
The Rostral Epidural Rete Mirabile: Functional Significance in Blood Flow Regulatory Mechanisms in Giraffe (Giraffa camelopardalis)

 ,
,  and
and 
Abstract
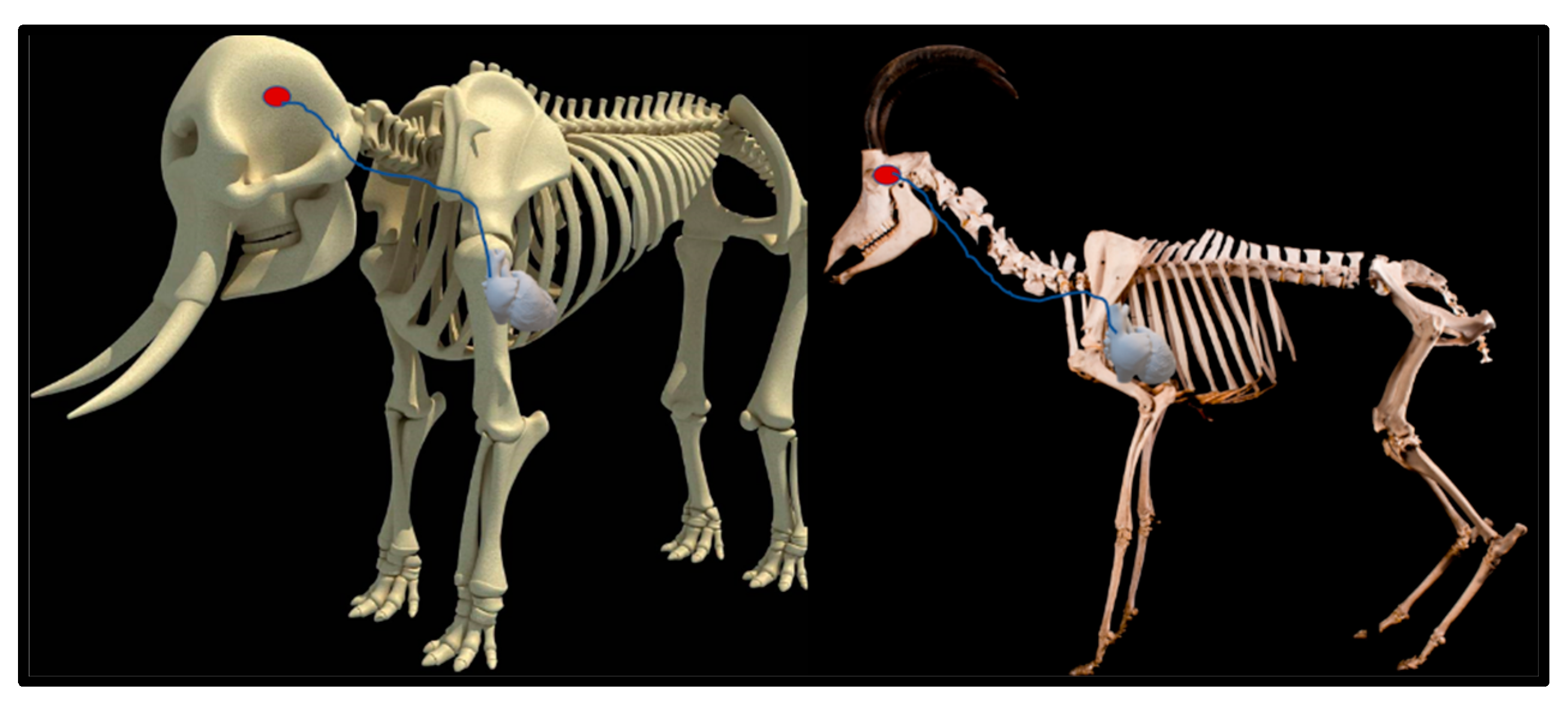
1. Introduction
2. Materials and Methods
2.1. Experimental Model and Subject Details
2.2. Animal Sources
2.3. Method Details
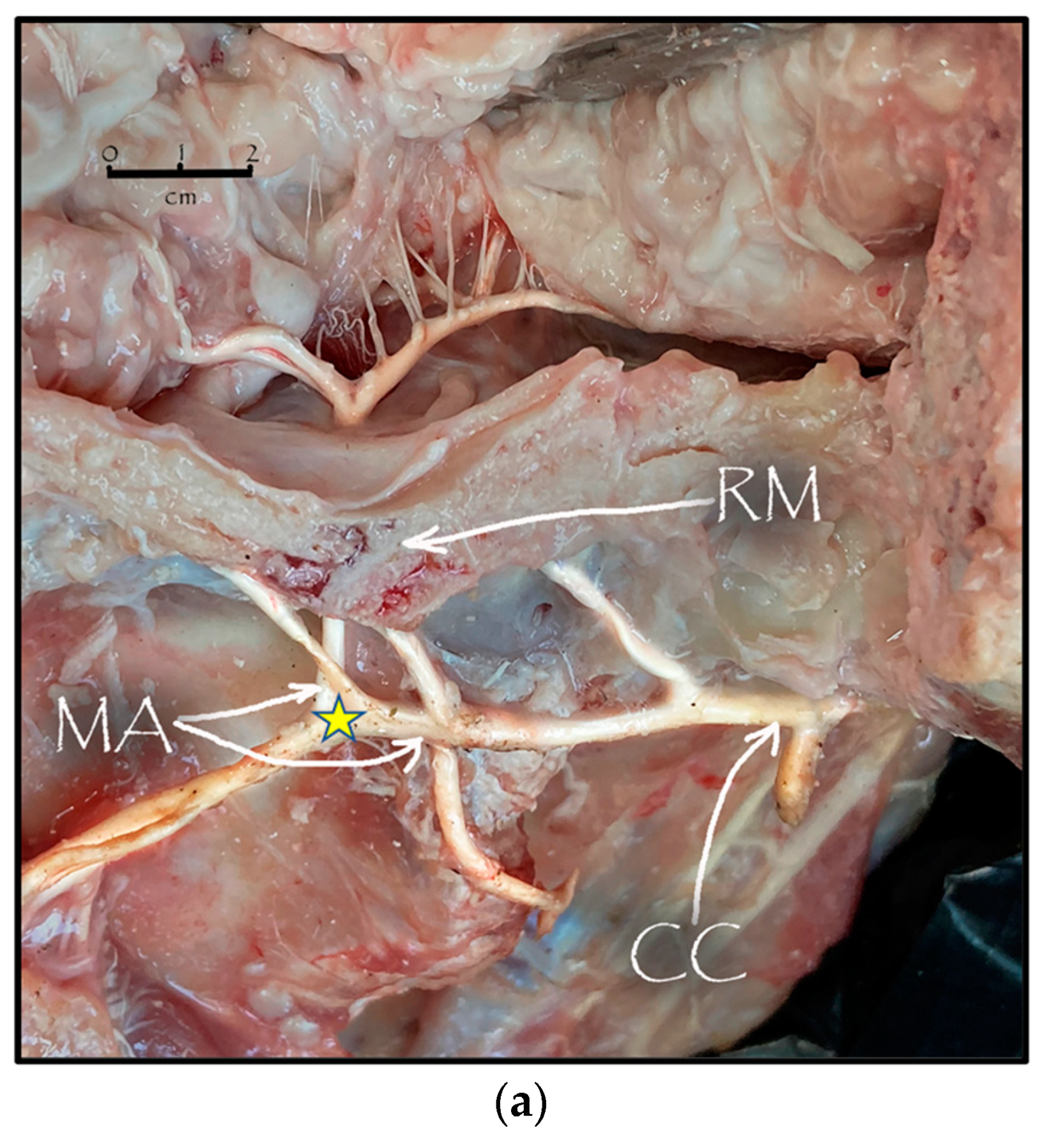
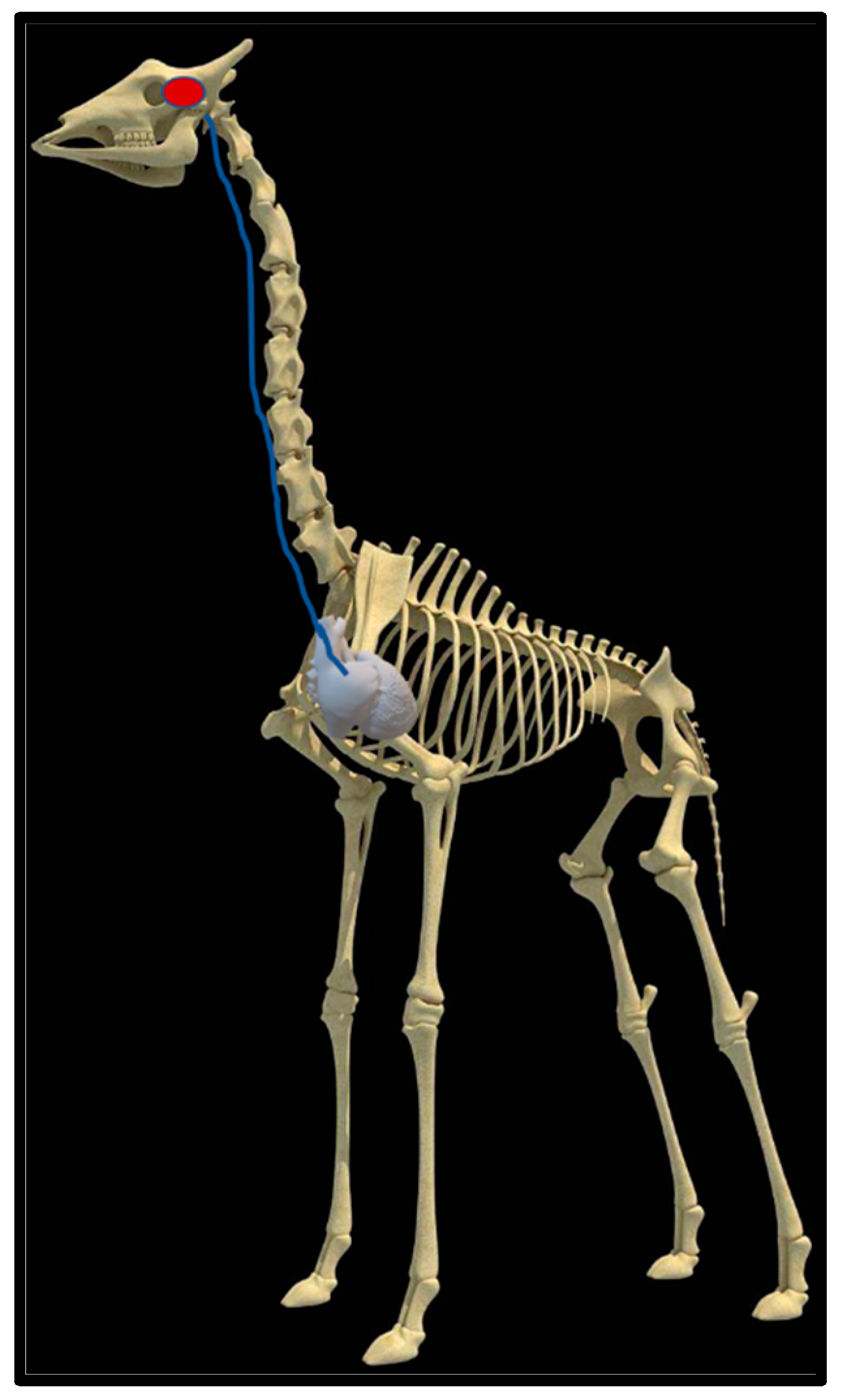
2.4. Dissection
2.5. Brain Endocast
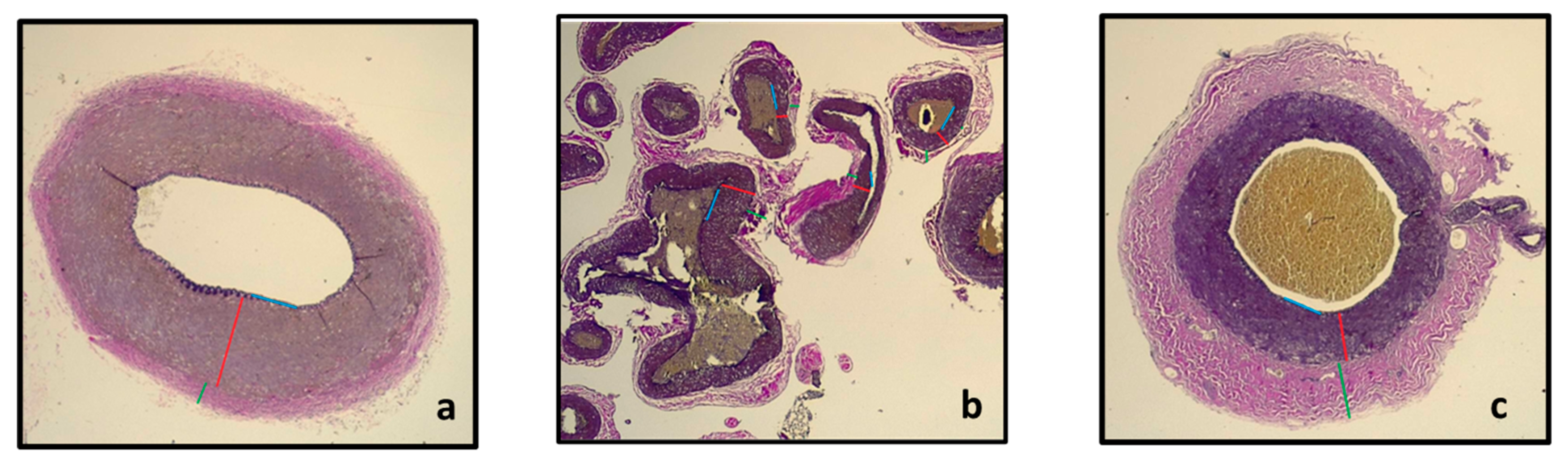
2.6. Histology Sample
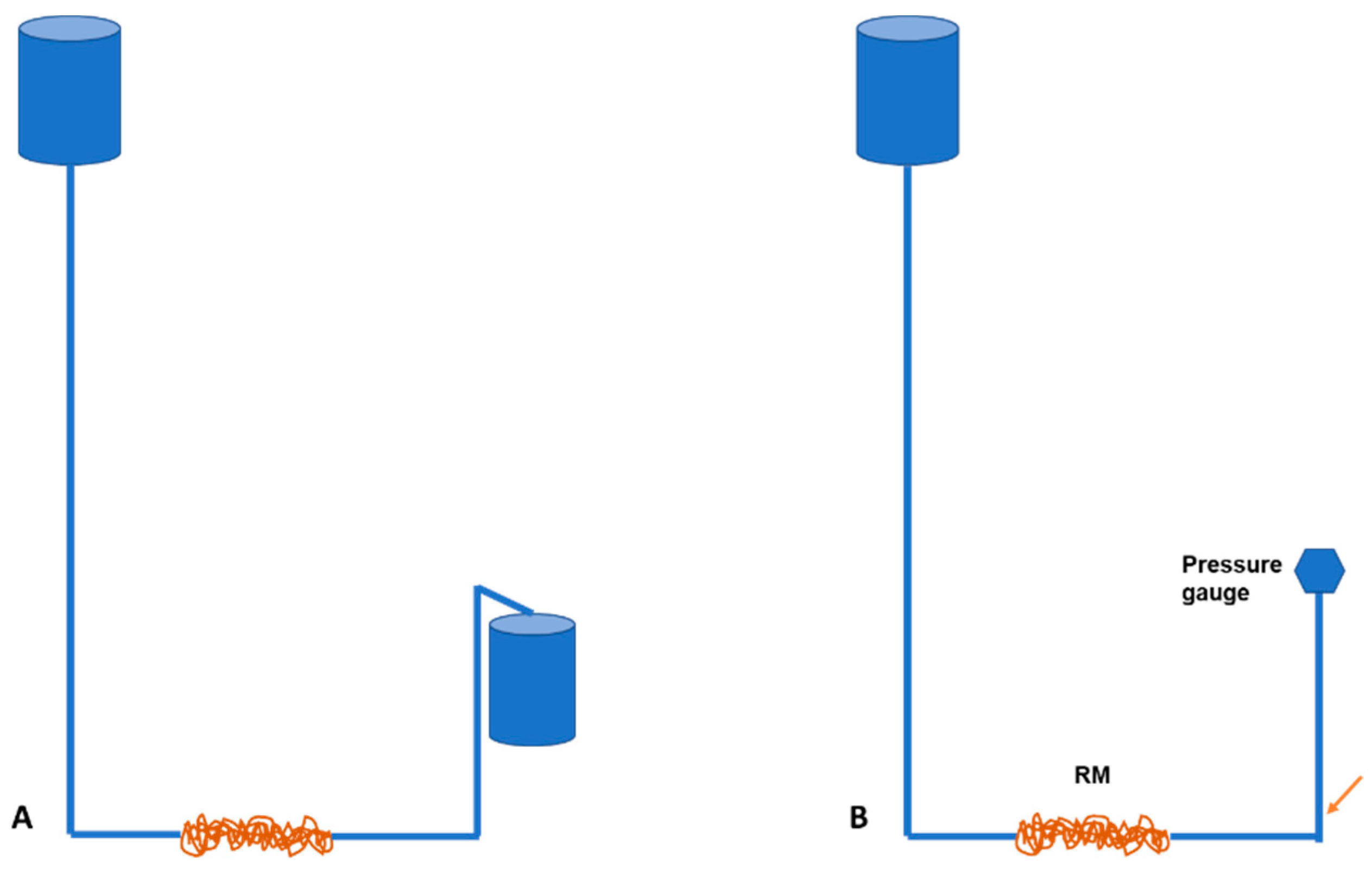
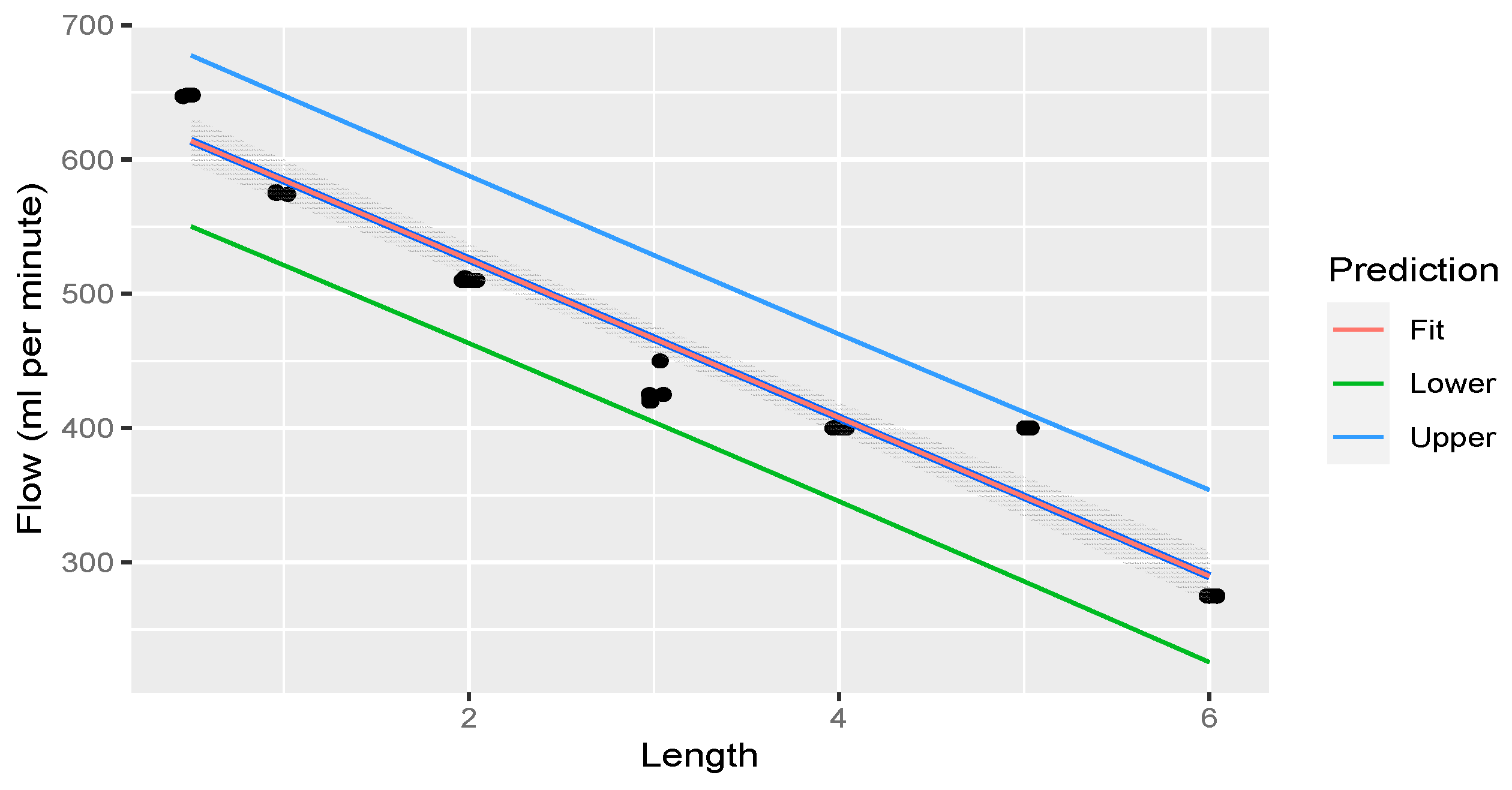
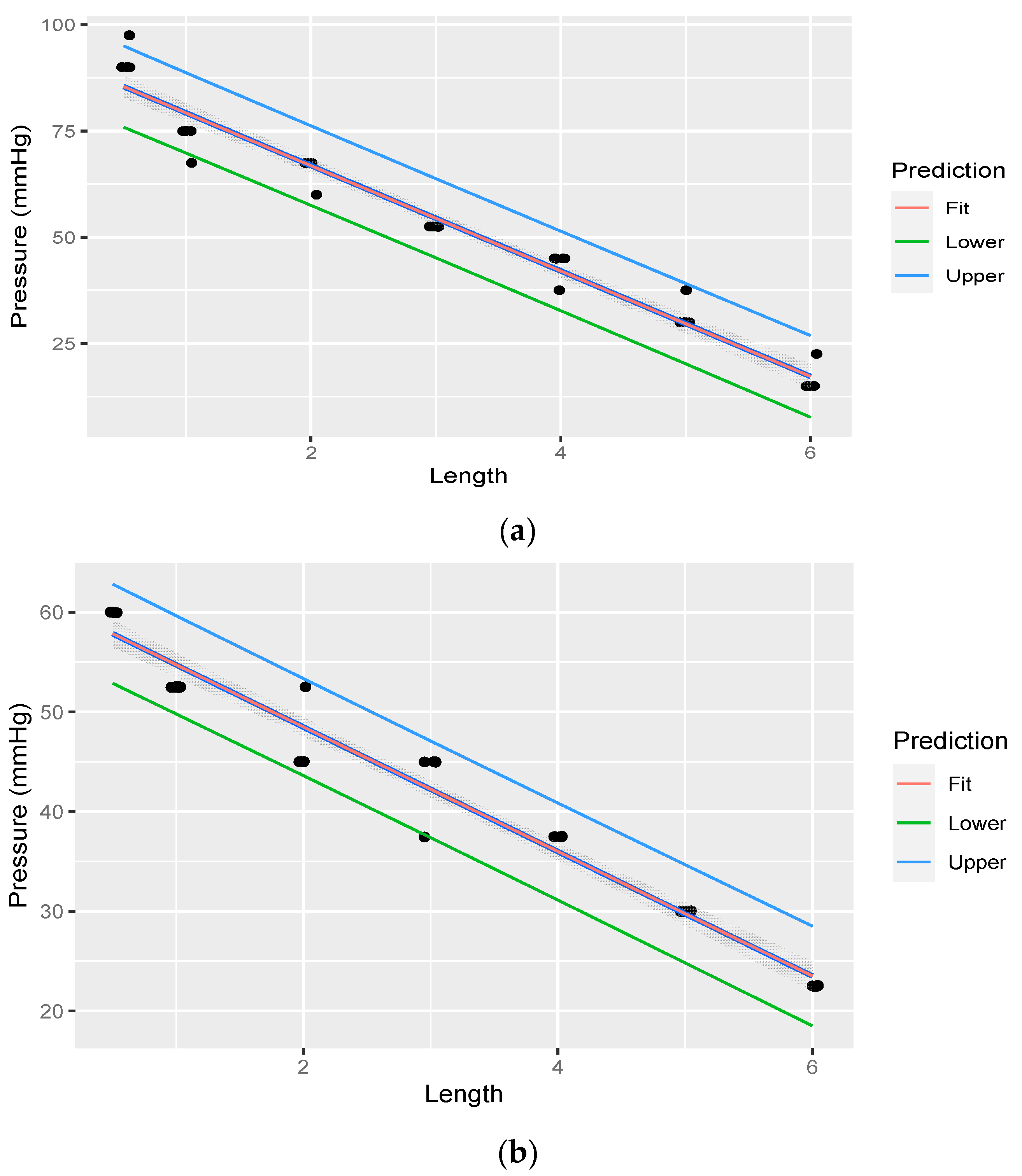
2.7. Pressure and Flow Tempo Experiment
3. Results
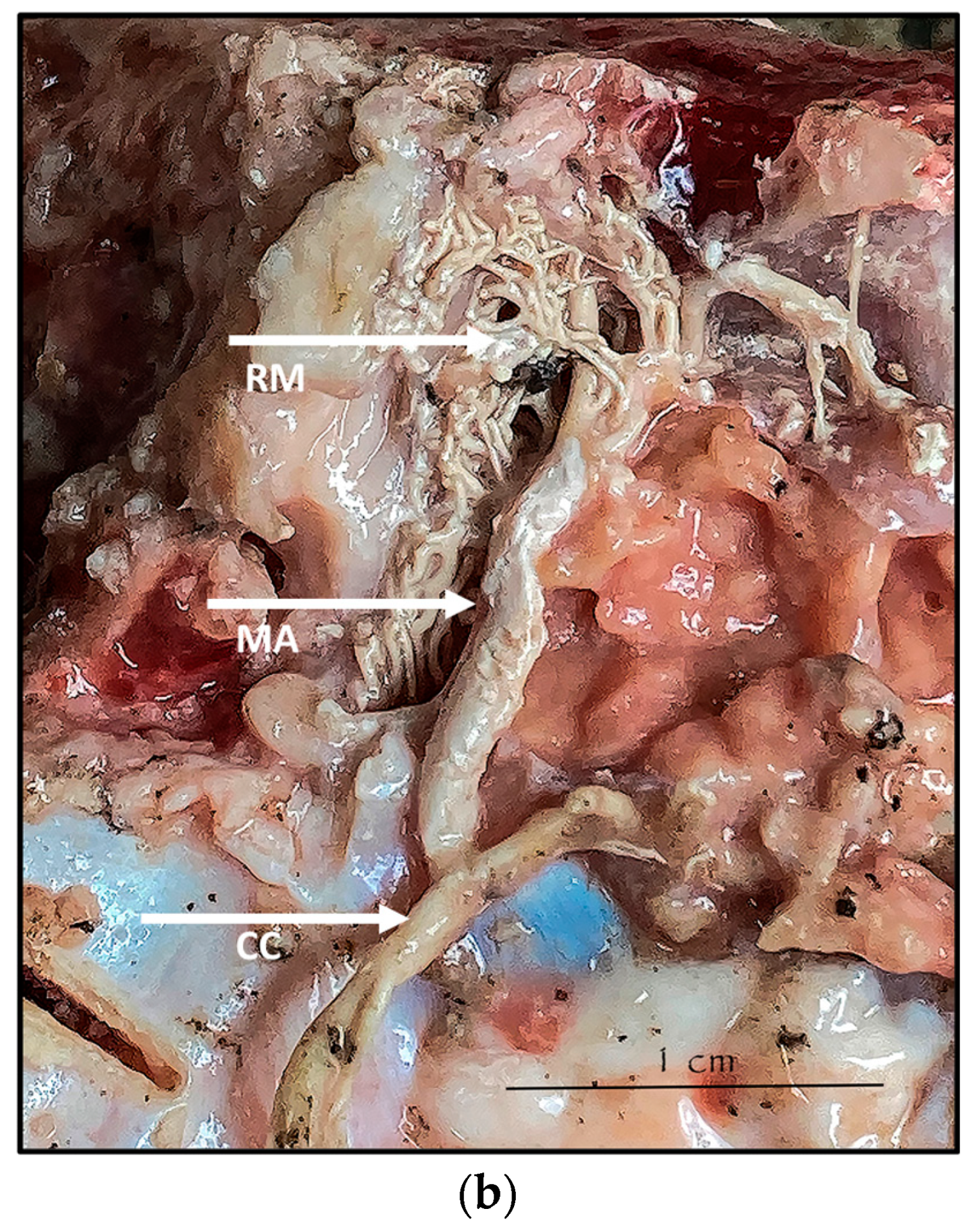
3.1. Dissection
3.2. Brain Volume Endocast
3.3. Histology
3.4. Pressure and Flow Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hargens, A.R.; Gershuni, D.H.; Danzig, L.A.; Millard, R.W.; Pettersson, K. Tissue Adaptations to gravitational stress: Newborn versus Adult Giraffes. Physiologist 1988, 31, S110–S113. [Google Scholar]
