubMed]
- Shami-Shah, A.; Travis, B.G.; Walt, D.R. Advances in extracellular vesicle isolation methods: A path towards cell-type specific EV isolation. Extracell. Vesicles Circ. Nucl. Acids 2023, 4, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yoon, J.Y.; Lee, J.H.; Lee, H.H.; Knowles, J.C.; Kim, H.W. Emerging biogenesis technologies of extracellular vesicles for tissue regenerative therapeutics. J. Tissue Eng. 2021, 12, 20417314211019015. [Google Scholar] [CrossRef]
- Dragovic, R.A.; Gardiner, C.; Brooks, A.S.; Tannetta, D.S.; Ferguson, D.J.; Hole, P.; Carr, B.; Redman, C.W.; Harris, A.L.; Dobson, P.J.; et al. Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomedicine 2011, 7, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Lyu, T.S.; Ahn, Y.; Im, Y.J.; Kim, S.S.; Lee, K.H.; Kim, J.; Choi, Y.; Lee, D.; Kang, E.; Jin, G.; et al. The characterization of exosomes from fibrosarcoma cell and the useful usage of Dynamic Light Scattering (DLS) for their evaluation. PLoS ONE 2021, 16, e0231994. [Google Scholar] [CrossRef]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 3, 22. [Google Scholar] [CrossRef]
- Nolte-‘t Hoen, E.; Cremer, T.; Gallo, R.C.; Margolis, L.B. Extracellular vesicles and viruses: Are they close relatives? Proc. Natl. Acad. Sci. USA 2016, 113, 9155–9161. [Google Scholar] [CrossRef]
- Vogel, R.; Coumans, F.A.; Maltesen, R.G.; Boing, A.N.; Bonnington, K.E.; Broekman, M.L.; Broom, M.F.; Buzas, E.I.; Christiansen, G.; Hajji, N.; et al. A standardized method to determine the concentration of extracellular vesicles using tunable resistive pulse sensing. J. Extracell. Vesicles 2016, 5, 31242. [Google Scholar] [CrossRef]
- Sina, A.A.; Vaidyanathan, R.; Wuethrich, A.; Carrascosa, L.G.; Trau, M. Label-free detection of exosomes using a surface plasmon resonance biosensor. Anal. Bioanal. Chem. 2019, 411, 1311–1318. [Google Scholar] [CrossRef]
- Guerrini, L.; Garcia-Rico, E.; O’Loghlen, A.; Giannini, V.; Alvarez-Puebla, R.A. Surface-Enhanced Raman Scattering (SERS) Spectroscopy for Sensing and Characterization of Exosomes in Cancer Diagnosis. Cancers 2021, 13, 2179. [Google Scholar] [CrossRef]
- Yurtsever, A.; Yoshida, T.; Badami Behjat, A.; Araki, Y.; Hanayama, R.; Fukuma, T. Structural and mechanical characteristics of exosomes from osteosarcoma cells explored by 3D-atomic force microscopy. Nanoscale 2021, 13, 6661–6677. [Google Scholar] [CrossRef]
- Daaboul, G.G.; Gagni, P.; Benussi, L.; Bettotti, P.; Ciani, M.; Cretich, M.; Freedman, D.S.; Ghidoni, R.; Ozkumur, A.Y.; Piotto, C.; et al. Digital Detection of Exosomes by Interferometric Imaging. Sci. Rep. 2016, 6, 37246. [Google Scholar] [CrossRef] [PubMed]
- Zieske, J.D.; Hutcheon, A.E.K.; Guo, X. Extracellular Vesicles and Cell-Cell Communication in the Cornea. Anat. Rec. 2020, 303, 1727–1734. [Google Scholar] [CrossRef] [PubMed]
- Ru, Y.; Zhang, Y.; Zhao, S. Expression and function of microRNAs in the cornea. [Zhonghua yan ke za Zhi] Chin. J. Ophthalmol. 2015, 51, 229–235. [Google Scholar]
- Yeung, V.; Zhang, T.C.; Yuan, L.; Parekh, M.; Cortinas, J.; Delavogia, E.; Hutcheon, A.E.K.; Guo, X.; Ciolino, J.B. Extracellular Vesicles Secreted by Corneal Myofibroblasts Promote Corneal Epithelial Cell Migration. Int. J. Mol. Sci. 2022, 23, 3136. [Google Scholar] [CrossRef]
- Saccu, G.; Menchise, V.; Gai, C.; Bertolin, M.; Ferrari, S.; Giordano, C.; Manco, M.; Dastrù, W.; Tolosano, E.; Bussolati, B.; et al. Bone Marrow Mesenchymal Stromal/Stem Cell-Derived Extracellular Vesicles Promote Corneal Wound Repair by Regulating Inflammation and Angiogenesis. Cells 2022, 11, 3892. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Wu, G.; Qi, P.; Zhang, Y.; Liu, Z.; Li, X.; Yu, Y.; Ye, X.; Li, Y.; et al. Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Deliver miR-21 to Promote Corneal Epithelial Wound Healing through PTEN/PI3K/Akt Pathway. Stem Cells Int. 2022, 2022, 1252557. [Google Scholar] [CrossRef]
- Zou, X.Y.; Yu, Y.; Lin, S.; Zhong, L.; Sun, J.; Zhang, G.; Zhu, Y. Comprehensive miRNA Analysis of Human Umbilical Cord-Derived Mesenchymal Stromal Cells and Extracellular Vesicles. Kidney Blood Press. Res. 2018, 43, 152–161. [Google Scholar] [CrossRef]
- Alli, A.A. Mechanisms of Extracellular Vesicle Biogenesis, Cargo Loading, and Release. In Extracellular Vesicles-Role in Diseases, Pathogenesis and Therapy; Paul, M.K., Ed.; Physiology; IntechOpen: London, UK, 2022; p. 338. [Google Scholar] [CrossRef]
