
Phylogenetic Placement and Taxonomic Reinvestigation of Endemic and Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae) †

 ,
,  ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experiments
2.1. Plant Materials
2.2. Morphological and Anatomical Analyses
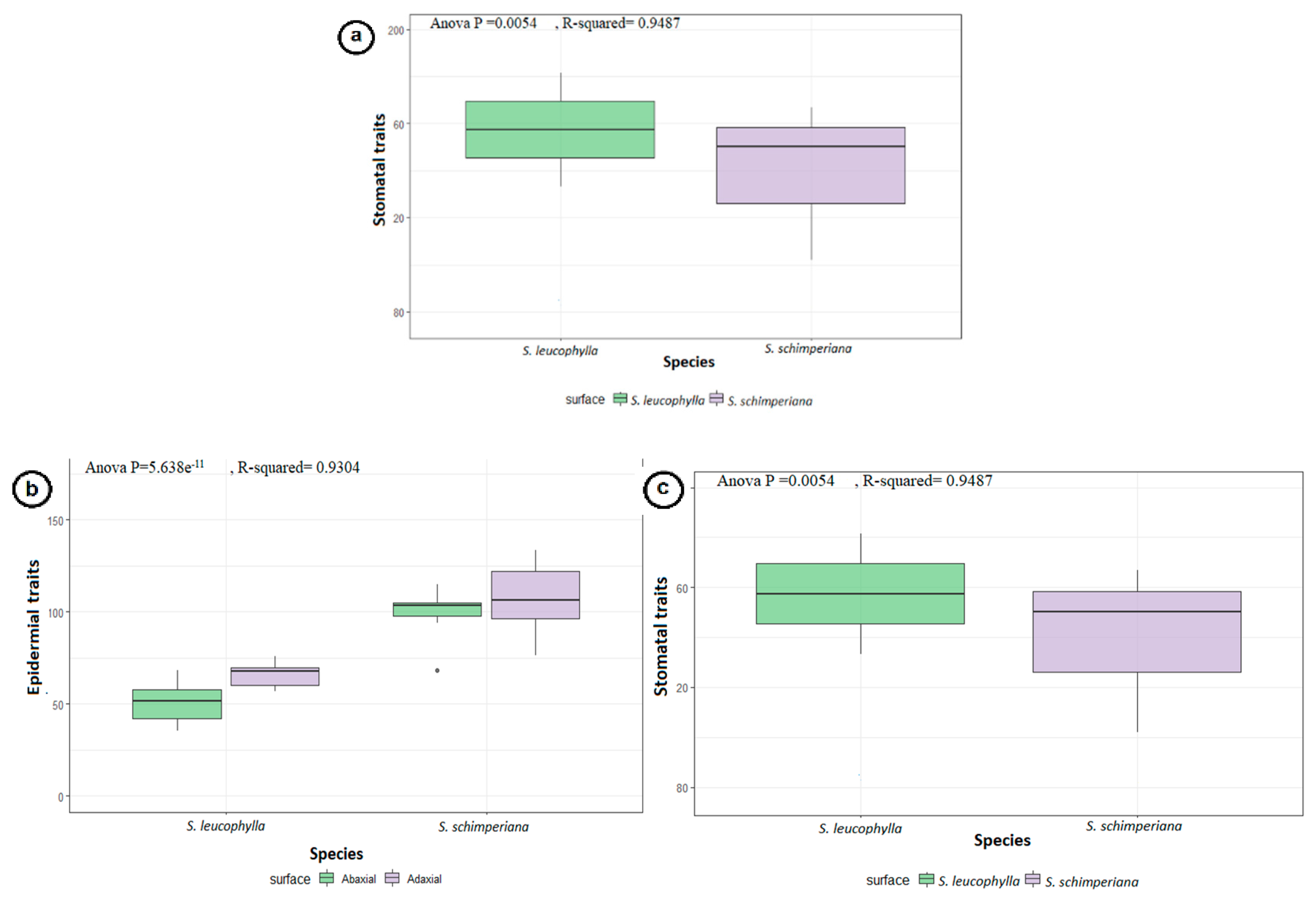
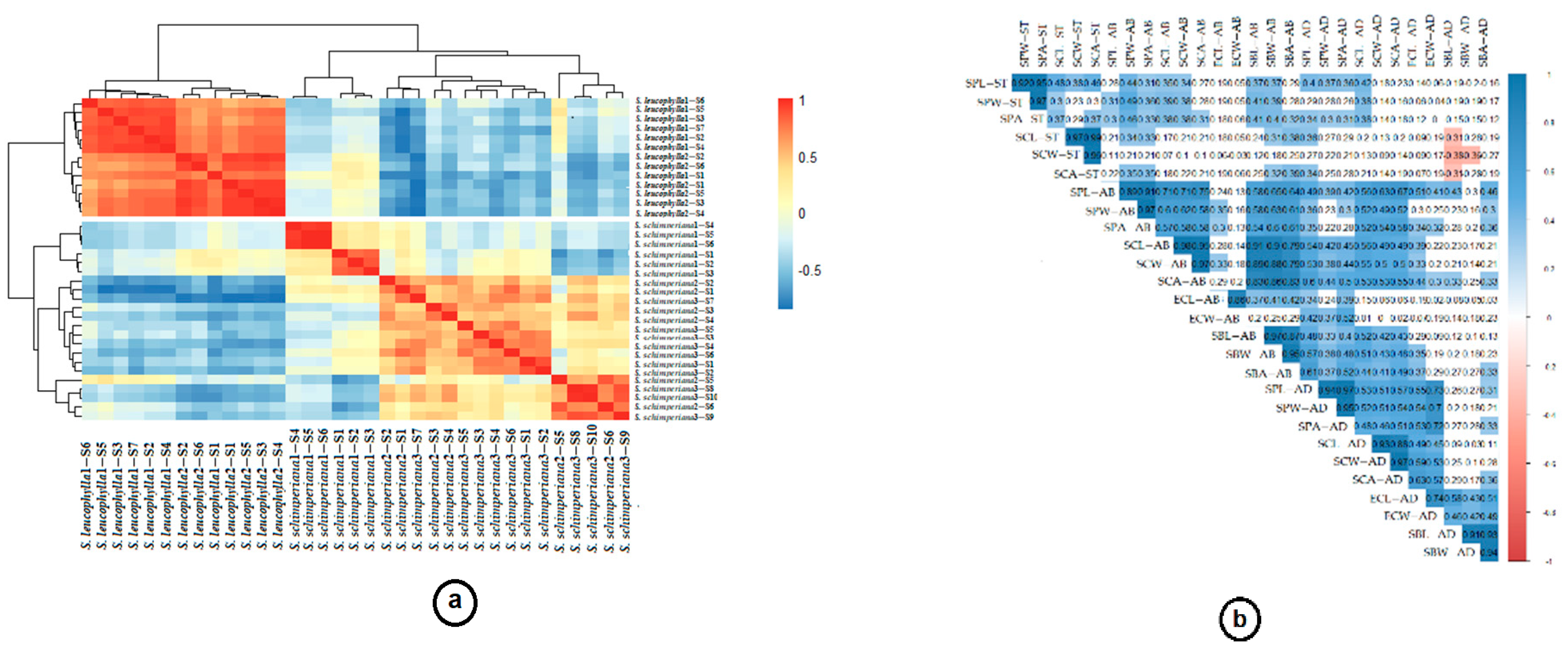
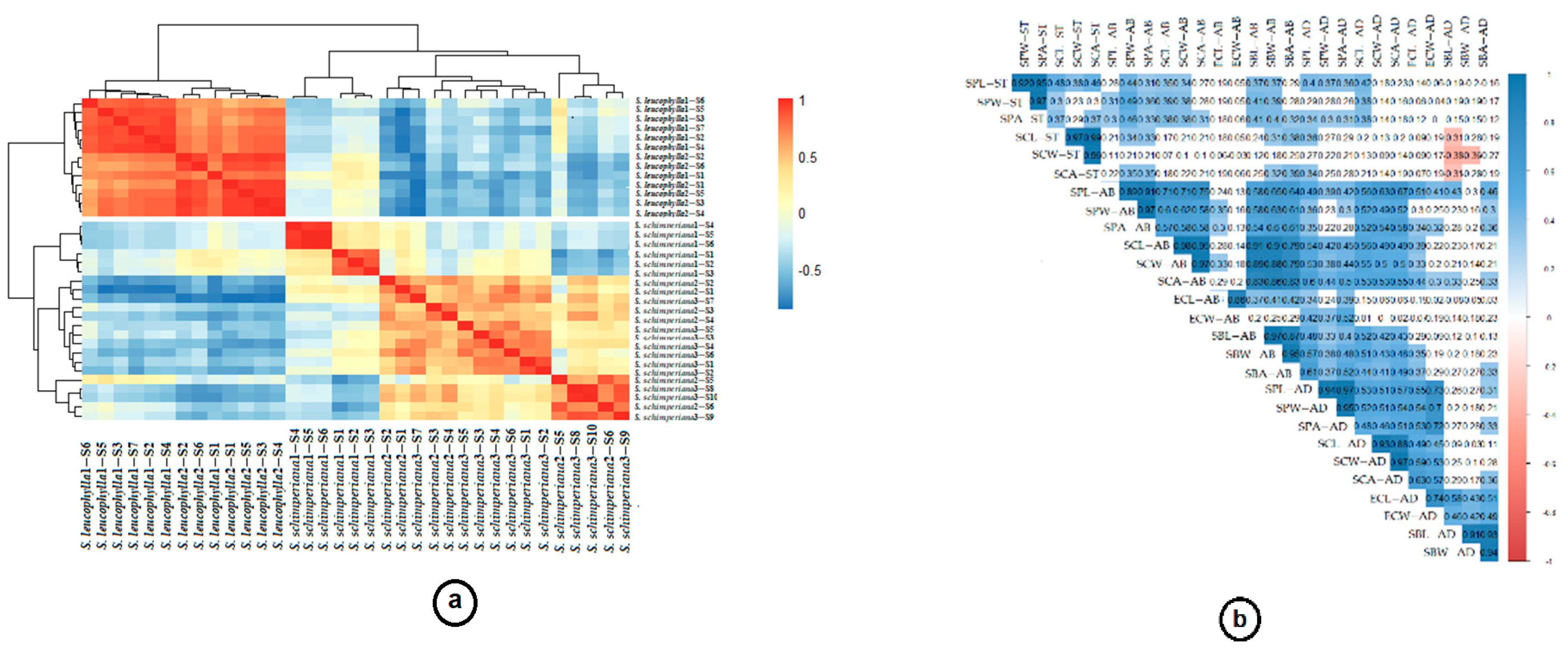
2.3. Statistical Analysis
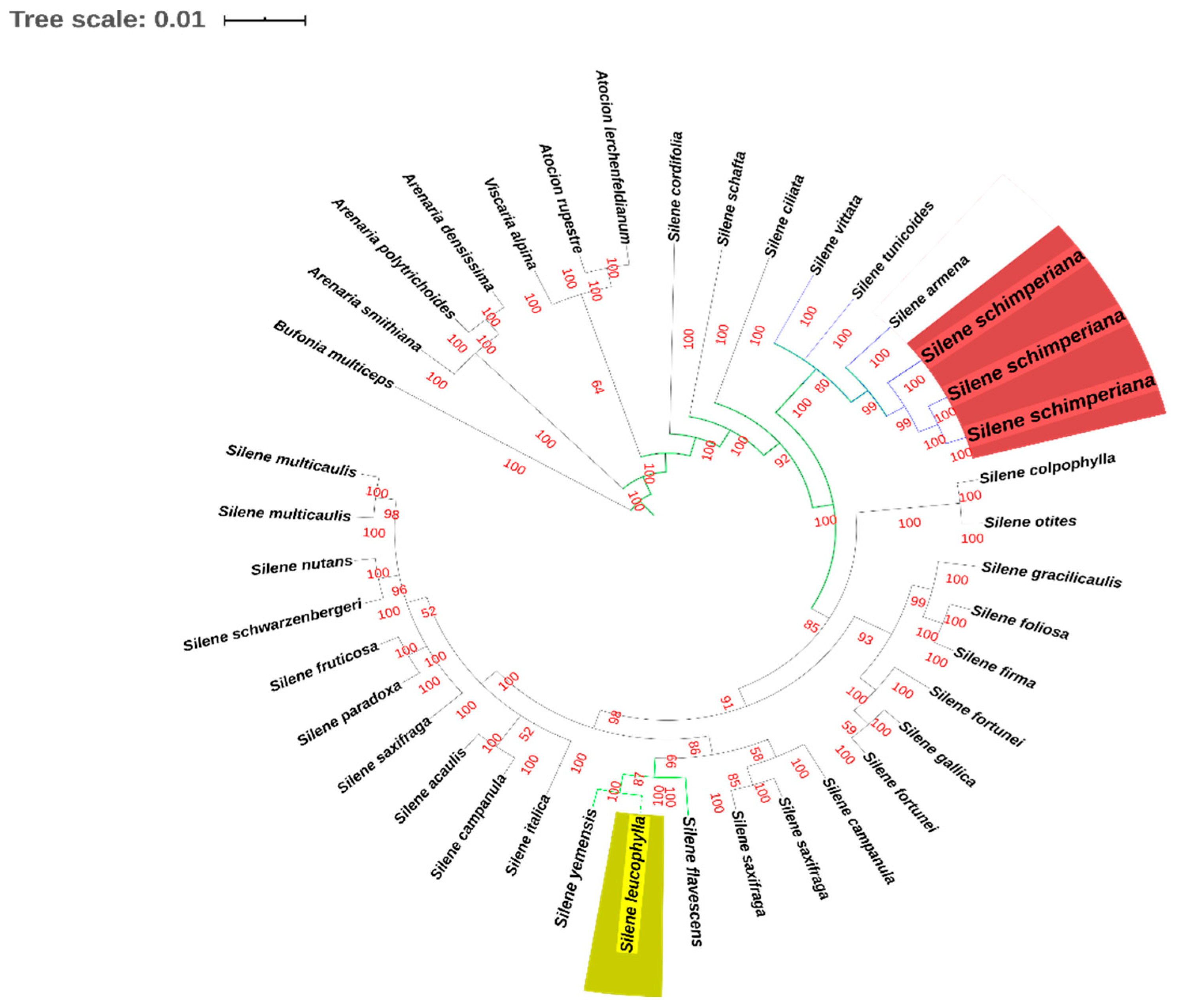
2.4. Phylogenetic Analysis
DNA Extraction, PCR Amplification, Sequencing, and Phylogenetic Analysis
3. Results
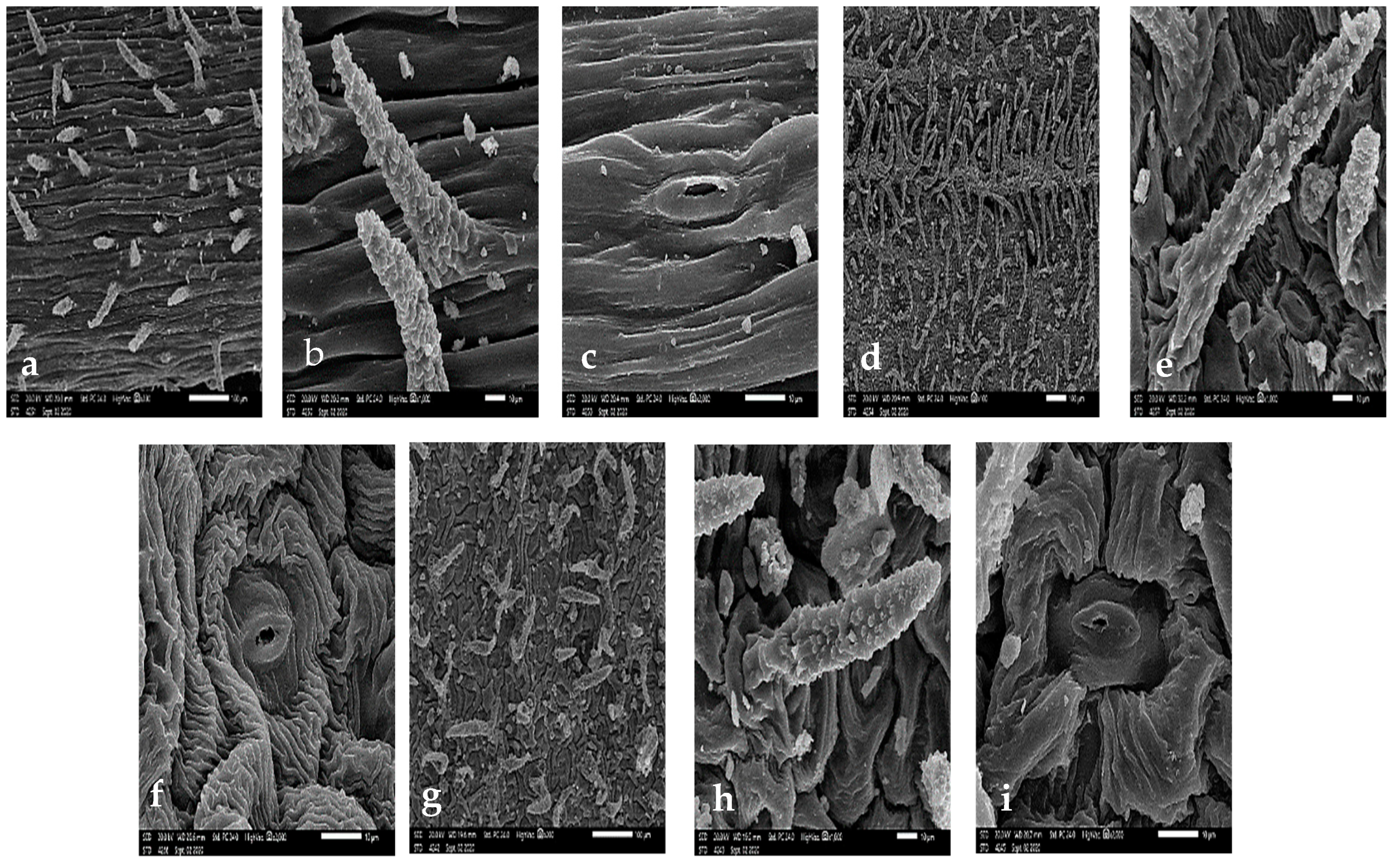
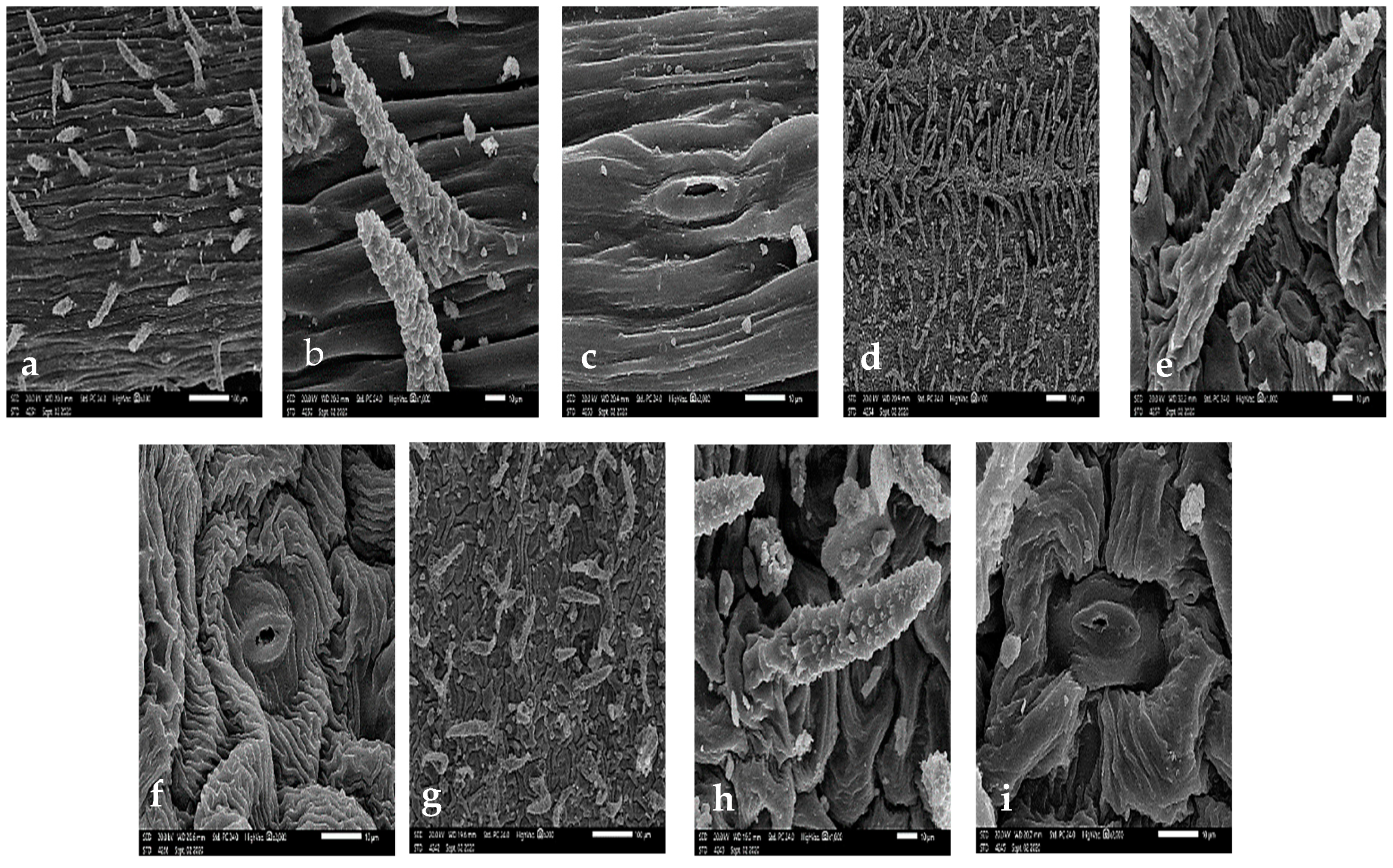
3.1. Stem Micromorphology
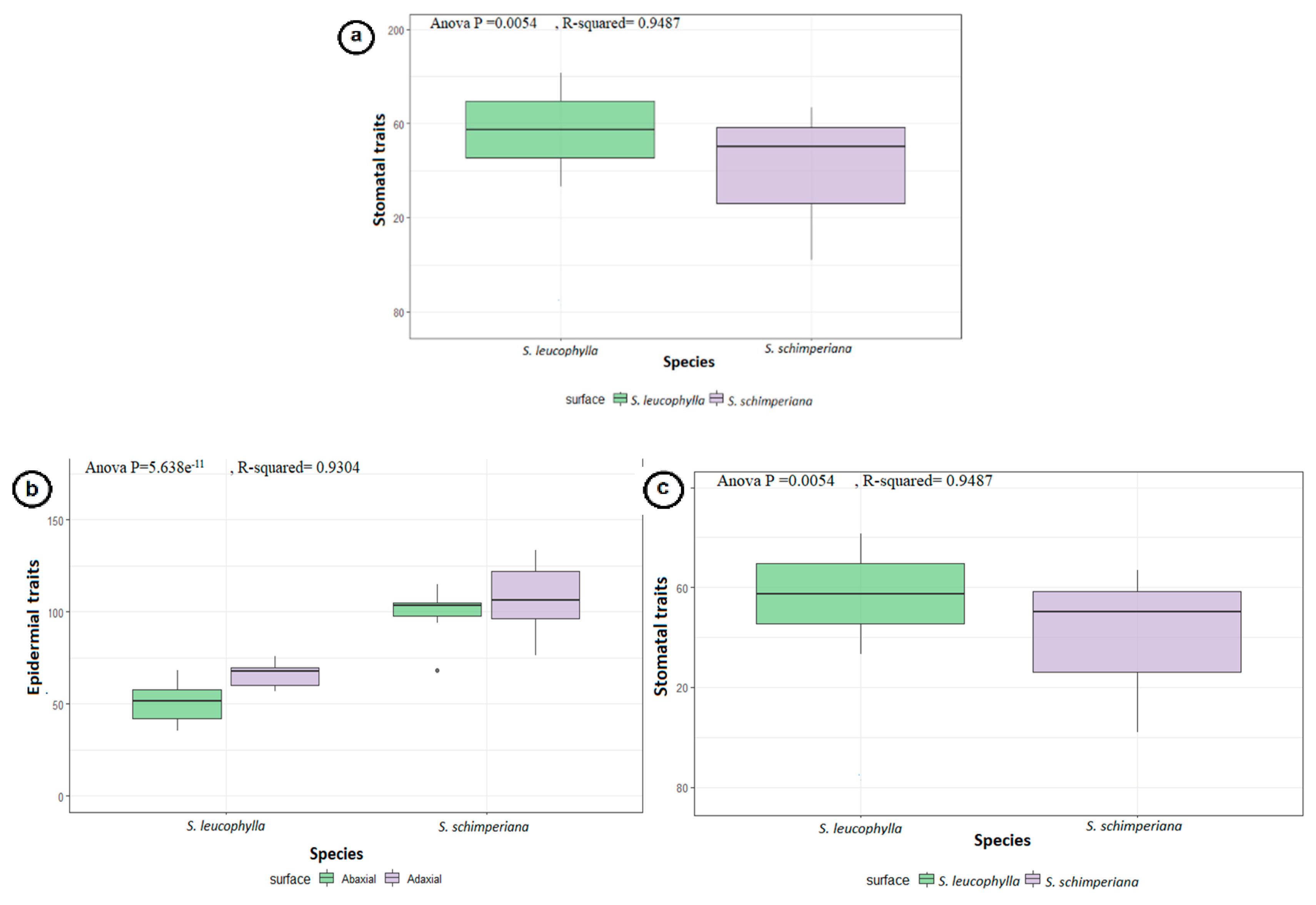
3.2. Leaf Epidermal Cells
3.3. Stomatal Complex
3.4. Phylogeny
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic Plant Species Conservation: Biotechnological Approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef] [Green Version]