- Ostergaard, K.H.; Bertelsen, M.F.; Brondum, E.T.; Alkjaer, C.; Hasenkam, J.M.; Smerup, M.; Wang, T.; Nyengaard, J.R.; Baandrup, U. Pressure profile and morphology of the arteries along the giraffe limb. J. Comp. Physiol. B 2011, 181, 691–698. [Google Scholar] [CrossRef]
- Peterson, K.K.; Horlyck, A.; Ostergaard, K.H.; Andresen, J.; Skovgaard, N.; Telinius, N.; Laher, I.; Bertelsel, M.F.; Grondahl, C.; Smerup, M.; et al. Protection against high intravascular pressure in giraffe legs. Am. J. Physiol. Regul. Integr. 2013, 305, R1021–R1030. [Google Scholar] [CrossRef] [PubMed]
- Aalkjaer, C.; Wang, T. The Remarkable Cardiovascular System of Giraffes. Annu. Rev. Physiol. 2021, 83, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.; Skinner, J.D. How Giraffe adapt to their extraordinary shape. Trans. R. Soc. S. Afr. 1993, 48, 207–2108. [Google Scholar] [CrossRef]
- Fukuta, L.; Kudo, H.; Sasaki, M.; Kiura, J.; Ismail, D.; Endo, H. Absence of rostral epidural rete mirabile in small tropical ruminants: Implication for the evolution of the arterial system in artiodactyls. Anatomy 2007, 210, 112–116. [Google Scholar] [CrossRef]
- Kieltyka-Kurc, A.; Frackowiak, H.; Zdun, M.; Nabzdyk, M.; Kowalczyk, K.; Tokacz, M. The arteries on the base of the brain in the camelids (Camelidae). Ital. J. Zool. 2014, 81, 215–220. [Google Scholar]
- O’Brien, H.D.; Gignac, P.M.; Hieronymus, T.L.; Witmer, L.M. A comparison of postnatal patterns in a growth series of giraffe (Artiodactyla: Giraffa Camelopardalis). Dev. Biol. Evol. Stud. Zool. 2016, 4, e1696. [Google Scholar]
- O’Brien, H.D. Cranial arterial patterns of the alpaca (Camelidae: Vicugna pacos). R. Soc. Open Sci. 2017, 4, 160967. [Google Scholar] [CrossRef]
- Daniel, P.M.; Dawes, J.D.K.; Prichard, M.M.L. Studies of the rostral epidural rete mirabile and its associated arteries. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1953, 237, 173–208. [Google Scholar]
- Dieguez, G.; Garcia-Villalon, A.L.; Gomez, B.; Lluch, L. Hemodynamic significance of the rostral epidural rete mirabile during changes in arterial blood pressure. Am. Physiol. Soc. 1988, 254, R770–R775. [Google Scholar]
- Melnikov, V.V. The Arterial System of the Sperm Whale (Physeter macrocephalus). J. Morphol. 1997, 234, 37–50. [Google Scholar] [CrossRef]
- Rommel, S.A.; Costidis, A.M.; Fernandez, A.; Jepson, P.D.; Pabst, D.A.; McLellan, W.A.; Houser, D.S.; Cranford, T.W.; Van Helden, A.L.; Allen, D.M.; et al. Elements of beaked whale anatomy and diving physiology and some hypothetical causes of sonar-related stranding. J. Cetacean Res. Manag. 2006, 7, 189–209. [Google Scholar] [CrossRef]
- Wang, X.R.; Liu, Y.; Guo, T.F.; Wu, J.P. Anatomic peculiarities of cerebral arterial system and blood supply in Yak (Bos grunniens). J. Anim. Vet. Adv. 2012, 11, 2533–2539. [Google Scholar]
