
Evidence of Exposure to Multiple Zoonotic Pathogens in Humans in Lusaka, Zambia: Insights from Metagenomic Next-Generation Sequencing


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Participant Enrollment
2.2. Specimen Collection and Laboratory Testing
2.3. Metagenomic Next-Generation Sequencing
2.3.1. Processing and Analysis of mNGS Data
2.3.2. Quality Control Metrics
2.3.3. Bioinformatic Analysis
2.4. Statistical Analysis
2.5. Ethical Considerations
3. Results
3.1. Demographic Characteristics
3.2. Diversity and Dominance of Microbes
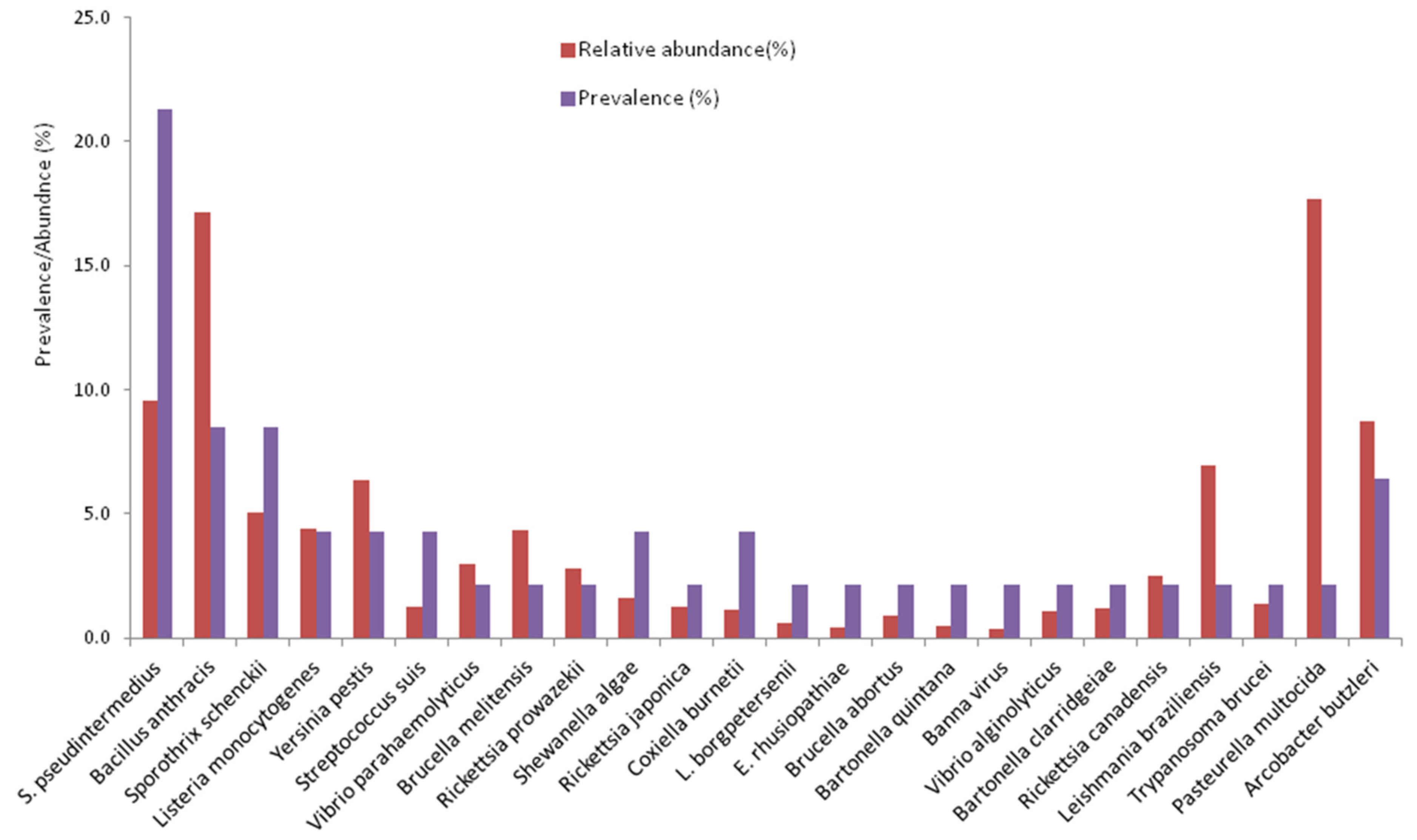
3.3. Prevalence and Abundance of Zoonotic Pathogens
3.4. Risk Factors for Infection with Zoonotic Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tagoe, J.N.A.; Yeboah, C.; Behene, E.; Kumordjie, S.; Nimo-Paintsil, S.; Attram, N.; Nyarko, E.O.; Carroll, J.A.; Fox, A.T.; Watters, C.; et al. Coinfection of Malaria and Bacterial Pathogens among Acute Febrile Patients in Selected Clinics in Ghana. Am. J. Trop. Med. Hyg. 2023, 109, 1036–1046. [Google Scholar] [CrossRef] [PubMed]
- Salyer, S.J.; Silver, R.; Simone, K.; Barton Behravesh, C. Prioritizing Zoonoses for Global Health Capacity Building-Themes from One Health Zoonotic Disease Workshops in 7 Countries, 2014–2016. Emerg. Infect. Dis. 2017, 23, S55–S64. [Google Scholar] [CrossRef]
- Ateudjieu, J.; Siewe Fodjo, J.N.; Ambomatei, C.; Tchio-Nighie, K.H.; Zoung Kanyi Bissek, A.-C. Zoonotic Diseases in Sub-Saharan Africa: A Systematic Review and Meta-Analysis. Zoonotic Dis. 2023, 3, 251–265. [Google Scholar] [CrossRef]
- Allan, K.J.; Biggs, H.M.; Halliday, J.E.B.; Kazwala, R.R.; Maro, V.P.; Cleaveland, S.; Crump, J.A. Epidemiology of Leptospirosis in Africa: A Systematic Review of a Neglected Zoonosis and a Paradigm for ‘One Health’ in Africa. PLOS Neglected Trop. Dis. 2015, 9, e0003899. [Google Scholar] [CrossRef]
- Vanderburg, S.; Rubach, M.P.; Halliday, J.E.B.; Cleaveland, S.; Reddy, E.A.; Crump, J.A. Epidemiology of Coxiella Burnetii Infection in Africa: A OneHealth Systematic Review. PLOS Neglected Trop. Dis. 2014, 8, e2787. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.A.; Nahin, S.; Bonna, A.S.; Kabir Rozars, M.F.; Hossain Hawlader, M.D. Leptospirosis and COVID-19 Co-Infection Case in Bangladesh. Heliyon 2022, 8, e11828. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.J.; Loman, N.; Bogaert, D.; O’Grady, J. Co-Infections: Potentially Lethal and Unexplored in COVID-19. Lancet Microbe 2020, 1, e11. [Google Scholar] [CrossRef]
