
Using Fish Assemblages to Assess the Ecological Effects of Marine Protection on Rocky Habitats in a Portuguese Natural Park

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
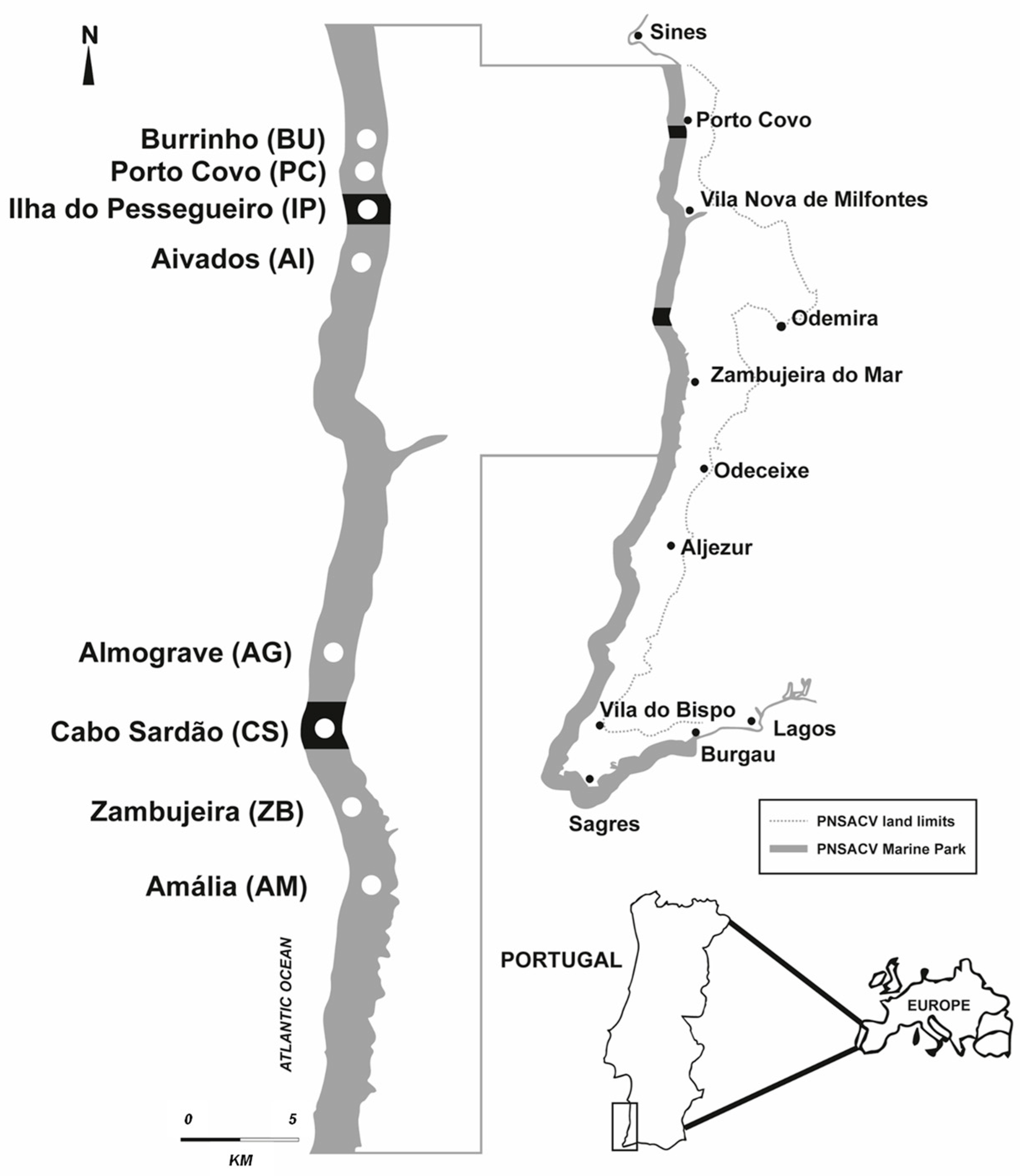
2.1. Study Area
2.2. Underwater Visual Censuses (UVC)
2.3. Substrate Characterization
