
Seed Desiccation Sensitivity Varies with Geographic Distribution in Two New Zealand Native Pittosporum Species


 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Sources
2.2. Seed Characterization Procedures
2.3. Seed Viability Test
2.4. Seed Germination Procedures
2.5. Seed Desiccation
2.6. Sorption Isotherm Procedures
2.7. Statistical Analysis
3. Results
3.1. Seed Morphology
3.2. Initial Seed Moisture Content, Viability, and Germination
3.3. Seed Desiccation Sensitivity Assessment
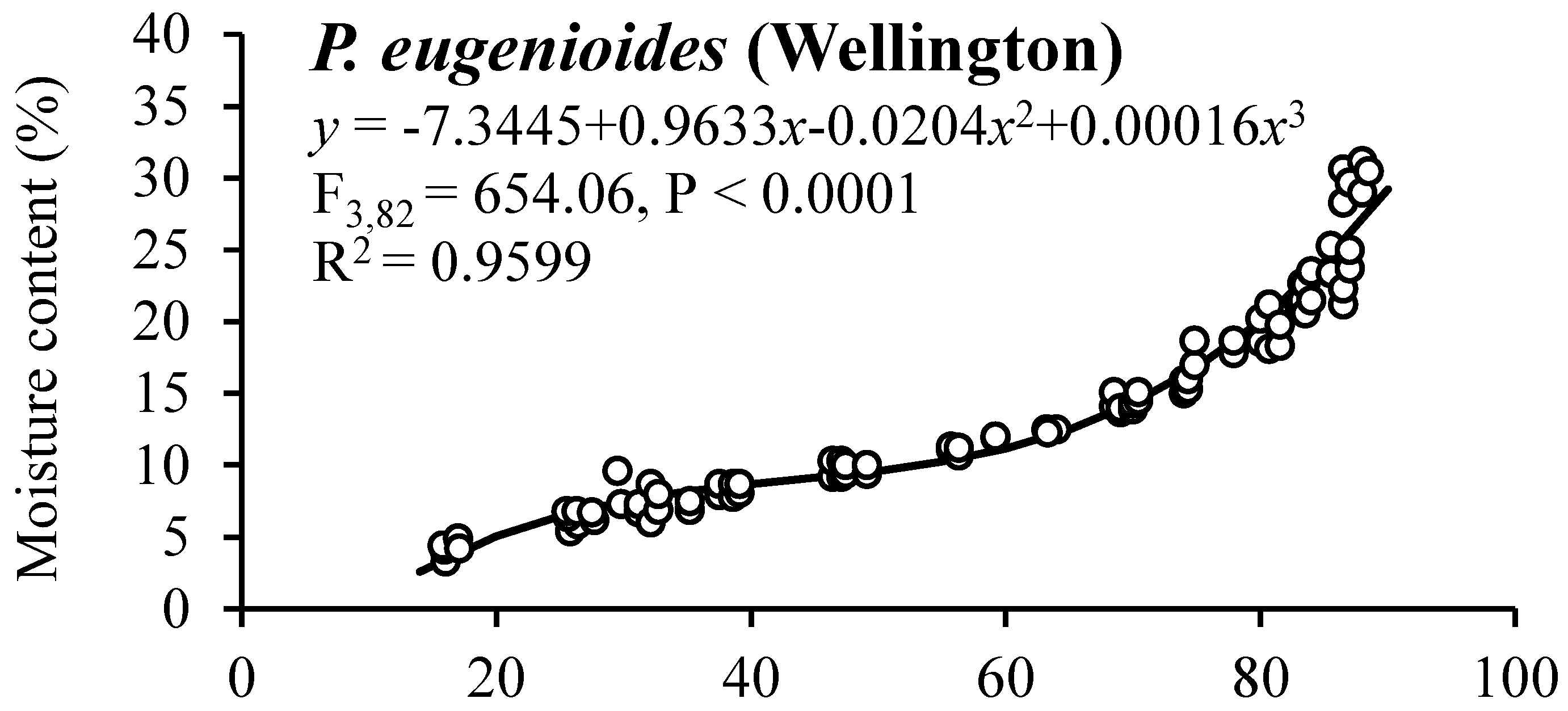
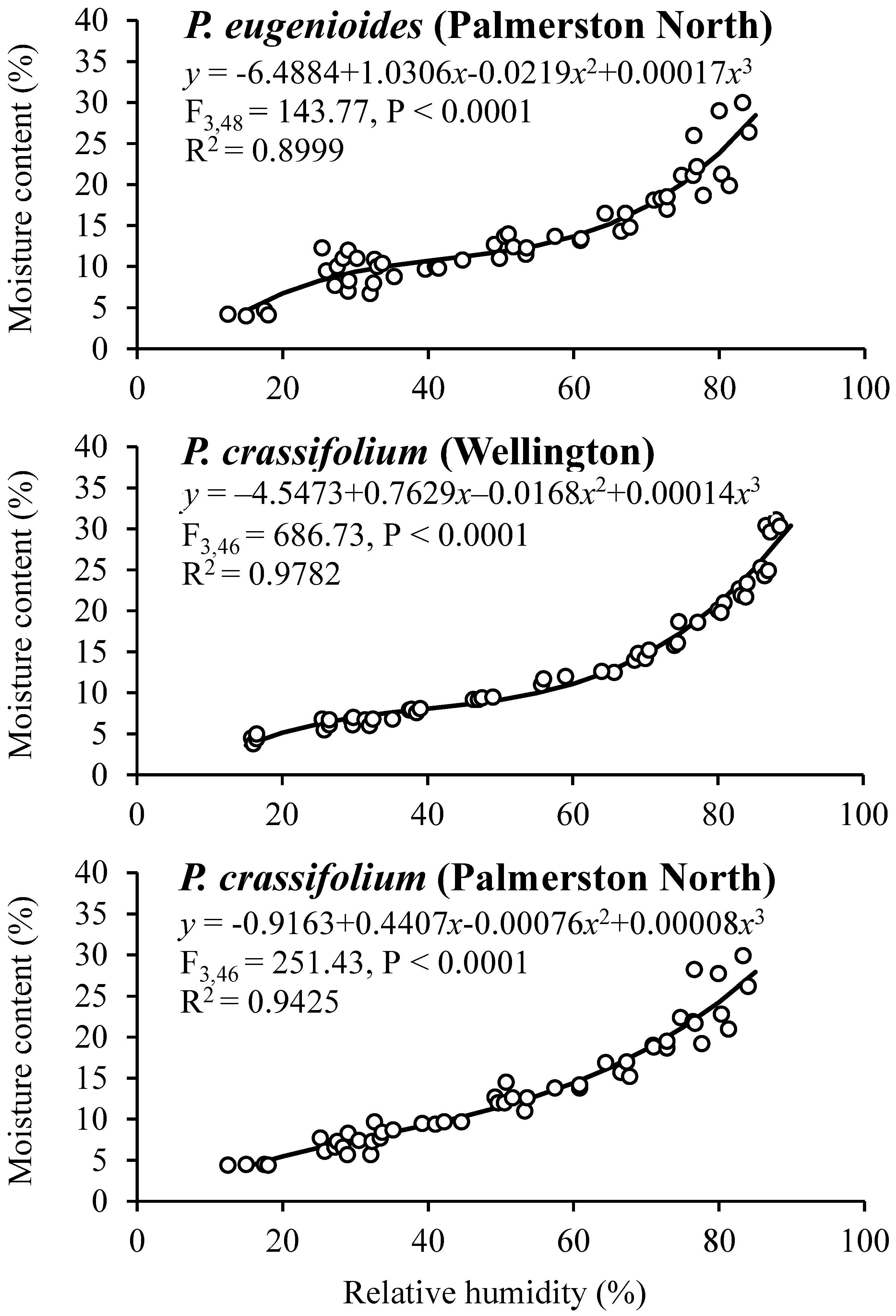
3.4. Seed Sorption Isotherms
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cayzer, L.W.; Crisp, M.D.; Telford, I.R. Revision of Pittosporum (Pittosporaceae) in Australia. Aust. Syst. Bot. 2000, 13, 845–902. [Google Scholar] [CrossRef]
- Haas, J.E. The pacific species of Pittosporum Banks ex Gaertn. (Pittosporaceae). Allertonia 1977, 1, 73–167. [Google Scholar]
- Wilson, E.O. Threats to biodiversity. Sci. Am. 1989, 261, 108–117. [Google Scholar] [CrossRef]
- Kingsford, R.; Watson, J.E.; Lundquist, C.; Venter, O.; Hughes, L.; Johnston, E.; Atherton, J.; Gawel, M.; Keith, D.A.; Mackey, B. Major conservation policy issues for biodiversity in Oceania. Conserv. Biol. 2009, 23, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Breitwieser, I.; Brownsey, P.J.; Nelson, W.A.; Smissen, R.; Wilton, A.D. (Eds.) Pittosporum Banks & Sol. ex Gaertn. 2010. Available online: http://www.nzflora.info/factsheet/Taxon/Pittosporum.html (accessed on 20 October 2020).
- De Lange, P.J.; Rolfe, J.R.; Barkla, J.W.; Courtney, S.P.; Champion, P.D.; Perrie, L.R.; Beadel, S.M.; Ford, K.A.; Breitwieser, I.; Schönberger, I.; et al. Conservation Status of New Zealand Indigenous Vascular Plants, 2017; Publishing Team, Department of Conservation: Wellington, New Zealand, 2018. [Google Scholar]
- Swarts, N.D.; Dixon, K.W. Perspectives on orchid conservation in botanic gardens. Trends Plant Sci. 2009, 14, 590–598. [Google Scholar] [CrossRef] [PubMed]
- FAO/IPGRI. Genebank Standards for Plant Genetic Resources for Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 1994. [Google Scholar]
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Prediction of desiccation sensitivity in seeds of woody species: A probabilistic model based on two seed traits and 104 species. Ann. Bot. 2006, 97, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Mayrinck, R.C.; Vilela, L.C.; Pereira, T.M.; Rodrigues-Junior, A.G.; Davide, A.C.; Vaz, T.A.A. Seed desiccation tolerance/sensitivity of tree species from Brazilian biodiversity hotspots: Considerations for conservation. Trees 2019, 33, 777–785. [Google Scholar] [CrossRef]
- Walters, C. Orthodoxy, recalcitrance and in-between: Describing variation in seed storage characteristics using threshold responses to water loss. Planta 2015, 242, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Ellis, R.H. A Protocol to Determine Seed Storage Behaviour; Bioversity International: Rome, Italy, 1996. [Google Scholar]
- Orwin, J. Shrubs and Small Trees of the Forest—Pittosporums. Available online: http://www.TeAra.govt.nz/en/shrubs-and-small-trees-of-the-forest/page-8 (accessed on 8 October 2020).
- Kew, R.B.G. Seed Information Database (SID); Version 7.1; Society for Ecological Restoration (SER): Washington, DC, USA, 2008. [Google Scholar]
- De Lange, P.J. Pittosporum tenuifolium Fact Sheet (Content Continuously Updated). Available online: https://www.nzpcn.org.nz/flora/species/pittosporum-tenuifolium/ (accessed on 21 October 2020).
- De Lange, P.J. Pittosporum eugenioides Fact Sheet (Content Continuously Updated). Available online: https://www.nzpcn.org.nz/flora/species/pittosporum-eugenioides/nzpcn.org.nz/flora_details.aspx?ID=1135 (accessed on 21 October 2020).
- Weston, R.J. Composition of essential oils from the leaves of seven New Zealand species of Pittosporum (Pittosporaceae). J. Essent. Oil Res. 2004, 16, 453–458. [Google Scholar] [CrossRef]
- De Lange, P.J. Pittosporum crassifolium Fact Sheet (Content Continuously Updated). Available online: https://www.nzpcn.org.nz/flora/species/pittosporum-crassifolium/ (accessed on 28 September 2019).
- Crowe, A. Which Native Tree? A Simple Guide to the Identification of New Zealand Native Trees; Viking Pacific: Delta, BC, Canada, 1992. [Google Scholar]
- NIWA. Climate Summaries. Available online: https://niwa.co.nz/education-and-training/schools/resources/climate/summary (accessed on 1 December 2019).
- ISTA. International Rules for Seed Testing 2015; The International Seed Testing Association: Washington, DC, USA, 2015. [Google Scholar]
- Yu, K. Dessication Response of Seed of Clianthus spp., Carmichaelia muritai, Pittosporum crassifolium and Pittosporum eugenoides. Master’s Thesis, (Presented in Partial Fulfilment of the Requirements). AgriScience in Horticulture, Massey University, Palmerston North, New Zealand, 2015. [Google Scholar]
- Madsen, M.D.; Davies, K.W.; Boyd, C.S.; Kerby, J.D.; Svejcar, T.J. Emerging seed enhancement technologies for overcoming barriers to restoration. Restor. Ecol. 2016, 24, S77–S84. [Google Scholar] [CrossRef]
- Moore, S.; Bannister, P.; Jameson, P.E. The effects of low temperatures on seed germination of some New Zealand species of Pittosporum. N. Z. J. Bot. 1994, 32, 483–485. [Google Scholar] [CrossRef]
- IPGRI-DFSC. The desiccation and storage protocol. In Comparative Storage Biology of Tropical Tree Seeds; Sacande, M., Joker, D., Dulloo, M.E., Thomsen, K.A., Eds.; International Plant Genetic Resources Institute (IPGRI): Rome, Italy, 2004; pp. 345–351. [Google Scholar]
- Gold, K.; Hay, F. Equilibrating seeds to specific moisture levels. In Technical Information Sheet_09; Royal Botanic Gardens Kew: London, UK, 2014. [Google Scholar]
- SAS. SAS/OR 9.3 User’s Guide: Mathematical Programming Examples; SAS Institute: Cary, NC, USA, 2012. [Google Scholar]
- Hong, T.; Ellis, R.; Astley, D.; Pinnegar, A.; Groot, S.; Kraak, H. Survival and vigour of ultra-dry seeds after ten years of hermetic storage. Seed Sci. Technol. 2005, 33, 449–460. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Plant population differences in dormancy and germination characteristics of seeds: Heredity or environment? Am. Midl. Nat. 1973, 90, 493–498. [Google Scholar] [CrossRef]
- Burrows, C. Germination behaviour of the seeds of six New Zealand woody plant species. N. Z. J. Bot. 1995, 33, 365–377. [Google Scholar] [CrossRef]
- Burrows, C. Germination behaviour of the seeds of seven New Zealand woody plant species. N. Z. J. Bot. 1996, 34, 355–367. [Google Scholar] [CrossRef]
- Burrows, C. Fruit types and seed dispersal modes of woody plants in Ahuriri Summit Bush, Port Hills, western Banks Peninsula, Canterbury, New Zealand. N. Z. J. Bot. 1994, 32, 169–181. [Google Scholar] [CrossRef]
- FAO. A Guide to Forest Seed Handling; FAO Forestry Paper 20/2; Food and Agriculture Organization: Rome, Italy, 1985. [Google Scholar]
- Vertucci, C.W.; Roos, E.E. Theoretical basis of protocols for seed storage II. The influence of temperature on optimal moisture levels. Seed Sci. Res. 1993, 3, 201–213. [Google Scholar] [CrossRef]
- Chen, C. Moisture sorption isotherms of pea seeds. J. Food Eng. 2003, 58, 45–51. [Google Scholar] [CrossRef]
- Menkov, N.D. Moisture sorption isotherms of lentil seeds at several temperatures. J. Food Eng. 2000, 44, 205–211. [Google Scholar] [CrossRef]
- Adkins, S.W.; Bellairs, S.M.; Loch, D.S. Seed dormancy mechanisms in warm season grass species. Euphytica 2002, 126, 13–20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| LiCl Weight (g) | Target RH (%) at 20 °C | RH (%) Measured at 20 °C | |
|---|---|---|---|
| Palmerston North Collection | Wellington Collection | ||
| 174 | 11 | 28.0 ± 1.04 | 26.7 ± 0.64 |
| 147 | 15 | 29.2 ± 0.76 | 28.4 ± 0.95 |
| 128 | 20 | 30.0 ± 0.76 | 32.1 ± 0.76 |
| 104 | 30 | 35.7 ± 1.24 | 38.1 ± 0.25 |
| 88 | 40 | 41.7 ± 0.70 | 47.4 ± 0.37 |
| 74 | 50 | 51.3 ± 0.52 | 58.4 ± 1.06 |
| 60 | 60 | 56.1 ± 1.89 | 66.0 ± 1.02 |
| 50 | 70 | 60.6 ± 2.63 | 72.0 ± 0.75 |
| 34 | 80 | 69.1 ± 1.31 | 78.1 ± 0.89 |
| 26 | 85 | 74.5 ± 1.06 | 82.7 ± 0.34 |
| 20 | 90 | 78.2 ± 1.01 | 85.7 ± 0.44 |
| 8 | 95 | 80.9 ± 1.13 | 87.6 ± 0.28 |
| Species | Location | Length (mm) | Breadth (mm) | Weight 1000 Seeds (g) |
|---|---|---|---|---|
| P. eugenioides | Palmerston North | 2.86 ± 0.029 A | 2.38 ± 0.027 A | 4.94 ± 0.560 A |
| Wellington | 2.87 ± 0.022 A | 2.21 ± 0.027 A | 5.56 ± 0.401 A | |
| P. crassifolium | Palmerston North | 4.38 ± 0.031 A | 3.33 ± 0.036 A | 26.46 ± 1.205 A |
| Wellington | 4.59 ± 0.035 A | 3.61 ± 0.032 A | 31.99 ± 3.034 A |
| Specie | Location | SMC (%) | Viability (%) | Germination (%) |
|---|---|---|---|---|
| P. eugenioides (mature) | Palmerston North | 10.8 ± 0.14 A | 89 ± 3.4 A | 16 ± 2.6 A |
| Wellington | 19.9 ± 0.27 B | 99 ± 1.0 B | 93 ± 1.3 B | |
| P. crassifolium | Palmerston North | 15.8 ± 0.64 A | 82 ± 4.2 A | 75 ± 2.9 A |
| Wellington | 16.8 ± 0.35 A | 82 ± 6.6 A | 53 ± 6.7 B |
| Species (Location) | Target SMC (%) | Drying Days | Actual SMC (%) | Germination (%) |
|---|---|---|---|---|
| P. eugenioides | ||||
| (Palmerston North) | 5 | 17 | 5.3 ± 0.19 C | 32 ± 4.4 B |
| 3 | 23 | 3.5 ± 0.01 C | 29 ± 4.0 BC | |
| Control * | No drying | 41.5 ± 0.98 A | 69 ± 1.7 A | |
| P. eugenioides | ||||
| (Wellington) | 10 | 9 | 10.1 ± 0.26 C | 87 ± 1.3 A |
| 5 | 32 | 4.6 ± 0.14 D | 56 ± 4.1 B | |
| 3 | 40 | 3.9 ± 0.31 D | 9 ± 2.2 C | |
| Control * | No drying | 41.8 ± 1.38 A | 93 ± 2.4 A | |
| P. crassifolium | ||||
| (Palmerston North) | 10 | 8 | 9.8 ± 0.06 C | 59 ± 2.1 AB |
| 5 | 11 | 5.4 ± 0.17 CD | 56 ± 1.8 AB | |
| 3 | 14 | 3.9 ± 0.07 D | 40 ± 1.4 B | |
| Control * | No drying | 35.8 ± 2.50 A | 68 ± 8.8 A | |
| P. crassifolium | ||||
| (Wellington) | 10 | 7 | 9.8 ± 0.32 C | 50 ± 3.4 B |
| 5 | 16 | 5.3 ± 0.14 D | 44 ± 7.5 B | |
| 3 | 34 | 3.3 ± 0.20 D | 36 ± 3.2 B | |
| Control * | No drying | 41.2 ± 1.60 A | 73 ± 3.6 A |
| Equilibrating with Silica Gel and Weight of LiCl (g) | Target RH (%) at 20 °C | SMC (%) | Germination (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| P. eugenioides | P. crassifolium | P. eugenioides | P. crassifolium | ||||||
| Palmerston North | Wellington | Palmerston North | Wellington | Palmerston North | Wellington | Palmerston North | Wellington | ||