- Baral, B.; Saini, V.; Kandpal, M.; Kundu, P.; Dixit, A.K.; Parmar, H.S.; Meena, A.K.; Trivedi, P.; Jha, H.C. The Interplay of Co-Infections in Shaping COVID-19 Severity: Expanding the Scope beyond SARS-CoV-2. J. Infect. Public Health 2024, 17, 102486. [Google Scholar] [CrossRef]
- Bohl, J.A.; Lay, S.; Chea, S.; Ahyong, V.; Parker, D.M.; Gallagher, S.; Fintzi, J.; Man, S.; Ponce, A.; Sreng, S.; et al. Discovering Disease-Causing Pathogens in Resource-Scarce Southeast Asia Using a Global Metagenomic Pathogen Monitoring System. Proc. Natl. Acad. Sci. USA 2022, 119, e2115285119. [Google Scholar] [CrossRef]
- Kelly-Cirino, C.D.; Nkengasong, J.; Kettler, H.; Tongio, I.; Gay-Andrieu, F.; Escadafal, C.; Piot, P.; Peeling, R.W.; Gadde, R.; Boehme, C. Importance of Diagnostics in Epidemic and Pandemic Preparedness. BMJ Glob. Health 2019, 4, e001179. [Google Scholar] [CrossRef]
- Chidzwondo, F.; Mutapi, F. Challenge of Diagnosing Acute Infections in Poor Resource Settings in Africa. AAS Open Res. 2024, 4, 28. [Google Scholar]
- Moonga, L.C.; Hayashida, K.; Nakao, R.; Lisulo, M.; Kaneko, C.; Nakamura, I.; Eshita, Y.; Mweene, A.S.; Namangala, B.; Sugimoto, C.; et al. Molecular Detection of Rickettsia Felis in Dogs, Rodents and Cat Fleas in Zambia. Parasites Vectors 2019, 12, 168. [Google Scholar] [CrossRef] [PubMed]
- Mubemba, B.; Mburu, M.M.; Changula, K.; Muleya, W.; Moonga, L.C.; Chambaro, H.M.; Kajihara, M.; Qiu, Y.; Orba, Y.; Hayashida, K.; et al. Current Knowledge of Vector-Borne Zoonotic Pathogens in Zambia: A Clarion Call to Scaling-up “One Health” Research in the Wake of Emerging and Re-Emerging Infectious Diseases. PLOS Neglected Trop. Dis. 2022, 16, e0010193. [Google Scholar] [CrossRef]
- Munjita, S.M.; Kajihara, M.; Mubemba, B.; Kalonda, A.; Tembo, J.; Chitanga, S.; Changula, K.; Tato, C.; Vanaerschot, M.; Munsaka, S.; et al. Evidence of Multiple Bacterial, Viral and Parasitic Infectious Disease Agents of Public Health Importance in Mastomys Natalensis Rodents in Riverine Areas in Selected Parts of Zambia. Ecol. Epidemiol. 2024, 15. [Google Scholar] [CrossRef]