- Leszczynska, A.; Kulkarni, M.; Ljubimov, A.V.; Saghizadeh, M. Exosomes from normal and diabetic human corneolimbal keratocytes differentially regulate migration, proliferation and marker expression of limbal epithelial cells. Sci. Rep. 2018, 8, 15173. [Google Scholar] [CrossRef] [PubMed]
- McKay, T.B.; Hutcheon, A.E.K.; Zieske, J.D.; Ciolino, J.B. Extracellular Vesicles Secreted by Corneal Epithelial Cells Promote Myofibroblast Differentiation. Cells 2020, 9, 1080. [Google Scholar] [CrossRef]
- Pucker, A.D.; Ngo, W.; Postnikoff, C.K.; Fortinberry, H.; Nichols, J.J. Tear Film miRNAs and Their Association With Human Dry Eye Disease. Curr. Eye Res. 2022, 47, 1479–1487. [Google Scholar] [CrossRef]
- Lee, S.; Ko, J.H.; Kim, S.N. The Extracellular MicroRNAs on Inflammation: A Literature Review of Rodent Studies. Biomedicines 2022, 10, 1601. [Google Scholar] [CrossRef]
- Yeung, V.; Boychev, N.; Farhat, W.; Ntentakis, D.P.; Hutcheon, A.E.K.; Ross, A.E.; Ciolino, J.B. Extracellular Vesicles in Corneal Fibrosis/Scarring. Int. J. Mol. Sci. 2022, 23, 5921. [Google Scholar] [CrossRef] [PubMed]
- Stunf Pukl, S. Are miRNAs Dynamic Biomarkers in Keratoconus? A Review of the Literature. Genes 2022, 13, 588. [Google Scholar] [CrossRef]
- Dou, S.; Wang, Q.; Zhang, B.; Wei, C.; Wang, H.; Liu, T.; Duan, H.; Jiang, H.; Liu, M.; Qi, X.; et al. Single-cell atlas of keratoconus corneas revealed aberrant transcriptional signatures and implicated mechanical stretch as a trigger for keratoconus pathogenesis. Cell Discov. 2022, 8, 66. [Google Scholar] [CrossRef]
- Wang, Y.M.; Ng, T.K.; Choy, K.W.; Wong, H.K.; Chu, W.K.; Pang, C.P.; Jhanji, V. Histological and microRNA Signatures of Corneal Epithelium in Keratoconus. J. Refract. Surg. 2018, 34, 201–211. [Google Scholar] [CrossRef]
- Ouyang, S.; Ma, J.; Sun, Q.; Li, J.; Chen, Y.; Luo, L. Comprehensive Bioinformatics Analysis to Reveal Key RNA Targets and Hub Competitive Endogenous RNA Network of Keratoconus. Front. Genet. 2022, 13, 896780. [Google Scholar] [CrossRef]
- Akoto, T.; Cai, J.; Nicholas, S.; McCord, H.; Estes, A.J.; Xu, H.; Karamichos, D.; Liu, Y. Unravelling the Impact of Cyclic Mechanical Stretch in Keratoconus—A Transcriptomic Profiling Study. Int. J. Mol. Sci. 2023, 24, 7437. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Asada, K.; Ueno, M.; Hiramoto, N.; Fujita, T.; Toda, M.; Sotozono, C.; Kinoshita, S.; Hamuro, J. Cellular Interplay Through Extracellular Vesicle miR-184 Alleviates Corneal Endothelium Degeneration. Ophthalmol. Sci. 2022, 2, 100212. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Chen, Q.; Wei, Z.; Xu, X.; Han, D.; Zhang, Y.; Chen, Z.; Liang, Q. MicroRNAs of extracellular vesicles derived from mesenchymal stromal cells alleviate inflammation in dry eye disease by targeting the IRAK1/TAB2/NF-kappaB pathway. Ocul. Surf. 2023, 28, 131–140. [Google Scholar] [CrossRef]
- Lozano, V.; Martín, C.; Blanco, N.; Alcalde, I.; Fernandez-Vega Cueto, L.; Merayo-Lloves, J.; Quirós, L.M. Exosomes Released by Corneal Stromal Cells Show Molecular Alterations in Keratoconus Patients and Induce Different Cellular Behavior. Biomedicines 2022, 10, 2348. [Google Scholar] [CrossRef]
- Hadvina, R.; Lotfy Khaled, M.; Akoto, T.; Zhi, W.; Karamichos, D.; Liu, Y. Exosomes and their miRNA/protein profile in keratoconus-derived corneal stromal cells. Exp. Eye Res. 2023, 236, 109642. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, S.; Liu, Y.; Wang, J.; Zhu, R.; Jin, X.; Zhang, H. Effect of Tear Derived Exosomes on Corneal Epithelial Cells During Diabetic Keraropathy. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Mansoor, H.; Ong, H.S.; Riau, A.K.; Stanzel, T.P.; Mehta, J.S.; Yam, G.H. Current Trends and Future Perspective of Mesenchymal Stem Cells and Exosomes in Corneal Diseases. Int. J. Mol. Sci. 2019, 20, 2853. [Google Scholar] [CrossRef]
- Tati, V.; Mitra, S.; Basu, S.; Shukla, S. Bone marrow mesenchymal stem cell-derived extracellular vesicles promote corneal epithelial repair and suppress apoptosis via modulation of Caspase-3 in vitro. FEBS Open Bio 2024, 14, 968–982. [Google Scholar] [CrossRef]
- Casado-Santos, A.; Gonzalez-Cubero, E.; Gonzalez-Fernandez, M.L.; Gonzalez-Rodriguez, Y.; Garcia-Rodriguez, M.B.; Villar-Suarez, V. Equine Corneal Wound Healing Using Mesenchymal Stem Cell Secretome: Case Report. Animals 2024, 14, 1842. [Google Scholar] [CrossRef]
