
Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams


Abstract
1. Introduction
2. Materials and Methods
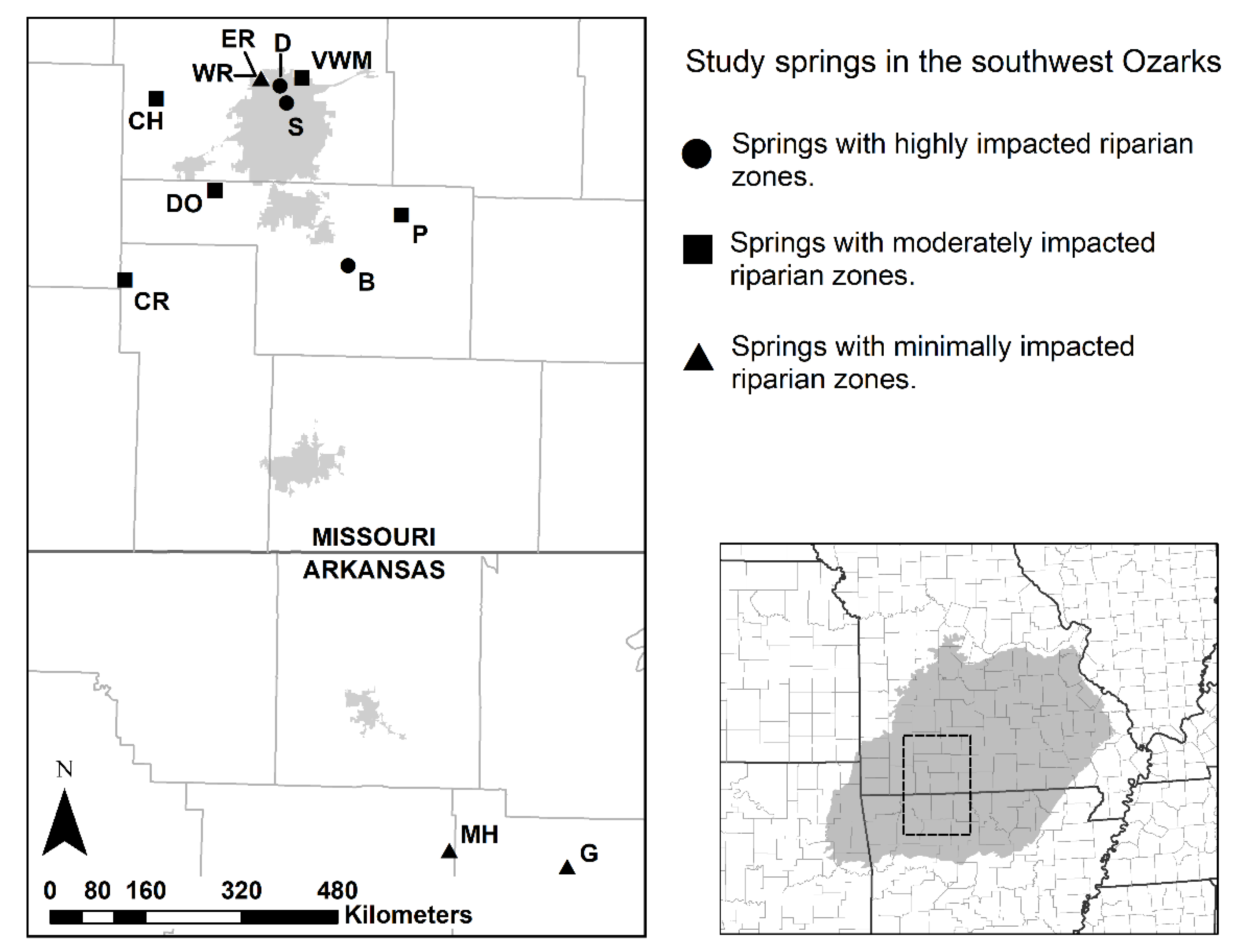
2.1. Study Sites
2.2. Data Collection
2.2.1. Environmental Variables
2.2.2. Biological Samples
2.3. Data Analysis
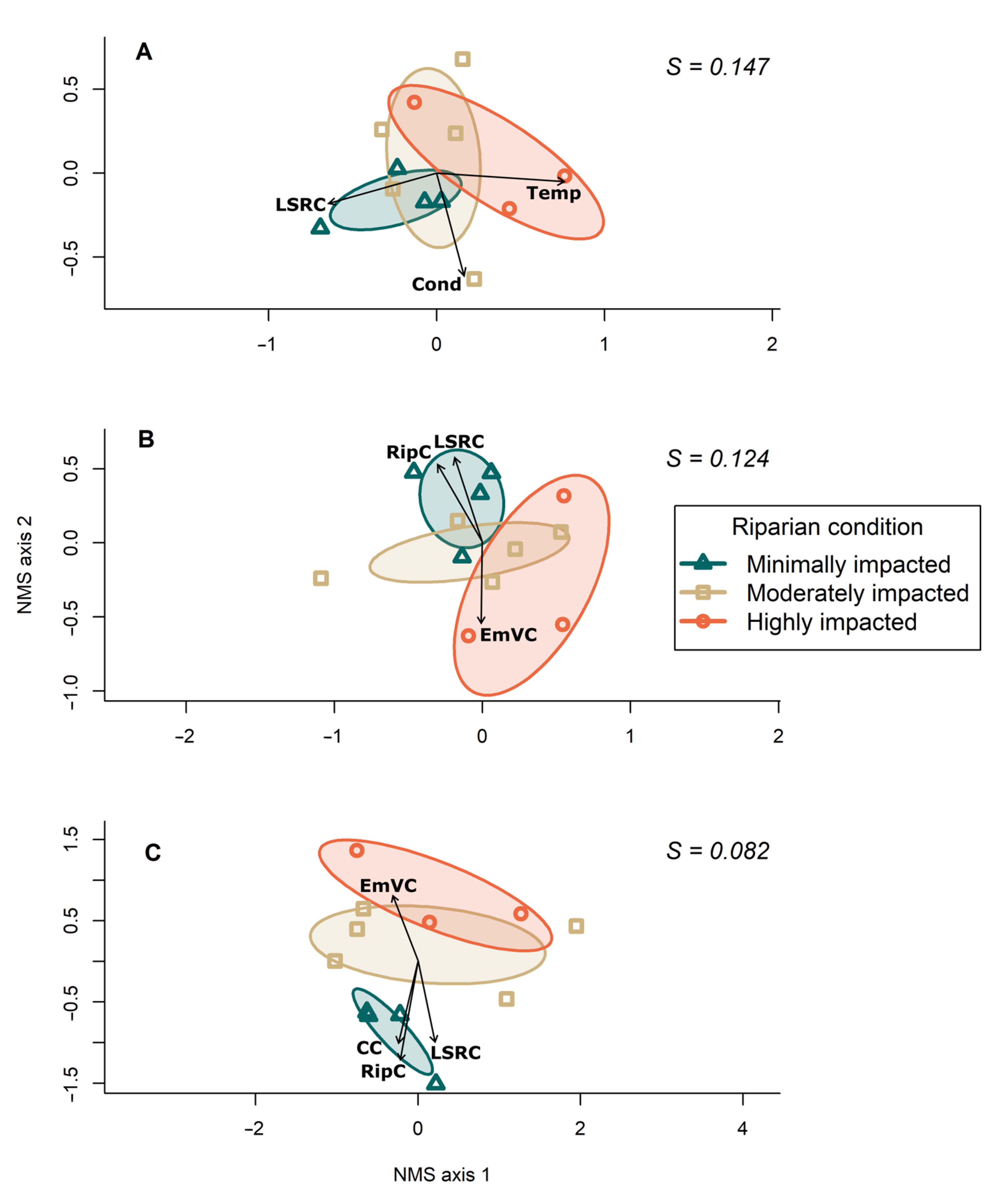
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, J.; Maser, C.; Rodiek, J. Wildlife Habitats in Managed Rangelands—The Great Basin of Southern Oregon: Riparian Zones; Pacific Northwest Forest and Range Experiment Station, US Department of Agriculture, Forest Service: Portland, OR, USA, 1979; p. 18. [Google Scholar]
- Gregory, S.V.; Swanson, F.J.; Mckee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones: Focus on links between land and water. Bioscience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Naiman, R.J.; Décamps, H. The ecology of interfaces: Riparian zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef]
- Richardson, J.S.; Danehy, R.J. A synthesis of the ecology of headwater streams and their riparian zones in temperate forests. For. Sci. 2007, 53, 131–147. [Google Scholar]
- Henley, W.F.; Patterson, M.A.; Neves, R.J.; Lemly, A.D. Effects of Sedimentation and Turbidity on Lotic Food Webs: A Concise Review for Natural Resource Managers. Rev. Fish. Sci. 2000, 8, 125–139. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Cummins, K.W.; Wilzbach, M.A.; Gates, D.M.; Perry, J.B.; Taliaferro, W.B. Shredders and Riparian Vegetation. Bioscience 1989, 39, 24–30. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols For Use in Streams And Wadeable Rivers: Periphyton, Benthic Macroinvertebrates, and Fish, 2nd ed.; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999. [Google Scholar]
- Chutter, F. An empirical biotic index of the quality of water in south African streams and rivers. Water Res. 1972, 6, 19–30. [Google Scholar] [CrossRef]
- Hilsenhoff, W.L. Use of Arthropods to Evaluate Water Quality of Streams, 100th ed.; Technical Bulletin-Wisconsin Department of Natural Resources, Division of Conservation: Madison, WI, USA, 1977; pp. 1–15. [Google Scholar]
- Zweig, L.D.; Rabeni, C.F. Biomonitoring for deposited sediment using benthic invertebrates: A test on 4 Missouri streams. J. North Am. Benthol. Soc. 2001, 20, 643–657. [Google Scholar] [CrossRef]
- Lenat, D.R.; Resh, V.H. Taxonomy and stream ecology—The benefits of genus- and species-level identifications. J. North Am. Benthol. Soc. 2001, 20, 287–298. [Google Scholar] [CrossRef]