- Foggi, B.; Viciani, D.; Baldini, R.M.; Carta, A.; Guidi, T. Conservation assessment of the endemic plants of the Tuscan Archipelago, Italy. Oryx 2015, 49, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Critical checklist of the endemic vascular plants of Egypt. Phytotaxa 2018, 360, 19–34. [Google Scholar] [CrossRef]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabeler, R.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Hernández-Ledesma, P.; Berendsohn, W.G.; Borsch, T.; von Mering, S.; Akhani, H.; Arias, S.; Castañeda-Noa, I.; Eggli, U.; Eriksson, R.; Flores-Olvera, H. A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales. Willdenowia 2015, 45, 281–383. [Google Scholar] [CrossRef] [Green Version]
- Radford, E.A.; Catullo, G.; Montmollin, B.D. Important Plant Areas of the South and East Mediterranean Region: Priority Sites for Conservation; IUCN/WWF: Gland, Switzerland, 2011. [Google Scholar]
- Fakhry, A.M.; El-Keblawy, A.; Shabana, H.A.; Gamal, I.E.; Shalouf, A. Microhabitats Affect Population Size and Plant Vigor of Three Critically Endangered Endemic Plants in Southern Sinai Mountains, Egypt. Land 2019, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative stress responses of some endemic plants to high altitudes by intensifying antioxidants and secondary metabolites content. Plants 2020, 9, 869. [Google Scholar] [CrossRef] [PubMed]
- Zahran, M.; Wafaa, A.; Samy, A.; Omran, G. Endemic species in Sinai Peninsula, Egypt, with particular reference to Saint Katherine protectorate: I-ecological features. J. Environ. Sci. 2015, 44, 589–609. [Google Scholar]
- Chowdhuri, P. Studies in the genus Silene. Notes from the RGB. Edinburgh 1957, 22, 221–278. [Google Scholar]
