Abstract
Pulse proteins are playing significant roles in the alternative protein space due to the demand for foods produced in an environmentally sustainable manner and, most importantly, due to the demand for foods of nutritious value. There has been extensive research to mimic animal-derived meat texture, flavour, mouthfeel, etc. However, there is still the perception that many of the plant-based proteins that have been texturized to mimic meat are still highly processed and contain chemicals or preservatives, reducing their appeal as being healthy and precluding any sustainable benefits. To counter this notion, the biotransformation of pulse proteins using enzymes or fermentation offers unique opportunities. Thus, this review will address the significance of pulse proteins in the alternative protein space and some of the processing aids leading to the isolation and modification of such protein concentrates in a sustainable manner. Fermentation-based valorization of pulse proteins will also be discussed as a “clean label” strategy (further adding to sustainable nutritious plant protein production), although some of the processes like the extensive use of water in submerged fermentation need to be addressed.
1. Introduction
The ubiquity of proteins in the diet of humans is immeasurable. Other than their contributions to the maintenance of a healthy outlook as the building blocks in muscles and bones, they also contribute to structural, physiological and functional roles in the body. Since the dawn of time, hunter–gatherers may have used animals (alongside plant-derived foods) as their major source of protein to subsist [1]. Animal meats continue to be a significant source of protein in the human diet, with poultry (mostly chicken), pig meat and beef accounting for more than 92% of global meat production [2], and, as of 2018, poultry accounted for the highest global production (Figure 1), excluding fish production. Production of the latter was estimated at 177 million tonnes for aquaculture fish and 90 million tonnes for captured fish as of 2020 [3]. The main reason for the current interest in plant-based protein sources is the sustainability challenges associated with the production, processing, cost and consumption of animal protein sources. Of utmost importance is the impact of animal sources on planetary health (e.g., energy, water, carbon footprint) and human health (e.g., obesity, cancers, cardiovascular diseases). For example, the ecological footprint of animal products takes up to 87% of agricultural land [4], the water footprint for 150 g of beef burger is 2350 L compared with soy protein burger at 158 L [5] and greenhouse gas and energy demands from animal sources are higher [6]. These factors have become market drivers for sourcing alternative proteins as substitutes for animal proteins in recent years. The catalyst to embrace plant-based proteins stems from the fact that there is a more conscious effort to source sustainably produced foods that are healthy and ethically produced [7,8]. It should also be noted that the demand for nutritious plant foods has increased over the years as a result of a saturation of the markets with ultra-processed foods containing chemical preservatives and chemical flavour enhancers [9,10]. Along with an abundance of unhealthy foods, other choices such as sedentary lifestyles, tobacco and alcohol consumption have led to an increase in the incidence of non-communicable diseases [11]. It is therefore not surprising that plant and plant-derived foods are being actively promoted and have become popular in recent years (if the surge in plant-based diets and plant-derived proteins as substitutes for meat proteins is any indication) [12].
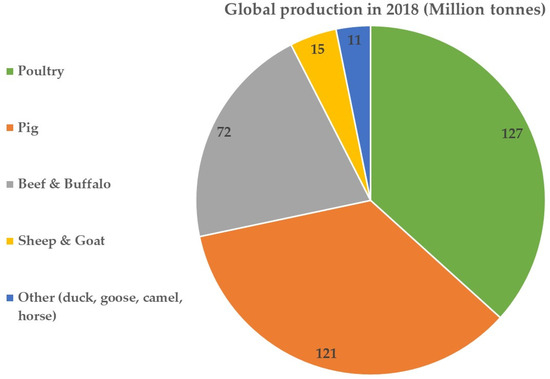
Figure 1.
Global meat production as of 2018. Source: [2].
Plant-based proteins as alternatives to animal-based proteins have now been firmly established in the marketplace, although they are not yet of significance in displacing animal-based proteins. This may be due to the fact that plant-based protein alternatives are being touted as mimics of animal-based proteins, which may still not qualify as healthy and sustainable due to the processing steps required to mimic the animal-based proteins [13]. Not being whole foods (i.e., those that are minimally processed), plant-based imitations of animal-based proteins attempt to emulate animal-based protein foods in terms of texture, appearance, taste, smell, functionality and cooking experience [14]. Thus, the adoption and acceptability of plant-based proteins as alternatives to animal-based proteins is likely to face an uphill challenge, especially from those still seeking minimally processed foods.
While the debate surrounding the merits and demerits of plant-based proteins is not likely to abate any time soon, there has nonetheless been a surging interest in pulse-based proteins to mimic animal-derived proteins. Pulses as such are not new and have been consumed for thousands of years. Mostly consumed in developing countries, pulses are leguminous crops harvested exclusively for the dry seeds and exclude those grown for oil extraction such as soybean and peanuts. Since they are leguminous crops, they are also vital in crop rotations due to their propensity for fixing nitrogen and therefore having a naturally sustainable nitrogen replenishing strategy. Pulses have received renewed interest in developed countries due to their high protein value, also evident in their increased consumption (Figure 2). The global production of pulses between 2018 and 2020 was about 90 million tonnes, with India being the largest producer, followed by Canada [15]. Interestingly, both India and Canada also had about the same consumption during the 2018–2020 period (Figure 2).
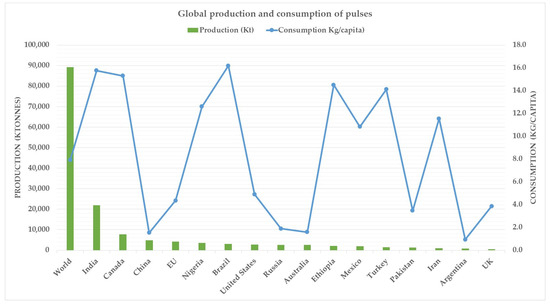
Figure 2.
Global production and consumption of pulses. Source: [15].
Plant protein sources are more abundant than animal protein sources, which makes leguminous crops a readily available protein option. For a long time, soybean was the most utilized plant protein source, but consumer concern due to allergens has led to the exploration of obtaining protein from other sources such as pulse crops. After soybeans, the most utilized plant-based protein sources are derived from pulse crops like field peas (Pisum sativum L.), lupins (Lupinus sp.), chickpeas (Cicer arietinum L.), common beans (Phaseolus vulgaris L.), lentils (Lens culinaris Medikus) and cowpeas (Vigna unguiculata L. Walp). This is because pulses are high in protein (>20%), dietary fibre and micronutrients, such as folate, iron, zinc, magnesium, potassium and selenium, with <3% fat content [16,17,18,19]. Pulses contain high-quality amino acids distributed proportionately within the different classes, and consequently benefit the consumer nutritionally, therapeutically and techno-functionally. Furthermore, pulses are a good source of bioactive compounds (i.e., phenolic compounds and phytosterols). Pulse protein is limited in sulphur-containing amino acids (i.e., cysteine and methionine) and high in lysine, glutamic acid and aspartic acid [20]. However, combining pulse proteins with other protein sources like cereals is an efficient way to improve the essential amino acid composition.
Even though the benefits of pulse proteins in the human diet have been established, there are still some inherent limitations impairing the use of these proteins in food systems. These include low net surface charge (affecting solubility), the presence of antinutritional compounds (causing low digestibility and bioavailability) and odour-causing components responsible for off-flavours. Nonetheless, several techniques (physical, chemical and biological) have been used to modify the structural and physicochemical properties of pulse proteins, thereby enhancing the nutritional, functional and sensory properties [21,22]. Fermentation technology is also an effective method that has been shown to improve the quality of pulse protein and to expand its usage as a food ingredient and additive [23,24].
In the context of healthy and sustainably produced dietary plant proteins, pulse proteins are undoubtedly the source crops of choice. However, as mentioned above, the reticence in consumption of plant-based proteins mimicking animal-based proteins may be perceived as being equivalent to consuming processed foods and therefore deemed not healthy. Faced with this prospect, exploring alternative ways to derive maximum benefits from pulse proteins would be valuable. Traditional and ethnic cooking approaches may not be enough to derive all the nutritional benefits sequestered within the pulse proteins. Therefore, the objective of this review is to provide an overview of pulse proteins, limitations of their nutritional benefits and modification approaches to valorize their nutritional profiles and their significance in the context of contributing to a sustainable environment, sustainable production and sustainable diet. In this regard, some of the primary processing technologies using chemical and physical approaches are revisited, with an outlook on potential sustainable practices. However, more sustainable biological approaches are likely to be the path forward. These include enzymatic and fermentation strategies, with the fermentation approach being the most preferred. While fermentation is a versatile approach for pulse protein modification, the major bottleneck in its implementation is addressing scale-up challenges and techno-economic sustainability. Indeed, the commercial success of fermentation-derived pulse proteins as sustainable ingredients in the alternative protein space will depend on successful and economical process optimization for scale-up.
2. Pulse Proteins in the Alternative Protein Space and Sustainability
As mentioned earlier, the use of pulse proteins as an option for a sustainable environment is very encouraging. Furthermore, as healthy and nutritional alternatives, pulse proteins are gaining importance. It is therefore important to highlight the attributes and processes that have allowed pulse proteins to attain such prominent roles in the alternative protein space. It is also imperative to understand the underlying processes that enable their transformation into high-quality proteins and their fit into the overall scheme of sustainability, even though animal protein sources (i.e., beef, poultry and pork) are still popular options in North America and are much preferred by consumers because of the innate sensory and techno-functional properties they offer [25]. However, animals are not a sustainable source of proteins due to their impacts on the following: (1) environment (global pressure on land, high water and energy consumption, greenhouse gas emission and high carbon foot print), (2) human health (high saturated fats, cholesterol and allergens) and (3) economy (cost/availability) [26]. Additionally, the production of animal protein is not efficient as some animals may require to be fed up to 15 Kg of plant materials to be able to produce 1 Kg of meat [27]. With the world population projected to reach 10 billion by 2050 [28], meat production has been projected to rise to 410 billion kg by the year 2050 [29]. Thus, this increased demand for more food could lead to an increased impact on the environment and increase the risks of food scarcity. From 2000 to 2018, the consumption of protein-rich food increased globally by 40% due to consumer awareness in benefits derived from a protein diet [25]. Current protein consumption by adults is estimated at 26 Kg of protein annually and has been projected to increase to 33 Kg by 2025 [25].
Pulse crops have several advantages over animal sources such as low allergens, low saturated fats, reduced emissions of greenhouse gases, lower carbon footprint, being readily available, having cost-efficient production and being an alternative for vegans and vegetarians [30,31]. For example, the partial replacement of lean meat with cooked lentil puree was shown to reduce the impact on the environment by ~33%, reduce production cost by ~26% and increase nutrient density by ~20% [32]. With their ability to fix nitrogen, pulses do not need fertilizers for growth and are important in crop rotations for the maintenance of soil health and moisture [33]. This is a significant incentive to grow more pulse crops as an alternative protein source for consumption. Furthermore, they contribute up to 10% of protein to human nutrition globally [34] and, in Canada, peas, lentils and chickpeas are having significant economic and commercial impacts, with about 5.5 million tonnes of pulses grown annually, cash receipt of CAN$1.7 billion and 75% production export [35]. The economic returns and production are likely to continue to increase, as will the demand for pulse proteins.