- Daigle, J.J. Florida Damselflies (Zygoptera): A Species Key to the Aquatic Larval Stages; Technical Series; Florida Department of Environmental Regulation: Tallahassee, FL, USA, 1991; Volume 11, pp. 1–15. [Google Scholar]
- Westfall, M.J.J.; May, M.L. Damselflies of North America; Scientific Publishers: Gainsville, FL, USA, 1996. [Google Scholar]
- Landwer, B.H.P.; Sites, R.W. The Larval Odonata of Ponds in the Prairie Region of Missouri. Trans. Am. Èntomol. Soc. 2010, 136, 1–105. [Google Scholar] [CrossRef]
- Tennessen, K.J. Dragonfly Nymphs of North America: An Identification Guide; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Bowles, D.E.; Williams, M.H.; Dodd, H.R.; Morrison, L.W.; Hinsey, J.A.; Ciak, C.E.; Rowell, G.A.; DeBacker, M.D.; Haack, J. Protocol for Monitoring Aquatic Invertebrates of Small Streams in the Heartland Inventory & Monitoring Network, Version 2.1. 2021. Available online: https://irma.nps.gov/DataStore/Reference/Profile/2284622 (accessed on 15 January 2022). [CrossRef]
- Smith, J.; Samways, M.J.; Taylor, S. Assessing Riparian Quality Using Two Complementary Sets of Bioindicators. Biodivers. Conserv. 2006, 16, 2695–2713. [Google Scholar] [CrossRef]
- Villalobos-Jimenez, G.; Dunn, A.M.; Hassall, C. Dragonflies and damselflies (Odonata) in urban ecosystems: A review. Eur. J. Èntomol. 2016, 113, 217–232. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- Bybee, S.; Córdoba-Aguilar, A.; Duryea, M.C.; Futahashi, R.; Hansson, B.; Lorenzo-Carballa, M.O.; Schilder, R.; Stoks, R.; Suvorov, A.; Svensson, E.I.; et al. Odonata (dragonflies and damselflies) as a bridge between ecology and evolutionary genomics. Front. Zool. 2016, 13, 1–20. [Google Scholar] [CrossRef]
- Corbet, P.S. A Biology of Dragonflies; Witherby: London, UK, 1962. [Google Scholar]
- Rantala, M.J.; Ilmonen, J.; Koskimäki, J.; Suhonen, J.; Tynkkynen, K. The macrophyte, Stratiotes aloides, protects larvae of dragonfly Aeshna viridis against fish predation. Aquat. Ecol. 2004, 38, 77–82. [Google Scholar] [CrossRef]
- Remsburg, A.J.; Olson, A.C.; Samways, M.J. Shade Alone Reduces Adult Dragonfly (Odonata: Libellulidae) Abundance. J. Insect Behav. 2008, 21, 460–468. [Google Scholar] [CrossRef]
- Hornung, J.P.; Rice, C.L. Odonata and wetland quality in southern Alberta, Canada: A preliminary study. Odonatologica 2003, 32, 119–129. [Google Scholar]
- García-García, P.L.; Vázquez, G.; Novelo-Gutiérrez, R.; Favila, M.E. Effects of land use on larval Odonata assemblages in cloud forest streams in central Veracruz, Mexico. Hydrobiologia 2016, 785, 19–33. [Google Scholar] [CrossRef]