- Kumar, A.; Li, Y.; Mallick, S.; Yang, E.; Dhaliwal, D.K.; Price, A.; Xie, T.; Du, Y. Stem Cell Secretome Promotes Scarless Corneal Wound Healing and Rescues Corneal Sensory Nerves. BioRxiv 2022, 2022, 490347. [Google Scholar] [CrossRef]
- Bhujel, B.; Oh, S.H.; Kim, C.M.; Yoon, Y.J.; Kim, Y.J.; Chung, H.S.; Ye, E.A.; Lee, H.; Kim, J.Y. Mesenchymal Stem Cells and Exosomes: A Novel Therapeutic Approach for Corneal Diseases. Int. J. Mol. Sci. 2023, 24, 10917. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Bojana Simovic, M.; Crissy, F.; Aleksandar, A.; Valentin, D.; Nebojsa, A.; Vladislav, V. Therapeutic Potential of Mesenchymal Stem Cell-Derived Exosomes in the Treatment of Eye Diseases. Cell Biol. Transl. Med. 2018, 1089, 47–57. [Google Scholar]
- Elyan, M.S.; Nada, H.F.; Hamam, G.G.; El Din Bayomi, N.S. Effect of Mesenchymal Stem Cells-Derived Exosomes on Healing of a Rabbit Model of Corneal Alkali Burn. Histological and Immunohistochemical Study. QJM Int. J. Med. 2024, 117, hcae070. [Google Scholar] [CrossRef]
- Ong, H.S.; Riau, A.K.; Yam, G.H.-F.; Yusoff, N.Z.B.M.; Han, E.J.Y.; Goh, T.-W.; Lai, R.C.; Lim, S.K.; Mehta, J.S. Mesenchymal Stem Cell Exosomes as Immunomodulatory Therapy for Corneal Scarring. Int. J. Mol. Sci. 2023, 24, 7456. [Google Scholar] [CrossRef]
- Verma, N.; Khare, D.; Poe, A.J.; Amador, C.; Ghiam, S.; Fealy, A.; Ebrahimi, S.; Shadrokh, O.; Song, X.Y.; Santiskulvong, C.; et al. MicroRNA and Protein Cargos of Human Limbal Epithelial Cell-Derived Exosomes and Their Regulatory Roles in Limbal Stromal Cells of Diabetic and Non-Diabetic Corneas. Cells 2023, 12, 2524. [Google Scholar] [CrossRef]
- Kistenmacher, S.; Schwammle, M.; Martin, G.; Ulrich, E.; Tholen, S.; Schilling, O.; Giessl, A.; Schlotzer-Schrehardt, U.; Bucher, F.; Schlunck, G.; et al. Enrichment, Characterization, and Proteomic Profiling of Small Extracellular Vesicles Derived from Human Limbal Mesenchymal Stromal Cells and Melanocytes. Cells 2024, 13, 623. [Google Scholar] [CrossRef]
- Kobal, N.; Marzidovsek, M.; Schollmayer, P.; Malicev, E.; Hawlina, M.; Marzidovsek, Z.L. Molecular and Cellular Mechanisms of the Therapeutic Effect of Mesenchymal Stem Cells and Extracellular Vesicles in Corneal Regeneration. Int. J. Mol. Sci. 2024, 25, 11121. [Google Scholar] [CrossRef] [PubMed]
- Elhusseiny, A.M.; Soleimani, M.; Eleiwa, T.K.; ElSheikh, R.H.; Frank, C.R.; Naderan, M.; Yazdanpanah, G.; Rosenblatt, M.I.; Djalilian, A.R. Current and Emerging Therapies for Limbal Stem Cell Deficiency. Stem Cells Transl. Med. 2022, 11, 259–268. [Google Scholar] [CrossRef]
- Tavakkoli, F.; Eleiwa, T.K.; Elhusseiny, A.M.; Damala, M.; Rai, A.K.; Cheraqpour, K.; Ansari, M.H.; Doroudian, M.; Keshel, S.H.; Soleimani, M.; et al. Corneal stem cells niche and homeostasis impacts in regenerative medicine; concise review. Eur. J. Ophthalmol. 2023, 33, 1536–1552. [Google Scholar] [CrossRef]
- Altman, J.; Jones, G.; Ahmed, S.; Sharma, S.; Sharma, A. Tear Film MicroRNAs as Potential Biomarkers: A Review. Int. J. Mol. Sci. 2023, 24, 3694. [Google Scholar] [CrossRef]
- Yu, X.; Wu, X. Exosomal let-7b-5p derived from Aspergillus fumigatus-treated human corneal epithelial cells promotes M1 macrophage activation via targeting SOCS-1. Front. Immunol. 2025, 16, 1548802. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Song, W.; Teng, L.; Huang, Y.; Liu, J.; Peng, Y.; Lu, X.; Yuan, J.; Zhao, X.; Zhao, Q.; et al. MiRNA 24-3p-rich exosomes functionalized DEGMA-modified hyaluronic acid hydrogels for corneal epithelial healing. Bioact. Mater. 2023, 25, 640–656. [Google Scholar] [CrossRef]
- Liu, J.; Gao, J.; Lu, P.; Wang, Y.; Xing, S.; Yan, Y.; Han, R.; Hao, P.; Li, X. Mesenchymal Stem Cell-Derived Exosomes as Drug Carriers for Delivering miRNA-29b to Ameliorate Inflammation in Corneal Injury Via Activating Autophagy. Investig. Ophthalmol. Vis. Sci. 2024, 65, 16. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Zheng, Q.; Luo, H.; Li, X.; Chen, Z.; Song, Z.; Zhou, G.; Hong, C. Exosomal miR-19a from adipose-derived stem cells suppresses differentiation of corneal keratocytes into myofibroblasts. Aging 2020, 12, 4093–4110. [Google Scholar] [CrossRef]
