
Altitudinal Genetic Variation of Pinus oocarpa Seedling Emergence in the Southern Mountains, Oaxaca, Mexico


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Cleaning of Seeds
2.2. Sowing and Experimental Design
2.3. Emergence Parameters Evaluated
2.4. Climatic Variables
2.5. Data Analysis
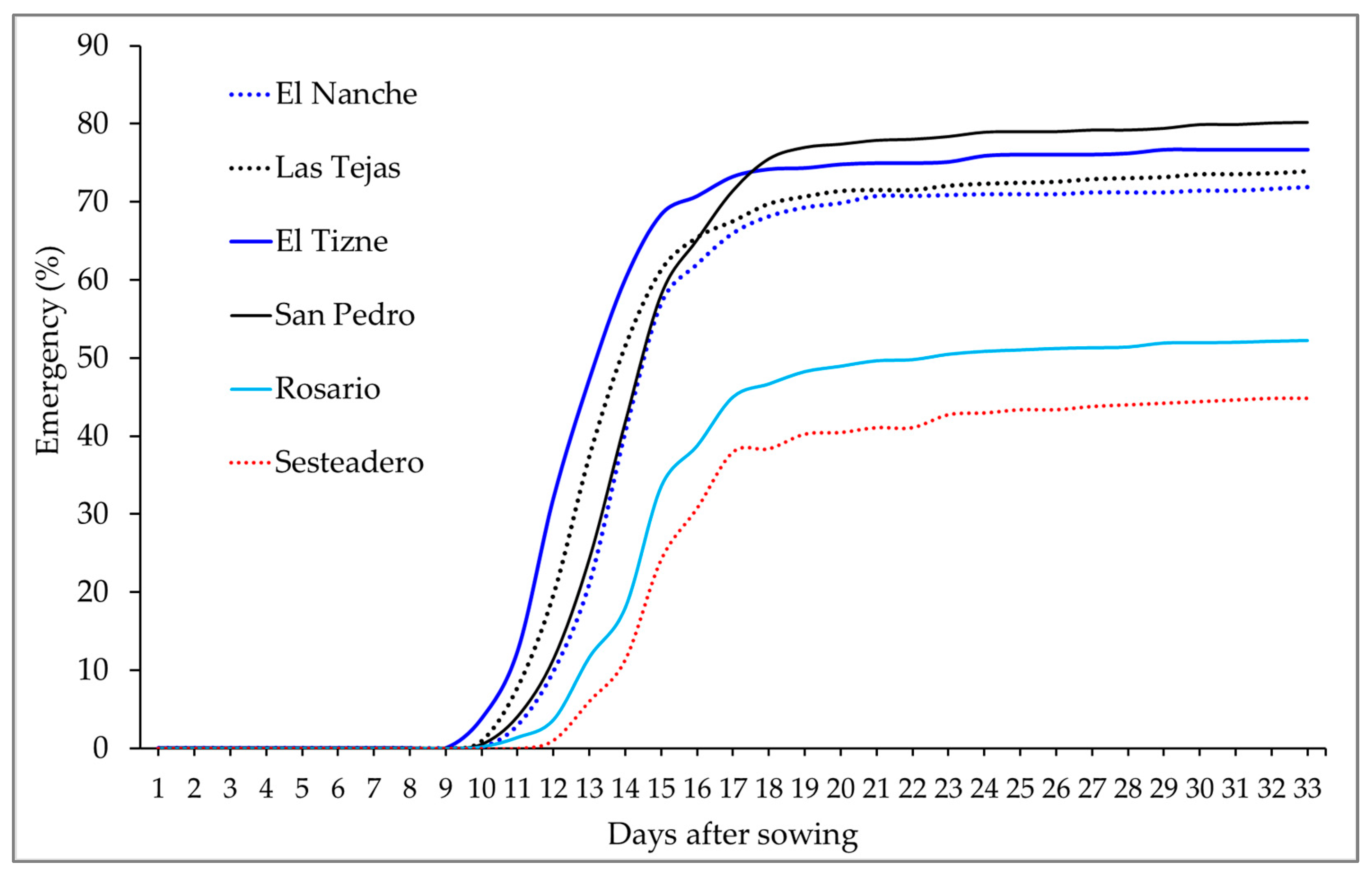
