Abstract
Large freshwater mussels (Unionida) are long-lived, have large bodies, and produce thousands to millions of larvae (glochidia) that usually must attach to host fish tissue to complete their life cycle. This is an obligate parasitic stage of mussel larvae. However, less than one in onemillion find a suitable host and survive. The degree of host specificity varies among unionid species, from specialists that can successfully parasitize only one or a few closely related fish species to generalists that can complete development on a taxonomically broad range of fish species. In addition, freshwater mussels are among the most threatened groups of animals. This is due to habitat destruction, the introduction of non-native species, and the loss of host fish on which their larvae (glochidia) are obligate parasites. Glochidiosis harms fish by affecting their growth; on the other hand, freshwater mussels play an important role in freshwaters by improving water quality and ridding the water of bacteria, algae, and pollutants, they are an indicator species of water quality. During our parasitological survey of fish from the Macedonian part of Lake Prespa in April 2022, many glochidia were found on the gills, skin, and fins of two endemic fishes, Prespa roach (Rutilus prespensis) and Prespa nase (Chondrostoma prespense), in the range of tens to thousands on one host. We thus recorded these two endemic species as new hosts of A. cygnea.
1. Introduction
Lake Prespa is a high-altitude basin (approximately 850 m asl.) situated in the southwestern region of R.N. Macedonia (Figure 1). It consists of two interlinked lakes, Macro Prespa (47.4 km2) and Micro Prespa (259.4 km2), which together constitute an inner-mountainous basin that has no natural surface outflow. The Macro Prespa Lake is shared between three countries: North Macedonia, Albania, and Greece. In the past, its maximum depth was 54.2 m and the average depth was 18.74 m, a situation that has drastically changed in recent years [1]. The lake has several small rivers as tributaries. In the past, its trophic state was on the border between oligotrophic and mesotrophic. Nowadays, the surface level of the lake has significantly decreased, especially in the last decade, due to a long-term dry period, which has resulted in eutrophication. Consequentially, at present, its trophic state has decreased to eutrophic and, in the near future, has the potential to become a hyper-eutrophic ecosystem [1]. The lake wasformed in Pliocene and is the same age as Lake Ohrid. Considering the fish fauna, this lake is a typical cyprinid lake, although the trout (Salmo peristericus) live in the rivers on the side of the Pelister Mountain and penetrate the lake waters, particularly in the summer.
Figure 1.
Topographic map of the Ohrid and Prespa lakes (from Wikimedia Commons).
Unfortunately, over recent years, the entire ecosystem has faced serious environmental challenges such as pollution, ineffective planning for land and water use, and poor preservation of rare and threatened species. The unsustainable agricultural, fishery, forest, water, wastewater, and solid waste management practices have had a harsh impact on the ecosystem’s health [2].
In addition, due to the unfavorable hydrological conditions, the system lost excessive quantities of freshwater which resulted in a 9-meter water level decline over a period of 25 years. This has not only severely affected valuable shoreline habitats but has also intensified major degradation processes [2].
The pollution and eutrophication processes have not only affected the region’s valuable biodiversity but also key sectors such as tourism, water, and fisheries, all of which have been imperative in ensuring the local population’s socio-economic well-being [2].
All of these processes are believed to be responsible for today’s negative demographic trends in the region. The local population’s migrations, inside and outside the country, are an additional major concern. The authorities and the local communities are becoming increasingly aware that the restoration and maintenance of ecosystem functions are of critical importance to ensure a sustainable future for the generations to come [2].
Luckily, the international community has been generously supporting these processes. Thanks to the technical and financial support from the UNDP, the Global Environment Facility (GEF), the Swiss Development Cooperation Agency (SDC), KfW, GTZ, and others, a variety of projects in the areas of agriculture, nature conservation, solid waste management, river restoration, infrastructure development, and forest regeneration have been successfully implemented. Numerous positive initiatives were and are still being piloted to help modify the unsustainable management practices, support the region’s people with long-term economic and social development, conserve the rich biodiversity, and protect the waters of the Prespa Lake Basin [2].