- O’Brien, H.D.; Bourke, J. Physical and computational fluid dynamics models for the hemodynamics of the artiodactyl carotid rete. J. Theor. Biol. 2015, 386, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Frackowiak, H.; Godynicki, S. Brain basal arteries in various species of Felidae. Pol. J. Vet. Sci. 2003, 6, 195–200. [Google Scholar] [PubMed]
- Jessen, C. Selective brain cooling in mammals and birds. Jpn. J. Physiol. 2001, 51, 291–301. [Google Scholar] [CrossRef]
- Mitchell, G.; Lust, A. The carotid rete and artiodactyl success. Biol. Lett. 2008, 4, 415–418. [Google Scholar] [CrossRef]
- Frackowiak, H.; Jakubowski, H. Arterial vascularization in the giraffe brain. Ann. Zool. Fenn. 2008, 45, 353–359. [Google Scholar] [CrossRef]
- Aiyan, A.A.; Menan, P.; AlDarwich, A.; Almuhairi, F.; Alnuaimi, S.; Bulshawareb, A.; Qablan, M.; Shebab, S. Descriptive Analysis of Cerebral Arterial Vascular Architecture in Dromedary Camel (Camelus dromedarius). Front. Neuroanat. 2019, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jerbi, H.; Vazquez, N.; Perez, W. Morphological Configuration and Topography of the brain arterial supply of the One-humped Camel (Camelus dromedarius, Linnaeus 1758). Int. J. Morphol. 2019, 37, 1095–1100. [Google Scholar] [CrossRef]
- Oliveira, J.C.D.; Campos, R. Systematic study of brain-based arteries in the wild boar (Sus scrofa scrofa). Anat. Histol. Embriol. 2005, 34, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Khamas, W.A.; Goshal, N.G.; Bal, H.S. Histomorphologic structure of the carotid rete-cavernous sinus complex and its functional importance in sheep (Ovis aries). Am. J. Vet. Res. 1983, 45, 156–158. [Google Scholar]
- Edelman, N.H.; Epstein, P.; Cherniak, N.S.; Fishman, A.P. Control of cerebral blood flow in the goat; role of the carotid rete. Am. J. Physiol. 1972, 223, 615–619. [Google Scholar] [CrossRef][Green Version]
- O’Brien, H.D. From Anomalous arteries to Selective Brain Cooling: Parallel Evolution of the Artiodactyl Carotid Rete. Anat. Rec. 2020, 303, 208–317. [Google Scholar] [CrossRef]
- Deacon, F.; Tutchings, A. The South African giraffe Giraffa camelopardalis giraffa: A conservation success story. Oryx 2019, 53, 45–48. [Google Scholar] [CrossRef]
- Kock, M.D.; Burroughs, R. Chemical and Physical Restraint of Wild Animals: A Training and Field Manual for African Species, 2nd ed.; IWVS (Africa): Greyton, South Africa, 2012. [Google Scholar]
- Lindstedt, S.L.; Schaeffer, P.J. Use of allometry in predicting anatomical and physiological parameters of mammals. Lab. Anim. 2002, 26, 1–19. [Google Scholar] [CrossRef]
- Sharma, R.; Sharma, S. Physiology, Blood Volume. In StatPearls, Treasure Island (F.L.); StatPearls: Tampa, FL, USA, 2021; Available online: https://europepmc.org/article/nbk/nbk526077#free-full-text (accessed on 16 October 2022).