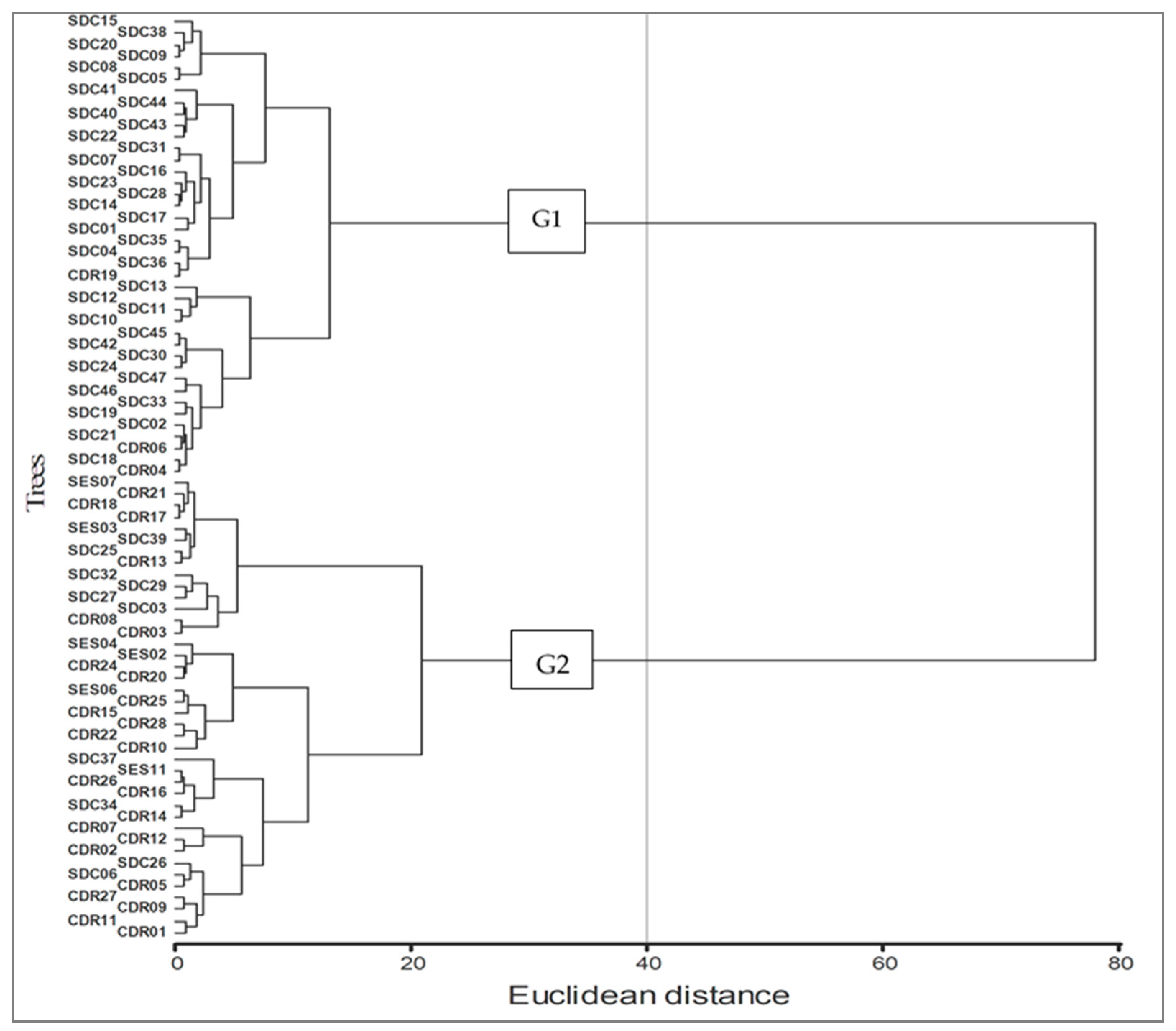
3. Results
3.1. Variation between Provenances and between Trees
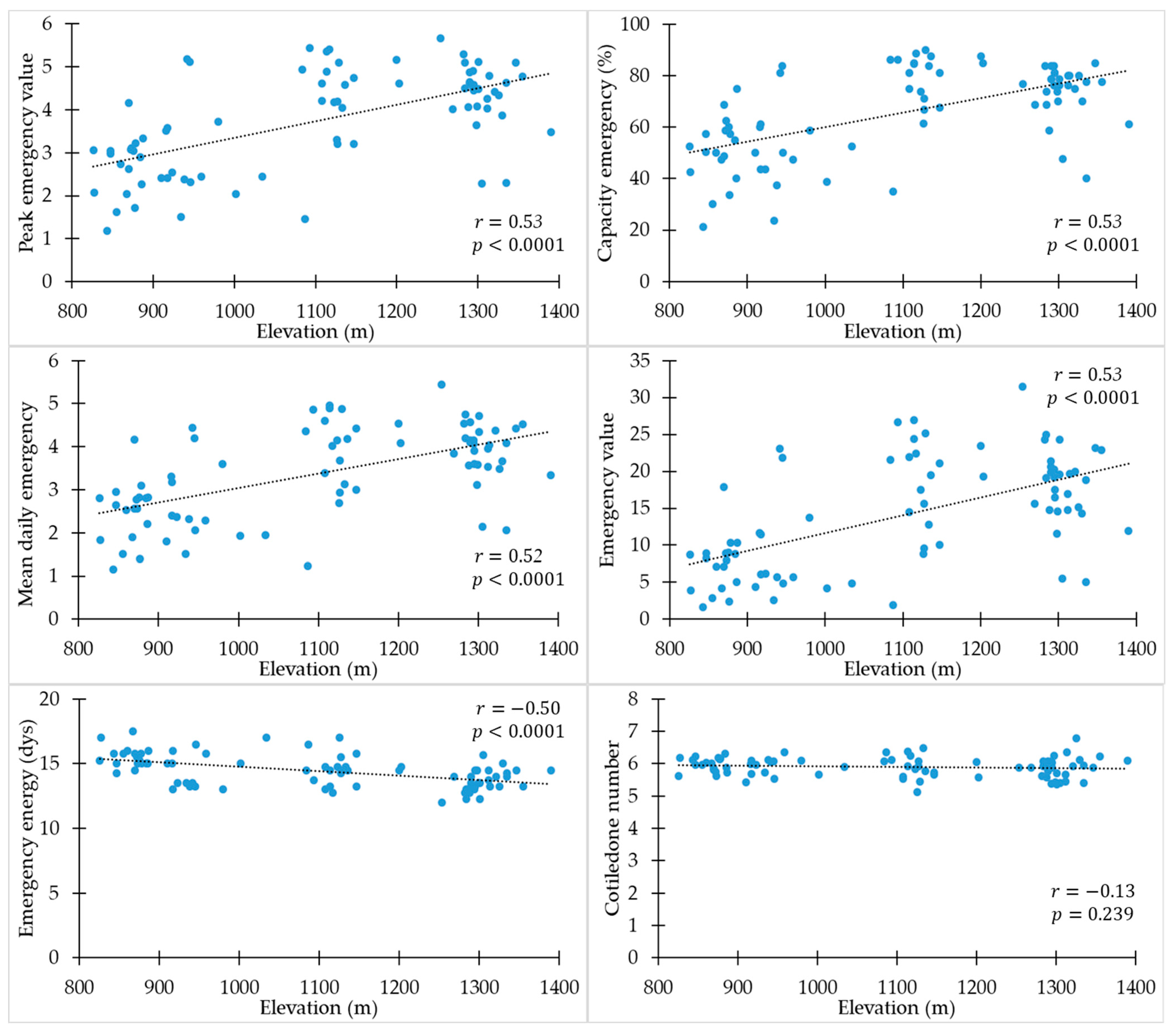
3.2. Association between Emergence and Cotyledons and Elevation and Climatic Variables
4. Discussion
4.1. Variation between Provenances and between Trees
4.2. Association between Emergence and Cotyledons and Elevation and Climatic Variables
4.3. Practical Applications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farjon, A.; Styles, B.T. Pinus (Pinaceae). Flora Neotrop. 1997, 75, 1–291. Available online: http://www.jstor.org/stable/4393881 (accessed on 31 October 2023).