The measures are based on the Watershed Management Plan’s recommendations from the UNDP project RESTORATION OF THE PRESPA LAKE ECOSYSTEM — Implementation of the Prespa Lake Watershed Management Plan, aiming at reducing the pressures from:
- -
- Agriculture (through the introduction of agro-ecological practices);
- -
- Forestry (erosion control by reforestation and control of torrents);
- -
- River pollution (wetland restoration techniques would be used for flood control and water filtering of the Golema Reka River);
- -
- Wastewaters (use of wetlands to upgrade the technology of the existing municipal wastewater treatment plant for nutrient removal);
- -
- Solid waste (upgrade of the agricultural waste management systems).
Freshwater mussels (order Unionoida) play an important role in lakes and rivers by constantly filtering water as they breathe and feed. They are filter feeders and improve water quality by filtering out bacteria, algae, organic matter, and pollutants. In the process, they accumulate contaminants in their bodies but do not break them down. They help remove excess nitrogen from water by incorporating it into their shells and tissues as they grow. Mussels move as much as 30 L of water per day through their internal filtration systems and play a key role in aquatic environments, modifying aquatic habitats as“ecosystem engineers”, and making them more suitable for themselves and other organisms. They are also an indicator of water quality since they are sensitive to excess siltation and low dissolved oxygen levels and cannot escape these unhealthy conditions. In addition, mussel shells provide an important substrate for algae and insect larvae to attach to.
Freshwater mussels represent one of the most severely endangered groups of animals due to habitat destruction, excess sedimentation, fragmentation from dams, pollution, the introduction of invasive non-native species, and, more recently, an intense drought in the southern plains which has contributed to the destruction of mussel beds. That is why mussels are very endangered; massive die-offs have occurred and their population hasdeclined by nearly 70 percent all over the world [3]. Rapid climatic and anthropogenic changes have led to the global decline of native freshwater mussels and an increased spread of those which are invasive. Efforts to maintain freshwater mussel diversity have prompted many programs for their conservation and management. One key point to such programs is the collection, maintenance, and use of mussel glochidium larvae, which require attachment to a fish host in a sensitive parasitic stage of their lifecycle. Loss of host fishes, to which their larvae (glochidia) are obligate parasites, endangers the existence of mussels as well. Understanding the thermal requirements and host-fish preferences of both native and non-native glochidia can increase knowledge of mussel larval survival and ultimately aid in the development of conservation and management programs [3].
Female mussels fertilize their eggs with sperm from a male. The fertilized eggs are brooded in the female’s gills, where they develop into tiny larvae called “glochidia”. Once mature, females may release their glochidia into the water. Large freshwater mussels (Unionida) have very long life spans, are large-bodied, and produce thousands to millions of larvae (glochidia), which typically must attach to host fish tissues (gills, fins, or skin) to complete their life cycle. It is an obligatory parasitic stage of mussel larvae to complete the reproductive process. However, fewer than one in one million find a suitable host and survive [4]. Over a few weeks to several months, the glochidia develop (metamorphose) into juvenile mussels while attached to the host fish. The degree of host specificity varies among species of unionids, from specialists able to successfully parasitize only one or a few closely related fish species to generalists which can complete development on a taxonomically wide range of fish species. The young mussel releases from the fish and falls into the sandy sediment of the river, where it will live for about 20 years.
Glochidiosis damages fish by impairing their growth, but on the other hand, freshwater mussels play an important role in freshwater by improving the water quality and filtering out bacteria, algae, and pollutants constantly. Therefore, they are an indicator species of water quality as well.
Although the relationship between glochidium larvae of freshwater mussels Unionoida and their obligatory fish hosts has been frequently described as phoretic commensalism (glochidia benefit from transportation services by fish, with no harm to fish) [5] or even symbiotic (both partners benefit [6]), increasing evidence has indicated that the relationship is parasitic. Stable isotope analysis, during the glochidial attachment to the fish host, indicates the acquirement of nutrients from the fish [7,8]. A heavy glochidia load can induce mortality of fish, indicating clear harm to the fish host [9]. When infected with glochidia, the fish host develops acquired immunity, indicating an activation of the immune system because of the attached glochidia [2,10,11]. Consequently, several adverse effects of glochidiosis on the fish host have been reported, including, for example, energetic cost and dysfunction of the liver, kidneys, and gills [12], altered behavior [13], and reduced expression of secondary sexual traits and decreased sperm quality [5]. The growth effect on brown trout can be expected to be low in natural infection intensities of less than 1000 glochidia of Margaritifera margaritifera per fish, but a lack of mortality effects is observed [14].