- Zhou, T.; He, C.; Lai, P.; Yang, Z.; Liu, Y.; Xu, H.; Lin, X.; Ni, B.; Ju, R.; Yi, W.; et al. miR-204-containing exosomes ameliorate GVHD-associated dry eye disease. Sci. Adv. 2022, 8, eabj9617. [Google Scholar] [CrossRef]
- Wang, G.; Zhu, Y.; Liu, Y.; Yang, M.; Zeng, L. Mesenchymal Stem Cells-Derived Exosomal miR-223-3p Alleviates Ocular Surface Damage and Inflammation by Downregulating Fbxw7 in Dry Eye Models. Investig. Ophthalmol. Vis. Sci. 2024, 65, 1. [Google Scholar] [CrossRef]
- Zeng, B.; Li, Y.; Xia, J.; Xiao, Y.; Khan, N.; Jiang, B.; Liang, Y.; Duan, L. Micro Trojan horses: Engineering extracellular vesicles crossing biological barriers for drug delivery. Bioeng. Transl. Med. 2024, 9, e10623. [Google Scholar] [CrossRef]
- Liu, G.-S.; Chen, H.-A.; Chang, C.-Y.; Chen, Y.-J.; Wu, Y.-Y.; Widhibrata, A.; Yang, Y.-H.; Hsieh, E.-H.; Delila, L.; Lin, I.C.; et al. Innovative platelet-derived extracellular vesicle drug delivery system for the treatment of corneal neovascularization. bioRxiv 2024, 2024, 614855. [Google Scholar] [CrossRef]
- Fan, Z.; Jiang, C.; Wang, Y.; Wang, K.; Marsh, J.; Zhang, D.; Chen, X.; Nie, L. Engineered extracellular vesicles as intelligent nanosystems for next-generation nanomedicine. Nanoscale Horiz. 2022, 7, 682–714. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.N.; Tian, C. Engineered Extracellular Vesicles: Emerging Therapeutic Strategies for Translational Applications. Int. J. Mol. Sci. 2023, 24, 15206. [Google Scholar] [CrossRef]
- Su, Y.; Chen, M.; Xu, W.; Gu, P.; Fan, X. Advances in Extracellular-Vesicles-Based Diagnostic and Therapeutic Approaches for Ocular Diseases. ACS Nano 2024, 18, 22793–22828. [Google Scholar] [CrossRef]
- Durmaz, E.; Dribika, L.; Kutnyanszky, M.; Mead, B. Utilizing extracellular vesicles as a drug delivery system in glaucoma and RGC degeneration. J. Control. Release 2024, 372, 209–220. [Google Scholar] [CrossRef]
- Lybecker, J.R.; Van de Ven, A.; Braesch-Andersen, K.; Juriga, D.; Norein, N.; Hansson, P.; Samanta, A. Comprehensive Characterization and Sustained, on-Demand Delivery of Corneal Epithelial Extracellular Vesicles for Healing Corneal Epithelium. ChemRxiv 2024. [Google Scholar] [CrossRef]
- Puistola, P.; Kethiri, A.; Nurminen, A.; Turkki, J.; Hopia, K.; Miettinen, S.; Moro, A.; Skottman, H. Cornea-Specific Human Adipose Stem Cell-Derived Extracellular Matrix for Corneal Stroma Tissue Engineering. ACS Appl. Mater. Interfaces 2024, 16, 15761–15772. [Google Scholar] [CrossRef]
- Prasathkumar, M.; Dhrisya, C.; Lin, F.H.; Sadhasivam, S. The Design and Developments of Protein-Polysaccharide Biomaterials for Corneal Tissue Engineering. Adv. Mater. Technol. 2023, 8, 2300171. [Google Scholar] [CrossRef]
- Ma, F.; Feng, J.; Liu, X.; Tian, Y.; Wang, W.J.; Luan, F.X.; Wang, Y.J.; Yang, W.Q.; Bai, J.Y.; Zhang, Y.Q.; et al. A synergistic therapeutic nano-eyedrop for dry eye disease based on ascorbic acid-coupled exosomes. Nanoscale 2023, 15, 1890–1899. [Google Scholar] [CrossRef]
- Zhao, W.; He, X.; Liu, R.; Ruan, Q. Accelerating corneal wound healing using exosome-mediated targeting of NF-kappaB c-Rel. Inflamm. Regen. 2023, 43, 6. [Google Scholar] [CrossRef]
- Yu, F.; Zhao, X.; Wang, Q.; Fang, P.H.; Liu, L.; Du, X.; Li, W.; He, D.; Zhang, T.; Bai, Y.; et al. Engineered Mesenchymal Stromal Cell Exosomes-Loaded Microneedles Improve Corneal Healing after Chemical Injury. ACS Nano 2024, 18, 20065–20082. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, Q.; Zhang, J.; Li, C.; Miao, Y.R.; Lei, Q.; Li, Q.; Guo, A.Y. EVmiRNA: A database of miRNA profiling in extracellular vesicles. Nucleic Acids Res. 2019, 47, D89–D93. [Google Scholar] [CrossRef] [PubMed]
- Wróbel-Dudzińska, D.; Kosior-Jarecka, E.; Ćwikińska-Haszcz, A.; Czop, M.; Kocki, J.; Żarnowski, T. miRNA profiling in the aqueous humour in patients with Fuch’s endothelial corneal dystrophy. Acta Ophthalmol. 2024, 102. [Google Scholar] [CrossRef]
- Al Bdour, M.; Sabbagh, H.M.; Jammal, H.M. Multi-modal imaging for the detection of early keratoconus: A narrative review. Eye Vis. 2024, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Torimura, A.; Kanei, S.; Shimizu, Y.; Baba, T.; Uotani, R.; Sasaki, S.I.; Nagase, D.; Inoue, Y.; Ochiya, T.; Miyazaki, D. Profiling miRNAs in tear extracellular vesicles: A pilot study with implications for diagnosis of ocular diseases. Jpn. J. Ophthalmol. 2024, 68, 70–81. [Google Scholar] [CrossRef]