- Schwarz, O.J.; Beaty, R.M.; Franco, E.O. Egg-Cone Pine (Pinus oocarpa Schiede). In Trees III. Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1991; Volume 16, pp. 304–316. [Google Scholar] [CrossRef]
- Leyva-Ovalle, A.; Velázquez-Martínez, A.; Aldrete, A.; Gómez-Guerrero, A.; Medina-Hernández, J.A. La Producción de Resina de Pino en México; CONAFOR: Guadalajara, Mexico, 2013; pp. 20–82. [Google Scholar]
- Reyes-Ramos, A.; Cruz de León, J.; Martínez-Palacios, A.; Lobit, P.C.M.; Ambríz-Parra, J.E.; Sánchez-Vargas, N.M. Ecological and dendrometric characters in which influence resin production of Pinus oocarpa of Michoacán, Mexico. Madera Bosques 2019, 25, e2511414. [Google Scholar] [CrossRef]
- Sarria-Villa, R.A.; Gallo-Corredor, J.A.; Benítez-Benítez, R. Characterization and determination of the quality of rosins and turpentines extracted from Pinus oocarpa and Pinus patula resin. Heliyon 2021, 7, e07834. [Google Scholar] [CrossRef]
- Fabián-Plesníková, I.; Sáenz-Romero, C.; de León, J.C.; Martínez-Trujillo, M.; Sánchez-Vargas, N.M. Growth trait genetic parameters in a progeny trial of Pinus oocarpa. Madera Bosques 2020, 26, e2632014. [Google Scholar] [CrossRef]
- Fabián-Plesníková, I.; Sáenz-Romero, C.; Cruz-De-León, J.; Martínez-Trujillo, M.; Sánchez-Vargas, N.M.; Terrazas, T. Heritability and characteristics of resin ducts in Pinus oocarpa stems in Michoacán, Mexico. IAWA J. 2021, 42, 258–278. [Google Scholar] [CrossRef]
- Muñoz-Flores, H.J.; Sáenz Reyes, J.T.; Gómez Cárdenas, M.; Hernández-Ramos, J.; Barrera, R. Variación morfológica en semillas de Pinus pseudostrobus Lindl. altamente productores de resina. Acta Univ. 2023, 33, e3549. [Google Scholar] [CrossRef]
- Romero-Sanchez, M.E.; Velasco-Garcia, M.V.; Perez-Miranda, R.; Velasco-Bautista, E.; Gonzalez-Hernandez, A. Different modelling approaches to determine suitable areas for conserving egg-cone pine (Pinus oocarpa Schiede) plus trees in the central part of Mexico. Forests 2022, 13, 2112. [Google Scholar] [CrossRef]
- Zobel, B.; Talbert, J. Applied Forest Tree Improvement; John Wiley & Sons: New York, NY, USA, 1984; p. 505. [Google Scholar]
- White, T.L.; Adams, W.T.; Neale, D.B. Forest Genetics; CABI Publishing: Cambrige, MA, USA, 2007; p. 682. [Google Scholar]
- Hernández-Anguiano, L.A.; López-Upton, J.; Ramírez-Herrera, C.; Romero-Manzanares, A. Variation in germination and vigor of Pinus cembroides and Pinus orizabensis Seeds. Agrociencia 2018, 52, 1161–1178. Available online: https://www.scielo.org.mx/pdf/agro/v52n8/1405-3195-agro-52-08-1161-en.pdf (accessed on 2 November 2023).