2.1. Advances in Fractionation of Pulse Proteins
While the importance of pulse crops in sustainable agriculture is unequivocal, the sustainable fractionation of pulse proteins to derive maximum nutritional benefits needs to be addressed. Methodologies hitherto developed for optimal benefits have centred around the enrichment of pulse proteins to mimic the propensities of animal proteins. Thus, fractionation methods have been developed to separate the proteins from the flours. Before fractionation, the seeds are pretreated by cleaning, drying, sorting, dehulling and milling into flour to preserve the functionalities of the protein ingredients [36]. These are well-established processes in the milling industries prior to proceeding to further downstream processes (Figure 3). Dehulling of pulse seeds, for example, has been shown to improve protein extractability, reduce antinutritional materials, increase digestibility and improve functional properties [37]. Some of these processes are therefore essential and finding alternatives is likely to be challenging. Where an important rethink of the scope of sustainable solutions can be envisaged is in the fractionation of pulse proteins. There are two methods employed in protein fractionation viz. dry or wet fractionation to produce protein ingredients with varying levels of purity and functionality (Figure 3).
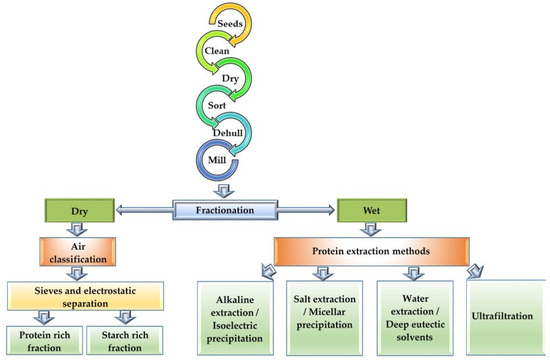
Figure 3.
Dry and wet protein fractionation techniques from pulses.
Dry fractionation involves two steps, namely milling, where the seeds are ground into powder to release the starch from the protein-rich matrix, and air classification, where sieves are used to separate the starch-rich fractions from the protein-rich fractions [37,38]. This method is guaranteed to produce pulse proteins with high nativity and good hydration properties, and only negligible levels of water and energy are required for processing, but the protein is of low purity (43–75%) [39,40]. Thus, despite the low purity, dry fractionation is a sustainable process with low resource requirements for pulse protein fractionation. Improvements to the basic dry fractionation process have been conducted to improve purity, wherein a two-step air classification technique coupled with electrostatic separation increased protein purity of dry fractionation products by 16% [38]. Another study [41] reported that a hybrid method performed by combining dry and wet fractionation produced proteins with 87% purity. The method used 5.5 times less water and energy than normally used in wet fractionation (discussed below). The separation of smaller protein bodies from larger starch granules using an air classifier is dependent on wheel speed, because smaller particles preferentially pass through the air classifier [42]. Consequently, protein-enriched fractions of peas, cowpeas and mung beans were achieved at higher wheel speeds, and the functional properties were dependent on the particle size of the fractions [42]. However, there is still a need for more studies regarding dry fractionation, as only a few recent studies have addressed the technical challenges related to efficiency improvement.
Wet fractionation, on the other hand, has the potential to produce protein with purity >90% and yield up to 90% [41]. However, due to high water and energy usage, the overall cost of production is high, making it more unsustainable compared with dry fractionation. The most common wet fractionation method is alkaline extraction–isoelectric precipitation (AE-IP), where solubilization is achieved at a high pH (8–11), causing denaturation and irreversible precipitation of some of the proteins at pH 4.5 [43,44]. In a study using reduced water with the AE-IP method, it was reported that the protein yield also decreased [45]. Irrespective of the water requirement, one study reported that the AE-IP extraction of pea protein at pH 9 reduced the beany flavour, with a decline in the level of flavour markers such as alcohols, aldehydes, ketones and pyrazine [46]. Unlike the dry fractionation methods, there have been advances in mild wet fractionation methods, which would allow for the preservation of protein nativity, the enhancement of functional properties and the reduction of production cost. One study [47] demonstrated that combining ultrasound technology with AE-IP improved the purity of lentil protein. Another study combined AE-IP with a modified salt-solubilization technique and led to the isolation of high purity (>80%) legumin and vicilin from pulse flours [43]. It is thus evident that, in wet fractionation, water requirement is important for high protein purity and for the enhancement of the functional properties of proteins. Therefore, more sustainable wet fractionation options need to be investigated. For example, it was reported that several washes of pulse flour with water at a neutral pH separated starch-rich fractions from protein-rich fractions and, with ultrafiltration, 75% protein could be recovered [48]. Although the purity of protein obtained by water fractionation was low, other solutes in the fraction enhanced the functionality of the pulse protein [48]. Furthermore, the processed water could be recycled to minimize waste [48]. Hybrid methods combining wet and dry fractionation may also be other options in the interim. The use of water and deep eutectic solvents (DESs) in mild wet fractionation methods also show promise. DESs are mild solvents used in the extraction of phenolic compounds and sugars from other plant protein sources [49,50,51]. Unlike acids and bases, DESs are considered green technology and more friendly to the protein ingredient and the environment, providing more sustainable alternatives for the wet fractionation of pulse proteins.
2.2. Pulse Protein Modification for Modulating Structure and Enhancing Functionality
As evident from the previous sections, pulse crops, while sustainably produced, do offer challenges when it comes to the fractionation of their proteins. Further downstream of fractionation, other challenges regarding enhancing the functionality of the pulse proteins are encountered, especially in terms of sustainable practices. In this section, we delve into modification techniques to enhance protein functionality. The main pulse protein fractions are the storage proteins or globulins (60–80%), metabolic proteins or albumins (10–25%) and a low percentage of prolamins and glutenins (3–5%) [52]. Consequently, native pulse protein has a very compact rigid structure and low surface net charge, with impaired functionalities like solubility, emulsification, foaming and gelation. Unlike animal proteins that have high α-helical structures, plant proteins contain higher β-sheets, further increasing the rigidity of the structure. Thus, structural modulation is required to improve flexibility. Other limitations include the presence of antinutritional factors that could impair digestibility and off-flavour compounds that affect the overall organoleptic properties of the protein. For these reasons, several modification methods have been devised to improve functionality. However, the sustainability of these methods still needs to be addressed.
The following three modification methods are currently available: physical (thermal, high-pressure, extrusion, microwave, sonication, cold plasma, ultrasound), chemical (phosphorylation, glycation, conjugation) and biological (germination, enzymatic hydrolysis, fermentation) (Table 1). These modification techniques can be conducted independently or in combinations thereof. For example, a hybrid treatment of yellow pea protein using heat and ultrafiltration (>50 kDa fractions) at different pHs enhanced solubility by 60% [53]. A cold plasma treatment of grass pea protein isolate at a higher voltage (18.6 kVpp) for 60 s compared with a lower voltage (9.4 kVpp) for 30 s improved the emulsification properties due to the increase in the density of charged and reactive species [54]. The higher voltage increased the content carbonyl groups, disulphide bonds, di-tyrosine crosslink and surface charge, which led to the formation of small oil droplet sizes surrounded by thick elastic interfacial layers [54]. In effect, cold plasma treatment would be considered a sustainable green technology approach for the pretreatment of plant proteins [55].

Table 1.
Different protein modification methods and their advantages and disadvantages.
A widely utilized physical modification technology for pulses is extrusion technology for the preparation of ready-to-eat (RTE) puff snacks, pasta and extruded meat replacements. Extrusion combines several processes such as mixing, cooking, kneading, shearing, shaping and forming in one unit. The structure and texture of extrudates are dependent on the extrusion parameters (moisture content, screw speed), cooking conditions (temperature, pressure) and chemical composition of the ingredients [74]. For example, high moisture extrusion is preferred for meat analogues from pulse protein because of the need for a moist and fibrous meat-like structure [74]. Extrusion cooking of pulses, like most thermal methods, reduces the quantity of flavour compounds in the raw flour. Extrusion cooking of pulse flours (red lentils, green pea and chickpeas) into puff snacks and pasta showed that phenolic compounds were reduced by 45%, while flavonoids were reduced by 41% [75]. However, cell wall bound phenolic compounds and antioxidant activity increased. In the same study, lentils were shown to exhibit high puff snack quality (high expansion 310 mm2 × 114 mm2) and a high content of flavonoid compounds after treatments [75]. On the other hand, low moisture extrusion cooking to produce puff snacks from composite flours of protein-rich fractions of pulses and pseudocereals resulted in reduced sectional expansion as the protein content increased [76]. Lupin, which is also high in protein, produced a protein-rich snack with optimal expansion and acceptable sensory properties in combination with buckwheat [76]. While extrusion has proven to have tremendously enhanced pulse protein functionality, the above examples may indicate that it is not a sustainable approach for pulse protein modification. However, this is not necessarily the case, as extrusion is considered an all-encompassing technology using little water for upcycling food by-products to valuable foods [77].
In chemical treatments, protein modification is achieved by selectively adding new moieties to the active groups (i.e., amino, carboxyl, disulphide, sulphide, sulphydryl, imidazole, indole, etc.) inherent in the protein through covalent and non-covalent interactions [78]. Acylation, one of the most commonly used chemical modification methods, increases protein flexibility by unfolding the structure through the transfer of the acyl group to the amine or hydroxyl group in the amino acid, wherein the aromatic–aliphatic residue balance increases [79]. Similarly, the chemical conjugation of pea protein with guar gum enhances the emulsification properties by 100% when compared with the emulsion capacity (58%) and stability (48%) of untreated pea protein [80].
Sustainable alternatives for the biotransformation of pulse proteins include enzyme treatments, germination and fermentation. Fermentation technology is an ancient traditional practice to improve edibility, palatability, flavour, digestibility and the shelf-life of food and is discussed further in Section 3. With regards to enzymes, which are generally purified from wild-type or bioengineered microorganisms in bioreactors, numerous modified food products have been produced, including the functional modification of pulse proteins, as reviewed in [81]. For example, deamidation with glutaminase effected conformational changes in pea protein isolates with increased structural flexibility, which could have been due to the reduction in β-sheets and β-antiparallel sheets and the increase in β-turns [82]. Given that the enzymes are produced in bioreactors adds to the cost of production, and also creates another layer of resource-demanding technology for production and enzyme purification.
The germination of pulse seeds has been reported to positively impact flavour, digestibility, palatability and the amino acid profile [83,84]. Germination leads to chemical changes in the seed, with the enzymatic breakdown of macromolecules such as starch, protein and lipids into smaller molecular weight compounds. Controlled sprouting of chickpea flour at the industrial scale significantly reduced antinutritional compounds, increased free minerals and vitamins and improved the release of peptides [83]. The germination of pulse seeds over 2 and 4 days showed a slight increase in the protein content (>3%), a significant increase in acidity and a decrease in phytic acid, which led to an increase in free minerals [85]. A 4-day germination of chickpeas increased the composition of phenolic compounds and antioxidant activity through a 100-fold increase in the isoflavones content [86]. Similarly, flours produced from pulse seeds that were germinated over 6 days exhibited enhanced pasting properties, water adsorption capacity, and protein content, while the lipid content was reduced [87]. The hybrid use of germination and extrusion (140 °C and 800 rpm) produced pulse-based high-moisture meat analogues (HMMAs), with a significant reduction in the bean-related flavours (i.e., 1-octen-3-ol and 2,4-decadienal, hexanal, 1-hexenol, 2-nonenal and (E,E)-2,4-nonadienal), as analyzed by gas chromatography–mass spectrometry/olfactory (Usman et al., 2023). However, some secondary flavour compounds were formed after the treatments, and germination slightly increased the umami and the bitter taste in the HMMAs [88]. The above discussion has certainly highlighted the need for improved sustainability of both physical and chemical modification methods for pulse proteins. However, the examples of germination-based pulse protein functionality alteration, while not completely effective, offer opportunities for further improvements in hybrid systems for a more sustainable approach.