- Al-Sharify, N.T.; Yussof, S.; Ghaeb, N.H.; Al-Sharify, Z.T.; Naser, H.Y.; Ahmed, S.M.; See, O.H.; Weng, L.Y. Advances in Corneal Diagnostics Using Machine Learning. Bioengineering 2024, 11, 1198. [Google Scholar] [CrossRef]
- Cruzat, A.; Qazi, Y.; Hamrah, P. In Vivo Confocal Microscopy of Corneal Nerves in Health and Disease. Ocul. Surf. 2017, 15, 15–47. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as Biomarkers and Diagnostics. J. Cell Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef]
- Cheng, H.L.; Fu, C.Y.; Kuo, W.C.; Chen, Y.W.; Chen, Y.S.; Lee, Y.M.; Li, K.H.; Chen, C.; Ma, H.P.; Huang, P.C.; et al. Detecting miRNA biomarkers from extracellular vesicles for cardiovascular disease with a microfluidic system. Lab Chip 2018, 18, 2917–2925. [Google Scholar] [CrossRef] [PubMed]
- Ramshani, Z.; Zhang, C.; Richards, K.; Chen, L.; Xu, G.; Stiles, B.L.; Hill, R.; Senapati, S.; Go, D.B.; Chang, H.C. Extracellular vesicle microRNA quantification from plasma using an integrated microfluidic device. Commun. Biol. 2019, 2, 189. [Google Scholar] [CrossRef]
- Paolini, A.; Baldassarre, A.; Bruno, S.P.; Felli, C.; Muzi, C.; Ahmadi Badi, S.; Siadat, S.D.; Sarshar, M.; Masotti, A. Improving the Diagnostic Potential of Extracellular miRNAs Coupled to Multiomics Data by Exploiting the Power of Artificial Intelligence. Front. Microbiol. 2022, 13, 888414. [Google Scholar] [CrossRef] [PubMed]
- Muthamilselvan, S.; Ramasami Sundhar Baabu, P.; Palaniappan, A. Microfluidics for Profiling miRNA Biomarker Panels in AI-Assisted Cancer Diagnosis and Prognosis. Technol. Cancer Res. Treat. 2023, 22, 15330338231185284. [Google Scholar] [CrossRef]
- Jeffet, J.; Mondal, S.; Federbush, A.; Tenenboim, N.; Neaman, M.; Deek, J.; Ebenstein, Y.; Bar-Sinai, Y. Machine-Learning-Based Single-Molecule Quantification of Circulating MicroRNA Mixtures. ACS Sens. 2023, 8, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Huang, C.-H.; Hsu, P.-W.; Chen, Y.-C.; Liu, Y.-H.; Wei, Z.-Y.; Chen, C.-H.; Ho, L.-H.; Yao, H.-W.; Lin, T.-Y.; et al. (Invited) liquid Biopsy Tool: Integrated Aiot Digital Bead-Based Sensor for Molecular-Fingerprint Profiling of Extracellular Vesicles. ECS Meet. Abstr. 2023, 243, 1914. [Google Scholar] [CrossRef]
- Al Sulaiman, D.; Juthani, N.; Doyle, P.S. Quantitative and Multiplex Detection of Extracellular Vesicle-Derived MicroRNA via Rolling Circle Amplification within Encoded Hydrogel Microparticles. Adv. Healthc. Mater. 2022, 11, e2102332. [Google Scholar] [CrossRef]
- Zhou, J.; Dong, J.; Hou, H.; Huang, L.; Li, J. High-throughput microfluidic systems accelerated by artificial intelligence for biomedical applications. Lab Chip 2024, 24, 1307–1326. [Google Scholar] [CrossRef]
- Gurnani, B.; Kaur, K.; Lalgudi, V.G.; Kundu, G.; Mimouni, M.; Liu, H.; Jhanji, V.; Prakash, G.; Roy, A.S.; Shetty, R.; et al. Role of artificial intelligence, machine learning and deep learning models in corneal disorders—A narrative review. J. Fr. Ophtalmol. 2024, 47, 104242. [Google Scholar] [CrossRef]
- Nguyen, T.; Ong, J.; Masalkhi, M.; Waisberg, E.; Zaman, N.; Sarker, P.; Aman, S.; Lin, H.; Luo, M.; Ambrosio, R.; et al. Artificial intelligence in corneal diseases: A narrative review. Cont. Lens Anterior Eye 2024, 47, 102284. [Google Scholar] [CrossRef]
- Adampourezare, M.; Hasanzadeh, M.; Seidi, F. Microfluidic assisted recognition of miRNAs towards point-of-care diagnosis: Technical and analytical overview towards biosensing of short stranded single non-coding oligonucleotides. Biomed. Pharmacother. 2022, 153, 113365. [Google Scholar] [CrossRef]
- Qu, J.-h.; Qin, X.-r.; Xie, Z.-j.; Qian, J.-h.; Zhang, Y.; Sun, X.-n.; Sun, Y.-z.; Peng, R.-m.; Xiao, G.-g.; Lin, J.; et al. Establishment of an automatic diagnosis system for corneal endothelium diseases using artificial intelligence. J. Big Data 2024, 11, 67. [Google Scholar] [CrossRef]
- Kryszan, K.; Wylegala, A.; Kijonka, M.; Potrawa, P.; Walasz, M.; Wylegala, E.; Orzechowska-Wylegala, B. Artificial-Intelligence-Enhanced Analysis of In Vivo Confocal Microscopy in Corneal Diseases: A Review. Diagnostics 2024, 14, 694. [Google Scholar] [CrossRef]
- Kosaka, N.; Yoshioka, Y.; Hagiwara, K.; Tominaga, N.; Katsuda, T.; Ochiya, T. Trash or Treasure: Extracellular microRNAs and cell-to-cell communication. Front. Genet. 2013, 4, 173. [Google Scholar] [CrossRef]