- Munjita, S.M.; Mubemba, B.; Changula, K.; Tembo, J.; Hamoonga, R.; Bates, M.; Chitanga, S.; Munsaka, S.; Simulundu, E. Unveiling the Hidden Threats: A Review of Pathogen Diversity and Public Health Risks from Bats, Rodents, and Non-Human Primates in Zambia (1990–2022). Front. Public Health 2024, 12, 1471452. [Google Scholar] [CrossRef]
- Tembo, J.; Egbe, N.F.; Maluzi, K.; Mulonga, K.; Chilufya, M.; Kapata, N.; Mukonka, V.; Simulundu, E.; Zumla, A.; Fwoloshi, S.; et al. Evaluation of SARS-CoV-2 Diagnostics and Risk Factors Associated with SARS-CoV-2 Infection in Zambia. Int. J. Infect. Dis. 2022, 120, 150–157. [Google Scholar] [CrossRef] [PubMed]
- In, J. Introduction of a Pilot Study. Korean J. Anesthesiol. 2017, 70, 601. [Google Scholar] [CrossRef]
- Munjita, S.M.; Mubemba, B.; Tembo, J.; Bates, M.; Munsaka, S. Rhipicephalus Simus Ticks: New Hosts for Phleboviruses. Parasitology 2024, 151, 962–970. [Google Scholar] [CrossRef]
- Munjita, S.M.; Moonga, G.; Mukubesa, A.N.; Ndebe, J.; Mubemba, B.; Vanaerschot, M.; Tato, C.; Tembo, J.; Kapata, N.; Chitanga, S.; et al. Luna Virus and Helminths in Wild Mastomys Natalensis in Two Contrasting Habitats in Zambia: Risk Factors and Evidence of Virus Dissemination in Semen. Pathogens 2022, 11, 1345. [Google Scholar] [CrossRef]
- Kalantar, K.L.; Carvalho, T.; de Bourcy, C.F.A.; Dimitrov, B.; Dingle, G.; Egger, R.; Han, J.; Holmes, O.B.; Juan, Y.-F.; King, R.; et al. IDseq—An Open Source Cloud-Based Pipeline and Analysis Service for Metagenomic Pathogen Detection and Monitoring. GigaScience 2020, 9, giaa111. [Google Scholar] [CrossRef]
- Munjita, S.M.; Abu, Y.E.; Mubemba, B. Total Microbiome of an African Filarial and Arbovirus Vector, Culex Quinquefasciatus: Insights into Composition and Prevalence of Human Pathogenic Microbes. J. Eur. Mosq. Control Assoc. 2025, 43, 1–9. [Google Scholar] [CrossRef]
- Young, T. Biodiversity Calculator for the Simpson and Shannon Indexes. Available online: https://www.alyoung.com/labs/biodiversity_calculator.html?srsltid=AfmBOor5T24oblgIGK-DVDa-O9GtqJ0uwfJshn3PtPU365GNfBdUAwmM (accessed on 14 December 2024).