- Calvão, L.B.; Juen, L.; Junior, J.M.B.D.O.; Batista, J.D.; Júnior, P.D.M. Land use modifies Odonata diversity in streams of the Brazilian Cerrado. J. Insect Conserv. 2018, 22, 675–685. [Google Scholar] [CrossRef]
- Córdoba-Aguilar, A.; Rocha-Ortega, M. Damselfly (Odonata: Calopterygidae) Population Decline in an Urbanizing Watershed. J. Insect Sci. 2019, 19, 1–6. [Google Scholar] [CrossRef]
- Subramanian, K.A.; Ali, S.; Ramachandra, T.V. Odonata as indicators of riparian ecosystem health a case study from south western Karnataka, India. Fraseria 2008, 7, 83–95. [Google Scholar]
- Miguel, T.B.; Oliveira-Junior, J.M.B.; Ligeiro, R.; Juen, L. Odonata (Insecta) as a tool for the biomonitoring of environmental quality. Ecol. Indic. 2017, 81, 555–566. [Google Scholar] [CrossRef]
- Pires, M.M.; Müller, N.F.D.; Stenert, C.; Maltchik, L. Influence of different riparian vegetation widths and substrate types on the communities of larval Odonata (Insecta) in southern Brazilian streams. Acta Limnol. Bras. 2020, 32, 1–16. [Google Scholar] [CrossRef]
- Silva, L.F.R.; Castro, D.M.P.; Juen, L.; Callisto, M.; Hughes, R.M.; Hermes, M.G. A matter of suborder: Are Zygoptera and Anisoptera larvae influenced by riparian vegetation in Neotropical Savanna streams? Hydrobiologia 2021, 848, 4433–4443. [Google Scholar] [CrossRef]
- de Oliveira-Junior, J.M.B.; Junior, P.D.M.; Dias-Silva, K.; Leitão, R.P.; Leal, C.; Pompeu, P.S.; Gardner, T.A.; Hughes, R.M.; Juen, L. Effects of human disturbance and riparian conditions on Odonata (Insecta) assemblages in eastern Amazon basin streams. Limnogical 2017, 66, 31–39. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Juen, L. Structuring of dragonfly communities (Insecta: Odonata) in eastern Amazon: Effects of environmental and spatial factors in preserved and altered streams. Insects 2019, 10, 322. [Google Scholar] [CrossRef]
- de Oliveira-Junior, J.M.B.; Shimano, Y.; Gardner, T.A.; Hughes, R.M.; de Marco Júnior, P.; Juen, L. Neotropical dragonflies (Insecta: Odonata) as indicators of ecological condition of small streams in the eastern Amazon. Austral Ecol. 2015, 40, 733–744. [Google Scholar] [CrossRef]
- Sims, G.G. A Distribution of Ozark Odonates; Central Wyoming College: Riverton, WY, USA, 2015; (Personal Communication; Unpublished Data). [Google Scholar]
- Trial, L. Atlas of Missouri Odonata-Revised 2005; Missouri Department of Conservation Report: Columbia, MO, USA, 2005. [Google Scholar]
- Zeller, M.M. Comparative Analysis of Aquatic Insect, Amphipod, and Isopod Community Composition Along Environmental Gradients in Rheocrene Spring Systems of Missouri; University of Missouri: Columbia, MO, USA, 2010. [Google Scholar]
- Brönmark, C.; Herrmann, J.; Malmqvist, B.; Otto, C.; Sjöström, P. Animal community structure as a function of stream size. Hydrobiologia 1984, 112, 73–79. [Google Scholar] [CrossRef]
- Minshall, G.W.; Robinson, C.T. Macroinvertebrate community structure in relation to measures of lotic habitat heterogeneity. Arch. Für Hydrobiol. 1998, 141, 129–151. [Google Scholar] [CrossRef]
- Meinzer, O.E. Large Springs in the United States; Geological Survey Water-Supply Paper 557, Government Printing Office: Washington, DC, USA, 1927; pp. 1–94. [Google Scholar]
- Fitzpatrick, F.A.; Waite, I.R.; D’Arconte, P.J.; Meador, M.R.; Maupin, M.A.; Gurtz, M.E. Revised Methods for Characterizing Stream Habitat in the National Water-Quality Assessment Program; US Geological Survey: Reston, VA, USA, 1998; Volume 98. [Google Scholar]
- ESRI. ArcGIS Version 10. 2012. Available online: http://www.esri.com (accessed on 24 October 2019).
- Bowles, D.E.; Dodd, H.R.; Williams, M.H.; Morrison, L.W.; James, K.; DeBacker, M.D.; Ciak, C.E.; Hinsey, J.A.; Rowell, G.A.; Haack, J.L. Protocol for Monitoring Spring Communities at Ozark National Scenic Riverways, Missouri; National Park Service: Fort Collins, CO, USA, 2008. [Google Scholar]
- Chovanec, A.; Waringer, J. Ecological integrity of river-floodplain systems? assessment by dragonfly surveys (Insecta: Odonata). Regul. Rivers: Res. Manag. 2001, 17, 493–507. [Google Scholar] [CrossRef]
- Moulton II, S.R.; Kennen, J.G.; Goldstein, R.M.; Hambrook, J.A. Revised Protocols for Sampling Algal, Invertebrate, and Fish Communities as Part of the National Water-Quality Assessment Program; Geological Survey (US): Reston, VA, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, L.G.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-5. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 8 March 2019).
- Oksanen, J. Vegan: An Introduction to Ordination. 2015. Available online: http://cran.r-project.org/web/packages/vegan/vignettes/introvegan.pdf (accessed on 8 March 2019).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.A. The effect of habitat structure on the spatial distribution of freshwater invertebrate populations. Habitat Struct. Phys. Arrange. Objects Space 1991, 8, 87–106. [Google Scholar] [CrossRef]
- Brito, J.S.; Michelan, T.S.; Juen, L. Aquatic macrophytes are important substrates for Libellulidae (Odonata) larvae and adults. Limnology 2020, 22, 139–149. [Google Scholar] [CrossRef]