- Caporali, A.; Miscianinov, V.; Saif, J.; Emanueli, C. MicroRNA transport in cardiovascular complication of diabetes. Biochim. Biophys. Acta 2016, 1861, 2111–2120. [Google Scholar] [CrossRef]
- Soccio, P.; Moriondo, G.; Lacedonia, D.; Tondo, P.; Quarato, C.M.I.; Foschino Barbaro, M.P.; Scioscia, G. EVs-miRNA: The New Molecular Markers for Chronic Respiratory Diseases. Life 2022, 12, 1544. [Google Scholar] [CrossRef]
- Rautavaara, Y. MSC-EV mechanisms in corneal wound healing and inflammation. Acta Ophthalmol. 2025, 103. [Google Scholar] [CrossRef]
- Deng, S.X.; Dos Santos, A.; Gee, S. Therapeutic Potential of Extracellular Vesicles for the Treatment of Corneal Injuries and Scars. Transl. Vis. Sci. Technol. 2020, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Ratuszny, D.; Gras, C.; Bajor, A.; Borger, A.K.; Pielen, A.; Borgel, M.; Framme, C.; Blasczyk, R.; Figueiredo, C. miR-145 Is a Promising Therapeutic Target to Prevent Cornea Scarring. Hum. Gene Ther. 2015, 26, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Palakurthi, S.S.; Shah, B.; Kapre, S.; Charbe, N.; Immanuel, S.; Pasham, S.; Thalla, M.; Jain, A.; Palakurthi, S. A comprehensive review of challenges and advances in exosome-based drug delivery systems. Nanoscale Adv. 2024, 6, 5803–5826. [Google Scholar] [CrossRef]
- Zeng, H.; Guo, S.; Ren, X.; Wu, Z.; Liu, S.; Yao, X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells 2023, 12, 1416. [Google Scholar] [CrossRef]
- Dave, K.M.; Pinky, P.P.; S Manickam, D. Molecular engineering of extracellular vesicles for drug delivery: Strategies, challenges, and perspectives. J. Control. Release 2025, 386, 114068. [Google Scholar] [CrossRef] [PubMed]
- Dowaidar, M. Gene therapy using extracellular vesicles loaded with miRNA derived from Bone Marrow Mesenchymal Stem Cells is a cell-free medication delivery method used in a variety of diseases. J. Nanobiotechnol. 2021, 19, 194. [Google Scholar]
- Hernandez, M.J.; Gaetani, R.; Pieters, V.M.; Ng, N.W.; Chang, A.E.; Martin, T.R.; Ingen, E.v.; Mol, E.A.; Dzieciatkowska, M.; Hansen, K.C.; et al. Decellularized Extracellular Matrix Hydrogels as a Delivery Platform for MicroRNA and Extracellular Vesicle Therapeutics. Adv. Ther. 2018, 1, 1800032. [Google Scholar] [CrossRef]
- Li, T.; Zhang, W.M.; Wang, J.; Liu, B.J.; Gao, Q.; Zhang, J.; Qian, H.D.; Pan, J.Y.; Liu, M.; Huang, Q.; et al. Circulating Small Extracellular Vesicles Involved in Systemic Regulation Respond to RGC Degeneration in Glaucoma. Adv. Sci. 2024, 11, e2309307. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Reclusa Asiain, P.; Durendez Saez, E.; Jantus-Lewintre, E.; Malarani, M.; Khan, S.; Fontana, S.; Naing, A.; Passiglia, F.; Raez, L.E.; et al. Extracellular Vesicles As miRNA Nano-Shuttles: Dual Role in Tumor Progression. Target. Oncol. 2018, 13, 175–187. [Google Scholar] [CrossRef]
- Groot, M.; Lee, H. Sorting Mechanisms for MicroRNAs into Extracellular Vesicles and Their Associated Diseases. Cells 2020, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Kumar, U.S.; Sadeghipour, N.; Massoud, T.F.; Paulmurugan, R. A Microfluidics-Based Scalable Approach to Generate Extracellular Vesicles with Enhanced Therapeutic MicroRNA Loading for Intranasal Delivery to Mouse Glioblastomas. ACS Nano 2021, 15, 18327–18346. [Google Scholar] [CrossRef]
- Verma, N.; Arora, S. Navigating the Global Regulatory Landscape for Exosome-Based Therapeutics: Challenges, Strategies, and Future Directions. Pharmaceutics 2025, 17, 990. [Google Scholar] [CrossRef]
- Ting, D.S.J.; Peh, G.S.L.; Adnan, K.B.; Mehta, J.S. Translational and Regulatory Challenges of Corneal Endothelial Cell Therapy: A Global Perspective. Tissue Eng. Part B Rev. 2020, 28, 52–62. [Google Scholar]
- Yadav, A.; Xuan, Y.; Sen, C.K.; Ghatak, S. Standardized Reporting of Research on Exosomes to Ensure Rigor and Reproducibility. Adv. Wound Care 2024, 13, 584–599. [Google Scholar] [CrossRef]
- Nelson, B.C.; Samantha, M.; Ghiran, G.I.; Jones, C.J.; DeRose, P.C.; Elzafir, E.; Vreeland, W.N.; Wang, L. Measurement and Standardization Challenges for Extracellular Vesicle Therapeutic Delivery Vectors. Nanomedicine 2020, 15, 2149–2170. [Google Scholar] [CrossRef]