2.4. Statistical Analyses
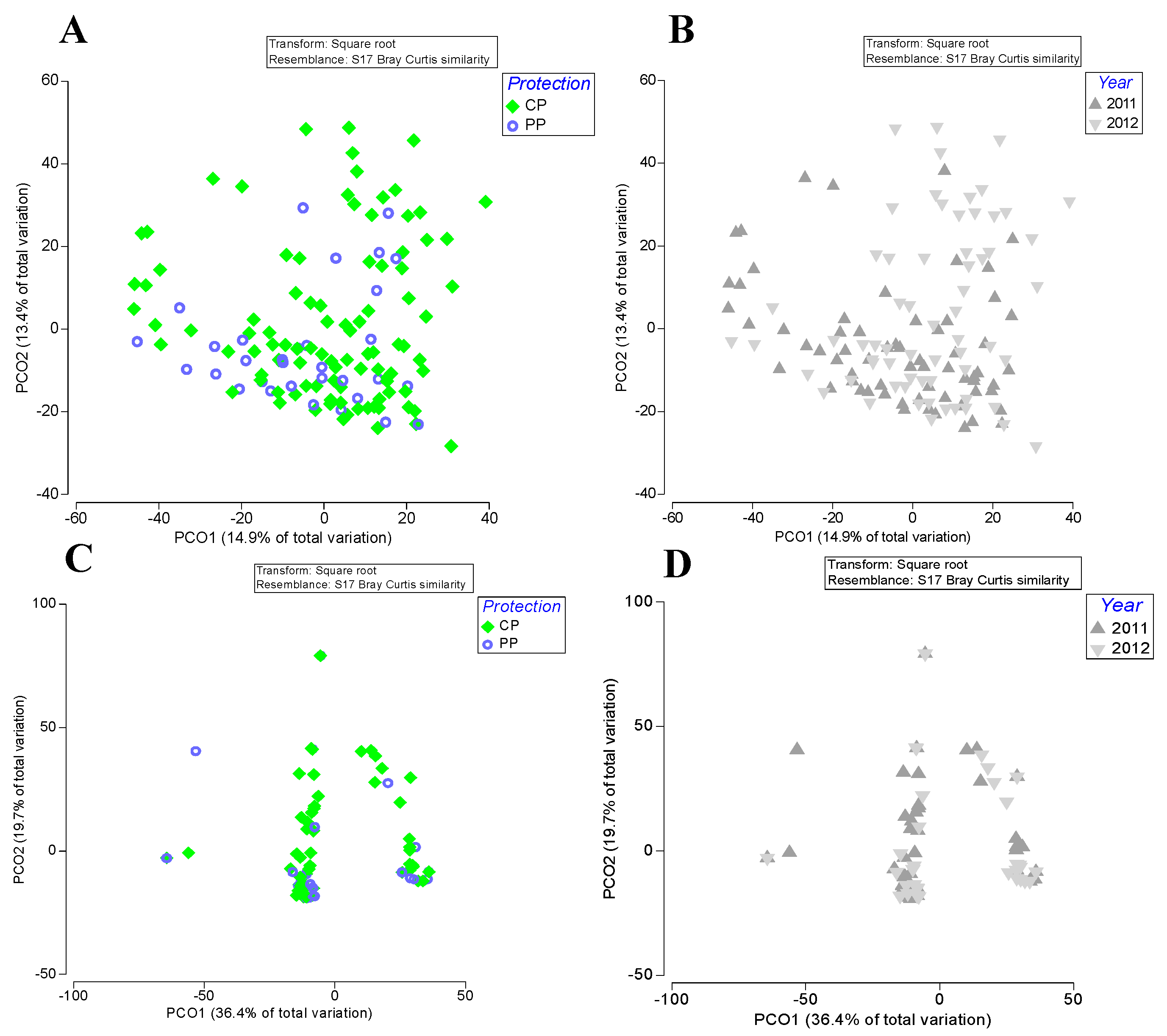
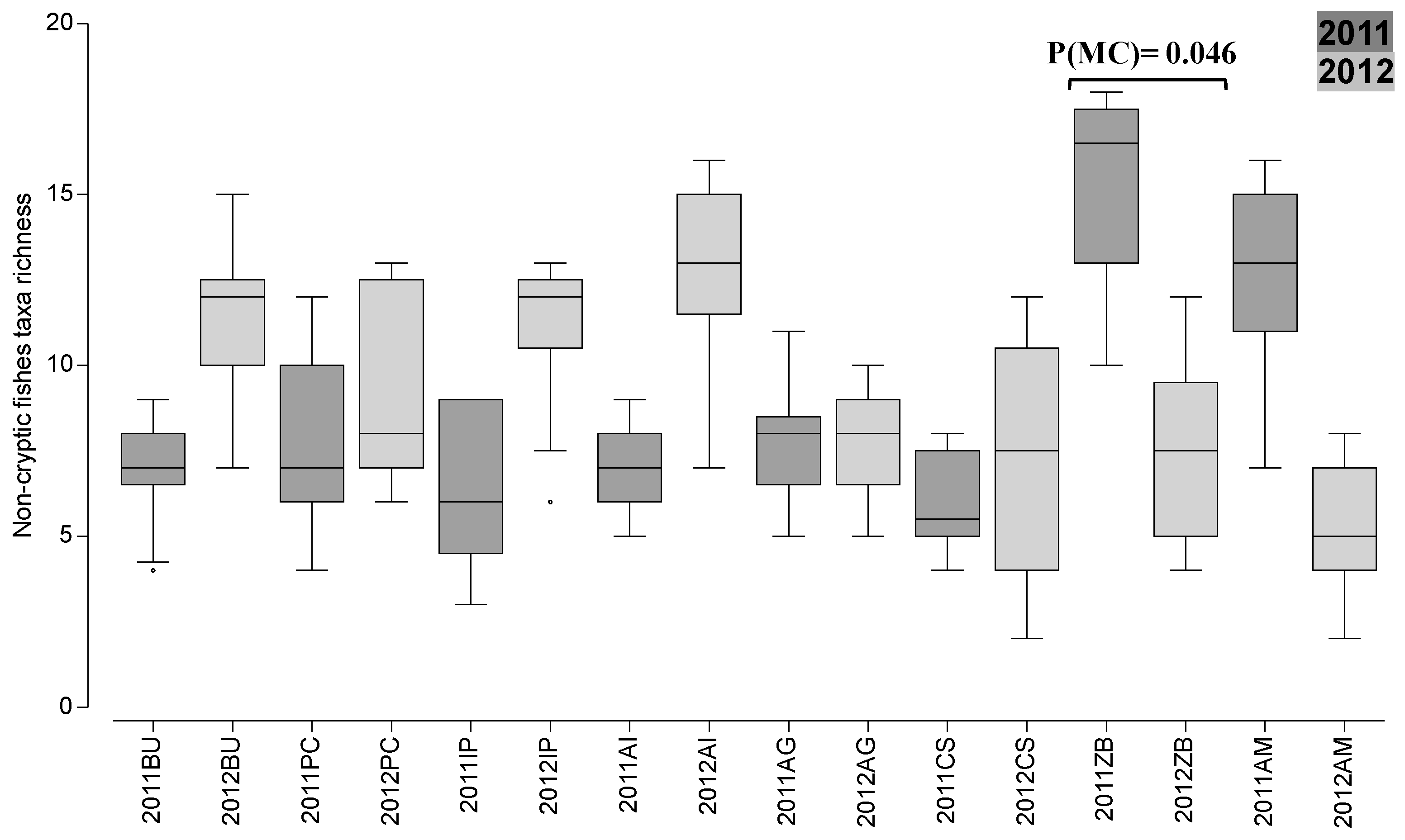
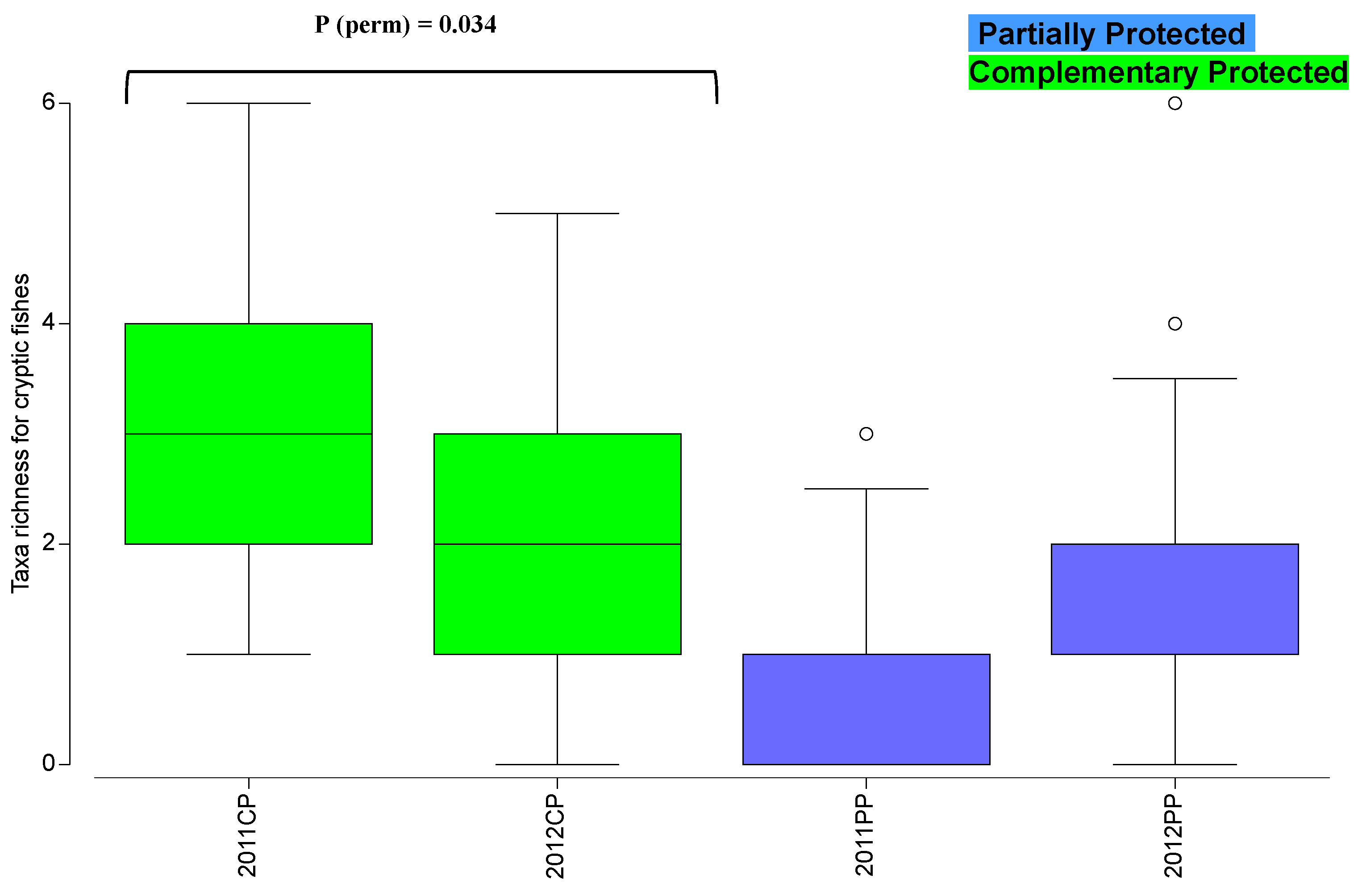