- Wijnhoven, S.; Van Riel, M.; Van Der Velde, G. Exotic and indigenous freshwater gammarid species: Physiological tolerance to water temperature in relation to ionic content of the water. Aquat. Ecol. 2003, 37, 151–158. [Google Scholar] [CrossRef]
- Sornom, P.; Felten, V.; Médoc, V.; Sroda, S.; Rousselle, P.; Beisel, J.-N. Effect of gender on physiological and behavioural responses of Gammarus roeseli (Crustacea Amphipoda) to salinity and temperature. Environ. Pollut. 2010, 158, 1288–1295. [Google Scholar] [CrossRef]
- Clements, W.H.; Kotalik, C. Effects of major ions on natural benthic communities: An experimental assessment of the US Environmental Protection Agency aquatic life benchmark for conductivity. Freshw. Sci. 2016, 35, 126–138. [Google Scholar] [CrossRef]
- Davis, R.K.; Brahana, J.V.; Johnston, J.S. Ground Water in Northwest Arkansas: Minimizing Nutrient Contamination from Non-Point Sources in Karst Terrane; Arkansas Water Resources Center: Fayetteville, AR, USA, 2000; pp. 1–69. [Google Scholar]
- Peterson, E.W.; Davis, R.K.; Brahana, J.; Orndorff, H.A. Movement of nitrate through regolith covered karst terrane, northwest Arkansas. J. Hydrol. 2002, 256, 35–47. [Google Scholar] [CrossRef]
- Carroll, T.M.; Thorp, J.H. Ecotonal shifts in diversity and functional traits in zoobenthic communities of karst springs. Hydrobiologia 2014, 738, 1–20. [Google Scholar] [CrossRef]
- Bick, G.H.; Bick, J.C. Substrate utilization during reproduction by Argia plana Calvert and Argia moesta (Hagen) (Odonata: Coenagrionidae). Odonatologica 1972, 1, 3–9. [Google Scholar]
- Alam, K.; Negishi, J.N.; Rahman, M.A.T.; Tolod, J.R. Stable isotope ratios of emergent adult aquatic insects can be used as indicators of water pollution in the hyporheic food web. Ecol. Indic. 2020, 118, 106738. [Google Scholar] [CrossRef]
- Kelly, D.W.; Dick, J.T.A.; Montgomery, W.I. The functional role of Gammarus (Crustacea, Amphipoda): Shredders, predators, or both? Hydrobiologia 2002, 485, 199–203. [Google Scholar] [CrossRef]
- Connell, J.H. The Influence of Interspecific Competition and Other Factors on the Distribution of the Barnacle Chthamalus Stellatus. Ecology 1961, 42, 710–723. [Google Scholar] [CrossRef]
- Flory, E.A.; Milner, A.M. Macroinvertebrate community succession in Wolf Point Creek, Glacier Bay National Park, Alaska. Freshw. Biol. 2000, 44, 465–480. [Google Scholar] [CrossRef]
- Madsen, P.B.; Morabowen, A.; Andino, P.; Espinosa, R.; Cauvy-Fraunié, S.; Dangles, O.; Jacobsen, D. Altitudinal distribution limits of aquatic macroinvertebrates: An experimental test in a tropical alpine stream. Ecol. Èntomol. 2015, 40, 629–638. [Google Scholar] [CrossRef]