These negative effects of glochidia can be expected to reduce the fitness of the fish host, thereby suggesting the role of freshwater mussels (glochidia) as a selective force, emphasizing the possibility of mussel-fish coevolution. Indeed, the recent findings of population-specific adaptations of mussels to infect certain host species or local fish hosts [15,16,17] support the view of (antagonistic) co-evolution between mussels and their fish hosts. In addition, these adverse effects may turn public opinion against the restoration of disappeared mussel populations if the fish host is an important target for commercial or recreational fisheries.
The aim of this research is thus to determine the infestation of cyprinid fish from Lake Prespa with the glochidia of freshwater mussels.
2. Materials and Methods
Fish material from a total of 27 specimens of two endemic cyprinid fish species was investigated parasitologically: Prespa roach Rutilus prespensis (15 specimens) and Prespa nase Chondrostoma prespense (12 specimens) were subjected to a parasitological investigation, during spring 2022. Only fresh fish were subjected to routine identification, dissection, and observation methods. Cleaned parasites were separated and put in fixatives (70% alcohol), and prepared for determination with staining and clearing techniques [18]. For the collection of glochidia, the stereomicroscope “Zeiss Stemi 305”and microscope “Zeiss Primovert” were used. For morphological examination, permanent slides of the whole individual parasite were prepared. Identification was made throughout the morphology of glochidia [18]. The difference in the intensity of infection (mean number of glochidia per infected fish) between the R. prespensis and the C. prespense was statistically analyzed using a t-test.
3. Results and Discussion
The native distribution of the swan mussel Anodonta cygnea (Linnaeus, 1758) is European-Siberian. It was previously found in Lake Prespa [19], which coincides with the findings of our parasitological investigations of fish and the sampling of adult mussels from Lake Prespa (Figure 2).

Figure 2.
Anodonta cygnea from Macedonian part of Lake Prespa—original photo.
A parasitological examination was performed on the fish from the Macedonian part of Lake Prespa in April 2022 (Figure 3). Two endemic cyprinid fish species were investigated parasitologically: Prespa roach Rutilus prespensis (Karaman, 1924)—15 specimens and Prespa nase—Chondrostoma prespense Karaman, 1924—12 specimens. Infestation with glochidia of A. cygnea was found with nine roaches (60%) and eight nases (66.67%) (Figure 4). This is the first evidence of A. cygnea glochidia on R. prespensis and C. prespense.
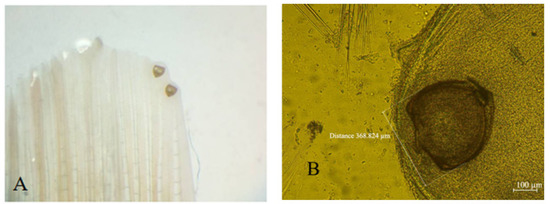
Figure 3.
Glochidia of Anodonta cygnea on the fins of Rutilus prespensis from the Macedonian part of Lake Prespa—original photos: (A)—Glochidia found on the fin of a fresh Rutilus prespensis specimen; (B)—microscopic slide of Glochidia.
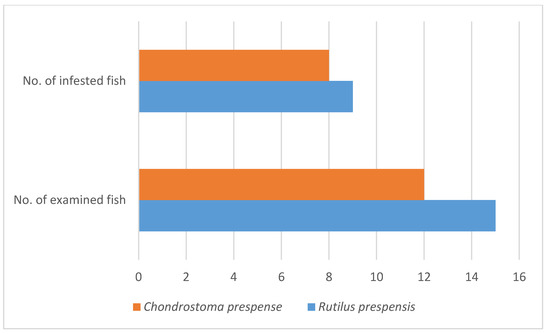
Figure 4.
Infestation of cyprinid fish Rutilus prespensis and Chondrostoma prespense from Lake Prespa with glochidia of Anodonta cygnea.
The mean (± S.E.) number of glochidia per infected fish was 40.0 ± 17.6 in the Prespa nase and 4.4 ± 0.9 in the Prespa roach. The difference was statistically marginally significant (t-test for non-equal variances, two-sided p = 0.083). This indicates that there is a trend for a higher intensity of glochidia infection in the Prespa nase than in the Prespa roach. However, further studies are needed to verify these preliminary results with respect to the possible difference between the nase and the roach in the suitability as a host of A. cygnea.
