
Physiological Characteristics of Expanding and Expanded Leaves of Vitis vinifera L. cv. Assyrtiko in Climate Change Conditions †

 ,
, 
Abstract
:1. Introduction
2. Experiments
2.1. Estimating Specific Leaf Area and Chlorophyll Content
2.2. In Situ Measurements of Optical Properties of Fresh Leaves
3. Results
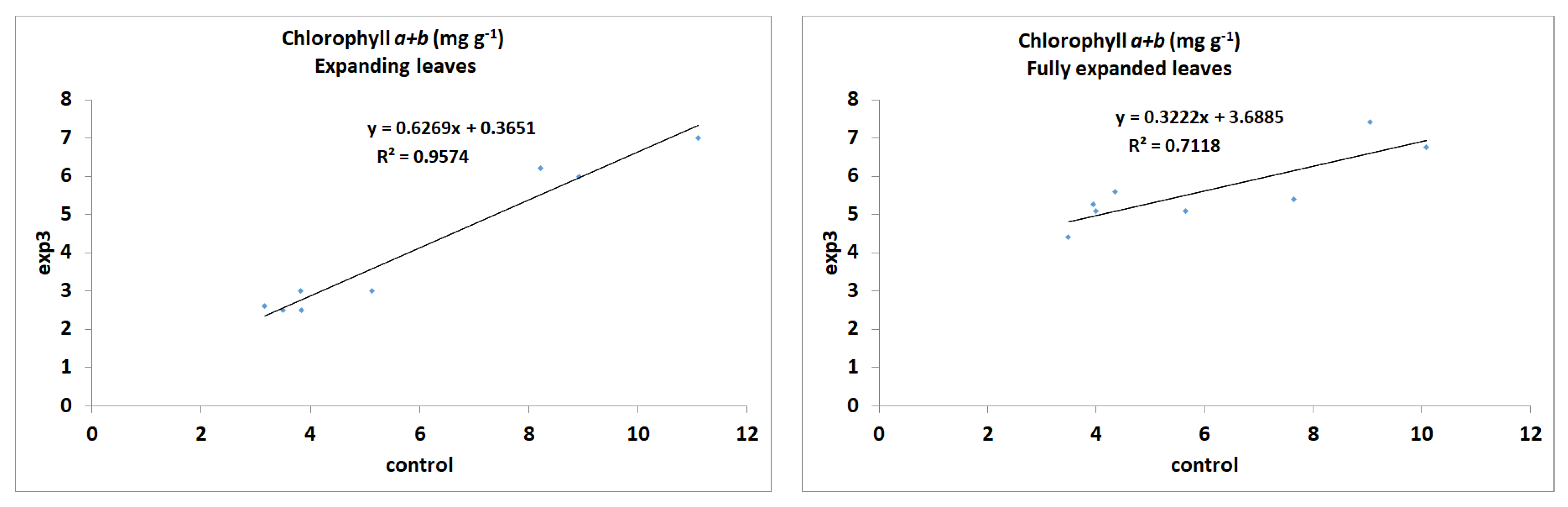
3.1. Chlorophyll Content
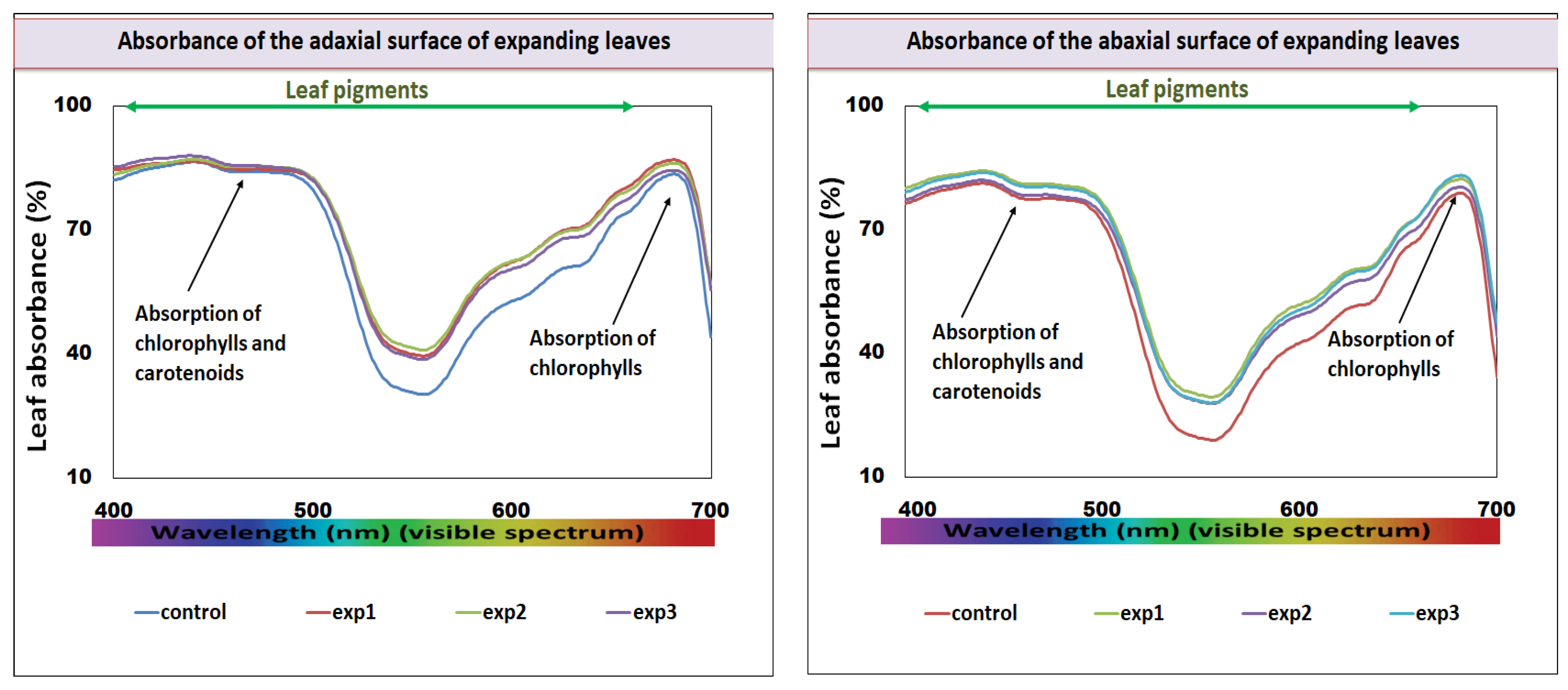
3.2. Leaf Absorbance
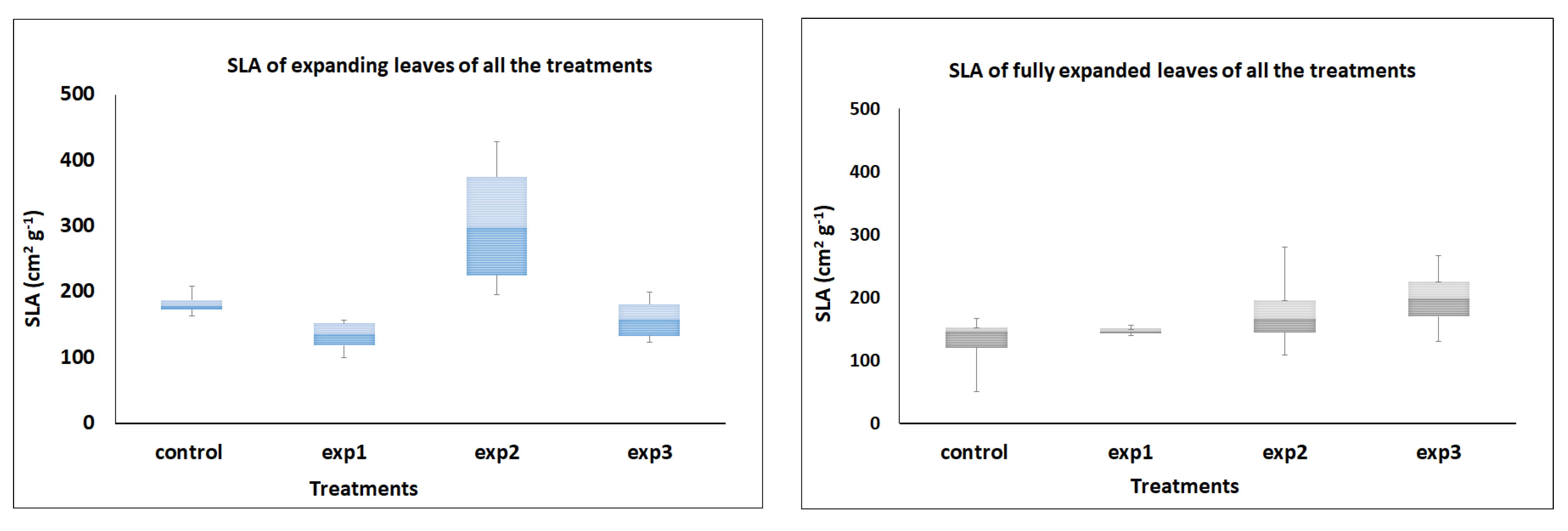
3.3. Specific Leaf Area (SLA)
4. Conclusions
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K. Intergovernmental Panel on Climate Change. In Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; DiBari, C.; Costafreda-Aumedes, S.; et al. A Review of the Potential Climate Change Impacts and Adaptation Options for European Viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Gauquelin, T.; Michon, G.; Joffre, R.; Duponnois, R.; Génin, D.; Fady, B.; Dagher-Kharrat, M.B.; Derridj, A.; Slimani, S.; Badri, W.; et al. Mediterranean forests, land use and climate change: A social-ecological perspective. Reg. Environ. Chang. 2018, 18, 623–636. [Google Scholar] [CrossRef]
- Del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Jara-Rojas, R.; Molina-Montenegro, M.A. Climate Change Impacts and Adaptation Strategies of Agriculture in Mediterranean-Climate Regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.J.; Ort, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 2001, 6, 36–42. [Google Scholar] [CrossRef]