- Caruso, T.; Pigino, G.; Bernini, F.; Bargagli, R.; Migliorini, M. The Berger–Parker Index as an Effective Tool for Monitoring the Biodiversity of Disturbed Soils: A Case Study on Mediterranean Oribatid (Acari: Oribatida) Assemblages. Biodivers. Conserv. Eur. 2008, 35–43. [Google Scholar]
- Beisel, J.-N.; Thomas, S.; Usseglio-Polatera, P.; Moreteau, J.-C. Assessing Changes in Community Structure by Dominance Indices: A Comparative Analysis. J. Freshw. Ecol. 1996, 11, 291–299. [Google Scholar] [CrossRef]
- Nyirenda, S.S.; Hang’ombe, B.M.; Mulenga, E.; Kilonzo, B.S. Serological and PCR Investigation of Yersinia Pestis in Potential Reservoir Hosts from a Plague Outbreak Focus in Zambia. BMC Res. Notes 2017, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Nakao, R.; Hang’ombe, B.M.; Sato, K.; Kajihara, M.; Kanchela, S.; Changula, K.; Eto, Y.; Ndebe, J.; Sasaki, M.; et al. Human Borreliosis Caused by a New World Relapsing Fever Borrelia–like Organism in the Old World. Clin. Infect. Dis. 2019, 69, 107–112. [Google Scholar] [CrossRef]
- Moonga, L.C.; Hayashida, K.; Mulunda, N.R.; Nakamura, Y.; Chipeta, J.; Moonga, H.B.; Namangala, B.; Sugimoto, C.; Mtonga, Z.; Mutengo, M.; et al. Molecular Detection and Characterization of Rickettsia Asembonensis in Human Blood, Zambia. Emerg. Infect. Dis. 2021, 27, 2237–2239. [Google Scholar] [CrossRef]
- Chitanga, S.; Simulundu, E.; Simuunza, M.C.; Changula, K.; Qiu, Y.; Kajihara, M.; Nakao, R.; Syakalima, M.; Takada, A.; Mweene, A.S.; et al. First Molecular Detection and Genetic Characterization of Coxiella Burnetii in Zambian Dogs and Rodents. Parasites Vectors 2018, 11, 40. [Google Scholar] [CrossRef]
- Simulundu, E.; Mweene, A.S.; Changula, K.; Monze, M.; Chizema, E.; Mwaba, P.; Takada, A.; Ippolito, G.; Kasolo, F.; Zumla, A.; et al. Lujo Viral Hemorrhagic Fever: Considering Diagnostic Capacity and Preparedness in the Wake of Recent Ebola and Zika Virus Outbreaks. Rev. Med. Virol. 2016, 26, 446–454. [Google Scholar] [CrossRef]
- National Center for Emerging and Zoonotic Infectious Diseases Respiratory Infections | CDC Yellow Book 2024. Available online: https://wwwnc.cdc.gov/travel/yellowbook/2024/posttravel-evaluation/respiratory-infections (accessed on 11 January 2025).
- Chieosilapatham, P.; Chuamanochan, M.; Chiewchavit, S.; Saikruatep, R.; Amornrungsun, E.; Preechasuth, K. Sporothrix Schenckii Sensu Stricto Related to Zoonotic Transmission in Thailand. Med. Mycol. Case Rep. 2023, 41, 44–47. [Google Scholar] [CrossRef]
- World Health Organisation Anthrax-Zambia. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2023-DON497 (accessed on 11 January 2025).
- Oberbeckmann, S.; Wichels, A.; Wiltshire, K.H.; Gerdts, G. Occurrence of Vibrio Parahaemolyticus and Vibrio Alginolyticus in the German Bight over a Seasonal Cycle. Antonie Van Leeuwenhoek 2011, 100, 291–307. [Google Scholar] [CrossRef]
- Nabeshima, T.; Nga, P.T.; Guillermo, P.; del Parquet, M.C.; Yu, F.; Thuy, N.T.; Trang, B.M.; Hien, N.T.; Nam, V.S.; Inoue, S.; et al. Isolation and Molecular Characterization of Banna Virus from Mosquitoes, Vietnam. Emerg. Infect. Dis 2008, 14, 1276–1279. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.J.; Quan, V.; Frean, J.; Knobel, D.L.; Rossouw, J.; Weyer, J.; Marcotty, T.; Godfroid, J.; Blumberg, L.H. Prevalence of Selected Zoonotic Diseases and Risk Factors at a Human-Wildlife-Livestock Interface in Mpumalanga Province, South Africa. Vector-Borne Zoonotic Dis. 2018, 18, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Koizumi, N.; Ohnuma, A.; Mutemwa, A.; Hang’ombe, B.M.; Mweene, A.S.; Takada, A.; Sugimoto, C.; Suzuki, Y.; Kida, H.; et al. Molecular Epidemiology of Pathogenic Leptospira Spp. in the Straw-Colored Fruit Bat (Eidolon Helvum) Migrating to Zambia from the Democratic Republic of Congo. Infect. Genet. Evol. 2015, 32, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Hang’ombe, B.M.; Sawa, H.; Kobayashi, S.; Orba, Y.; Ishii, A.; Thomas, Y.; Isozumi, R.; Yoshimatsu, K.; Mweene, A.S.; et al. Cross-Reactivity of Secondary Antibodies against African Rodents and Application for Sero-Surveillance. J. Vet. Med. Sci. 2013, 75, 819–825. [Google Scholar] [CrossRef]
- Masunga, D.S.; Rai, A.; Abbass, M.; Uwishema, O.; Wellington, J.; Uweis, L.; El Saleh, R.; Arab, S.; Onyeaka, C.V.P.; Onyeaka, H. Leptospirosis Outbreak in Tanzania: An Alarming Situation. Ann. Med. Surg. 2022, 80, 104347. [Google Scholar] [CrossRef]
- Squarre, D.; Chambaro, H.M.; Hayashida, K.; Moonga, L.C.; Qiu, Y.; Goto, Y.; Oparaocha, E.; Mumba, C.; Muleya, W.; Bwalya, P.; et al. Autochthonous Leishmania Infantum in Dogs, Zambia, 2021. Emerg. Infect. Dis. 2022, 28, 888–890. [Google Scholar] [CrossRef]
- Prouzet-Mauléon, V.; Labadi, L.; Bouges, N.; Ménard, A.; Mégraud, F. Arcobacter Butzleri: Underestimated Enteropathogen. Emerg. Infect. Dis. 2006, 12, 307–309. [Google Scholar] [CrossRef]