- Fuentes-Amaro, S.L.; Rodríguez-Laguna, R.; Razo-Zárate, R.; Meza-Rangel, J.; Jiménez-Casas, M. Altitudinal variation in emergence and initial growth of Pinus patula plants. Rev. Fitotec. Mex. 2021, 44, 655–660. Available online: https://revistafitotecniamexicana.org/documentos/44-4/19a.pdf (accessed on 30 October 2023).
- Gutiérrez-Vázquez, B.N.; Flores-Montaño, A. Patrón y magnitud de la variación de la densidad de la madera en rodales semilleros de Pinus oocarpa. Madera Bosques 2019, 25, e2531615. [Google Scholar] [CrossRef]
- Viveros-Viveros, H.; Sáenz-Romero, C.; Guzmán-Reyna, R.R. Pattern and magnitude of the variation of the wood density in seed production stands of Pinus oocarpa. Rev. Fitotec. Mex. 2005, 28, 333–338. [Google Scholar] [CrossRef]
- Xu, Y.; Cai, N.; He, B.; Zhang, R.; Zhao, W.; Mao, J.; Duan, A.; Li, Y.; Woeste, K. Germination and early seedling growth of Pinus densata Mast. provenances. J. For. Res. 2016, 27, 283–294. [Google Scholar] [CrossRef]
- López-López, A.; Palacios-Rangel, M.I.; Sáenz-Romero, C.; Villanueva-Morales, A.; Pacheco-Almaraz, V. Variación clinal de caracteres fenotípicos y fisiológicos en Pinus hartwegii Lindl., para la estación forestal experimental zoquiapan, México. Polibotánica 2023, 56, 62–79. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Viveros-Viveros, H.; Guzmán-Reyna, R.R. Altitudinal genetic variation among Pinus oocarpa populations on Michoacán, Western México. Preliminary Results from a Nursery Test. For. Genet. 2004, 11, 343–349. Available online: https://www.uv.mx/personal/heviveros/files/2014/11/SaenzRomero_et-al_2004_PoocarpaVivero_ForestGenetics.pdf (accessed on 27 October 2023).
- Sáenz-Romero, C.; Guzmán-Reyna, R.R.; Rehfeldt, G.E. Altitudinal genetic variation among Pinus oocarpa populations in Michoacán, Mexico: Implications for seed zoning, conservation, tree breeding and global warming. For. Ecol. Manag. 2006, 229, 340–350. [Google Scholar] [CrossRef]
- Capilla-Dinorin, E.; López-Upton, J.; Jiménez-Casas, M.; Rebolledo-Camacho, V. Reproductive characteristics and seed quality in fragmented populations of Pinus chiapensis (Martínez) Andresen. Rev. Fitotec. Mex. 2021, 44, 211–219. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, biogeography, and evolution of dormancy, and germination. Plant Ecol. 2001, 152, 204–205. [Google Scholar] [CrossRef]
- Mercer, K.L.; Alexander, H.M.; Snow, A.A. Selection on seedling emergence timing and size in an annual plant, Helianthus annuus (Common Sunflower, Asteraceae). Am. J. Bot. 2011, 98, 975–985. [Google Scholar] [CrossRef]
- Zamora-Martínez, M.C.; Velasco Bautista, E.; Muñoz Flores, H.J.; Romero Sánchez, M.E. Modelos Predictivos Para la Producción de Productos Forestales no Maderables: Resina de Pino; Manual Técnico No. 9; INIFAP: Distrito Federal, Mexico, 2013. [Google Scholar]
- Czabator, F.J. Germination value: An index combining speed and completeness of pine seed germination. For. Sci. 1962, 8, 386–396. [Google Scholar] [CrossRef]
- Juárez-Agis, A.; López-Upton, J.; Vargas-Hernández, J.J.; Sáenz-Romero, C. Geographic variation in germination and initial seedling growth of Pseudotsuga menziesii of méxico. Agrociencia 2006, 40, 783–792. Available online: https://www.redalyc.org/pdf/302/30240610.pdf (accessed on 30 October 2023).