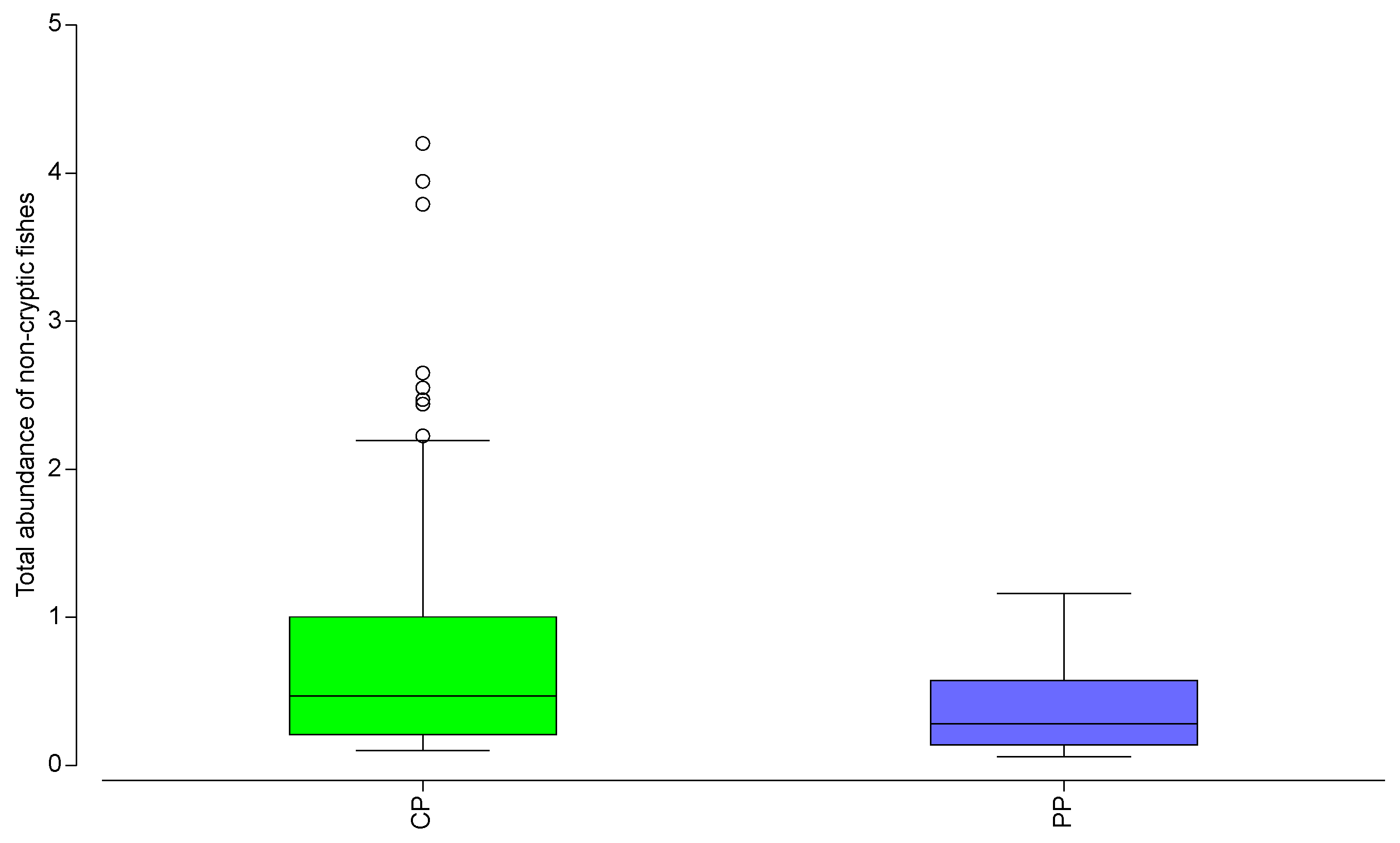
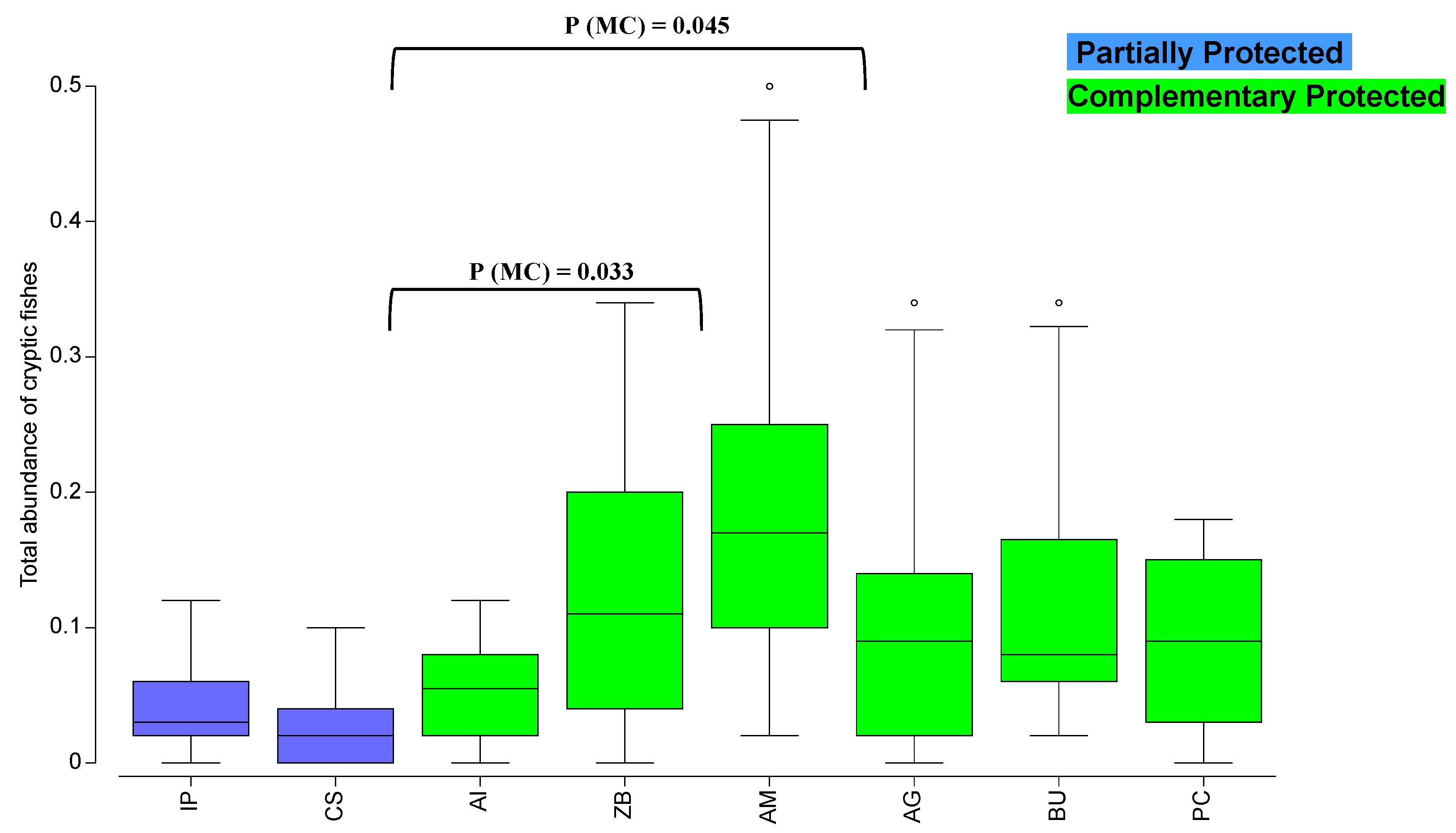
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef]
- Ojaveer, H.; Galil, B.S.; Minchin, D.; Olenin, S.; Amorim, A.; Canning-Clode, J.; Chainho, P.; Copp, G.H.; Gollasch, S.; Jelmert, A.; et al. Ten recommendations for advancing the assessment and management of non-indigenous species in marine ecosystems. Mar. Policy 2014, 44, 160–165. [Google Scholar] [CrossRef]
- Nielsen, P.; Nielsen, M.N.; McLaverty, C.; