1. Introduction
Scots pine (
Pinus sylvestris L.) has a wide range of habitat [
1] and is one of the primary sources of wood raw materials in boreal countries. The seeds of Scots pine as integral components of forest reproductive material (FRM) [
2] are valuable products, moved by trade operations, as shown in [
3], for significant distances, and to some extent determining the rate of forest landscape restoration technology (FLR technology) [
4,
5]. Thus, the leading importer of Scots pine seeds in Europe is Sweden (about 175 kg per year, the total amount for Europe is 272.76 kg), and the origin of these seeds belongs exclusively to Finland. Quality, defined by appropriate indicators, characterizes any product. Improvement of quality indicators and competitiveness of forest seeds is one of the promising directions of the development of forest seed production. This direction assumes the active interaction of private forest users with forest seed producers, which is consistent with the realities in the forest legislation and the basic provisions of Restoration Opportunities Assessment Methodology [
6] for use in global reforestation initiatives [
7,
8,
9].
One of the important quality indicators for the seeds of Scots pine is ground germination. Germination depends on the «year of stand formation, stratification substrate, and individual characteristics of mother trees» [
10], temperature conditions [
11] and seed viability [
12]. The outdoor germination test carried out in a container nursery [
12] differs sharply from the test values in the open ground only when the seed viability is low. Germination in the ground of conifer seeds is closely related to temperature conditions [
11] and occurs more slowly and unstably [
13] compared to seeds of agricultural crops, which leads to additional costs for greenhouse heating in nurseries.
The operation of conifer seed sorting [
14] before sowing [
15] to improve the quality of produced seedlings is laid by the world’s leading manufacturers of forest nursery equipment (for example, BCC AB, Sweden), setting the pace of development of forest reproductive material technology. Traditionally, seeds are consistently separated in such technology using a standard machine [
16] according to geometrical and gravitational, i.e., quantitative, traits. However, the spectrometric properties [
17,
18,
19,
20,
21,
22] of seeds should not be neglected either, since their use provides non-invasive quality (seed viability) control [
23].
The basis for the study was the relatively small amount of data on germination in containers of Scots pine seeds separated using optical graders [
24,
25] by the seed coat colour and quantitative traits. Novikov et al. [
26], conducting theoretical studies of the single seed detection process of Scots pine, point out that «the seed spectrometric feature, both in the visible (VIS) and near-infrared (NIR) regions has a low identification error [
27], and is best suited for automating seed grading processes [
28]».
The seeds of Scots pine within the range are characterized by a large variety of seed coat colours [
29,
30]. The individuality of seed colour of Scots pine and its stability in the course of pine ontogenesis [
31] has been noted in studies [
32,
33,
34,
35,
36]. Seed colouration is a phenotypic marker in the taxonomy and ecological and geographical differentiation of this species. Therefore, this trait is used to characterize the genotypic composition of populations representing this polymorphic species and other morphophysiological traits. Currently, 50 variations of this trait have been identified in pine. It is also known that the colouration of mature, full-grown seeds is uniform within the crown of one tree and does not change in ontogeny, which allows effective use of this trait of the maternal individual, preserved in vegetative progeny.
Various explanations for seed polymorphism of
Pinus sylvestris L. are known: protection of seeds from eating by birds [
37], the influence of growing conditions [
35], and the impact of environmental factors. Studies on the relationship between seed colouration and germination are heterogeneous and contradictory [
38,
39]. Several forms of
Pinus sylvestris L. are distinguished [
35]: black—
Pinus sylvestris L. var.
melanosperma Litv.; yellow—
Pinus sylvestris L. var.
leucosperma Litv.; brown—
Pinus sylvestris L. var.
phoeosperma Litv.; and mottled—
Pinus sylvestris L. f.
cinnamomeosperma mihi. In her dissertation, Pinaevskaya [
40], according to the literature data, provides information on the forms of pine with mono- and polychromatic colour variants, noting that many authors establish at least three forms—black, brown, and light/yellow/white.
The idea of using seed colour polymorphism is to form a forest plantation of pine trees with one form (i.e., with seeds of the same colour), which will stabilize the seed quality in each lot. If there are differences in sowing qualities of seeds of different colours, using seed colour sorting devices can produce more productive plantations. If there are no differences in seed quality, sorting by colour makes sense for more complete control of the origin of the seed.
Thus, the study aims to determine the effect of seed coat colour, seed size, and seed provenance on the seed quality of Scots pine species.
4. Discussion
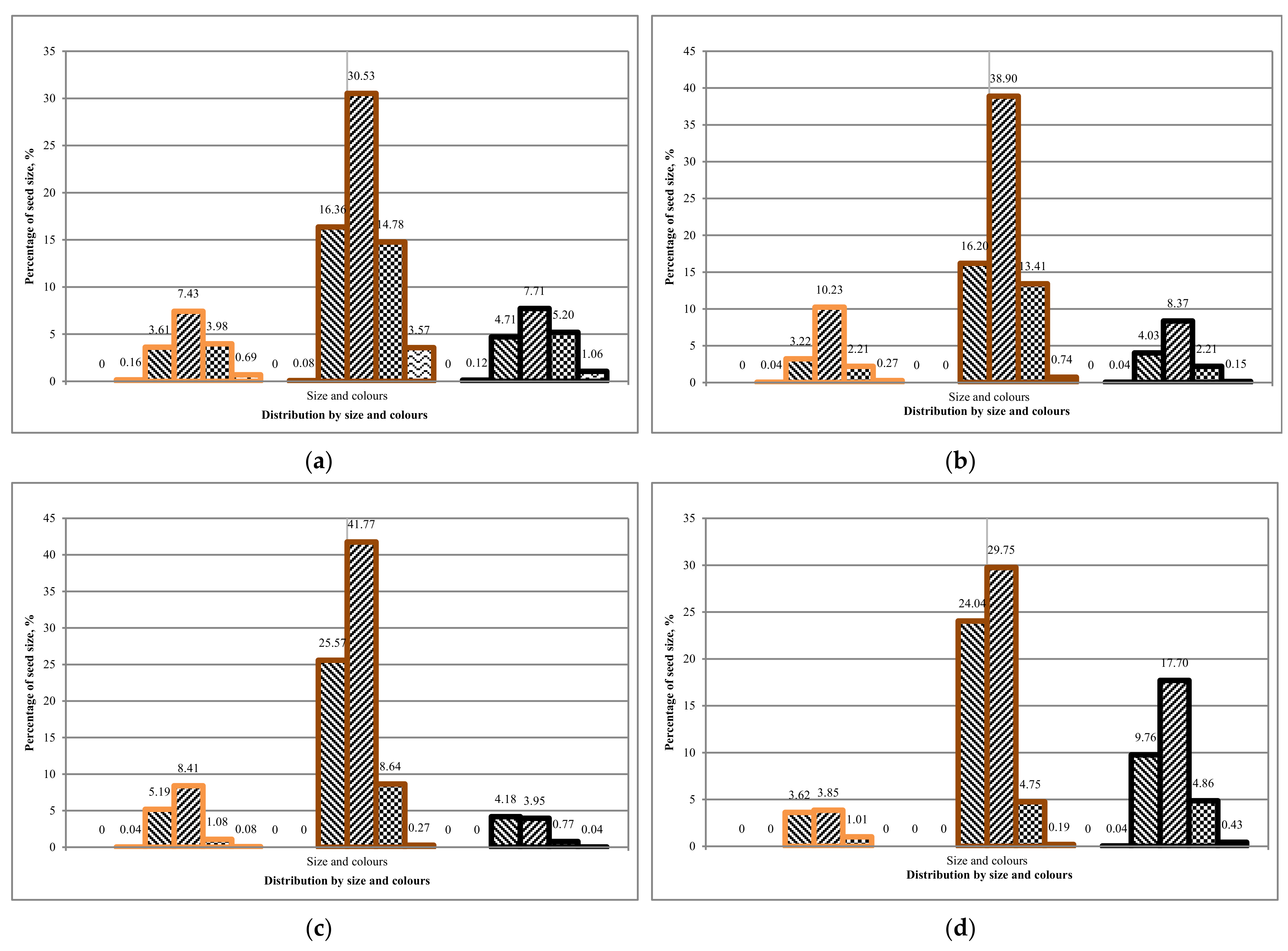
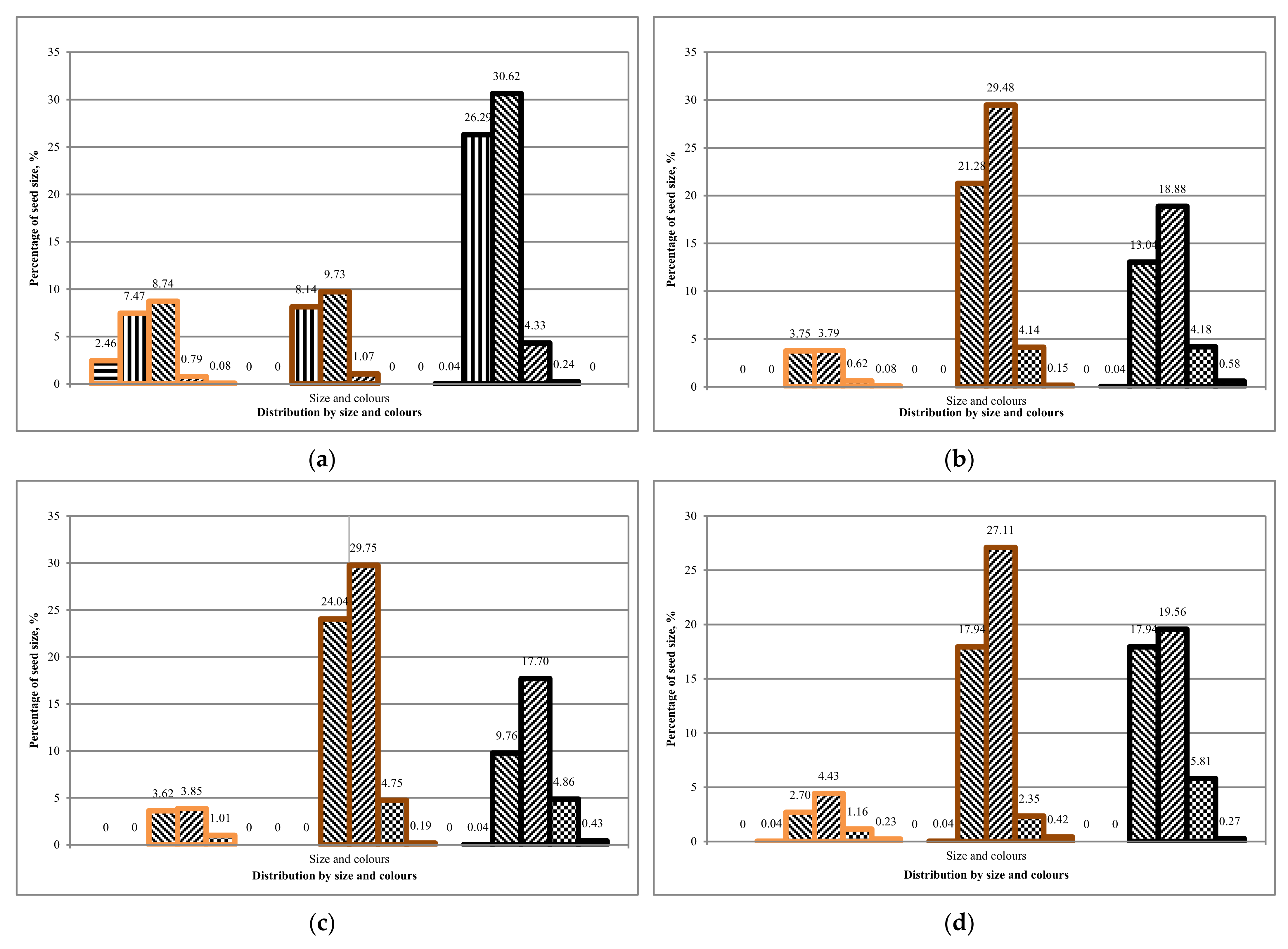
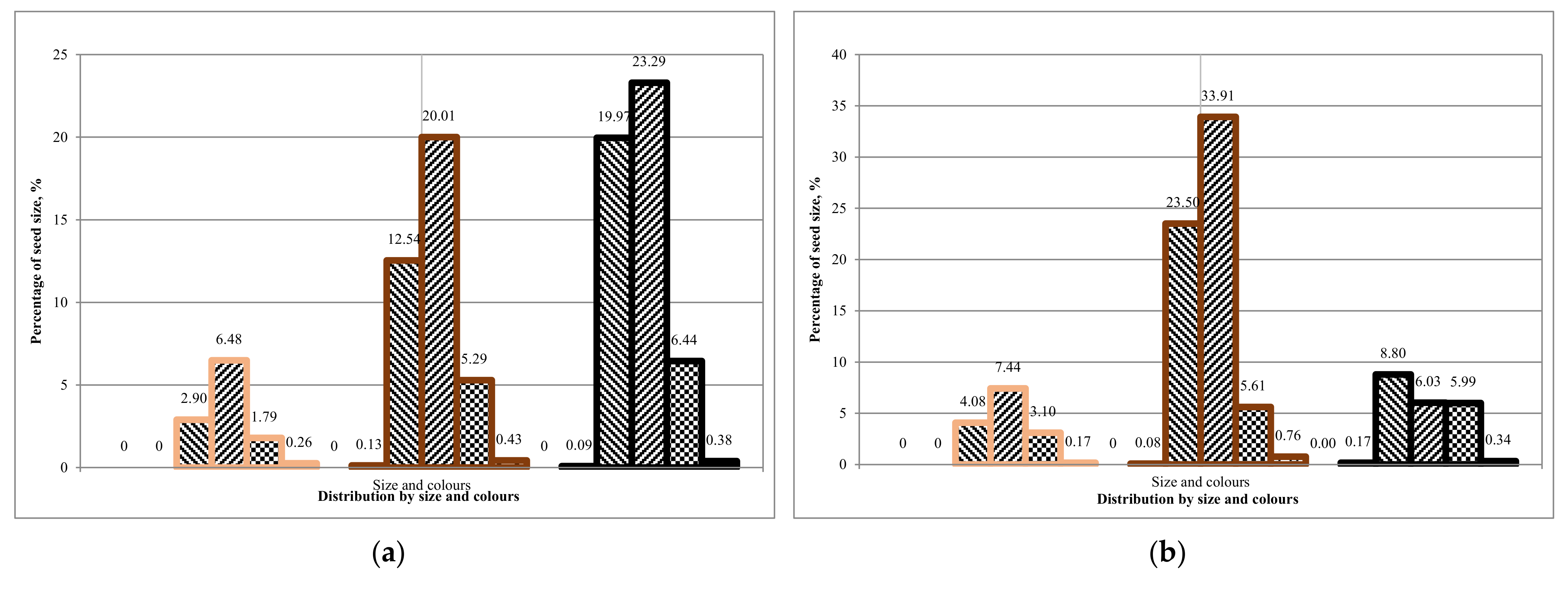
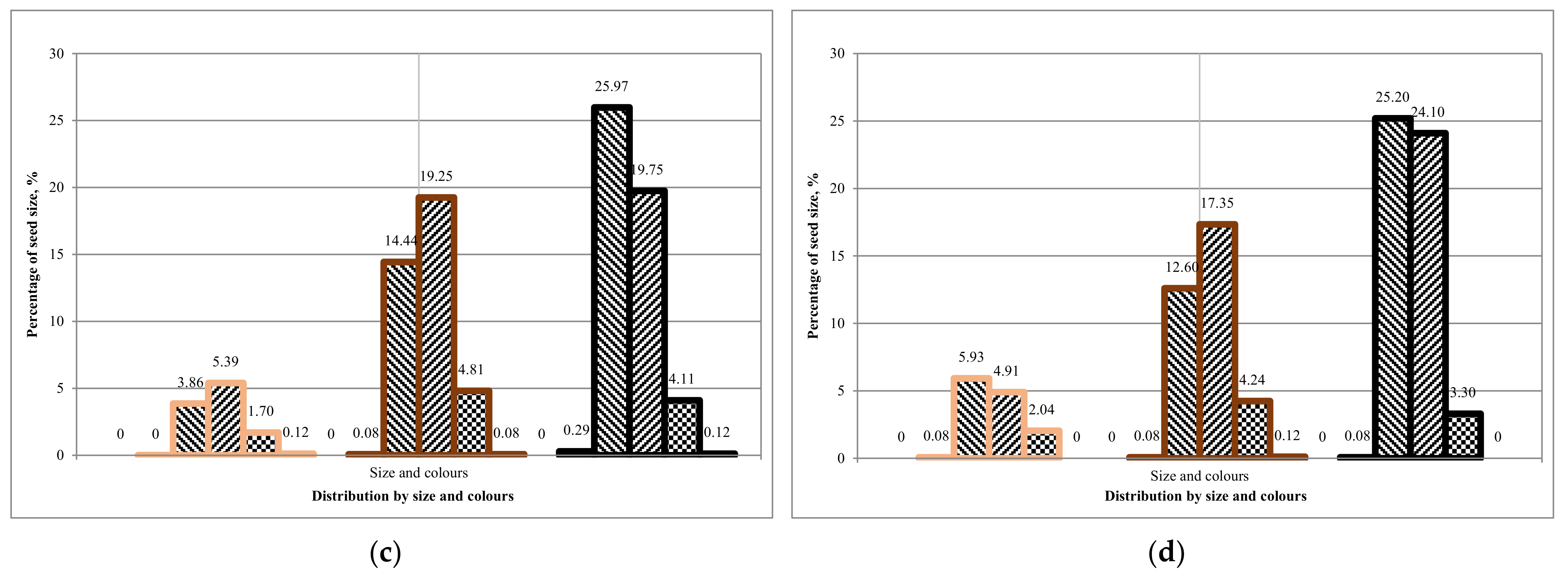
Comparing the first and second rotation of Tambov seeds, we can conclude that the average ground germination is 25 ± 5%. The average ground germination of seeds from Komi is 15 ± 5%. It can be concluded that there is a difference between the sowing qualities of seeds from different regions,
Table 13. However, is there a dependence on seed coat colour and seed size?
The results obtained on the soil germination of seeds showed that it is more expedient to use unsorted seeds from the Komi region for growing in the first rotation. Seeds from the Tambov region can be grown according to a two-rotation scheme for the growing season.
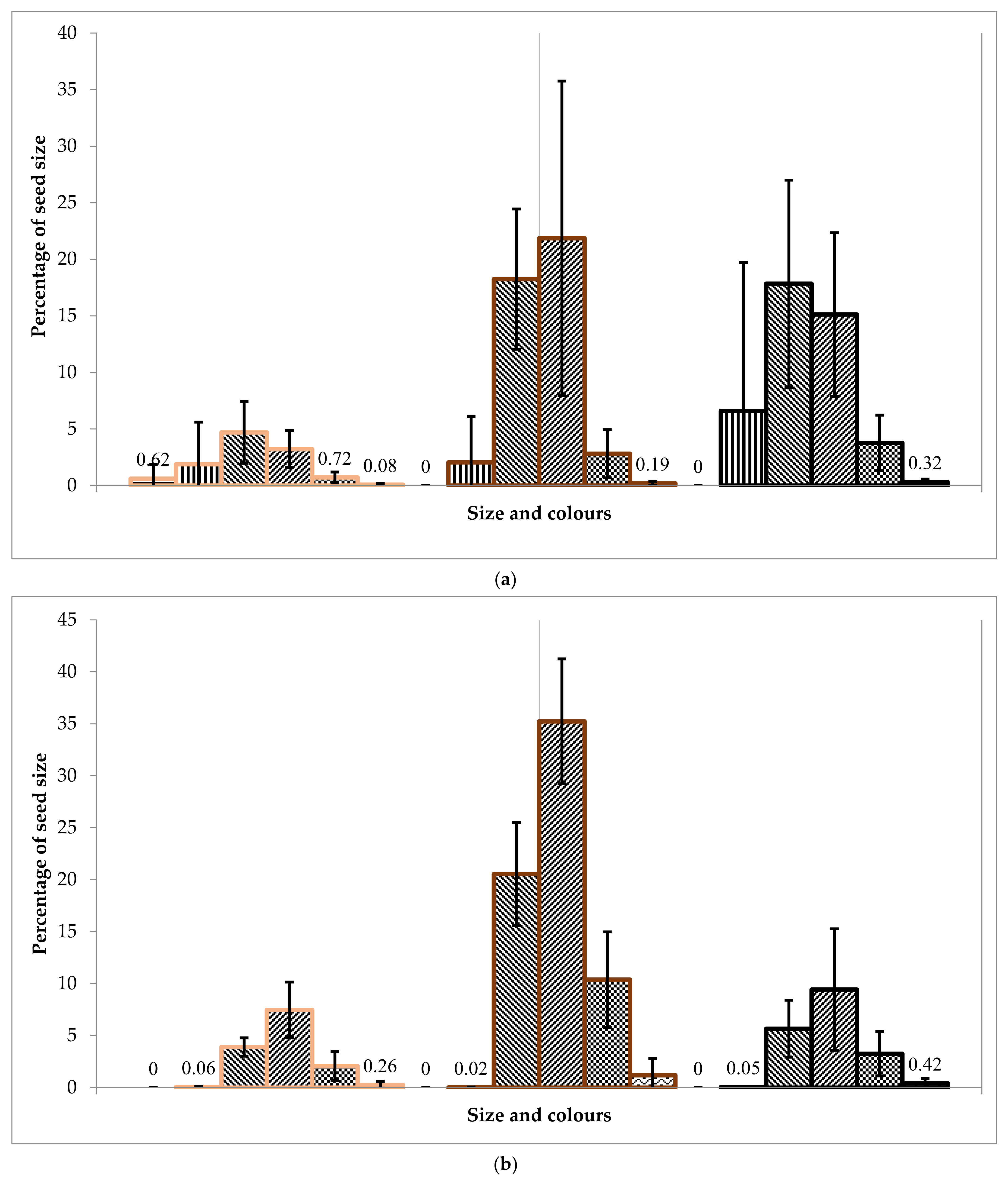
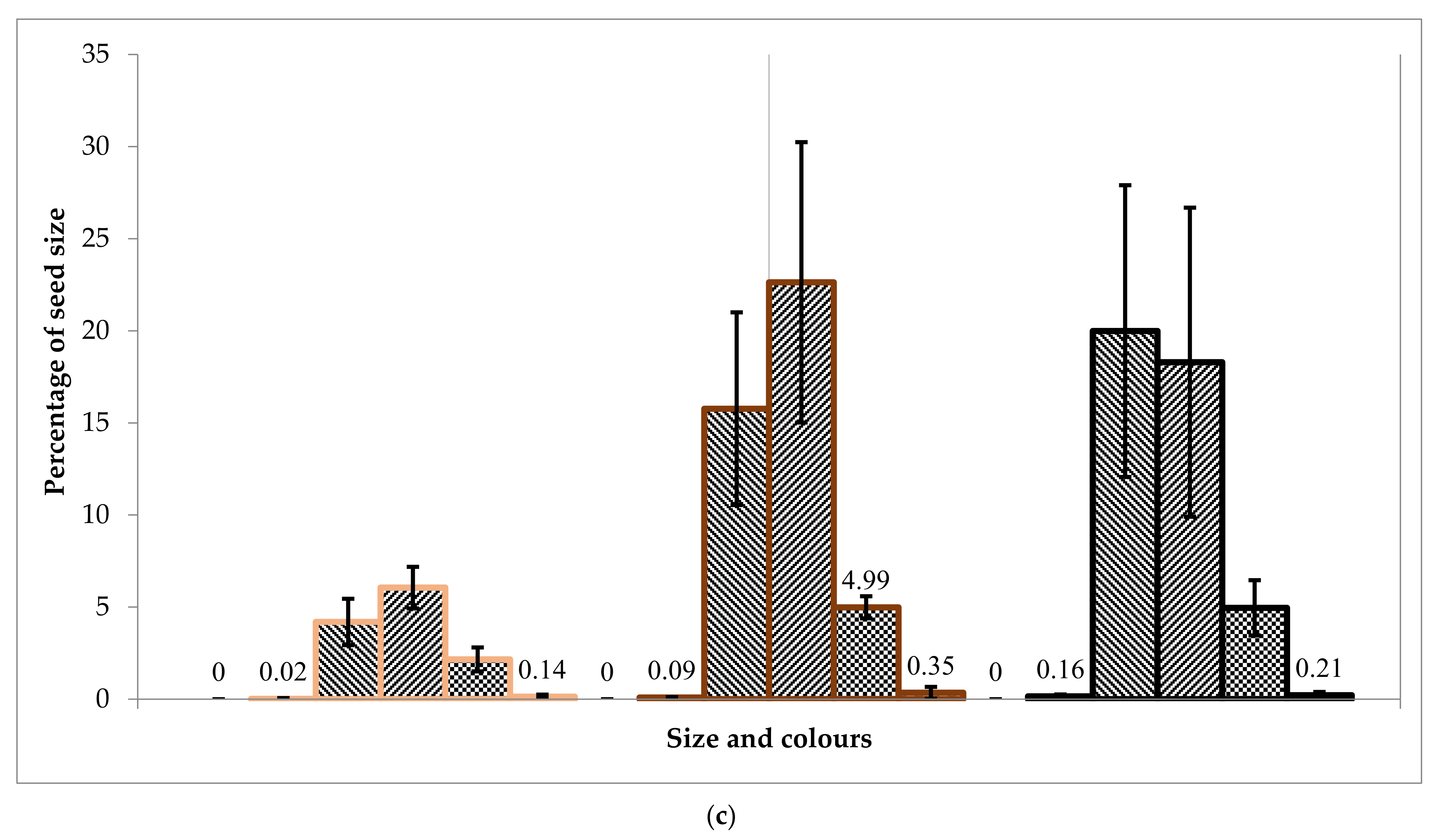
A laboratory experiment showed that the seed size distribution corresponds to the Gaussian distribution. The fraction of 1.7 to 2.0 mm (medium-sized seeds) prevails in all cases and colours in 30–40%, except white for each batch. The overall low germination efficiency of seeds was recorded in all the collected seed batches.
The laboratory germination of the Tambov’s seedlot is higher than the Lisino’s seedlot, and its soil germination is higher than the soil germination in the batch of seeds from Komi, which is explained by the more southern region of seed origin. Since the Tambov’s seedlot was stored for two years in the seed storage, and the Lisino’s seedlot for four years, it can be assumed that the decrease in germination is due to the duration of the storage period. However, at the same time, the germination of medium and large seeds of the brown fraction decreases slightly, due to a larger supply of nutrients in the endosperm.
The colour of pine seeds may well have a certain breeding value. Thus, in Estonia, there was less ground germination in light seeds and worse growth of seedlings from black seeds [
45]. In Ukraine, 40-year-old crops planted from 12 groups of seeds by colour were studied, and only cultures from light seeds were preserved significantly worse [
46]. In the study of pine seeds from 93 points [
47], light seeds were characterized by reduced germination energy, which was 75% versus 81% for black and variegated seeds. In Belarus, in the Minsk region, there was less damage to the crops of shoots created from dark seeds of pine seedlings [
48], and seedlings from dark seeds are also resistant to fusarium [
49].
However, offspring from light seeds do not grow worse in all regions. In the north-west of Russia, seedlings from light seeds are equivalent and even surpass in height plants obtained from seeds of other colours [
50].
Some researchers believe that the colour of the seeds is associated with the conditions of the place of growth and light tones are inherent in fresh and dry pine forests, and seeds with a predominance of dark shade are inherent in moist growing conditions [
35]. In phenetic studies of pine, the study of the variability of the seed colour structures showed that they are determined by three layers and have a high degree of heritability, and the frequency of phenols of the third colour layer of seeds can serve as markers of pine populations [
51]. In Rayevsky’s dissertation [
52], on the basis of a system of methods for isolating and evaluating phenes of Vidyakin’s Scots pine, five phenotypes were isolated for the conditions of the Petrozavodsk forest seed plantation (Republic of Karelia, Russia) and it was found that clones with dark brown and brown seed colouration have the highest seed productivity.
Studies have shown that plantations with different proportions of dark and light-coloured seeds are represented within the boundaries of the area of Scots pine. At the same time, trees with dark-coloured seeds predominate in most plantings. A particularly large representation of pine trees with dark seeds is characteristic of rich and sufficiently moist ecotopes. Pine trees with light-coloured seeds are more common in the dry forests of the forest-steppe zone (southern regions), as well as in pessimal conditions characterized by a cold thermal regime and excessive moistening of plantings near the northern border of the range [
51]. The productivity of such pine forests is usually estimated at no higher than the III class of yield. A significant proportion of trees with light seeds are characteristic of pine swamp habitats, while their number increases in “dystrophic areas” and decreases as the ecological situation improves in other areas of the ecotope. In a number of studies, the relationship between the colour of seeds and their weight has not been revealed [
53]. However, most of the works devoted to the study of the variability of seed colouration emphasize the breeding significance of this trait and its connection with economically valuable forestry and breeding indicators of individual trees and plantings [
54]. Thus, S. Kurdiani, when studying a pine plantation in the Kiev region, found various levels of damage mainly in trees producing light seeds [
55].
In a some papers devoted to the study of the effect of seed colouration on the growth and stability of offspring, the advantage of seedlings grown from light seeds was noted [
56,
57]. Other authors, on the contrary, concluded that the best growth and stability are characteristic of seedlings grown from dark-coloured seeds. In the conditions of the Moscow region, there was an increased resistance to fusarium seedlings from black seeds [
58].
Experiments to study the influence of the conditions of origin and cultivation on the growth and preservation of offspring from colour-seed forms of Scots pine were also conducted in the forest-steppe regions of Siberia. Thus, in the south of the Krasnoyarsk Territory, ecological cultures representing the offspring of populations differentiated by seed colour were studied in arid conditions [
35,
59]. As a result of the research, differences were found in the survival rate and growth of offspring of populations with the advantage of different colour-seed groups-with black, brown, and light grey shades of the seed peel.
During the study by M.V. Rogozin, pine seeds were divided into gradations: black, dark, variegated, brown and light. In all gradations, different frequencies of the best families with intrafamily selection of phenotypically best individuals were observed, but a stable advantage was shown by variegated seeds, in which in eight experiments out of 11 the frequency was 129–227% of the norm. The most variable offspring were from light seeds with a frequency of the best families from 0 to 400%. Seeds with a dark colour germinated more slowly than variegated and brown ones, and the germination time of light seeds in different years varied from the longest (2.4 days) to the shortest (1.13 days). The author suggested that it is likely that trees with light seeds play the role of a mobile element in the gene pool and influence the adaptation of pine trees during its settlement to the south and north, which is confirmed by other studies [
60].
Other researchers found that dark seeds germinate better than light ones [
61,
62,
63], while others found no difference [
64]. Baldwin [
61] classified pine seeds (
Pinus sylvestris L.) into four colour classes: light, brown, dark and variegated. A possible reason for the different colour of pine seeds used in forestry may be that they are subjected to abrasion during industrial extraction, the worn seeds are paler [
65]. However, Steven et al. [
62] noticed that pine seeds have a naturally variable colour. They estimated the percentage of different colour classes in all major local forests as 10.2% light, 49.1% brown, 36.7% dark and 4.0% variegated. Since colour variations are widespread, the question of how this affects germination remains important.
Mukassabi et al. did not find a significant relationship between the percentage of dark, full seeds and such characteristics as the length of the cone, the number of seeds per cone, the height of the tree [
39]. However, it was found that the mass of dark seeds can be accurately predicted by the mass of random samples from 100 mixed seeds. Moreover, the number of dark seeds can be predicted by the average weight of 100 samples of dark seeds. These ratios are likely to be valid for other seed sources and are of direct practical importance for a rapid method of assessing seed germination. The study also shows that a large proportion of dark, viable seeds can be obtained from young trees.
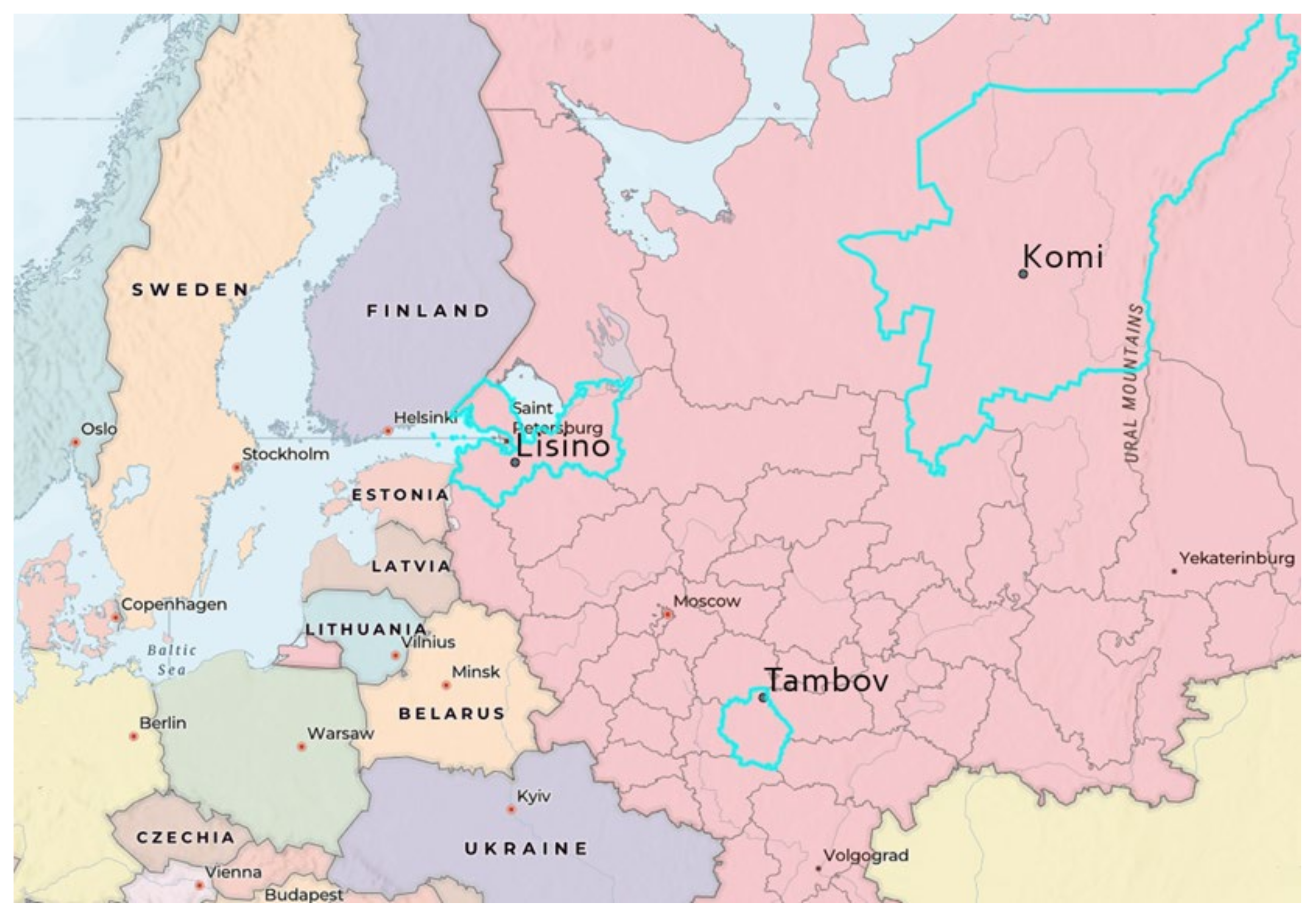
In Russia, the following practice is legally fixed: in accordance with the order of the Ministry of Natural Resources of the Russian Federation No. 909 of 9 November 2020, zoned seeds harvested within the boundaries of the municipal district should be used for forest reproduction; in their absence, seeds of forest plants—the source of origin of which is located within the territory of the forestry—should be used. Forest-seeded areas are fixed by the order of the Ministry of Natural Resources of the Russian Federation, dated 8 October 2015. All the seeds studied belong to different forest-seed areas; therefore, the justification for the use of Tambov seed material in the Leningrad region will require the creation of geographical crops and further research.
In the context of climate change, the tendency to move forest seeds from south to north and the revision of the concept of functional zoning [
66] may be necessary in the future. In 2022, it is planned to lay field experiments with Lisino’s seeds and take into account seedlings from the provenances of Komi and Tambov based on the results of wintering under snow.
In future studies, it is planned to take into account the colour of the coating as an influence parameter when stored in containers, in a controlled substrate.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