3. Fermentation-Based Pulse Protein Valorization
Despite the recognized value of pulse proteins in the alternative protein space, there is still a need to enhance their techno-functional properties to further extend applications in the food industry and, importantly, to sustainably produce these proteins. Furthermore, there is still a perception that pulse proteins are highly processed and not healthier than other processed foods; therefore, addressing a sustainable diet strategy is also imperative. Sustainable diets are considered to have minimal impacts on the environment while also addressing food and nutritional security and preserving biodiversity among other sustainable practices, as reviewed in [89]. Thus, to further improve pulse protein functionality, fermentation-based strategies have been explored as “clean label” alternatives, potentially fitting into a sustainable diet scheme. Fermentation has been in existence for thousands of years, mainly as a means of food preservation and shelf-life extension. Unbeknownst to the practitioners of those times were the underlying scientific principles that governed the textural, sensorial and other improved properties associated with the fermented foods. Simply stated, fermentation is a process by which microorganisms are able to breakdown complex molecules into simpler more bioavailable forms and, in so doing, are also able to derive nutrients and energy for their own growth. Lactic acid bacteria (LAB) are the most commonly known of these microorganisms in the preservation and production of value derived foods [90]. Other than LAB, many fungal and yeast strains can also be used for the production of fermented foods. The science behind the fermentation process has advanced tremendously over the years, with growing sophistication in the engineering designs of fermenters for the efficient production of fermented foods. Strain selection and engineering have also led to further improvements in fermentation, leading to the emergence of precision fermentation wherein synthetic biology has been driving the production of specific biomolecules in microorganisms. The fermentation process, other than altering texture and aroma profiles, can also affect food composition due to the metabolic processes and release of enzymes and other chemicals, which can breakdown complex molecules within the food product [91]. LAB also release small aromatic compounds and exopolysaccharides that contribute to the aroma and flavour of the fermented foods [92]. Fungal strains such as Aspergillus and Rhizopus contribute to the texturization of the food, in addition to contributing to mycoproteins that further enhance the nutritional value of the fermented foods [93].
The fermentation process is in itself simplistic and, in recent years, has been touted as a more sustainable approach to food production [94]. However, it requires careful manipulation to prevent undesired or harmful microorganisms from overtaking the microorganism of interest. This is straightforward in a closed fermentation system such as submerged fermentation (SmF). Solid state fermentation (SSF), on the other hand, is more prone to contamination, especially on larger scales. These two systems of fermentation have their advantages and disadvantages. SmF uses high moisture content (free-flowing culture medium), while SSF uses low moisture content (no free-flowing liquid). In SSF, the substrate itself can be the source of nutrients or the substrate can be inert with nutrient media immobilized to it. In terms of resources, SmF requires high energy due to the large volumes of water required, alongside the operational controls associated with fermenters. SSF generally requires less resources and low energy (Table 2).
Table 2.
Differences between solid state fermentation (SSF) and submerged fermentation (SmF) (modified from [95,96]).
In terms of growth effectiveness of microorganisms, fungal species are more amenable for cultivation in SSF than bacterial strains, which are more adapted to higher levels of water content for efficient growth [95]. Downstream processing of products post-fermentation is also less resource demanding in SSF compared with SmF [95]. In the latter, a dewatering step by centrifugation or some other separation techniques would be necessary if the final product is the biomass. For SSF-derived products, downstream processing is simple, involving drying or direct processing or incorporation into food products. However, one of the main disadvantages of SSF is the challenge in maintaining uniform temperature distribution and aeration on a large scale. Nonetheless, due to its lower resource requirements, it is sometimes more desirable for a lower cost of operation. For pulse protein fermentation, depending on the objective, either SSF or SmF can be used. SSF is more appropriate for a texturized protein, whereas SmF is better suited for the production of fermented beverages. Both LAB and fungal species can be used for pulse protein valorization. Generally, whether LAB or fungal strains are used, pulse protein biotransformation through fermentation influences a number of attributes such as increases in bioavailable proteins and peptides, changes in protein solubility, changes in emulsification and foaming properties, changes in water and oil holding capacities, changes in flavour and taste (e.g., decrease in bitterness, beany taste, increase in umami) and change in aroma profiles (Figure 4). Thus, from a sustainable perspective, SSF would be a preferred strategy compared with SmF, but eventually efficiency and economic feasibility and returns will determine the choice of which fermentation method to use. To gain a better perspective on the significance of fermentation in valorizing pulse proteins in the alternative protein space, in the following sections, we discuss some specific applications.
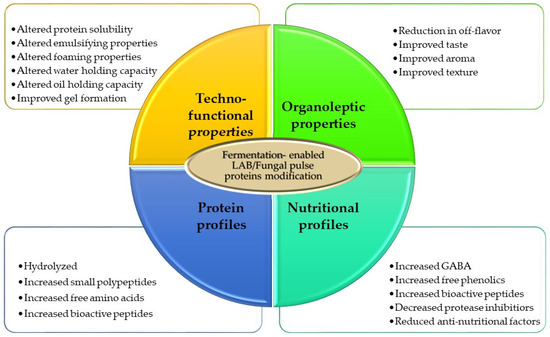
Figure 4.
Changes in pulse proteins as a result of fermentation with LAB and/or fungal strains.
3.1. Lactic Acid Bacteria (LAB) Fermentation of Pulse Proteins
The benefits of lactic acid bacteria (LAB) fermentation of pulse proteins include increases in bioactive peptides, resistant starches, soluble fibres, antioxidants and γ-aminobutyric acid (GABA), while leading to decreases in tannins, phytases, saponins, raffinose family oligosaccharides, vicine, convicine and mycotoxins [97]. LAB also produce antimicrobials in the form of organic acids (lactic acid, citric acid), bacteriocins and antifungal peptides. Other than contributing to these added sensorial, nutritional and functional properties, LAB (being categorized as GRAS (generally recognized as safe)) are versatile microorganisms for use in the food industry, and newer strains with additional attributes are continually being identified [98].
One of the main deterrents to the consumption of pulses is the occurrence of antinutritional factors (ANFs) such as raffinose oligosaccharides (RFO), phytic acid, tannins, saponins, lectins, protease inhibitors, etc., which can be toxic, make the pulses unpalatable, reduce the bioavailability of nutrients or cause stomach or intestinal discomfort [99]. In a study of the fermentation of air-classified faba bean protein-rich fraction by Lactobacillus plantarum strain VTT E-133328 [100], it was shown that the vicine and convicine concentrations were significantly decreased by more than 91%, with an 86% reduction in trypsin inhibitor activity and a 40% reduction in tannins. Fermentation also led to an increase in free amino acids, particularly essential amino acids and GABA, with improvement in in vitro protein digestibility and a decreased hydrolysis index. Similarly, using a consortium of LAB strains, 67% and 27% reductions in phytic acid and RFOs, respectively, were achieved in Lupinus mutabilis, a high protein value crop in South America [101]. Trypsin inhibitor activity was also reduced in chickpea (desi and kabuli) and faba bean by fermentation with LAB consortia from lyophilized yogurt culture, with a corresponding increase in protein digestibility [102]. Fermentation with other LAB such as Lactobacillus plantarum and Lactobacillus brevis in kidney bean, chickpea, pea, lentil and grass pea led to decreased RFOs and condensed tannins [103]. LAB SSF of chickpea protein concentrate also led to a decrease in raffinose and stachyose by 88.3–99.1%, with verbascose being undetectable [104]. Thus, compared with physical and chemical treatments, LAB fermentation can sustainably reduce ANFs and improve pulse protein functionality.
While eliminating or reducing ANFs leads indirectly to improved nutritional profiles, the production of other valuable compounds as a result of fermentation contribute directly to improved nutritional attributes, without the need for supplementation, which may lead to the perception of overprocessing or unsustainable practices. For example, bioactive compounds were shown in extracts from Lactobacillus plantarum-fermented kidney bean (Phaseolus vulgaris var. Pinto), wherein increased GABA content and angiotensin converting enzyme inhibitory (ACEI) activity were observed, an indication of potential antihypertensive activity [105]. Similar observations in increased ACEI activities were made with navy bean milk fermentation using several LAB strains [106]. Bioactive peptides were similarly produced during fermentation of legume proteins due to hydrolysis of the complex proteins or even released by the microorganisms being used [107]. In a study of several fermented legume species, sourdough using Lactobacillus plantarum C48 and Lactobacillus brevis AM7, lunasin-like polypeptides were observed in the extracts and shown to inhibit the proliferation of human adenocarcinoma Caco-2 cells [108].
3.2. Fungal Fermentation of Pulse Proteins
LAB fermentation-type profile changes also occur as a result of fungal fermentation. For example, fermentation of common beans with Rhizopus oligosporus led to a reduction in ANFs [109]. A reduction in ANFs such as phytic acid was also observed in lentils with the edible oyster mushroom, Pleurotus ostreatus, under SSF conditions [110]. Using three different Rhizopus species, independently or in combinations, it was demonstrated that ANFs such as tannins, phytates and trypsin inhibitors could be reduced in Bambara nut (Voandzeia subterranean L.) [111]. Other studies with Rhizopus oligosporus in pulses such as desi and kabuli chickpea and pigeon pea resulted in an increase in proteins and essential and non-essential amino acids [112], similar to some of the techno-functional changes observed with LAB fermentation. Increases in protein content were also observed with Aspergillus oryzae fermentation of a pea-processing by-product still containing about 18% protein [113]. However, the study attributed the additional protein to the fungus, thereby creating a vegan mycoprotein. This presents an opportunity for upcycling side stream pulse proteins for additional protein production and fits into a circular economy concept in the total use of by-products from the grain processing industry.
In addition to the contribution of mycoproteins to the already protein-rich pulses, a combination of fungal and pulse proteins further enhances the texture of the fermented product. This has become particularly desirable in the alternative protein space to mimic the texture of animal-based meats. The most extensively researched and studied mycoprotein is that derived from Fusarium venenatum and commonly marketed under the brand name Quorn [114]. In the UK, there was a 10-year evaluation period prior to the approval of mycoprotein for food in 1983 and, in the US, the FDA approved it with GRAS designation in 2002 [114]. Since then, a number of other fungal species have been shown to be rich in mycoproteins for consumption. Irrespective of the mycoprotein content of such fungal species, the overarching advantage of such fungal–pulse protein interactions is the production of a texturized nutrient- and flavour-enhanced food. From conventional tempeh-type fungi, such as Rhizopus sp., and koji-type fungi such as Aspergillus sp., edible mushrooms are now also being used for the biotransformation of plant proteins. For example, a pea–rice protein hybrid blend fermented with shiitake mushroom mycelia exhibits improved protein digestibility, enhanced nutritional value and improved organoleptic properties [115,116]. These examples further provide compelling evidence for the use of pulse proteins as sustainable alternatives when combined with fungal fermentation systems.
3.3. Mixed Culture Fermentation of Pulse Proteins
Mixed culture fermentation is also receiving renewed interest due to synergistic effects in the biotransformation of food products for sustainable diets. Fermentation-based enhancement of the organoleptic properties of foods is well recognized. With the advent of genomics tools, many of the microorganisms used have been characterized and their roles elucidated. However, this deeper understanding of the microorganisms and their metabolic activities and interactions with the food matrices has also led to the use of single species/strains in food fermentation [117]. While this has brought about tailored food products with strain-specific biotransformation, the benefits derived from the consortia of microorganisms have been lost. Indeed, just as the domestication of plants and animals occurred at the dawn of the agricultural revolution [118], so too did the domestication of bacteria, yeast and fungi much later, leading to further improvements in food organoleptic properties and, importantly, preservation, but achieving unknowingly consortia of microorganisms or “starter cultures” [119,120].