- Pua, H.H.; Happ, H.C.; Gray, C.J.; Mar, D.J.; Chiou, N.T.; Hesse, L.E.; Ansel, K.M. Increased Hematopoietic Extracellular RNAs and Vesicles in the Lung during Allergic Airway Responses. Cell Rep. 2019, 26, 933–944.e4. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Ludwig, R.G.; Garcia-Martin, R.; Brandao, B.B.; Kahn, C.R. Extracellular miRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef]
- Cheng, L.; Vella, L.J.; Barnham, K.J.; McLean, C.; Masters, C.L.; Hill, A.F. Small RNA fingerprinting of Alzheimer’s disease frontal cortex extracellular vesicles and their comparison with peripheral extracellular vesicles. J. Extracell. Vesicles 2020, 9, 1766822. [Google Scholar] [CrossRef]
- Reis, M.; Mavin, E.; Nicholson, L.; Green, K.; Dickinson, A.M.; Wang, X.N. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Attenuate Dendritic Cell Maturation and Function. Front. Immunol. 2018, 9, 2538. [Google Scholar] [CrossRef]
- Min, L.; Zhu, S.; Chen, L.; Liu, X.; Wei, R.; Zhao, L.; Yang, Y.; Zhang, Z.; Kong, G.; Li, P.; et al. Evaluation of circulating small extracellular vesicles derived miRNAs as biomarkers of early colon cancer: A comparison with plasma total miRNAs. J. Extracell. Vesicles 2019, 8, 1643670. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Di, K.; Fan, B.; Wu, J.; Gu, X.; Sun, Y.; Khan, A.; Li, P.; Li, Z. MicroRNAs in extracellular vesicles: Sorting mechanisms, diagnostic value, isolation, and detection technology. Front. Bioeng. Biotechnol. 2022, 10, 948959. [Google Scholar] [CrossRef] [PubMed]
- Lo, T.-W.; Zhang, Y.; Liu, J.; Kim, Y.; Dong, H.; Lucien, F.; Liu, Y. Abstract A030: Rapid extraction and detection of extracellular vesicle-derived PD-L1 in a microfluidic platform. Clin. Cancer Res. 2024, 30, A30. [Google Scholar]
- Bracht, J.W.P.; Los, M.; van der Pol, E.; Verkuijlen, S.; van Eijndhoven, M.A.J.; Pegtel, D.M.; Nieuwland, R. Choice of size-exclusion chromatography column affects recovery, purity, and miRNA cargo analysis of extracellular vesicles from human plasma. Extracell. Vesicles Circ. Nucleic Acids 2024, 5, 497–508. [Google Scholar] [CrossRef]
- Miceli, R.T.; Chen, T.Y.; Nose, Y.; Tichkule, S.; Brown, B.; Fullard, J.F.; Saulsbury, M.D.; Heyliger, S.O.; Gnjatic, S.; Kyprianou, N.; et al. Extracellular vesicles, RNA sequencing, and bioinformatic analyses: Challenges, solutions, and recommendations. J. Extracell. Vesicles 2024, 13, e70005. [Google Scholar] [CrossRef]
- Srinivasan, S.; Duval, M.X.; Kaimal, V.; Cuff, C.; Clarke, S.H. Assessment of methods for serum extracellular vesicle small RNA sequencing to support biomarker development. J. Extracell. Vesicles 2019, 8, 1684425. [Google Scholar] [CrossRef] [PubMed]
- Nieuwland, R.; Falcon-Perez, J.M.; Thery, C.; Witwer, K.W. Rigor and standardization of extracellular vesicle research: Paving the road towards robustness. J. Extracell. Vesicles 2020, 10, e12037. [Google Scholar] [CrossRef] [PubMed]
- Saint-Pol, J.; Culot, M. Minimum information for studies of extracellular vesicles (MISEV) as toolbox for rigorous, reproducible and homogeneous studies on extracellular vesicles. Toxicol. In Vitro 2025, 106, 106049. [Google Scholar] [CrossRef]
- Li, D.; Yao, X.; Yue, J.; Fang, Y.; Cao, G.; Midgley, A.C.; Nishinari, K.; Yang, Y. Advances in Bioactivity of MicroRNAs of Plant-Derived Exosome-Like Nanoparticles and Milk-Derived Extracellular Vesicles. J. Agric. Food Chem. 2022, 70, 6285–6299. [Google Scholar] [CrossRef]
- Zhong, H.; Guan, G.; Jin, Y. Roles of helminth extracellular vesicle-derived let-7 in host-parasite crosstalk. Front. Immunol. 2024, 15, 1449495. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Rao, J.; Liang, Z.; Xu, X.; Lin, F.; Lin, Y.; Wang, C.; Chen, C. Efficacy of miRNA-modified mesenchymal stem cell extracellular vesicles in spinal cord injury: A systematic review of the literature and network meta-analysis. Front. Neurosci. 2022, 16, 989295. [Google Scholar] [CrossRef]
- Takakura, Y.; Hanayama, R.; Akiyoshi, K.; Futaki, S.; Hida, K.; Ichiki, T.; Ishii-Watabe, A.; Kuroda, M.; Maki, K.; Miura, Y.; et al. Quality and Safety Considerations for Therapeutic Products Based on Extracellular Vesicles. Pharm. Res. 2024, 41, 1573–1594. [Google Scholar] [CrossRef]
- Lim, Y.J.; Seo, M.S.; Park, W.T.; Park, S.; Lee, G.W. Extracellular vesicle-derived MicroRNAs as potential therapies for spinal cord and peripheral nerve injuries. RNA Biol. 2025, 22, 1–9. [Google Scholar] [CrossRef]
- DaCunza, J.T.; Wickman, J.R.; Ajit, S.K. miRNA packaging into small extracellular vesicles and implications in pain. Pain Rep. 2024, 9, e1198. [Google Scholar] [CrossRef]