- Oxelman, B.; Rautenberg, A.; Thollesson, M.; Larsson, A.; Frajman, B.; Eggens, F.; Petri, A.; Aydin, Z.; Töpel, M.; Brandtberg-Falkman, A. Sileneae taxonomy and systematics. Cited 2013, 4, 2014. [Google Scholar]
- Hosny, A.; el Hadidi, M.; Shamso, E. Taxonomic Studies of Silenoideae (Caryophyllaceae) in Egypt. 1. Systematic revision of the genus Silene L. Taeckholmia 1992, 14, 1–36. [Google Scholar]
- Zarrinkamar, F. Foliar anatomy of the Caryophyllaceae family in Arasbaran, NW. J. Iran. Iran. J. Bot. 2001, 9, 93–102. [Google Scholar]
- Ahmad, K.; Khan, M.A.; Ahmad, M.; Zafar, M.; Arshad, M.; Ahmad, F. Taxonomic diversity of stomata in dicot flora of a district tank (NWFP) in Pakistan. Afr. J. Biotechnol. 2009, 8, 1052–1055. [Google Scholar]
- Keshavarzi, M.; Bozchaloyi, S.E. Leaf and stem comparative anatomical analysis of three genera of Alsinoideae (Caryophyllaceae). Iran. J. Bot. 2014, 20, 71–79. [Google Scholar]