Thus, such starter culture concepts have been revisited and several studies have demonstrated synergistic effects in the improved attributes of the fermented products. For example, using bacterial strains of Lactobacillus plantarum L1047 and Pediococcus pentosaceous P113, Schindler et al. [121] demonstrated improvements in aroma profiles due to a decrease in n-hexanal content, which led to a reduction in or masking of off-flavours. With a hybrid cow milk–pea protein strategy for yogurt consistency assessment using starter cultures of LAB, higher ratios of pea proteins led to higher acidity, greater syneresis and lower firmness, but a 10% pea protein approached the consistency of yogurt [122]. Similarly, with consortia including yeast species, LAB and other bacteria, 100% pea protein emulsions were shown to emanate a roasted/grilled aroma due to the release of volatile compounds, and 50:50 pea–milk emulsions showed a fruity and lactic acid aroma [123]. Again, a reduction in hexanal was observed. Co-fermentation with yeasts and LAB also led to a reduction in off-flavours in 4% pea protein solutions, with additional yeasts contributing to the production of esters [124]. Therefore, fermentation with consortia of interspecific microorganisms offers new opportunities for pulse flavour and taste and aroma profile enhancement while simultaneously contributing to other benefits such as protein hydrolysis and reduction in ANFs, etc.
4. Scale-Up Challenges in Pulse Protein Fermentation and Implementation of Sustainable Practices
One of the most overlooked aspects in fermentation is the translation from small-scale shake flask research to large-scale production. Often, the cost of production is not envisaged on large scales and the cost of production quickly becomes extensive due to the use of expensive ingredients at the process development and optimization stage, as reviewed in [125], thus rendering the process unsustainable. With regard to pulse proteins, integrating the fractionated/isolated proteins into a large-scale fermentation process is still not fully realized, whether SmF or SSF. For example, one of the important aspects of microbial fermentation is the maintenance of sterility. However, very few studies have looked at the effects of autoclaving or high temperatures on the techno-functional properties of pulse proteins. Many studies have indicated that autoclaving is in fact beneficial for the removal of antinutritional factors, as reviewed in [126,127]. While it is conceivable that the fermented protein will be heat-treated/cooked, any microbial contamination during fermentation may not be of any consequence. Similarly, if the fermentation duration is short, whether SmF or SSF, non-autoclaved protein substrates may be used. It is therefore imperative that some baseline studies be established for pulse protein fermentation and the effects of autoclaving, especially on structure and function. Some early research into autoclaving of pulse proteins have indicated a reduced lysine content but no appreciable changes in other amino acids [128,129]. However, another study did not find any changes in amino acid composition due to autoclaving [130]. However, the other overarching concern is that autoclaving and high temperatures imply high energy resources and therefore unsustainability. Further studies need to be conducted to address aspects of pulse protein fermentation under more of a pasteurization type of process to avoid the use of high energy resource-demanding processes.
Another aspect to be considered in scale-up in SmF systems is the high volumes of water used. If the pulse protein is destined to be used as a fermented beverage, this is likely not an issue. However, in applications where the pulse protein together with the microbial biomass needs to be used for texturized alternatives, then significant dewatering needs to be undertaken. Attempts to reuse the water from post-fermentation processes are now gaining interest to render fermentation more sustainable. For example, cheese whey permeate was used for the production of oil from oleaginous yeast Cutaneotrichosporon oleaginosus [131]. Such approaches regarding the reuse or treatment of fermentation waste broth will make SmF more sustainable on larger scales.
5. Conclusions
The need for sustainable sources of proteins and sustainable diets has led to new opportunities to explore plant-based proteins. Pulse proteins have become major sources of such research and discovery efforts. The challenge to date, however, in fully utilizing the potential of pulse proteins has been due to their flavour profiles, aromas, digestibility, antinutritional factors, etc. With different processing methods, such discrepancies can be addressed to some extent but may lead to extensive resource requirements, making the process unsustainable, or may add to the cost of processing. Furthermore, such processing approaches are sometimes viewed negatively due to the use of chemicals, even if they are considered food safe. Thus, approaches employing biological transformations such as enzymes and fermentation offer “clean-label” solutions to alter pulse protein functionality and quality. Fermentation-based approaches have recently gained renewed interest precisely because of their ability to address the multitude of undesirable quality attributes in pulse proteins by judiciously selecting the appropriate bacteria or fungal GRAS strains for biotransformation. An added advantage of fungal strains is that they also contribute to further enhance the protein content of the final product by contributing their own proteins. However, the techno-functional transformation of pulse proteins using fermentation will only be fully realized with a more targeted approach to enhance flavour and reduce ANFs. Furthermore, the commercial success of pulse protein fermentation will only by attainable if there is a seamless transition from fractionation to large-scale fermentation and making the process more sustainable, considering that in these processes such as SmF large volumes of water are used.
Author Contributions
Conceptualization, S.G., N.A., Y.W., M.Ç.T. and M.T.N.; writing-original draft preparation, S.G., N.A., Y.W., M.Ç.T. and M.T.N.; writing-review and editing, S.G., N.A., Y.W., M.Ç.T. and M.T.N.; project administration, S.G.; funding acquisition, M.Ç.T. All authors have read and agreed to the published version of the manuscript.
Funding
Research at Saskatchewan Food Industry Development Centre is supported by funds from the Saskatchewan Ministry of Agriculture, the Saskatchewan Pulse Growers and Protein Industries Canada.
Conflicts of Interest
Seedhabadee Ganeshan, Yingxin Wang, Mehmet Ç. Tülbek are employed by the Saskatchewan Food Industry Development Centre, a non-profit organization, which supports the agi-food industry. The above authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. The funding agencies had no role in the design of the study; in the collection, analysis, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Bunn, H.T.; Ezzo, J.A. Hunting and Scavenging by Plio-Pleistocene Hominids: Nutritional Constraints, Archaeological Patterns, and Behavioural Implications. J. Archaeol. Sci. 1993, 20, 365–398. [Google Scholar] [CrossRef]
- TheWorldCounts. The World Counts—Impact through Awareness. Available online: https://www.theworldcounts.com/challenges/consumption/foods-and-beverages/world-consumption-of-meat (accessed on 26 May 2024).
- FAO. The State of World Fisheries and Aquaculture. 2022. Available online: https://www.fao.org/3/cc0461en/online/sofia/2022/world-fisheries-aquaculture.html (accessed on 22 May 2024).
- Espinosa-Marrón, A.; Adams, K.; Sinno, L.; Cantu-Aldana, A.; Tamez, M.; Marrero, A.; Bhupathiraju, S.N.; Mattei, J. Environmental Impact of Animal-Based Food Production and the Feasibility of a Shift Toward Sustainable Plant-Based Diets in the United States. Front. Sustain. 2022, 3, 841106. [Google Scholar] [CrossRef]
- Ercin, A.E.; Aldaya, M.M.; Hoekstra, A.Y. The water footprint of soy milk and soy burger and equivalent animal products. Ecol. Indic. 2012, 18, 392–402. [Google Scholar] [CrossRef]
- Xu, X.; Lan, Y. A comparative study on carbon footprints between plant- and animal-based foods in China. J. Clean. Prod. 2016, 112, 2581–2592. [Google Scholar] [CrossRef]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Barreiro, M.F.; Morales, P.; Ferreira, I.C.F.R. Adding Molecules to Food, Pros and Cons: A Review on Synthetic and Natural Food Additives. Compr. Rev. Food Sci. Food Saf. 2014, 13, 377–399. [Google Scholar] [CrossRef] [PubMed]
- McClements, D. Future Foods: How Modern Science Is Transforming the Way We Eat; Copernicus: Cham, Switzerland, 2019; p. 366. [Google Scholar]
- WHO. Noncommunicable Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases (accessed on 22 May 2024).
- McClements, D.J.; Grossmann, L. A brief review of the science behind the design of healthy and sustainable plant-based foods. npj Sci. Food 2021, 5, 17. [Google Scholar] [CrossRef]
- Al-Heeti, A. Whole Foods CEO Says Plant-Based ‘Meat’ Is Unhealthy. Available online: https://www.cnet.com/health/nutrition/whole-foods-ceo-says-plant-based-meat-is-unhealthy/ (accessed on 5 September 2023).
- Bryant, C.J. Plant-based animal product alternatives are healthier and more environmentally sustainable than animal products. Future Foods 2022, 6, 100174. [Google Scholar] [CrossRef]
- OECD; FAO. Table C.46—Pulses Projections: Production and Food Consumption. In OECD-FAO Agricultural Outlook 2021–2030; OECD Publishing: Paris, France, 2021. [Google Scholar] [CrossRef]
- Brun, P.; Camacho, M.; Perea, F.; Rubio, M.J.; Rodríguez-Navarro, D.N. Characterization of Spanish chickpea genotypes (Cicer arietinum L.): Proximate, mineral, and phenolic compounds composition. Eur. Food Res. Technol. 2024, 250, 1007–1016. [Google Scholar] [CrossRef]
- Johnson, N.; Boatwright, J.L.; Bridges, W.; Thavarajah, P.; Kumar, S.; Thavarajah, D. Targeted improvement of plant-based protein: Genome-wide association mapping of a lentil (Lens culinaris Medik.) diversity panel. Plants People Planet 2024, 6, 640–655. [Google Scholar] [CrossRef]
- Carvalho, M.; Carnide, V.; Sobreira, C.; Castro, I.; Coutinho, J.; Barros, A.; Rosa, E. Cowpea Immature Pods and Grains Evaluation: An Opportunity for Different Food Sources. Plants 2022, 11, 2079. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, D.; Vurro, F.; Santamaria, M.; Garzon, R.; Rosell, C.M.; Summo, C.; Pasqualone, A. Effect of dry-fractionated pea protein on the physicochemical properties and the nutritional features of gluten-free focaccia flat bread. LWT 2023, 182, 114873. [Google Scholar] [CrossRef]
- Sá, A.G.A.; Wan, Z.; Jha, A.B.; Gali, K.K.; Warkentin, T.D.; House, J.D. Influence of different amino acid scoring patterns on the protein quality of field peas. J. Food Compos. Anal. 2024, 127, 105938. [Google Scholar] [CrossRef]
- Locali-Pereira, A.R.; Caruso, Í.P.; Rabesona, H.; Laurent, S.; Meynier, A.; Kermarrec, A.; Birault, L.; Geairon, A.; Le Gall, S.; Thoulouze, L.; et al. Pre-treatment effects on the composition and functionalities of pigeon pea seed ingredients. Food Hydrocoll. 2024, 152, 109923. [Google Scholar] [CrossRef]
- Dnyaneshwar Patil, N.; Bains, A.; Kaur, S.; Yadav, R.; Ali, N.; Patil, S.; Goksen, G.; Chawla, P. Influence of dual succinylation and ultrasonication modification on the amino acid content, structural and functional properties of Chickpea (Cicer arietinum L.) protein concentrate. Food Chem. 2024, 445, 138671. [Google Scholar] [CrossRef] [PubMed]
- Matysek, J.; Baier, A.; Kalla-Bertholdt, A.-M.; Grebenteuch, S.; Rohn, S.; Rauh, C. Effect of ultrasound and fibre enrichment on aroma profile and texture characteristics of pea protein-based yoghurt alternatives. Innov. Food Sci. Emerg. Technol. 2024, 93, 103610. [Google Scholar] [CrossRef]
- Greulich, O.; Duedahl-Olesen, L.; Mikkelsen, M.S.; Smedsgaard, J.; Bang-Berthelsen, C.H. Fourier Transform Infrared Spectroscopy Tracking of Fermentation of Oat and Pea Bases for Yoghurt-Type Products. Fermentation 2024, 10, 189. [Google Scholar] [CrossRef]
- FIAL. Protein Market: Size of the Prize Analysis for Australia. Available online: https://www.fial.com.au/blogs/post/protein-market-size-of-the-prize-analysis-for-australia (accessed on 13 September 2023).