- Mpundu, P.; Muma, J.B.; Mukubesa, A.N.; Kainga, H.; Mudenda, S.; Bumbangi, F.N.; Muleya, W.; Katemangwe, P.; Munyeme, M. Antibiotic Resistance Patterns of Listeria Species Isolated from Broiler Abattoirs in Lusaka, Zambia. Antibiotics 2022, 11, 591. [Google Scholar] [CrossRef] [PubMed]
- Mpundu, P.; Muma, J.B.; Mukumbuta, N.; Mukubesa, A.N.; Muleya, W.; Kapila, P.; Hang’ombe, B.M.; Munyeme, M. Isolation, Discrimination, and Molecular Detection of Listeria Species from Slaughtered Cattle in Namwala District, Zambia. BMC Microbiol. 2022, 22, 160. [Google Scholar] [CrossRef]
- Munjita, S.M.; Chileshe, M.; Mutemwa, S. Ebola Virus Disease in West Africa: A Call to Overhaul Health Systems in Sub-Saharan Africa. Int. J. Med. Sci. Public Health 2015, 4, 873–877. [Google Scholar] [CrossRef]
 .
.
.
.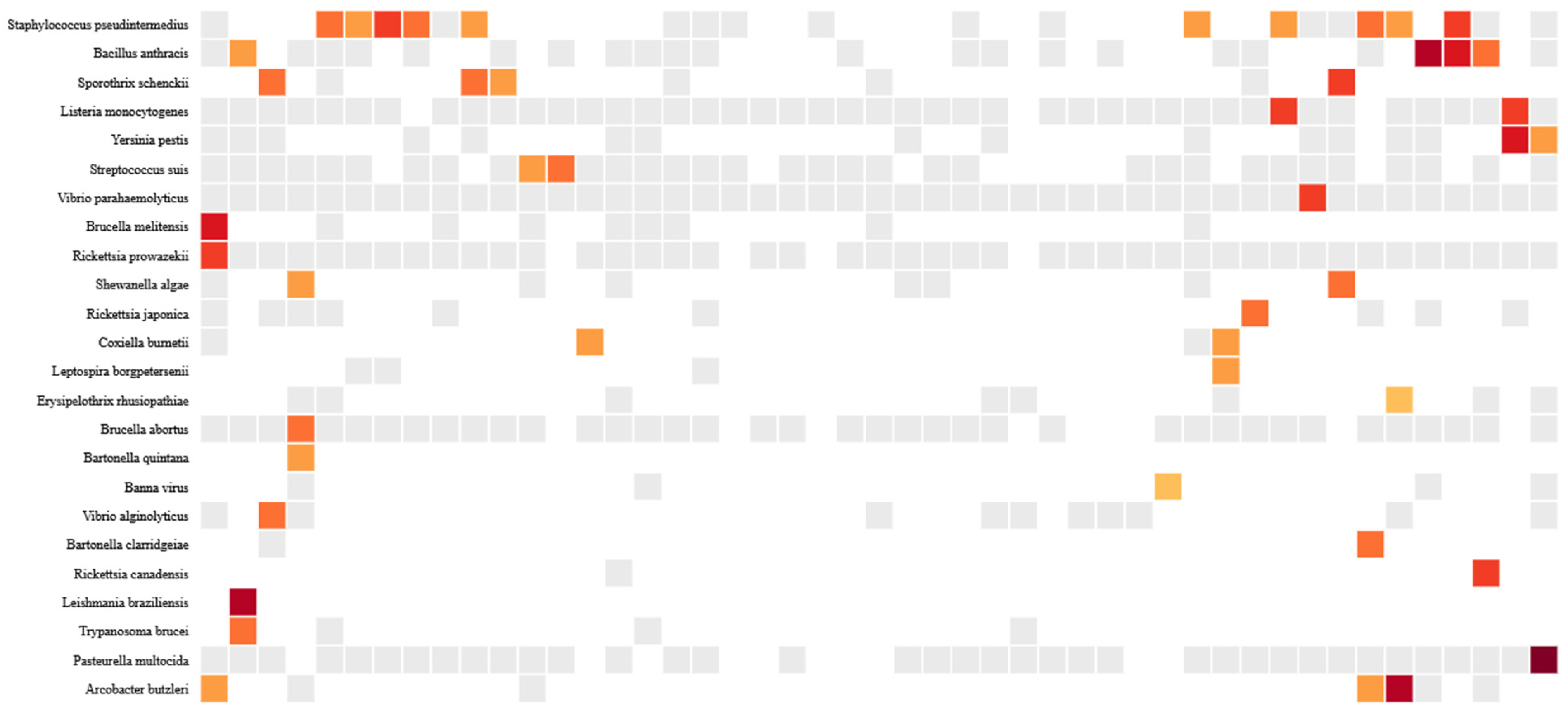

{kind=link}
{kind=link}
| Variable | Chi-Square (χ2) Test | Multivariate Logistic Regression | |||
|---|---|---|---|---|---|
| p-Value | Level | Odds Ratio | 95% CI * | p-Value | |
| Gender | 0.502 | Male | 0.951 | 0.280–3.227 | 0.936 |
| Female | Ref | ||||
| Age | 0.306 | Adult | 1.191 | 0.300–4.723 | 0.804 |
| Youth | Ref | ||||
| Comorbidities | 0.434 | Present | 2.667 | 0.441–16.147 | 0.286 |
| Absent | Ref | ||||
| HIV | 0.242 | Positive | 0.179 | 0.020–1.593 | 0.179 |
| Negative | Ref | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munjita, S.M.; Tembo, J.; Muleya, W.; Bates, M. Evidence of Exposure to Multiple Zoonotic Pathogens in Humans in Lusaka, Zambia: Insights from Metagenomic Next-Generation Sequencing. Zoonotic Dis. 2025, 5, 13. https://doi.org/10.3390/zoonoticdis5020013
Munjita SM, Tembo J, Muleya W, Bates M. Evidence of Exposure to Multiple Zoonotic Pathogens in Humans in Lusaka, Zambia: Insights from Metagenomic Next-Generation Sequencing. Zoonotic Diseases. 2025; 5(2):13. https://doi.org/10.3390/zoonoticdis5020013
Chicago/Turabian StyleMunjita, Samuel Munalula, John Tembo, Walter Muleya, and Matthew Bates. 2025. "Evidence of Exposure to Multiple Zoonotic Pathogens in Humans in Lusaka, Zambia: Insights from Metagenomic Next-Generation Sequencing" Zoonotic Diseases 5, no. 2: 13. https://doi.org/10.3390/zoonoticdis5020013
APA StyleMunjita, S. M., Tembo, J., Muleya, W., & Bates, M. (2025). Evidence of Exposure to Multiple Zoonotic Pathogens in Humans in Lusaka, Zambia: Insights from Metagenomic Next-Generation Sequencing. Zoonotic Diseases, 5(2), 13. https://doi.org/10.3390/zoonoticdis5020013