- Parent, A.; Carpenter, M.B. Carpenters Human Neuroanatomy, 9th ed.; Williams and Wilkins Publisher: Philadelphia, PA, USA, 1995. [Google Scholar]
- Jerbi, H.; Khaldi, S.; Perez, W. Morphometric study of the rostral epidural rete mirabile in the dromedary (Camelus dromedarius). Int. J. Morphol. 2016, 34, 1429–1435. [Google Scholar] [CrossRef]
- Dormer, K.J.; Denn, M.J.; Stone, J.L. Cerebral blood flow in the sea lion (Zalophus californianus) during voluntary dives. Comp. Biochem. Physiol. 1977, 58, 11–18. [Google Scholar] [CrossRef]
- Blix, A.S.; Walloe, L.; Messelt, E.B. On how whales avoid decompression sickness and why they sometimes strand. J. Exp. Biol. 2013, 216, 3385–3387. [Google Scholar] [CrossRef]
- Kieltyka-Kurc, A.; Frackowiak, H.; Brudnicki, W. The arteries of brain base in species of the cervid family. Anat. Rec. 2015, 298, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Maloney, S.K.; Mitchell, D.; Blache, D. The contribution of rostral epidural rete mirabilevariability to brain temperature variability in sheep in a thermoneutral environment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1298–R1305. [Google Scholar] [CrossRef] [PubMed]
- De Salles, A.A.F.; Solberg, T.D.; Mischel, P.; Massoud, T.F.; Plasencia, A.; Goetsch, S.; De Souza, E.; Vinuela, F. Arteriovenous Malformation Animal Model for Radiosurgery: The Rete Mirabile. Am. J. Neuroradiol. 1996, 17, 1451–1458. [Google Scholar] [PubMed]
- Jayanti, S. Bends, Flow and Pressure Drop in; Begell House: Danbury, CT, USA, 2011. [Google Scholar] [CrossRef]
- Vogl, A.W.; Fisher, H.D. Arterial retia related to supply of the cerebral nervous system in two small toothed whales–narwhal (Monodon monoceros) and beluga (Delphinapterus leucas). J. Morphol. 1982, 174, 41–56. [Google Scholar] [CrossRef]
- Barnett, C.H.; Marsden, C.D. Functions of the Mammalian Rostral epidural rete mirabile. Nature 1961, 191, 88–89. [Google Scholar] [CrossRef]
- Grinberg, L.; Cheever, E.; Anor, T.; Madsen, J.R.; Karniadakis, G.E. Modelling Blood Flow Circulation in Intracranial Arterial Networks: A Comparative 3D/1D simulation study. Ann. Biomed. Eng. 2011, 39, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Ackerman, J.J.H.; Sukstanskii, A.L.; Yablonskiy, D.A. How the body controls brain temperature: The temperature shielding effect of cerebral blood flow. J. Appl. Physiol. 2006, 101, 1481–1488. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T. Media | T. Media + T. Intima | T. Media + T. Intima + T. Adventitia | T. Media Thickness: Total Thickness (%) | |
|---|---|---|---|---|
| Pre-carotid rete | 0.53–0.60 mm | 0.55–0.63 mm | 0.63–0.73 mm | 83% |
| Carotid rete | 0.09–0.18 mm | 0.10–0.19 mm | 0.15–0.28 mm | 62% |
| Post-carotid rete | 0.28–0.33 mm | 0.30–0.35 mm | 0.43–0.68 mm | 54% |
| CR Length (m) | mL Per Minute (mL/min) |
|---|---|
| 6 | 275.00 |
| 5 | 400.00 |
| 4 | 400.40 |
| 3 | 428.00 |
| 2 | 510.40 |
| 1 | 575.00 |
| 0.5 | 647.80 |
| Term | Estimate | Std. Error | t-Statistic | p-Value |
|---|---|---|---|---|
| Intercept | 643.347 | 9.682 | 66.450 | p < 0.001 |
| Length | −58.922 | 2.682 | −21.97 | p < 0.001 |
| Rete Length (m) | Average Pressure (mmHg) Head-Up | Average Pressure (mmHg) Head-Down |
|---|---|---|
| 6 | 313.53 | 157.51 |
| 5 | 298.52 | 150.01 |
| 4 | 286.52 | 142.51 |
| 3 | 277.52 | 136.51 |
| 2 | 264.02 | 133.51 |
| 1 | 256.52 | 127.51 |
| 0.5 | 238.52 | 120.01 |
| Term | Estimate | Std. Error | t-Statistic | p-Value |
|---|---|---|---|---|
| Intercept | 91.650 | 1.451 | 63.184 | p < 0.001 |
| Length | −12.396 | 0.402 | −30.856 | p < 0.001 |
| Term | Estimate | Std. Error | t-Statistic | p-Value |
|---|---|---|---|---|
| Intercept | 60.963 | 0.756 | 80.668 | p < 0.001 |
| Length | −6.243 | 0.209 | −29.825 | p < 0.001 |
| Radius | Area (mm2) | Number of Vessels | Total Area (mm2) |
|---|---|---|---|
| 5 | 78.54 | 1 × 10 | 78.54 |
| 2.5 | 19.64 | 2 × 5 | 39.28 |
| 2.5 | 19.64 | 4 × 5 | 78.56 |
| Radius | Circumference | Number of Vessels | Total Circumference |
|---|---|---|---|
| 5 | 31.42 | ×1 | 31.42 |
| 2.5 | 15.70 | ×4 | 62.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Walt, M.S.; Daffue, W.; Goedhals, J.; van der Merwe, S.; Deacon, F. The Rostral Epidural Rete Mirabile: Functional Significance in Blood Flow Regulatory Mechanisms in Giraffe (Giraffa camelopardalis). Anatomia 2023, 2, 138-155. https://doi.org/10.3390/anatomia2020013
van der Walt MS, Daffue W, Goedhals J, van der Merwe S, Deacon F. The Rostral Epidural Rete Mirabile: Functional Significance in Blood Flow Regulatory Mechanisms in Giraffe (Giraffa camelopardalis). Anatomia. 2023; 2(2):138-155. https://doi.org/10.3390/anatomia2020013
Chicago/Turabian Stylevan der Walt, Marna S., Willem Daffue, Jacqueline Goedhals, Sean van der Merwe, and Francois Deacon. 2023. "The Rostral Epidural Rete Mirabile: Functional Significance in Blood Flow Regulatory Mechanisms in Giraffe (Giraffa camelopardalis)" Anatomia 2, no. 2: 138-155. https://doi.org/10.3390/anatomia2020013
APA Stylevan der Walt, M. S., Daffue, W., Goedhals, J., van der Merwe, S., & Deacon, F. (2023). The Rostral Epidural Rete Mirabile: Functional Significance in Blood Flow Regulatory Mechanisms in Giraffe (Giraffa camelopardalis). Anatomia, 2(2), 138-155. https://doi.org/10.3390/anatomia2020013