Many glochidia on the gills, skin, and fins of the two endemic fishes, Prespa roach—Rutilus prespensis (Karaman, 1924) and Prespa nase—Chondrostoma prespense (Karaman, 1924), were found, in the range of tens to thousands on a single fish host (Figure 4).
The heavy glochidia load found in our investigations induces clear harm to the fish host (Figure 3 and Figure 4).
Indeed, it is not surprising that mussel species that attach to fish for several months, growing remarkably in size during that time [14,18], activate the fish immune system [2], intake nutrients from the host [5], cause respiratory burden and damage to gills [19,20], and possibly impair the feeding efficiency [21] of the fish host, resulting in the significantly lower weight of the fish [22]. A heavy glochidia load can even induce mortality of the fish [7].
The adverse effects of glochidia parasites on a host are usually density-dependent [15]. For example, a negative effect of Margaritifera margaritifera glochidia on the critical swimming speed of brown trout increased with the intensity of infection and was evident only when it exceeded 10,000 glochidia per host [15]. Inhibition of respiration reduces the capacity of oxygen transportation to the muscle, which reduces nutrient intake due to the reduced anaerobic metabolism. In the current natural populations, the mean intensity of M. margaritifera infection in salmonids only rarely exceeds 1000 glochidia per fish. Yet, it is possible that a negative growth effect of M. margaritifera glochidia on the fish host in natural infection intensities of less than 1000 glochidia is also evident, although less prominent [15].
The record of glochidia of swan musselsin endemic fish species from the Lake Prespa, Prespa roach and Prespa nase in the present study, is considered the first in N. Macedonia. At the same time, both fish species represent new hosts for this glochidia worldwide.
The problem with introducing fish and other animal species to non-native regions of the world is that almost completely unknown species of parasites are then introduced to them. Data on non-native fish parasites are important for evaluating the health status and their general impact on native fish populations. The introduction of non-native species can also have notable effects on the populations of native species and the ecosystem and may also result in socio-economic consequences for a country due to major damage to fisheries and aquaculture.
Most parasitic organisms are in a co-evolutionary arms race with their host species. In horizontally transmitted endoparasites, this generally results in relatively low parasitic virulence for the parasites’ adult stages in the final host. Additionally, invasive species can contribute to the life cycle of native parasites. If an invasive species can serve as a suitable (e.g., intermediate, paratenic, or final) host for local parasites, these parasites may “spill back” to other local hosts which will consequently increase their infection rates within the native host populations in newly invaded habitats [23].
Globalization, the transfer and the invasion of non-indigenous species, has caused widespread biotic homogenization and the replacement of local species, resulting in a worldwide biodiversity loss. Several mechanisms, such as different environmental tolerance, higher reproduction rates, or different aggression and mutual predation, can be involved in the regulation of the competitive interactions between native and non-indigenous species. Following the replacement of the native fauna, non-indigenous species can transform habitats and even threaten entire ecosystems. They can alter ecosystem processes, causing serious problems to the environment and major economic losses. The invasion of a new habitat by a host species infected with parasites can have different effects on the local parasite fauna: (1) loss of the original parasite burden of the invader (enemy release hypothesis), (2) introduction of a new parasite species with the invader (parasite spillover), (3) the invasive species can successfully act as intermediate hosts or vectors for existing parasites or diseases (parasite spillback), and (4) loss of the local parasite species, if the invader replaces local host species but cannot act as intermediate or definitive host in the parasite life cycles (dilution effect) [24].
During the investigations that will follow, we expect to discover more data on the rate of infestation and the effect of glochidia on the health status of fish. The previously reported host fishes of A. cygnea include species such as Salmo trutta, Gobio gobio, Leuciscus idus, Scardinius erythrophthalmus, Gymnocephalus cernua, Perca fluviatilis, and Lepomis gibbosus [25]. Now, two new fish species can be added to the list of suitable hosts of A. cygnea, Prespa roach Rutilus prespensis and Prespa nase Chondrostoma prespense.