- Yuan, L.L. Estimation and Application of Macroinvertebrate Tolerance Values. US EPA, ORD; National Center for Environmental Assessment: Washington, DC, USA, 2006. [Google Scholar]
- Huggins, D.G.; Moffett, M. Proposed Biotic and Habitat Indices for Use in Kansas Streams: Report No. 35 of the Kansas Biological Survey; Kansas Biological Survey: Lawrence, KS, USA, 1988. [Google Scholar]
- Lenat, D.R. A Biotic Index for the Southeastern United States: Derivation and List of Tolerance Values, with Criteria for Assigning Water-Quality Ratings. J. North Am. Benthol. Soc. 1993, 12, 279–290. [Google Scholar] [CrossRef]
- Hilsenhoff, W.L. An improved biotic index of organic stream pollution. Gt. Lakes Entomol. 1987, 20, 7. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Stream | Stream Code | Spring Magnitude | Source to Reach (m) | Reach Length (m) | RipC Width (m) | Riparian Impactedness |
|---|---|---|---|---|---|---|
| West Ritter | WR | 3rd | 16.5 | 68 | 13.6 | Minimal |
| Valley Water Mill | VWM | 3rd | 43.0 | 124 | 24.8 | Moderate |
| Silver | S | 3rd | 0.0 | 41 | 17.2 | High |
| Patterson | P | 3rd | 72.0 | 68 | 13.6 | Moderate |
| Chiles | CH | 3rd | 0.0 | 33 | 10.6 | Moderate |
| East Ritter | ER | 3rd | 260.0 | 62 | 12.4 | Minimal |
| Doling | D | 4th | 180.0 | 34 | 6.8 | High |
| Mitch Hill | MH | 3rd | 0.0 | 82 | 16.4 | Minimal |
| Gilbert | G | 3rd | 11.0 | 74 | 14.8 | Minimal |
| Double | DO | 3rd | 0.0 | 56 | 11.2 | Moderate |
| Brown | B | 3rd | 140.0 | 54 | 10.8 | High |
| Crane Creek | CR | 4th | 0.0 | 24 | 4.8 | Moderate |
| Species | WR | VWM | S | P | CH | ER | D | MH | G | DO | B | CR | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anisoptera | |||||||||||||
| Aeshna umbrosa Walker, 1908 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16 A | 0 | 0 | 0 | 0 | 16 |
| Anax junius Drury, 1773 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Basiaeschna janata Say, 1840 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 7 |
| Boyeria vinosa Say, 1840 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 4 |
| Cordulegaster obliqua Say, 1840 | 14 | 0 | 0 | 0 | 0 | 2 A | 0 | 9 | 8 | 0 | 0 | 0 | 33 |
| Epitheca cyanosura Say, 1840 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Erythemis simplicicollis Say, 1840 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 1 A | 0 | 4 |