- Nejati, E.M.; Ghahremaninejad, F.; Attar, F. Foliar anatomy of the genus Silene L. (Caryophyllaceae) at sectional level in Iran. Iran. J. Bot 2016, 22, 2. [Google Scholar]
- Rolland, J.; Cadotte, M.W.; Davies, J.; Devictor, V.; Lavergne, S.; Mouquet, N.; Pavoine, S.; Rodrigues, A.; Thuiller, W.; Turcati, L. Using Phylogenies in Conservation: New Perspectives; The Royal Society: London, UK, 2012. [Google Scholar]
- Rodrigues, A.S.; Gaston, K.J. Maximising phylogenetic diversity in the selection of networks of conservation areas. Biol. Conserv. 2002, 105, 103–111. [Google Scholar] [CrossRef]
- Hoveka, L.N.; van der Bank, M.; Bezeng, B.S.; Davies, T.J. Identifying biodiversity knowledge gaps for conserving South Africa’s endemic flora. Biodivers. Conserv. 2020, 29, 2803–2819. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots (Version R Package Version 0.3.0) [Computer Software]; R Development Core Team: Vienna, Austria, 2020. [Google Scholar]
- Soetewey, A. Correlation Coefficient and Correlation Test in R. 2020. Available online: https://www.statsandr.com/blog/correlation-coefficient-and-correlation-test-in-r/ (accessed on 20 February 2021).
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.R.; Zhang, Y.H.; Nakamura, K.; Guan, B.C.; Qiu, Y.X. Developing DNA barcodes for species identification in Podophylloideae (Berberidaceae). J. Syst. Evol. 2014, 52, 487–499. [Google Scholar] [CrossRef]
- Stace, C.A. Cuticular Studies as an Aid to Plant Taxonomy; Bulletin of the British Museum (Natural History); British Museum (Natural History): London, UK, 1965. [Google Scholar]
- Parkhurst, D.F. The adaptive significance of stomatal occurrence on one or both surfaces of leaves. J. Ecol. 1978, 66, 367–383. [Google Scholar] [CrossRef]