Author Contributions
Conceptualization, D.B.-D., S.S. (Stojmir Stojanovski), J.T., S.S. (Stoe Smiljkov) and B.R.; methodology, D.B.-D., S.S. (Stojmir Stojanovski), J.T., S.S. (Stoe Smiljkov) and B.R.; software, S.S. (Stojmir Stojanovski); validation, D.B.-D., S.S. (Stojmir Stojanovski), J.T. and B.R.; formal analysis, D.B.-D., S.S. (Stojmir Stojanovski), J.T. and B.R.; investigation, D.B.-D., S.S. (Stojmir Stojanovski), J.T., S.S. (Stoe Smiljkov) and B.R.; resources, S.S. (Stojmir Stojanovski) and B.R.; data curation, S.S. (Stojmir Stojanovski), J.T. and B.R.; writing—original draft preparation, D.B.-D., S.S. (Stojmir Stojanovski), J.T. and B.R.; writing—review and editing, S.S. (Stojmir Stojanovski), J.T. and B.R.; visualization, D.B.-D., S.S. (Stojmir Stojanovski), J.T., S.S. (Stoe Smiljkov) and B.R.; supervision, D.B.-D., S.S. (Stojmir Stojanovski), J.T. and S.S. (Stoe Smiljkov); project administration, S.S. (Stojmir Stojanovski); funding acquisition, S.S. (Stojmir Stojanovski); All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by PONT (Prespa Ohrid Nature Trust) “Understanding anthropogenic pressures on Lake Prespa” and the EU Co-operation in Science and Technology (COST) project Confremus (CA18239).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Data are available from the authors upon request.
Acknowledgments
We would like to thank for the grant support of PONT (PrespaOhrid Nature Trust) for the project section “Understanding anthropogenic pressures on Lake Prespa” as well as the EU Co-operation in Science and Technology (COST) project Confremus (CA18239) which enabled us to gain experience working on this research. Through CONFREMUS, Dijana Blazhekovikj-Dimovska and Stojmir Stojanovski completed their STSM at the University of Jyväskylä related on research on parasites of mussels in N. Macedonia and learning techniques of freshwater mussel parasite examination.
Conflicts of Interest
The authors declare no conflict of interest.
References and Note
- Petrovska, M.; Conevski, T.; Krstić, S.; Blinkov, I.; Stavric, V.; Cukaliev, O.; Longholt, J.; Ristovski, I.; Kočovski, Z.; Minčev, I.; et al. Prespa Lake Watershed Management Plan; Report; Ministry of Environment and Physical Planning: Skopje, North Macedonia, 2014.
- Restoration of the Prespa Lake Ecosystem-Implmention of the Prespa Lake Watershed Management Plan, UNDP Project Final Report, 2012.
- Benedict, A.; Geist, J. Early life-Cycle in Freshwater Mussel Conservation and Management; Book of Abstracts; The European Congress of Malacological Societes: Prague, Czech Republic, 2021; p. 55. [Google Scholar]
- Ćmiel, A.M.; Dołęga, J.; Aldridge, D.C.; Lipińska, A.; Tang, F.; Zając, K.; Lopes-Lima, M.; Zając, T. The size and shape of parasitic larvae of naiads (Unionidae) are not dependent on female size. Sci. Rep. 2021, 11, 23755. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.M.R.; Marjomäki, T.J.; Taskinen, J. Effect of glochidia infection on growth of fish: Freshwater pearl mussel Margaritifera margaritifera and brown trout Salmo trutta. Hydrobiologia 2019, 848, 3179–3189. [Google Scholar] [CrossRef]
- Horký, P.; Douda, K.; Maciak, M.; Závorka, L.; Slavik, O. Parasite-induced alterations of host behaviour in a riverine fish: The effects of glochidia on host dispersal. Freshw. Biol. 2014, 59, 1452–1461. [Google Scholar] [CrossRef]
- Taeubert, J.E.; Geist, J. Critical swimming speed of brown trout (Salmo trutta) infected with freshwater pearl mussel (Margaritifera margaritifera) glochidia and implications for artificial breeding of an endangered mussel species. Parasitol. Res. 2013, 112, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Douda, K.; Liu, H.-Z.; Yu, D.; Rouchet, R.; Liu, F.; Tang, Q.-Y.; Methling, C.; Smith, C.; Reichard, M. The role of local adaptation in shaping fish-mussel coevolution. Freshw. Biol. 2017, 62, 1858–1868. [Google Scholar] [CrossRef]
- Rogers-Lowery, C.L.; Dimock, R.V.; Kuhn, R.E. Antibody response of bluegill sunfish during development of acquired resistance against the larvae of the freshwater mussel Utterbackia imbecillis. Dev. Comp. Immunol. 2006, 31, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G.; Vogel, C. The parasitic stage of the fresh- water pearl mussel (Margaritifera margaritifera L.) I. Host response to Glochidiosis. Arch. Für Hydrobiol. 1987, 76, 393–402. [Google Scholar]
- Ooue, K.; Terui, A.; Urabe, H.; Nakamura, F. A delayed effect of the aquatic parasite Margaritifera laevis on the growth of the salmonid host fish Oncorhynchus masou masou. Limnology 2017, 18, 345–351. [Google Scholar] [CrossRef]
- Thomas, G.R.; Taylor, J.; de Leaniz, C.G. Does the parasitic freshwater pearl mussel M. margaritifera harm its host? Hydrobiologia 2014, 735, 191–201. [Google Scholar] [CrossRef]
- Fritts, M.W.; Fritts, A.K.; Carleton, S.A.; Bringolf, R.B. Shifts in stable-isotope signatures confirm parasitic relationship of freshwater mussel glochidia attached to host fish. J. Molluscan Stud. 2013, 79, 163–167. [Google Scholar] [CrossRef][Green Version]
- Hastie, L.C.; Young, M.R. Freshwater pearl mussel (Margaritifera margaritifera) glochidiosis in wild and farmed salmonid stocks in Scotland. Hydrobiologia 2001, 445, 109–119. [Google Scholar] [CrossRef]
- Chowdhury, M.R.; Salonen, J.K.; Marjomäki, T.J.; Taskinen, J. Interaction between the endangered freshwater pearl mussel Margaritifera margaritifera, the duck mussel Anodonta anatina and the fish host (Salmo): Acquired and cross-immunity. Hydrobiologia 2017, 810, 273–281. [Google Scholar] [CrossRef]
- Denic, M.; Taeubert, J.-E.; Geist, J. Trophic relationships between the larvae of two freshwater mussels and their fish hosts. Invertebr. Biol. 2015, 134, 129–135. [Google Scholar] [CrossRef]
- Salonen, J.K.; Taskinen, J. Electrofishing as a new method to search for unknown populations of the endangered freshwater pearl mussel Margaritifera margaritifera. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 27, 115–127. [Google Scholar] [CrossRef]
- Österling, E.M.; Ferm, J.; Piccolo, J.J. Parasitic freshwater pearl mussel larvae (Margaritifera margaritifera L.) reduce the drift-feeding rate of juvenile brown trout (Salmo trutta L.). Environ. Biol. Fishes 2014, 97, 543–549. [Google Scholar] [CrossRef]
- Albrecht, C.; Hauffe, T.; Schreiber, K.; Wilke, T. Mollusc biodiversity in a European ancient lake system: Lakes Prespa and Mikri Prespa in the Balkans. Hydrobiologia 2011, 682, 47–59. [Google Scholar] [CrossRef]
- Vasiljkov, G.V. Gelmintozi ryb. Izdateljstvo “Kolos”; Moskva: Moscow, Russia, 1983; pp. 45–50. [Google Scholar]
- Modesto, V.; Ilarri, M.; Souza, A.T.; Lopes-Lima, M.; Douda, K.; Clavero, M.; Sousa, R. Fish and mussels: Importance of fish for freshwater mussel conservation. Fish Fish. 2017, 19, 244–259. [Google Scholar] [CrossRef]
- Kekäläinen, J.; Pirhonen, J.; Taskinen, J. Do highly ornamented and less parasitized males have high quality sperm?—An experimental test for parasite-induced reproductive trade-offs in European minnow (Phoxinus phoxinus). Ecol. Evol. 2014, 4, 4237–4246. [Google Scholar] [CrossRef] [PubMed]
- Nachev, M.; Hohenadler, M.; Bröckers, N.; Grabner, D.; Sures, B. Role of Invasive Gobies for Transmission of Acanthocephalans of the Genus Pomphorhynchus in the River Rhine. 2022; in press. [Google Scholar] [CrossRef]
- Brian, J.; Aldridge, D. Endosymbionts: An overlooked threat in the conservation of freshwater mussels? Biol. Conserv. 2019, 237, 155–165. [Google Scholar] [CrossRef]
- Jansen, W.; Bauer, G.; Zahner-Meike, E. Glochidial Mortality in Freshwater Mussels. In Ecology and Evolution of the Freshwater Mussels Unionoida; Bauer, G., Wächtler, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 145, pp. 185–211. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).