- van Zanten, H.H.E.; Mollenhorst, H.; Klootwijk, C.W.; van Middelaar, C.E.; de Boer, I.J.M. Global food supply: Land use efficiency of livestock systems. Int. J. Life Cycle Assess. 2016, 21, 747–758. [Google Scholar] [CrossRef]
- van der Spiegel, M.; Noordam, M.Y.; van der Fels-Klerx, H.J. Safety of Novel Protein Sources (Insects, Microalgae, Seaweed, Duckweed, and Rapeseed) and Legislative Aspects for Their Application in Food and Feed Production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef]
- Borlaug, N.E. Feeding a world of 10 billion people: The miracle ahead. In Vitro Cell. Dev. Biol.-Plant 2002, 38, 221–228. [Google Scholar] [CrossRef]
- Boland, M.J.; Rae, A.N.; Vereijken, J.M.; Meuwissen, M.P.M.; Fischer, A.R.H.; van Boekel, M.A.J.S.; Rutherfurd, S.M.; Gruppen, H.; Moughan, P.J.; Hendriks, W.H. The future supply of animal-derived protein for human consumption. Trends Food Sci. Technol. 2013, 29, 62–73. [Google Scholar] [CrossRef]
- Onwezen, M.C.; Bouwman, E.P.; Reinders, M.J.; Dagevos, H. A systematic review on consumer acceptance of alternative proteins: Pulses, algae, insects, plant-based meat alternatives, and cultured meat. Appetite 2021, 159, 105058. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.; de Moura, A.P.; Cunha, L.M. Increasing Pulse Consumption to Improve Human Health and Food Security and to Mitigate Climate Change. In Climate Change-Resilient Agriculture and Agroforestry: Ecosystem Services and Sustainability; Castro, P., Azul, A.M., Leal Filho, W., Azeiteiro, U.M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 21–35. [Google Scholar]
- Chaudhary, A.; Tremorin, D. Nutritional and Environmental Sustainability of Lentil Reformulated Beef Burger. Sustainability 2020, 12, 6712. [Google Scholar] [CrossRef]
- MacWilliam, S.; Parker, D.; Marinangeli, C.P.F.; Trémorin, D. A meta-analysis approach to examining the greenhouse gas implications of including dry peas (Pisum sativum L.) and lentils (Lens culinaris M.) in crop rotations in western Canada. Agric. Syst. 2018, 166, 101–110. [Google Scholar] [CrossRef]
- AGT. History of Pulses. Available online: http://www.agtfoods.com/about-pulses/history-of-pulses.html (accessed on 30 March 2024).
- Balasubramanian, P. Pulse Crops. Available online: https://www.thecanadianencyclopedia.ca/en/article/pulse-crops (accessed on 13 September 2023).
- Pelgrom, P.J.M.; Wang, J.; Boom, R.M.; Schutyser, M.A.I. Pre- and post-treatment enhance the protein enrichment from milling and air classification of legumes. J. Food Eng. 2015, 155, 53–61. [Google Scholar] [CrossRef]
- Fernando, S. Production of protein-rich pulse ingredients through dry fractionation: A review. LWT 2021, 141, 110961. [Google Scholar] [CrossRef]
- Xing, Q.; Utami, D.P.; Demattey, M.B.; Kyriakopoulou, K.; de Wit, M.; Boom, R.M.; Schutyser, M.A.I. A two-step air classification and electrostatic separation process for protein enrichment of starch-containing legumes. Innov. Food Sci. Emerg. Technol. 2020, 66, 102480. [Google Scholar] [CrossRef]
- Li, L.; Yuan, T.Z.; Setia, R.; Raja, R.B.; Zhang, B.; Ai, Y. Characteristics of pea, lentil and faba bean starches isolated from air-classified flours in comparison with commercial starches. Food Chem. 2019, 276, 599–607. [Google Scholar] [CrossRef]
- Allotey, D.K.; Kwofie, E.M.; Adewale, P.; Lam, E.; Ngadi, M. A meta-analysis of pulse-protein extraction technologies: Impact on recovery and purity. J. Food Eng. 2022, 327, 111048. [Google Scholar] [CrossRef]
- Dumoulin, L.; Jacquet, N.; Malumba, P.; Richel, A.; Blecker, C. Dry and wet fractionation of plant proteins: How a hybrid process increases yield and impacts nutritional value of faba beans proteins. Innov. Food Sci. Emerg. Technol. 2021, 72, 102747. [Google Scholar] [CrossRef]
- Schlangen, M.; Taghian Dinani, S.; Schutyser, M.A.I.; van der Goot, A.J. Dry fractionation to produce functional fractions from mung bean, yellow pea and cowpea flour. Innov. Food Sci. Emerg. Technol. 2022, 78, 103018. [Google Scholar] [CrossRef]
- Chang, L.; Lan, Y.; Bandillo, N.; Ohm, J.-B.; Chen, B.; Rao, J. Plant proteins from green pea and chickpea: Extraction, fractionation, structural characterization and functional properties. Food Hydrocoll. 2022, 123, 107165. [Google Scholar] [CrossRef]
- Tanger, C.; Engel, J.; Kulozik, U. Influence of extraction conditions on the conformational alteration of pea protein extracted from pea flour. Food Hydrocoll. 2020, 107, 105949. [Google Scholar] [CrossRef]
- Higa, F.A.; Boyd, L.; Sopiwnyk, E.; Nickerson, M.T. Effect of particle size, flour:water ratio and type of pulse on the physicochemical and functional properties of wet protein extraction. Cereal Chem. 2022, 99, 1049–1062. [Google Scholar] [CrossRef]
- Gao, Z.; Shen, P.; Lan, Y.; Cui, L.; Ohm, J.-B.; Chen, B.; Rao, J. Effect of alkaline extraction pH on structure properties, solubility, and beany flavor of yellow pea protein isolate. Food Res. Int. 2020, 131, 109045. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.G.; Speranza, P.; Kurozawa, L.E.; Kawazoe Sato, A.C. Lentil protein: Impact of different extraction methods on structural and functional properties. Heliyon 2022, 8, e11775. [Google Scholar] [CrossRef] [PubMed]
- Möller, A.C.; Li, J.; van der Goot, A.J.; van der Padt, A. A water-only process to fractionate yellow peas into its constituents. Innov. Food Sci. Emerg. Technol. 2022, 75, 102894. [Google Scholar] [CrossRef]
- Yue, J.; Zhu, Z.; Yi, J.; Lan, Y.; Chen, B.; Rao, J. Structure and functionality of oat protein extracted by choline chloride–dihydric alcohol deep eutectic solvent and its water binary mixtures. Food Hydrocoll. 2021, 112, 106330. [Google Scholar] [CrossRef]
- Bowen, H.; Durrani, R.; Delavault, A.; Durand, E.; Chenyu, J.; Yiyang, L.; Lili, S.; Jian, S.; Weiwei, H.; Fei, G. Application of deep eutectic solvents in protein extraction and purification. Front. Chem. 2022, 10, 912411. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Wang, X.; Liu, W. Application of Deep Eutectic Solvents in Food Analysis: A Review. Molecules 2019, 24, 4594. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.A.; Rinaldi, M.; Menga, V.; Codianni, P.; Giuzio, L.; Fares, C.; Flagella, Z. Influence of Organic and Conventional Farming on Grain Yield and Protein Composition of Chickpea Genotypes. Agronomy 2021, 11, 191. [Google Scholar] [CrossRef]
- Asen, N.D.; Aluko, R.E. Physicochemical and Functional Properties of Membrane-Fractionated Heat-Induced Pea Protein Aggregates. Front. Nutr. 2022, 9, 852225. [Google Scholar] [CrossRef] [PubMed]
- Mahdavian Mehr, H.; Koocheki, A. Effect of atmospheric cold plasma on structure, interfacial and emulsifying properties of Grass pea (Lathyrus sativus L.) protein isolate. Food Hydrocoll. 2020, 106, 105899. [Google Scholar] [CrossRef]
- Basak, S.; Annapure, U.S. Recent trends in the application of cold plasma for the modification of plant proteins—A review. Future Foods 2022, 5, 100119. [Google Scholar] [CrossRef]
- Maria Medeiros Theóphilo Galvão, A.; Lamy Rasera, M.; de Figueiredo Furtado, G.; Grossi Bovi Karatay, G.; Tavares, G.M.; Dupas Hubinger, M. Lentil protein isolate (Lens culinaris) subjected to ultrasound treatment combined or not with heat-treatment: Structural characterization and ability to stabilize high internal phase emulsions. Food Res. Int. 2024, 183, 114212. [Google Scholar] [CrossRef] [PubMed]
- Asen, N.D.; Aluko, R.E. Effect of Heat Treatment on Yellow Field Pea (Pisum sativum) Protein Concentrate Coupled with Membrane Ultrafiltration on Emulsification Properties of the Isolated >50 kDa Proteins. Membranes 2023, 13, 767. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuan, J.-j.; Zhang, Y.-r.; Chen, X.; Wang, J.-l.; Chen, B.; Li, K.; Bai, Y.-h. Unraveling the effect of combined heat and high-pressure homogenization treatment on the improvement of chickpea protein solubility from the perspectives of colloidal state change and structural characteristic modification. Food Chem. 2024, 442, 138470. [Google Scholar] [CrossRef] [PubMed]
- Gallego, M.A.-O.X.; Arnal, M.; Barat, J.M.; Talens, P.A.-O. Effect of Cooking on Protein Digestion and Antioxidant Activity of Different Legume Pastes. Foods 2020, 10, 47. [Google Scholar] [CrossRef]
- Nasrollahzadeh, F.; Alexi, N.; Skov, K.B.; Roman, L.; Sfyra, K.; Martinez, M.M. Texture profiling of muscle meat benchmarks and plant-based analogues: An instrumental and sensory design approach with focus on correlations. Food Hydrocoll. 2024, 151, 109829. [Google Scholar] [CrossRef]
- Hooper, S.D.; Bassett, A.; Wiesinger, J.A.; Glahn, R.P.; Cichy, K.A. Extrusion and drying temperatures enhance sensory profile and iron bioavailability of dry bean pasta. Food Chem. Adv. 2023, 3, 100422. [Google Scholar] [CrossRef]
- Silvestre-De-León, R.; Espinosa-Ramírez, J.; Pérez-Carrillo, E.; Serna-Saldívar, S.O. Extruded chickpea flour sequentially treated with alcalase and α-amylase produces dry instant beverage powders with enhanced yield and nutritional properties. Int. J. Food Sci. Technol. 2021, 56, 5178–5189. [Google Scholar] [CrossRef]
- Santhosh, R.; Babu, D.M.; Thakur, R.; Nath, D.; Hoque, M.; Gaikwad, K.K.; Ahmed, J.; Sarkar, P. Effect of atmospheric cold plasma treatment on structural, thermal, and mechanical properties of pea protein isolate edible films. Sustain. Chem. Pharm. 2024, 37, 101398. [Google Scholar] [CrossRef]
- Rout, S.; Srivastav, P.P. Modification of soy protein isolate and pea protein isolate by high voltage dielectric barrier discharge (DBD) atmospheric cold plasma: Comparative study on structural, rheological and techno-functional characteristics. Food Chem. 2024, 447, 138914. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, G.; Flamminii, F.; Faieta, M.; Prete, R.; Di Michele, A.; Pittia, P.; Di Mattia, C.D. High pressure homogenization to boost the technological functionality of native pea proteins. Curr. Res. Food Sci. 2023, 6, 100499. [Google Scholar] [CrossRef] [PubMed]
- Yaver, E.; Bilgiçli, N. Effect of ultrasound-accelerated debittering method on total alkaloid and total carotenoid content of lupin seeds (Lupinus albus L.) and storage stability of thermally treated lupin flours. J. Food Meas. Charact. 2023, 17, 3378–3389. [Google Scholar] [CrossRef]
- Osemwota, E.C.; Alashi, A.A.-O.; Aluko, R.A.-O. Comparative Study of the Structural and Functional Properties of Membrane-Isolated and Isoelectric pH Precipitated Green Lentil Seed Protein Isolates. Membranes 2021, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Zha, F.; Dong, S.; Rao, J.; Chen, B. The structural modification of pea protein concentrate with gum Arabic by controlled Maillard reaction enhances its functional properties and flavor attributes. Food Hydrocoll. 2019, 92, 30–40. [Google Scholar] [CrossRef]
- Mamilla, R.K.; Mishra, V.K. Effect of germination on antioxidant and ACE inhibitory activities of legumes. LWT 2017, 75, 51–58. [Google Scholar] [CrossRef]
- Karabulut, G.; Yildiz, S.; Karaca, A.C.; Yemiş, O. Ultrasound and enzyme-pretreated extraction for the valorization of pea pod proteins. J. Food Process Eng. 2023, 46, e14452. [Google Scholar] [CrossRef]
- Massmann, C.M.; Berhow, M.; Gibbons, W.R.; Karki, B. The effects of fungal bioprocessing on air-classified pea protein concentrates. LWT 2022, 154, 112686. [Google Scholar] [CrossRef]
- Valtonen, A.; Aisala, H.; Nisov, A.; Nikinmaa, M.; Honkapää, K.; Sozer, N. Synergistic use of fermentation and extrusion processing to design plant protein-based sausages. LWT 2023, 184, 115067. [Google Scholar] [CrossRef]
- Rämö, S.; Kahala, M.; Joutsjoki, V. Aflatoxin B1 Binding by Lactic Acid Bacteria in Protein-Rich Plant Material Fermentation. Appl. Sci. 2022, 12, 12769. [Google Scholar] [CrossRef]
- Ferawati, F.; Zahari, I.; Barman, M.; Hefni, M.; Ahlström, C.; Witthöft, C.; Östbring, K. High-Moisture Meat Analogues Produced from Yellow Pea and Faba Bean Protein Isolates/Concentrate: Effect of Raw Material Composition and Extrusion Parameters on Texture Properties. Foods 2021, 10, 843. [Google Scholar] [CrossRef] [PubMed]
- Blandino, M.; Bresciani, A.; Locatelli, M.; Loscalzo, M.; Travaglia, F.; Vanara, F.; Marti, A. Pulse type and extrusion conditions affect phenolic profile and physical properties of extruded products. Food Chem. 2023, 403, 134369. [Google Scholar] [CrossRef]
- Martin, A.; Schmidt, V.; Osen, R.; Bez, J.; Ortner, E.; Mittermaier, S. Texture, sensory properties and functionality of extruded snacks from pulses and pseudocereal proteins. J. Sci. Food Agric. 2022, 102, 5011–5021. [Google Scholar] [CrossRef] [PubMed]
- Pennells, J.; Bless, I.; Juliano, P.; Ying, D. Extrusion Processing of Biomass By-Products for Sustainable Food Production. In From Biomass to Biobased Products; Jacob-Lopes, E., Zepka, Q.L., Dias, R.R., Eds.; IntechOpen: Rijeka, Croatia, 2023. [Google Scholar]
- Zha, F.; Rao, J.; Chen, B. Modification of pulse proteins for improved functionality and flavor profile: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3036–3060. [Google Scholar] [CrossRef]
- Charoensuk, D.; Brannan, R.G.; Chanasattru, W.; Chaiyasit, W. Physicochemical and emulsifying properties of mung bean protein isolate as influenced by succinylation. Int. J. Food Prop. 2018, 21, 1633–1645. [Google Scholar] [CrossRef]
- Shen, Y.; Hong, S.; Singh, G.; Koppel, K.; Li, Y. Improving functional properties of pea protein through “green” modifications using enzymes and polysaccharides. Food Chem. 2022, 385, 132687. [Google Scholar] [CrossRef]
- Boukid, F.; Ganeshan, S.; Wang, Y.; Tülbek, M.Ç.; Nickerson, M.T. Bioengineered Enzymes and Precision Fermentation in the Food Industry. Int. J. Mol. Sci. 2023, 24, 10156. [Google Scholar] [CrossRef]
- Fang, L.; Xiang, H.; Sun-Waterhouse, D.; Cui, C.; Lin, J. Enhancing the Usability of Pea Protein Isolate in Food Applications through Modifying Its Structural and Sensory Properties via Deamidation by Glutaminase. J. Agric. Food Chem. 2020, 68, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Marengo, M.; Carpen, A.; Bonomi, F.; Casiraghi, M.C.; Meroni, E.; Quaglia, L.; Iametti, S.; Pagani, M.A.; Marti, A. Macromolecular and Micronutrient Profiles of Sprouted Chickpeas to Be Used for Integrating Cereal-Based Food. Cereal Chem. 2017, 94, 82–88. [Google Scholar] [CrossRef]
- Avezum, L.; Rondet, E.; Mestres, C.; Achir, N.; Madode, Y.; Gibert, O.; Lefevre, C.; Hemery, Y.; Verdeil, J.-L.; Rajjou, L. Improving the nutritional quality of pulses via germination. Food Rev. Int. 2022, 39, 6011–6044. [Google Scholar] [CrossRef]
- Atudorei, D.; Stroe, S.-G.; Codină, G.G. Impact of Germination on the Microstructural and Physicochemical Properties of Different Legume Types. Plants 2021, 10, 592. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Song, L.; Feng, S.; Liu, Y.; He, G.; Yioe, Y.; Liu, S.Q.; Huang, D. Germination Dramatically Increases Isoflavonoid Content and Diversity in Chickpea (Cicer arietinum L.) Seeds. J. Agric. Food Chem. 2012, 60, 8606–8615. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of germination on the chemical composition, thermal, pasting, and moisture sorption properties of flours from chickpea, lentil, and yellow pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.; Swanson, G.; Chen, B.; Xu, M. Sensory profile of pulse-based high moisture meat analogs: A study on the complex effect of germination and extrusion processing. Food Chem. 2023, 426, 136585. [Google Scholar] [CrossRef] [PubMed]
- Burlingame, B.; Dernini, S.; Charrondiere, U.; Stadlmayr, B.; Mondovi, S.; Dop, M. Biodiversity and Sustainable Diets; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; Available online: https://hdl.handle.net/10568/104606 (accessed on 27 May 2024).
- Steinkraus, K.H. Lactic acid fermentation in the production of foods from vegetables, cereals and legumes. Antonie Van Leeuwenhoek 1983, 49, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Cui, C.; Ruan, Z. Fermentation-enabled wellness foods: A fresh perspective. Food Sci. Hum. Wellness 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Wuyts, S.; Van Beeck, W.; Allonsius, C.N.; van den Broek, M.F.L.; Lebeer, S. Applications of plant-based fermented foods and their microbes. Curr. Opin. Biotechnol. 2020, 61, 45–52. [Google Scholar] [CrossRef]
- Ahmad, M.I.; Farooq, S.; Alhamoud, Y.; Li, C.; Zhang, H. A review on mycoprotein: History, nutritional composition, production methods, and health benefits. Trends Food Sci. Technol. 2022, 121, 14–29. [Google Scholar] [CrossRef]
- Rastogi, Y.R.; Thakur, R.; Thakur, P.; Mittal, A.; Chakrabarti, S.; Siwal, S.S.; Thakur, V.K.; Saini, R.V.; Saini, A.K. Food fermentation—Significance to public health and sustainability challenges of modern diet and food systems. Int. J. Food Microbiol. 2022, 371, 109666. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Berovič, M.; Krieger, N. Solid-State Fermentation Bioreactor Fundamentals: Introduction and Overview. In Solid-State Fermentation Bioreactors: Fundamentals of Design and Operation; Mitchell, D.A., Berovič, M., Krieger, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–12. [Google Scholar]
- Berovic, M. Cultivation of Medicinal Mushroom Biomass by Solid-State Bioprocessing in Bioreactors. In Solid State Fermentation: Research and Industrial Applications; Steudler, S., Werner, A., Cheng, J.J., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–25. [Google Scholar]
- Verni, M.; Pontonio, E.; Montemurro, M.; Rizzello, C.G. Fermentation as Strategy for Improving Nutritional, Functional, Technological, and Sensory Properties of Legumes. In Legumes Research; Jose, C.J.-L., Alfonso, C., Eds.; IntechOpen: Rijeka, Croatia, 2022; Chapter 13. [Google Scholar]
- Klupsaite, D.; Juodeikiene, G.; Zadeike, D.; Bartkiene, E.; Maknickiene, Z.; Liutkute, G. The influence of lactic acid fermentation on functional properties of narrow-leaved lupine protein as functional additive for higher value wheat bread. LWT 2017, 75, 180–186. [Google Scholar] [CrossRef]
- Kumar, Y.; Basu, S.; Goswami, D.; Devi, M.; Shivhare, U.S.; Vishwakarma, R.K. Anti-nutritional compounds in pulses: Implications and alleviation methods. Legume Sci. 2022, 4, e111. [Google Scholar] [CrossRef]
- Coda, R.; Melama, L.; Rizzello, C.G.; Curiel, J.A.; Sibakov, J.; Holopainen, U.; Pulkkinen, M.; Sozer, N. Effect of air classification and fermentation by Lactobacillus plantarum VTT E-133328 on faba bean (Vicia faba L.) flour nutritional properties. Int. J. Food Microbiol. 2015, 193, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Romero-Espinoza, A.M.; Serna-Saldivar, S.O.; Vintimilla-Alvarez, M.C.; Briones-García, M.; Lazo-Vélez, M.A. Effects of fermentation with probiotics on anti-nutritional factors and proximate composition of lupin (Lupinus mutabilis sweet). LWT 2020, 130, 109658. [Google Scholar] [CrossRef]
- Chandra-Hioe, M.V.; Wong, C.H.M.; Arcot, J. The Potential Use of Fermented Chickpea and Faba Bean Flour as Food Ingredients. Plant Foods Hum. Nutr. 2016, 71, 90–95. [Google Scholar] [CrossRef]
- Curiel, J.A.; Coda, R.; Centomani, I.; Summo, C.; Gobbetti, M.; Rizzello, C.G. Exploitation of the nutritional and functional characteristics of traditional Italian legumes: The potential of sourdough fermentation. Int. J. Food Microbiol. 2015, 196, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Xing, Q.; Dekker, S.; Kyriakopoulou, K.; Boom, R.M.; Smid, E.J.; Schutyser, M.A.I. Enhanced nutritional value of chickpea protein concentrate by dry separation and solid state fermentation. Innov. Food Sci. Emerg. Technol. 2020, 59, 102269. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Torino, M.I.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef]
- Rui, X.; Wen, D.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Enrichment of ACE inhibitory peptides in navy bean (Phaseolus vulgaris) using lactic acid bacteria. Food Funct. 2014, 6, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Emkani, M.; Oliete, B.; Saurel, R. Effect of Lactic Acid Fermentation on Legume Protein Properties, a Review. Fermentation 2022, 8, 244. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Curiel, J.A.; Pinto, D.; Marzani, B.; Coda, R.; Gobbetti, M. Italian legumes: Effect of sourdough fermentation on lunasin-like polypeptides. Microb. Cell Factories 2015, 14, 168. [Google Scholar] [CrossRef]
- Valdez-González, F.; Gutiérrez-Dorado, R.; Hernández-Llamas, A.; García-Ulloa, M.; Sánchez-Magaña, L.; Cuevas-Rodríguez, B.; Rodríguez-González, H. Bioprocessing of common beans in diets for tilapia: In vivo digestibility and antinutritional factors. J. Sci. Food Agric. 2017, 97, 4087–4093. [Google Scholar] [CrossRef]
- Sánchez-García, J.; Asensio Grau, A.; García-Hernández, J.; Heredia, A.; Andrés, A. Nutritional and antioxidant changes in lentils and quinoa through fungal solid-state fermentation with Pleurotus ostreatus. Bioresour. Bioprocess. 2022, 9, 51. [Google Scholar] [CrossRef]
- Olanipekun, B.F.; Otunola, E.T.; Oyelade, O.J. Effect of fermentation on antinutritional factors and in vitro protein digestibility of Bambara nut (Voandzeia subterranean L.). Food Sci. Qual. Manag 2015, 39. Available online: https://api.semanticscholar.org/CorpusID:82228002 (accessed on 27 May 2024).