- Vico, G.; Way, D.A.; Hurry, V.; Manzoni, S. Can leaf net photosynthesis acclimate to rising and more variable temperatures? Plant Cell Environ. 2019, 42, 1913–1928. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, G.; Nikolopoulos, D.; Klouvatou, A.; Vekkos, K.-A.; Manetas, Y.; Karabourniotis, G. The Photoprotective Role of Epidermal Anthocyanins and Surface Pubescence in Young Leaves of Grapevine (Vitis vinifera). Ann. Bot. 2006, 98, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koufos, G.; Mavromatis, T.; Koundouras, S.; Fyllas, N.M.; Jones, G.V. Viticulture-climate relationships in Greece: The impacts of recent climate trends on harvest date variation. Int. J. Clim. 2014, 34, 1445–1459. [Google Scholar] [CrossRef]
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3530–3535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponti, L.; Gutierrez, A.P.; Boggia, A.; Neteler, M. Analysis of Grape Production in the Face of Climate Change. Climate 2018, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Bertamini, M.; Zulini, L.; Muthuchelian, K.; Nedunchezhian, N. Effect of water deficit on photosynthetic and other physiological responses in grapevine (Vitis vinifera L. cv. Riesling) plants. Photosynthetica 2006, 44, 151–154. [Google Scholar] [CrossRef]
- Meletiou-Christou, M.S.; Rhizopoulou, S. Leaf functional traits of four evergreen species growing in Mediterranean environmental conditions. Acta Physiol. Plant. 2017, 39, 34. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Stratakis, E.; Ranella, A.; Farsari, M.; Fotakis, C. Laser-based micro/nanoengineering for biological applications. Prog. Quantum Electron. 2009, 33, 127–163. [Google Scholar] [CrossRef]