| Silica gel | 5 | 4.3 ± 0.15 H | 4.0 ± 0.29 J | 4.4 ± 0.03 I | 4.3 ± 0.22 I | 17 ± 4.9 A | 60 ± 4.9 B | 62 ± 6.7 A | 49 ± 5.3 B |
| 174 | 11 | 8.9 ± 1.30 G | 6.1 ± 0.11 IJ | 6.3 ± 0.48 HI | 6.1 ± 0.26 HI | 18 ± 3.6 A | 85 ± 4.9 A | 72 ± 2.9 A | 49 ± 4.4 B |
| 147 | 15 | 9.7 ± 1.07 FG | 7.8 ± 0.76 HI | 7.2 ± 0.40 GH | 6.3 ± 0.13 H | 25 ± 3.5 A | 91 ± 2.4 A | 65 ± 5.0 A | 64 ± 5.9 AB |
| 128 | 20 | 9.8 ± 0.57 FG | 7.4 ± 1.16 HI | 7.6 ± 0.18 FGH | 6.8 ± 0.09 H | 23 ± 4.7 A | 88 ± 0.8 A | 68 ± 6.1 A | 66 ± 1.2 AB |
| 104 | 30 | 10.1 ± 0.27 FG | 8.7 ± 0.06 GH | 9.1 ± 0.36 FG | 7.9 ± 0.06 GH | 19 ± 3.5 A | 91 ± 1.9 A | 73 ± 5.2 A | 74 ± 3.5 AB |
| 88 | 40 | 10.1 ± 0.27 FG | 10.1 ± 0.06 FG | 9.5 ± 0.05 F | 9.3 ± 0.06 G | 22 ± 5.2 A | 90 ± 2.1 A | 78 ± 2.2 A | 77 ± 6.6 A |
| 74 | 50 | 12.1 ± 0.58 EFG | 11.7 ± 0.32 EF | 11.9 ± 0.32 E | 11.5 ± 0.41 F | 25 ± 4.9 A | 89 ± 2.2 A | 82 ± 3.8 A | 77 ± 4.1 A |
| 60 | 60 | 12.9 ± 0.28 EF | 13.3 ± 0.65 DE | 13.3 ± 0.43 DE | 13.2 ± 0.42 F | 21 ± 1.7 A | 94 ± 1.5 A | 79 ± 5.2 A | 81 ± 5.3 A |
| 50 | 70 | 14.2 ± 0.22 DE | 14.7 ± 0.34 D | 14.6 ± 0.26 D | 15.3 ± 0.41 E | 24 ± 6.4 A | 94 ± 0.5 A | 80 ± 2.5 A | 77 ± 5.5 A |
| 34 | 80 | 17.0 ± 0.44 CD | 17.7 ± 0.35 C | 17.6 ± 0.44 C | 19.6 ± 0.62 D | 24 ± 5.7 A | 94 ± 1.5 A | 79 ± 1.9 A | 78 ± 3.5 A |
| 26 | 85 | 19.4 ± 0.93 BC | 20.3 ± 0.72 B | 19.5 ± 0.36 C | 21.7 ± 0.67 C | 25 ± 4.0 A | 96 ± 1.2 A | 74 ± 2.6 A | 78 ± 6.0 A |
| 20 | 90 | 20.9 ± 0.32 B | 22.3 ± 0.61 B | 21.8 ± 0.31 B | 24.4 ± 0.40 B | 22 ± 3.3 A | 90 ± 2.8 A | 70 ± 4.2 A | 79 ± 3.8 A |
| 8 | 95 | 28.0 ± 1.03 A | 29.2 ± 0.48 A | 28.0 ± 0.77 A | 30.3 ± 0.29 A | 26 ± 3.8 A | 91 ± 1.3 A | 73 ± 4.5 A | 86 ± 3.5 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kai, Y.; Alfaro Pinto, A.; Clavijo McCormick, A.; Nadarajan, J.; He, X.Z.; MacKay, M.; McGill, C. Seed Desiccation Sensitivity Varies with Geographic Distribution in Two New Zealand Native Pittosporum Species. Seeds 2023, 2, 370-381. https://doi.org/10.3390/seeds2030028
Kai Y, Alfaro Pinto A, Clavijo McCormick A, Nadarajan J, He XZ, MacKay M, McGill C. Seed Desiccation Sensitivity Varies with Geographic Distribution in Two New Zealand Native Pittosporum Species. Seeds. 2023; 2(3):370-381. https://doi.org/10.3390/seeds2030028
Chicago/Turabian StyleKai, Yu, Alejandra Alfaro Pinto, Andrea Clavijo McCormick, Jayanthi Nadarajan, Xiong Zhao He, Marion MacKay, and Craig McGill. 2023. "Seed Desiccation Sensitivity Varies with Geographic Distribution in Two New Zealand Native Pittosporum Species" Seeds 2, no. 3: 370-381. https://doi.org/10.3390/seeds2030028
APA StyleKai, Y., Alfaro Pinto, A., Clavijo McCormick, A., Nadarajan, J., He, X. Z., MacKay, M., & McGill, C. (2023). Seed Desiccation Sensitivity Varies with Geographic Distribution in Two New Zealand Native Pittosporum Species. Seeds, 2(3), 370-381. https://doi.org/10.3390/seeds2030028