- Toor, B.S.; Kaur, A.; Kaur, J. Fermentation of legumes with Rhizopus oligosporus: Effect on physicochemical, functional and microstructural properties. Int. J. Food Sci. Technol. 2022, 57, 1763–1772. [Google Scholar] [CrossRef]
- Souza Filho, P.F.; Nair, R.B.; Andersson, D.; Lennartsson, P.R.; Taherzadeh, M.J. Vegan-mycoprotein concentrate from pea-processing industry byproduct using edible filamentous fungi. Fungal Biol. Biotechnol. 2018, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, T.J.A.; Wall, B.T.; Wilde, P.J.; Stephens, F.B.; Taylor, S.L.; Freedman, M.R. Mycoprotein: The Future of Nutritious Nonmeat Protein, a Symposium Review. Curr. Dev. Nutr. 2019, 3, nzz021. [Google Scholar] [CrossRef]
- Clark, A.J.; Soni, B.K.; Sharkey, B.; Acree, T.; Lavin, E.; Bailey, H.M.; Stein, H.H.; Han, A.; Elie, M.; Nadal, M. Shiitake mycelium fermentation improves digestibility, nutritional value, flavor and functionality of plant proteins. LWT 2022, 156, 113065. [Google Scholar] [CrossRef]
- Soni, B.K.; Kelly, B.J.; Langan, J.P.; Davis, H.; Hahn, A.D. Methods for the Production and Use of Myceliated High Protein Food Compositions. U.S. Patent 20170295837A1, 3 July 2018. Available online: https://patents.google.com/patent/US20170295837A1 (accessed on 27 May 2024).
- Sieuwerts, S.; de Bok Frank, A.M.; Hugenholtz, J.; van Hylckama Vlieg Johan, E.T. Unraveling Microbial Interactions in Food Fermentations: From Classical to Genomics Approaches. Appl. Environ. Microbiol. 2008, 74, 4997–5007. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.G.; Rinker, D.C. The genomics of microbial domestication in the fermented food environment. Curr. Opin. Genet. Dev. 2015, 35, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.L.; Klaenhammer, T.R. Genomic Evolution of Domesticated Microorganisms. Annu. Rev. Food Sci. Technol. 2010, 1, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Verstrepen, K.J. Taming Wild Yeast: Potential of Conventional and Nonconventional Yeasts in Industrial Fermentations. Annu. Rev. Microbiol. 2014, 68, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Schindler, S.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Improvement of the Aroma of Pea (Pisum sativum) Protein Extracts by Lactic Acid Fermentation. Food Biotechnol. 2012, 26, 58–74. [Google Scholar] [CrossRef]