| Gomphurus sp. Needham, 1901 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Libellula incesta Hagen, 1861 | 0 | 1 E | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Somatochlora tenebrosa Say, 1840 | 0 | 1 | 0 | 0 | 0 | 11 A | 0 | 2 | 0 | 0 | 0 | 0 | 14 |
| Sympetrum corruptum Hagen, 1861 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Zygoptera | |||||||||||||
| Argia funebris Hagen, 1861 | 1 A | 0 | 244 A | 2 A | 0 | 0 | 85 A | 2 A | 0 | 51 A | 0 | 48 A | 433 |
| Argia translata Hagen in Selys, 1865 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 6 |
| Calopteryx maculata Palisot de Beauvois, 1807 | 19 A | 33 A | 0 | 24 A | 0 | 42 A | 18 A | 18 | 28 A | 10 A | 2 A | 2 A | 196 |
| Enallagma basidens Calvert, 1902 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 |
| Enallagma divagans Selys, 1876 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Enallagma geminatum Kellicott, 1895 | 0 | 6 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 |
| Enallagma signatum Hagen, 1861 | 0 | 3 | 0 | 0 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19 |
| Enallagma vesperum Calvert, 1919 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Ischnura hastata Say, 1840 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| Ischnura posita Hagen, 1861 | 0 | 13 | 0 | 0 | 22 | 0 | 5 | 0 | 0 | 0 | 43 | 0 | 83 |
| Ischnura verticalis Say, 1840 | 0 | 2 | 0 | 0 | 18 A | 0 | 0 | 0 | 0 | 0 | 2 A | 0 | 22 |
| Total | 34 | 87 | 244 | 26 | 71 | 55 | 114 | 61 | 36 | 61 | 48 | 50 |
| Stream | Q (m3/s) | pH | ORP (mV) | DO (%) | Cond (µS/cm) | TDS (mg/L) | Temp (°C) | NO3− (mg/L) | PO43− (mg/L) | Embed (%) | D50 (mm) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| West Ritter | 0.107 | 6.93 | 261.7 | 73.2 | 586 | 293 | 14.99 | 13.2 | 0.08 | 75 | 38.5 |
| Valley Water Mill | 0.082 | 6.88 | 277.7 | 107.1 | 596 | 298 | 16.50 | 8.8 | 0.00 | 50 | 46.5 |
| Silver | 0.030 | 6.93 | 308.3 | 70.1 | 499 | 250 | 17.37 | 8.8 | 0.04 | 100 | 27.3 |
| Patterson | 0.170 | 7.16 | 246.2 | 73.2 | 316 | 158 | 14.07 | 4.4 | 0.00 | 25 | 54.5 |
| Chiles | 0.029 | 7.03 | 313.9 | 72.9 | 434 | 217 | 14.54 | 17.6 | 0.14 | 75 | 54.5 |
| East Ritter | 0.039 | 7.05 | 204.4 | 52.9 | 616 | 308 | 16.24 | 13.2 | 0.04 | 75 | 38.5 |
| Doling | 0.006 | 7.59 | 231.7 | 56.8 | 594 | 297 | 26.63 | 8.8 | 0.00 | 100 | 77.0 |
| Mitch Hill | 0.100 | 6.77 | 328.7 | 62.1 | 457 | 229 | 15.73 | 13.2 | 0.00 | 75 | 54.5 |
| Gilbert | 0.033 | 7.47 | 264.5 | 84.3 | 317 | 159 | 18.27 | 8.8 | 0.06 | 100 | 38.5 |
| Double | 0.060 | 7.16 | 275.2 | 70.6 | 357 | 178 | 17.29 | 17.6 | 0.04 | 50 | 54.5 |
| Brown | 0.053 | 7.17 | 265.8 | 79.0 | 448 | 224 | 16.00 | 22.0 | 0.02 | 50 | 46.5 |