- Crookston, N. Research on Forest Climate Change: Potential Effects of Global Warming on Forests and Plant Climate Relationships in Western North America and Mexico; VirginiaTech: Blacksburg, VA, USA, 2006; Available online: https://charcoal2.cnre.vt.edu/climate/ (accessed on 6 September 2023).
- Tuhkanen, S. Climatic parameters and indices in plant geography. Acta Phytogeogr. Suec. 1980, 67, 1–110. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Rehfeldt, G.E.; Crookston, N.L.; Pierre, D.; StAmant, R.; Beaulieu, J.; Richardson, B. Spline models of contemporary, 2030, 2060 and 2090 climates for Mexico and their use in understanding climate-change impacts on the vegetation. Clim. Chang. 2010, 102, 595–623. [Google Scholar] [CrossRef]
- Conover, W.J. The rank transformation-an easy and intuitive way to connect many nonparametric methods to their parametric counterparts for seamless teaching introductory statistics courses. Wiley Interdiscip. Rev. Comput. Stat. 2012, 4, 332–338. [Google Scholar] [CrossRef]
- SAS Institute Inc. Statistical Analysis System; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Gómez-Jiménez, D.; Ramírez-Herrera, C.; Jasso-Mata, J.; López-Upton, J. Variación en características reproductivas y germinación de semillas de Pinus leiophylla Schiede Ex Schltdl. & Cham. Rev. Fitotec. Mex. 2010, 33, 297–304. Available online: https://revistafitotecniamexicana.org/documentos/33-4/3r.pdf (accessed on 28 October 2023).
- López-Upton, J.; Mendoza-Herrera, A.J.; Jasso-Mata, J.; Vargas-Hernández, J.J.; Gómez-Guerrero, A. Variación morfológica de plántulas e influencia del pH del agua de riego en doce poblaciones de Pinus greggii Engelm. Madera Bosques 2016, 6, 81–94. [Google Scholar] [CrossRef]
- Ulusan, M.D.; Bilir, N. Broad-sense heritability for seedling characters and its importance for breeding in scots pine. Sdü Fen Edeb. Fakültesi Fen 2008, 3, 133–138. Available online: https://dergipark.org.tr/tr/download/article-file/116270 (accessed on 29 October 2023).
- Alvarez, R.; Valbuena, L.; Calvo, L. Influence of tree age on seed germination response to environmental factors and inhibitory substances in Pinus pinaster. Int. J. Wildland Fire 2005, 14, 277–284. [Google Scholar] [CrossRef]
- Álvarez-Cisneros, O.; Pérez-Reyes, C.M.; Bonilla-Vichot, M. Evaluación de la viabilidad en semillas de Pinus tropicalis Morelet con diferente tiempo de almacenamiento. Avances 2020, 22, 97–109. Available online: https://www.redalyc.org/journal/6378/637869115007/ (accessed on 26 October 2023).