- Yousseef, M.; Lafarge, C.; Valentin, D.; Lubbers, S.; Husson, F. Fermentation of cow milk and/or pea milk mixtures by different starter cultures: Physico-chemical and sensorial properties. LWT—Food Sci. Technol. 2016, 69, 430–437. [Google Scholar] [CrossRef]
- Ben-Harb, S.; Saint-Eve, A.; Panouillé, M.; Souchon, I.; Bonnarme, P.; Dugat-Bony, E.; Irlinger, F. Design of microbial consortia for the fermentation of pea-protein-enriched emulsions. Int. J. Food Microbiol. 2019, 293, 124–136. [Google Scholar] [CrossRef] [PubMed]
- El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.-C.; Helinck, S.; Landaud, S. Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts. Foods 2020, 9, 349. [Google Scholar] [CrossRef] [PubMed]
- Ganeshan, S.; Kim, S.H.; Vujanovic, V. Scaling-up production of plant endophytes in bioreactors: Concepts, challenges and perspectives. Bioresour. Bioprocess. 2021, 8, 63. [Google Scholar] [CrossRef]
- Pedrosa, M.M.; Guillamón, E.; Arribas, C. Autoclaved and Extruded Legumes as a Source of Bioactive Phytochemicals: A Review. Foods 2021, 10, 379. [Google Scholar] [CrossRef]
- Kårlund, A.; Gómez-Gallego, C.; Korhonen, J.; Palo-oja, O.-M.; El-Nezami, H.; Kolehmainen, M. Harnessing Microbes for Sustainable Development: Food Fermentation as a Tool for Improving the Nutritional Quality of Alternative Protein Sources. Nutrients 2020, 12, 1020. [Google Scholar] [CrossRef] [PubMed]
- del Cueto, A.G.; Martinez, W.H.; Frampton, V.L. Heat Effects on Peas, Effect of Autoclaving on the Basic Amino Acids and Proteins of the Chick Pea. J. Agric. Food Chem. 1960, 8, 331–332. [Google Scholar] [CrossRef]
- Srihara, P.; Alexander, J.C. Protein Quality of Raw and Autoclaved Plant Protein Blends. Can. Inst. Food Sci. Technol. J. 1983, 16, 63–67. [Google Scholar] [CrossRef]
- Khattab, R.Y.; Arntfield, S.D.; Nyachoti, C.M. Nutritional quality of legume seeds as affected by some physical treatments, Part 1: Protein quality evaluation. LWT—Food Sci. Technol. 2009, 42, 1107–1112. [Google Scholar] [CrossRef]
- Donzella, S.; Fumagalli, A.; Arioli, S.; Pellegrino, L.; D’Incecco, P.; Molinari, F.; Speranza, G.; Ubiali, D.; Robescu, M.S.; Compagno, C. Recycling Food Waste and Saving Water: Optimization of the Fermentation Processes from Cheese Whey Permeate to Yeast Oil. Fermentation 2022, 8, 341. [Google Scholar] [CrossRef]
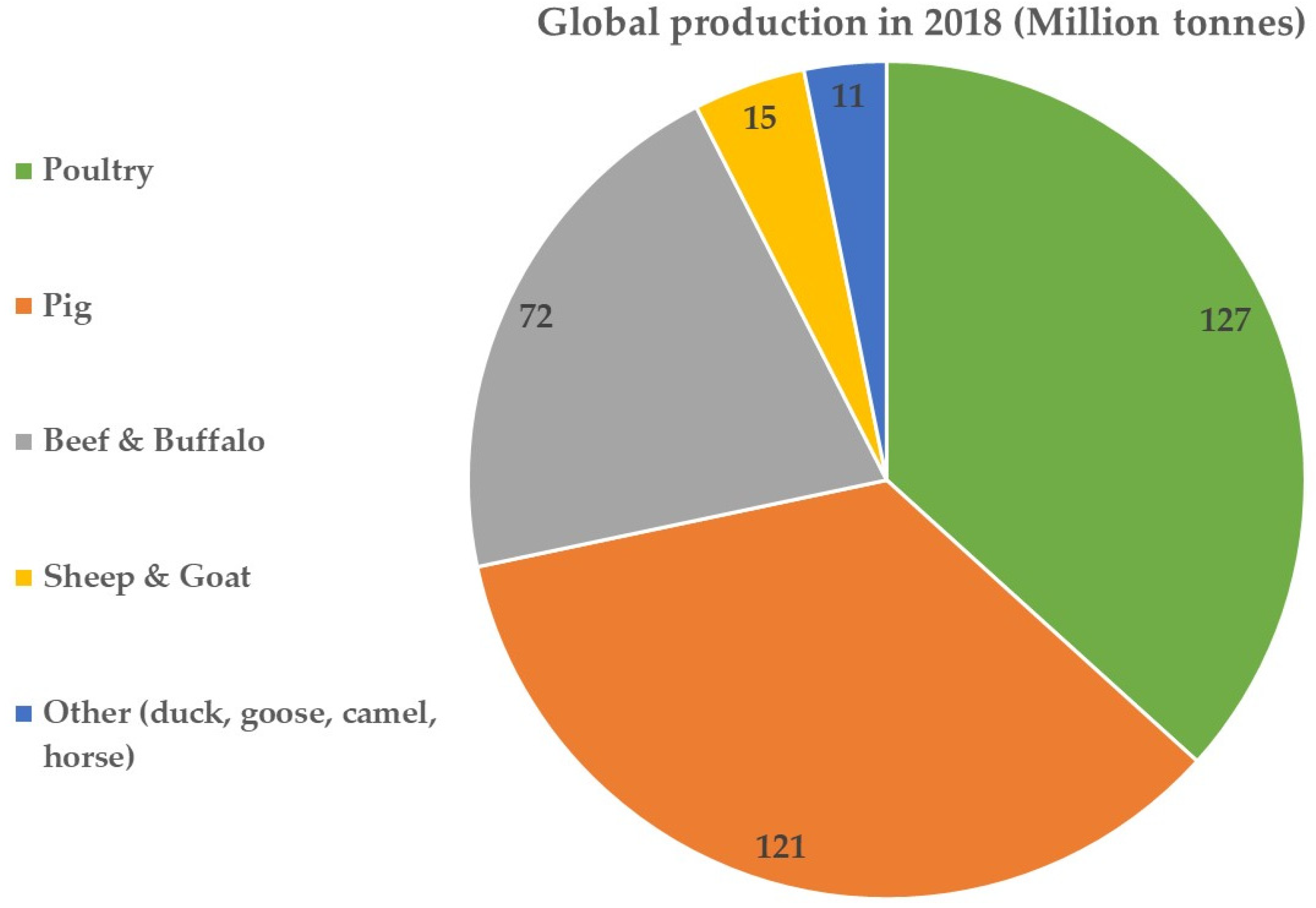
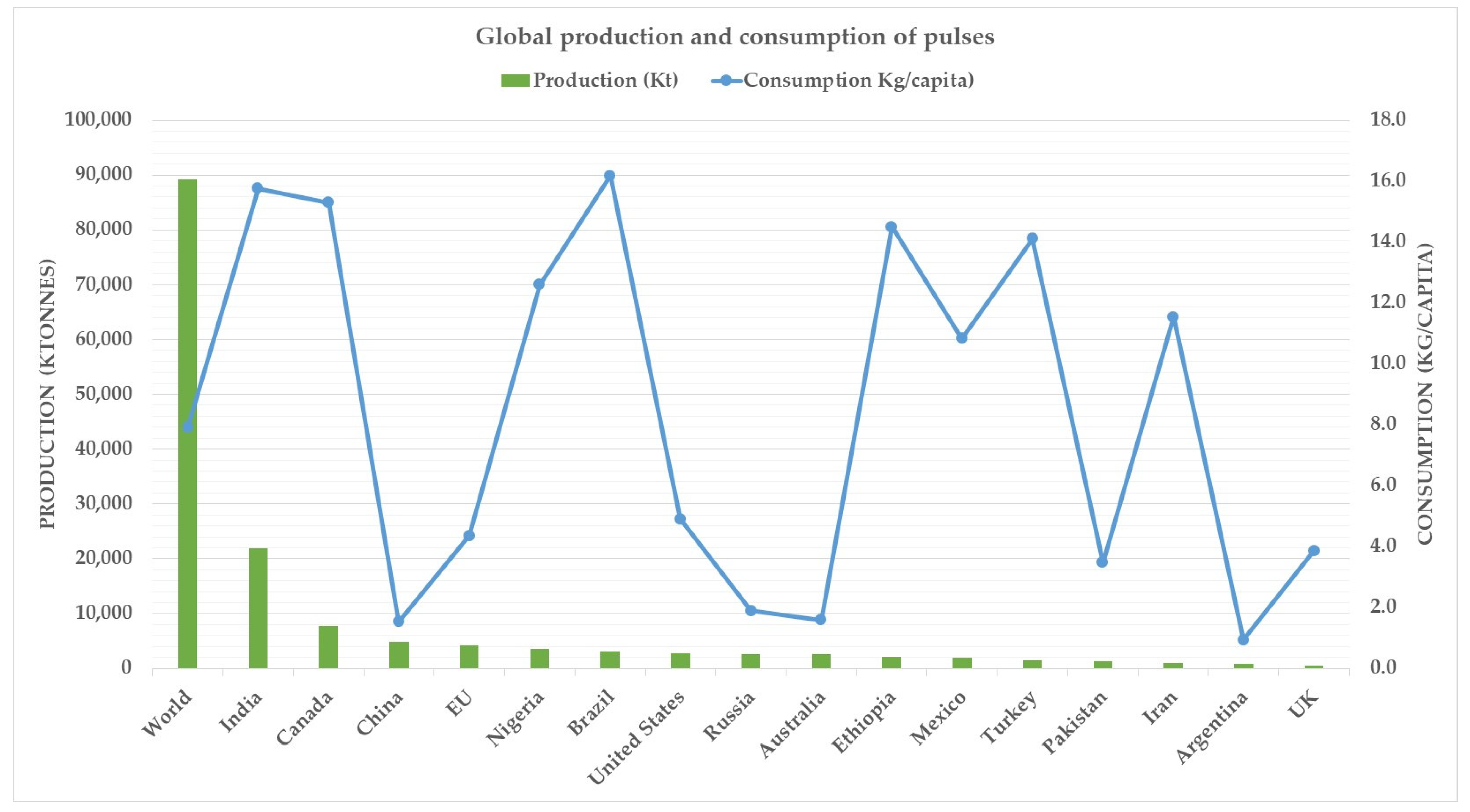
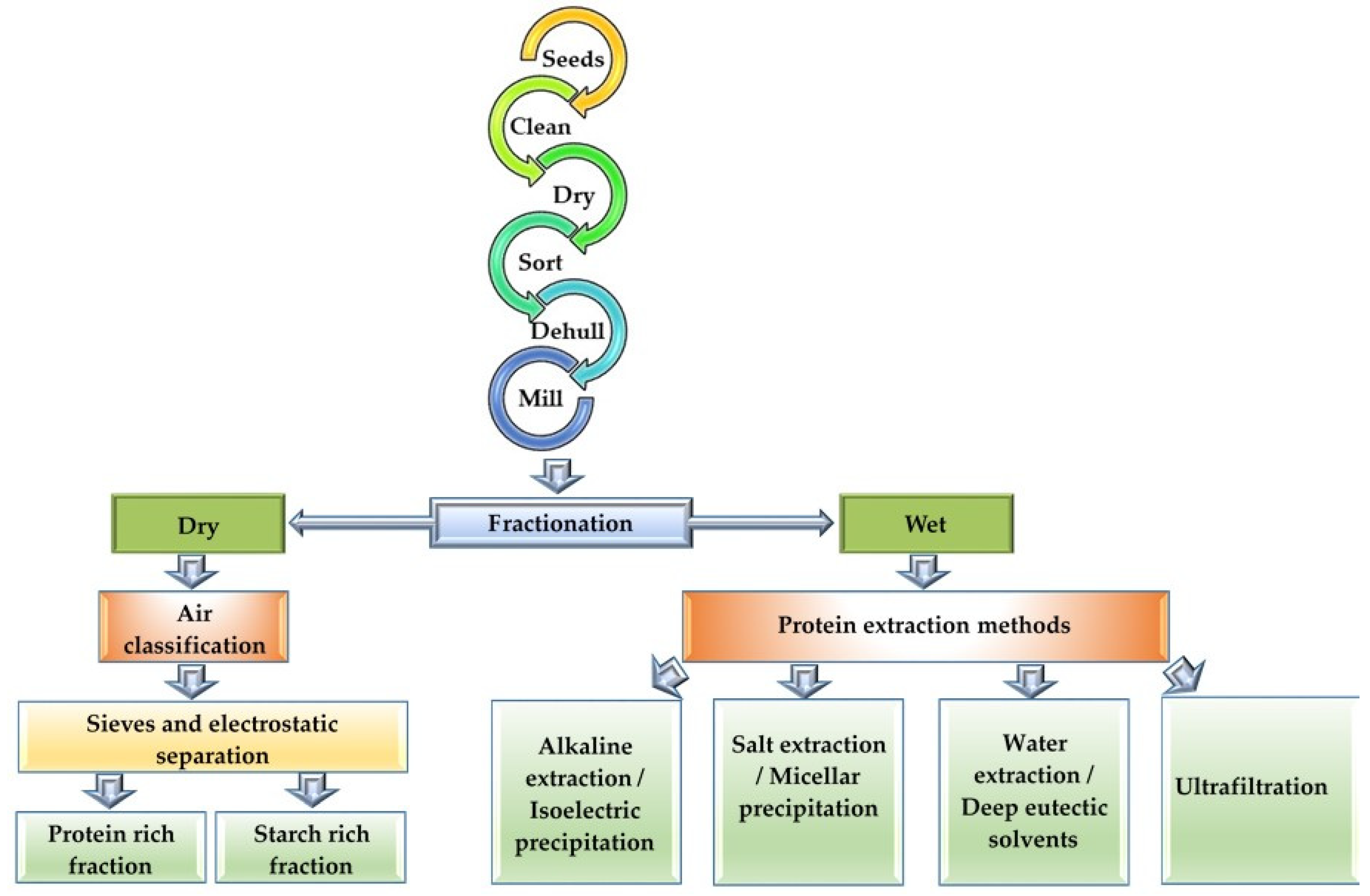
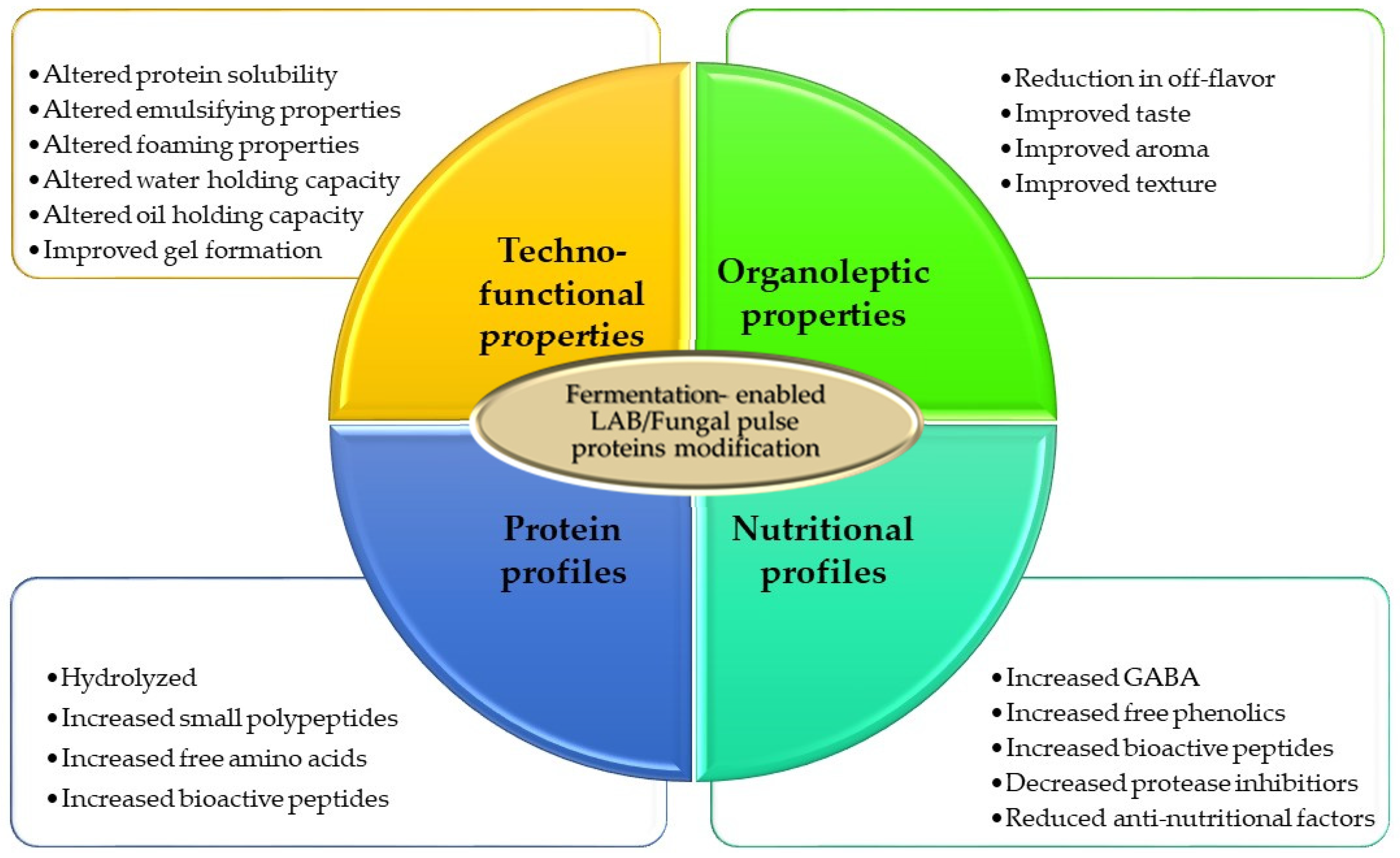
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).