| Crane Creek | 0.009 | 6.86 | 321.1 | 58.9 | 405 | 203 | 14.96 | 17.6 | 0.04 | 75 | 38.5 |
| Stream | RipC (% Undisturbed) | LSRC (% Undisturbed) | CC (% Cover) | EmVC (% Cover) |
|---|---|---|---|---|
| West Ritter | 95 | 78 | 75 | 20 |
| Valley Water Mill | 65 | 65 | 50 | 70 |
| Silver | 0 | 0 | 0 | 90 |
| Patterson | 45 | 60 | 50 | 50 |
| Chiles | 40 | 50 | 25 | 20 |
| East Ritter | 95 | 80 | 50 | 50 |
| Doling | 20 | 15 | 50 | 35 |
| Mitch Hill | 85 | 100 | 50 | 10 |
| Gilbert | 100 | 75 | 100 | 0 |
| Double | 50 | 10 | 25 | 90 |
| Brown | 10 | 80 | 0 | 35 |
| Crane Creek | 40 | 75 | 0 | 60 |
| Fitted Vector | NMS A r | NMS A p Value | NMS B r | NMS B p Value | NMS C r | NMS C p Value |
|---|---|---|---|---|---|---|
| RipC | 0.39 | 0.105 | 0.54 | 0.0292 * | 0.82 | 0.0003 *** |
| LSRC | 0.46 | 0.0616 · | 0.53 | 0.0322 * | 0.56 | 0.0277 * |
| CC | 0.26 | 0.250 | 0.24 | 0.280 | 0.58 | 0.0203 * |
| EmVC | 0.24 | 0.284 | 0.43 | 0.0772 · | 0.41 | 0.0965 · |
| Temp | 0.60 | 0.0044 ** | 0.32 | 0.175 | 0.036 | 0.896 |
| Cond | 0.41 | 0.0898 · | 0.27 | 0.241 | 0.091 | 0.641 |
| Odonate Taxa | rpb | p Value | Impactedness |
|---|---|---|---|
| Cordulegaster obliqua | 0.84 | 0.0071** | Minimally impacted |
| Argia funebris | 0.61 | 0.048* | Highly impacted |
| Calopteryx maculata | 0.58 | 0.14 | Minimally impacted |
| Enallagma geminatum | 0.54 | 0.28 | Moderately impacted |
| Argia translata | 0.50 | 0.24 | Highly impacted |
| Somatochlora tenebrosa | 0.49 | 0.15 | Minimally impacted |
| Enallagma signatum | 0.44 | 0.48 | Moderately impacted |
| Ischnura posita | 0.42 | 0.32 | Highly impacted |
| Aeshna umbrosa | 0.42 | 0.57 | Minimally impacted |
| Boyeria vinosa | 0.42 | 0.57 | Minimally impacted |
| Ischnura verticalis | 0.38 | 0.73 | Moderately impacted |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheri, C.R.; Finn, D.S. Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams. Hydrobiology 2023, 2, 260-276. https://doi.org/10.3390/hydrobiology2010017
Cheri CR, Finn DS. Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams. Hydrobiology. 2023; 2(1):260-276. https://doi.org/10.3390/hydrobiology2010017
Chicago/Turabian StyleCheri, Cameron R., and Debra S. Finn. 2023. "Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams" Hydrobiology 2, no. 1: 260-276. https://doi.org/10.3390/hydrobiology2010017
APA StyleCheri, C. R., & Finn, D. S. (2023). Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams. Hydrobiology, 2(1), 260-276. https://doi.org/10.3390/hydrobiology2010017