- Leavitt, R.J.; Limoli, C.L.; Baulch, J.E. miRNA-based therapeutic potential of stem cell-derived extracellular vesicles: A safe cell-free treatment to ameliorate radiation-induced brain injury. Int. J. Radiat. Biol. 2019, 95, 427–435. [Google Scholar] [CrossRef]
- Al-Modawi, R.N.; Brinchmann, J.E.; Karlsen, T.A. Immunological Off-Target Effects of microRNA Control Sequences. Res. Sq. 2020. [Google Scholar] [CrossRef]
- O’Brien, K.P.; Khan, S.; Gilligan, K.E.; Zafar, H.; Lalor, P.; Glynn, C.; O’Flatharta, C.; Ingoldsby, H.; Dockery, P.; De Bhulbh, A.; et al. Employing mesenchymal stem cells to support tumor-targeted delivery of extracellular vesicle (EV)-encapsulated microRNA-379. Oncogene 2018, 37, 2137–2149. [Google Scholar] [CrossRef]
- Albanese, M.; Chen, Y.A.; Huls, C.; Gartner, K.; Tagawa, T.; Mejias-Perez, E.; Keppler, O.T.; Gobel, C.; Zeidler, R.; Shein, M.; et al. MicroRNAs are minor constituents of extracellular vesicles that are rarely delivered to target cells. PLoS Genet. 2021, 17, e1009951. [Google Scholar] [CrossRef]
- Jiang, L.; Vader, P.; Schiffelers, R.M. Extracellular vesicles for nucleic acid delivery: Progress and prospects for safe RNA-based gene therapy. Gene Ther. 2017, 24, 157–166. [Google Scholar] [CrossRef]
- Drees, E.E.E.; Groenewegen, N.J.; Verkuijlen, S.; van Eijndhoven, M.A.J.; Ramaker, J.; Veenstra, P.; Hussain, M.; Groothuis-Oudshoorn, C.G.M.; de Jong, D.; Zijlstra, J.M.; et al. Towards IVDR-compliance by implementing quality control steps in a quantitative extracellular vesicle-miRNA liquid biopsy assay for response monitoring in patients with classic Hodgkin lymphoma. J. Extracell. Biol. 2024, 3, e164. [Google Scholar] [CrossRef] [PubMed]
- Abeysinghe, P.; Turner, N.; Mitchell, M.D. A comparative analysis of small extracellular vesicle (sEV) micro-RNA (miRNA) isolation and sequencing procedures in blood plasma samples. Extracell. Vesicles Circ. Nucl. Acids 2024, 5, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Gouin, K.; Peck, K.; Antes, T.; Johnson, J.L.; Li, C.; Vaturi, S.D.; Middleton, R.; de Couto, G.; Walravens, A.S.; Rodriguez-Borlado, L.; et al. A comprehensive method for identification of suitable reference genes in extracellular vesicles. J. Extracell. Vesicles 2017, 6, 1347019. [Google Scholar] [CrossRef]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Vigano, M.; Colombini, A.; Lugano, G.; Bollati, V.; de Girolamo, L. miR-22-5p and miR-29a-5p Are Reliable Reference Genes for Analyzing Extracellular Vesicle-Associated miRNAs in Adipose-Derived Mesenchymal Stem Cells and Are Stable under Inflammatory Priming Mimicking Osteoarthritis Condition. Stem Cell Rev. Rep. 2019, 15, 743–754. [Google Scholar] [CrossRef]
- Buschmann, D.; Kirchner, B.; Hermann, S.; Marte, M.; Wurmser, C.; Brandes, F.; Kotschote, S.; Bonin, M.; Steinlein, O.K.; Pfaffl, M.W.; et al. Evaluation of serum extracellular vesicle isolation methods for profiling miRNAs by next-generation sequencing. J. Extracell. Vesicles 2018, 7, 1481321. [Google Scholar] [CrossRef] [PubMed]
- Maqueda, J.J.; Giovanazzi, A.; Rocha, A.M.; Rocha, S.; Silva, I.; Saraiva, N.; Bonito, N.; Carvalho, J.; Maia, L.; Wauben, M.H.M.; et al. Adapter dimer contamination in sRNA-sequencing datasets predicts sequencing failure and batch effects and hampers extracellular vesicle-sRNA analysis. J. Extracell. Biol. 2023, 2, e91. [Google Scholar] [CrossRef] [PubMed]
- Shukuya, T.; Ghai, V.; Amann, J.M.; Okimoto, T.; Shilo, K.; Kim, T.K.; Wang, K.; Carbone, D.P. Circulating MicroRNAs and Extracellular Vesicle-Containing MicroRNAs as Response Biomarkers of Anti-programmed Cell Death Protein 1 or Programmed Death-Ligand 1 Therapy in NSCLC. J. Thorac. Oncol. 2020, 15, 1773–1781. [Google Scholar] [CrossRef]
- Chen, X.; Jin, Y.; Feng, Y. Evaluation of Plasma Extracellular Vesicle MicroRNA Signatures for Lung Adenocarcinoma and Granuloma With Monte-Carlo Feature Selection Method. Front. Genet. 2019, 10, 367. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, L.; Cheng, L.; Wang, Y.; Li, M.; Yu, J.; Ma, Z.; Ho, P.C.; Sethi, G.; Chen, X.; et al. Extracellular vesicle-derived biomarkers in prostate cancer care: Opportunities and challenges. Cancer Lett. 2024, 601, 217184. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).