- Rossatto, D.R.; Kolb, R.M. Gochnatia polymorpha (Less.) Cabrera (Asteraceae) changes in leaf structure due to differences in light and edaphic conditions. Acta Bot. Bras. 2010, 24, 605–612. [Google Scholar] [CrossRef]
- Ullah, F.; Zafar, M.; Ahmad, M.; Shah, S.N.; Razzaq, A.; Sohail, A.; Zaman, W.; Çelik, A.; Ayaz, A.; Sultana, S. A systematic approach to the investigation of foliar epidermal anatomy of subfamily Caryophylloideae (Caryophyllaceae). Flora 2018, 246, 61–70. [Google Scholar] [CrossRef]
- Rohrbach, P. Conspectus systematicus specierum generis Silene Ann. Sci. Nat. Bot. 1868, 5, 369–382. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Banhawy, A.; Ellmouni, F.Y.; Nour, I.H.; Faried, A.; Olwey, A.O.; ElKordy, A. Phylogenetic Placement and Taxonomic Reinvestigation of Endemic and Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae). Biol. Life Sci. Forum 2021, 4, 99. https://doi.org/10.3390/IECPS2020-08619
El-Banhawy A, Ellmouni FY, Nour IH, Faried A, Olwey AO, ElKordy A. Phylogenetic Placement and Taxonomic Reinvestigation of Endemic and Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae). Biology and Life Sciences Forum. 2021; 4(1):99. https://doi.org/10.3390/IECPS2020-08619
Chicago/Turabian StyleEl-Banhawy, Ahmed, Faten Y. Ellmouni, Iman H. Nour, Ahmed Faried, Asmaa O. Olwey, and Ahmed ElKordy. 2021. "Phylogenetic Placement and Taxonomic Reinvestigation of Endemic and Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae)" Biology and Life Sciences Forum 4, no. 1: 99. https://doi.org/10.3390/IECPS2020-08619
APA StyleEl-Banhawy, A., Ellmouni, F. Y., Nour, I. H., Faried, A., Olwey, A. O., & ElKordy, A. (2021). Phylogenetic Placement and Taxonomic Reinvestigation of Endemic and Endangered Plant Species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae). Biology and Life Sciences Forum, 4(1), 99. https://doi.org/10.3390/IECPS2020-08619