- Rivera, V.J.C. Germinación de las semillas de Pinus oocarpa Schiede utilizando riego con agua residual sedimentada. Agua Saneam. Ambiente 2012, 7, 36–40. [Google Scholar] [CrossRef]
- Cervantes-Machuca, M.M.; Rodríguez-Ortiz, G.; Enríquez del Valle, J.R.; Rodríguez-Vásquez, M.E. Caracterización morfológica de semillas y eficiencia germinativa de Pinus patula var. longepedunculata y P. Pseudostrobus var. oaxacana. E-CUCBA 2023, 10, 44–51. [Google Scholar] [CrossRef]
- Romero-Rangel, S.; Rubio-Licona, L.E.; Chávez-Serrano, L.; Rojas-Zenteno, E.C.; García-Pineda, M. Germination and early growth of Pinus devoniana and Pinus pseudostrobus (Pinaceae). Biocyt 2017, 10, 749–756. [Google Scholar] [CrossRef]
- Tíscar, P.; Lucas, M. Seed mass variation, germination time and seedling performance in a population of Pinus nigra subsp. salzamannii. For. Syst. 2010, 19, 344–353. [Google Scholar] [CrossRef]
- Romero-Arenas, O.; Rivera-Tapia, J.A.; Lopez-Olguín, J.F.; Espino-Barros, O.A.V.; Huerta-Lara, M.; Parraguirre-Lezama, C. Seed germination of Pinus patula in waste Nutshell (Juglans regia L.) in nursery. CIBA Rev. Iber. Cien. Biol. Y Agrop. 2013, 2, 1–17. [Google Scholar] [CrossRef]
- Bustamante-García, V.; Prieto-Ruíz, J.A.; Merlín-Bermudes, E.; Álvarez-Zagoya, R.; Carrillo-Parra, A.; Hernández-Díaz, J.C. Potential and efficiency of seed production of Pinus engelmannii Carr., in three seed stands of the state of Durango, Mexico. Madera Bosques 2012, 18, 7–21. Available online: https://www.scielo.org.mx/pdf/mb/v18n3/v18n3a2.pdf (accessed on 23 October 2023).
- Ghildiyal, S.K.; Sharma, C.M.; Gairola, S. Environmental variation in seed and seedling characteristics of Pinus roxburghii Sarg. from Uttarakhand, India. Appl. Ecol. Environ. Res. 2009, 17, 121–129. [Google Scholar] [CrossRef]
- Fuentes-Amaro, S.L.; Rodríguez-Laguna, R.; Razo-Zárate, R.; Meza-Rangel, J.; Jiménez-Casas, M.; López-Zepeda, G.A. Energía germinativa en árboles selectos de Pinus patula Schl. et Cham. Rev. Iberoam.Cienc. 2020, 7, 69–78. Available online: http://www.reibci.org/publicados/2020/jul/3900106.pdf (accessed on 1 November 2023).
- Isik, K. Altitudinal variation in Pinus brutia Ten.: Seed and seedling characteristics. Silvae Genet. 1986, 35, 58–67. Available online: https://www.thuenen.de/media/institute/fg/PDF/Silvae_Genetica/1986/Vol._35_Heft_2-3/35_2-3__58.pdf (accessed on 1 November 2023).
- Ortiz-Bibian, M.A.; Castellanos-Acuña, D.; Gómez-Romero, M.; Lindig-Cisneros, R.; Silva-Farías, M.A.; Sáenz-Romero, C. Variation among Abies religiosa (H.B.K.) Schl. et Cham populations along an altitudinal Gradient. I. seed germination capacity. Rev. Fitotec. Mex. 2019, 42, 301–308. [Google Scholar] [CrossRef]
- Parker, W.C.; Noland, T.L.; Morneault, A.E. The effects of seed mass on germination, seedling emergence, and early seedling growth of eastern white pine (Pinus strobus L.). New. For. 2006, 32, 33–49. [Google Scholar] [CrossRef]
- Verdú, M.; Traveset, A. Early emergence enhances plant fitness: A phylogenetically controlled meta-analysis. Ecology 2005, 86, 1385–1394. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | Community | Number of Trees | Altitude (m asl) | Collection Date | Identification Key of Superior Trees |
|---|---|---|---|---|---|
| Rosario | Constancia del Rosario | 27 | 902 | 14 to 15 March 2020 | CDR01, CDR02, …, CDR28 |
| Sesteadero | Putla Villa de Guerrero | 6 | 905 | 16 March 2020 | SES01, SES02, SES03, SES06, SES07, SES11 |
| San Pedro | Santo Domingo Coatlán | 18 | 1130 | 23 to 24 March 2020 | SDC30, SDC31, …, SDC47 |
| El Tizne | Santo Domingo Coatlán | 8 | 1288 | 19 to 20 March 2020 | SDC11, SDC12, …, SDC18 |
| Las Tejas | Santo Domingo Coatlán | 10 | 1302 | 21 to 22 Marzo 2020 | SDC01, SDC02, …, SDC10 |
| El Nanche | Santo Domingo Coatlán | 11 | 1325 | 17 to 18 Marzo 2020 | SDC19, SDC20, …, SDC29 |
| Variable 1 | Mean | Contribution to Total Variance (%) and Significance (p) | Total Variance | ||||
|---|---|---|---|---|---|---|---|
| Provenance | p | Tree (Provenance) | p | Error | |||
| CE | 66.88 | 46.85 | <0.0001 | 28.59 | <0.0001 | 24.55 | 413.03 |
| VPE | 3.84 | 48.65 | <0.0001 | 30.20 | <0.0001 | 21.15 | 1.82 |
| EMD | 3.45 | 39.66 | <0.0001 | 22.52 | <0.0001 | 37.82 | 1.71 |
| VE | 14.71 | 41.07 | <0.0001 | 24.00 | <0.0001 | 34.93 | 84.04 |
| EE | 14.28 | 33.85 | <0.0001 | 40.63 | <0.0001 | 25.52 | 2.16 |
| NCOT | 5.91 | 0.00 | 0.0006 | 11.71 | <0.0001 | 88.29 | 0.66 |
| Provenances | Emergence Capacity (%) | Peak Emergence Value | Average Daily Emergence | Emergence Value | Emergence Energy (Day) | Number of Cotyledons |
|---|---|---|---|---|---|---|
| San Pedro | 81.10 a | 4.59 ab | 4.12 a | 19.53 ab | 14.34 b | 5.90 bc |
| El Tizne | 76.63 b | 4.76 a | 4.20 a | 20.51 a | 13.10 a | 5.90 b |
| Las Tejas | 74.55 bc | 4.36 b | 3.93 bc | 17.99 bc | 13.49 c | 5.81 c |
| El Nanche | 71.93 c | 4.11 c | 3.75 c | 16.11 c | 14.14 c | 5.94 ab |
| Rosario | 52.22 d | 2.86 d | 2.61 d | 8.42 d | 15.01 d | 5.94 ab |
| Sesteadero | 44.84 e | 2.36 e | 2.12 e | 5.71 e | 15.63 e | 6.00 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velasco-García, M.V.; Hernández-Hernández, A. Altitudinal Genetic Variation of Pinus oocarpa Seedling Emergence in the Southern Mountains, Oaxaca, Mexico. Seeds 2024, 3, 1-15. https://doi.org/10.3390/seeds3010001
Velasco-García MV, Hernández-Hernández A. Altitudinal Genetic Variation of Pinus oocarpa Seedling Emergence in the Southern Mountains, Oaxaca, Mexico. Seeds. 2024; 3(1):1-15. https://doi.org/10.3390/seeds3010001
Chicago/Turabian StyleVelasco-García, Mario Valerio, and Adán Hernández-Hernández. 2024. "Altitudinal Genetic Variation of Pinus oocarpa Seedling Emergence in the Southern Mountains, Oaxaca, Mexico" Seeds 3, no. 1: 1-15. https://doi.org/10.3390/seeds3010001
APA StyleVelasco-García, M. V., & Hernández-Hernández, A. (2024). Altitudinal Genetic Variation of Pinus oocarpa Seedling Emergence in the Southern Mountains, Oaxaca, Mexico. Seeds, 3(1), 1-15. https://doi.org/10.3390/seeds3010001