- De Jong, S.M.; Addink, E.A.; Doelman, J.C. Detecting leaf-water content in Mediterranean trees using high-resolution spectrometry. Int. J. Appl. Earth Obs. Geoinf. 2014, 27, 128–136. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| SLA | Treatment | p-Value |
|---|---|---|
| Expanding leaves | control & exp1 | 0.00002 |
| control & exp2 | 0.0002 | |
| control & exp3 | 0.0025 | |
| Fully expanded leaves | control & exp1 | 0.1286 |
| control & exp2 | 0.0273 | |
| control & exp3 | 0.0012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolyva, F.; Rhizopoulou, S.; Meletiou-Christou, M.-S.; Stratakis, E. Physiological Characteristics of Expanding and Expanded Leaves of Vitis vinifera L. cv. Assyrtiko in Climate Change Conditions. Biol. Life Sci. Forum 2021, 4, 55. https://doi.org/10.3390/IECPS2020-08882
Kolyva F, Rhizopoulou S, Meletiou-Christou M-S, Stratakis E. Physiological Characteristics of Expanding and Expanded Leaves of Vitis vinifera L. cv. Assyrtiko in Climate Change Conditions. Biology and Life Sciences Forum. 2021; 4(1):55. https://doi.org/10.3390/IECPS2020-08882
Chicago/Turabian StyleKolyva, Foteini, Sophia Rhizopoulou, Maria-Sonia Meletiou-Christou, and Emmanuel Stratakis. 2021. "Physiological Characteristics of Expanding and Expanded Leaves of Vitis vinifera L. cv. Assyrtiko in Climate Change Conditions" Biology and Life Sciences Forum 4, no. 1: 55. https://doi.org/10.3390/IECPS2020-08882
APA StyleKolyva, F., Rhizopoulou, S., Meletiou-Christou, M.-S., & Stratakis, E. (2021). Physiological Characteristics of Expanding and Expanded Leaves of Vitis vinifera L. cv. Assyrtiko in Climate Change Conditions. Biology and Life Sciences Forum, 4(1), 55. https://doi.org/10.3390/IECPS2020-08882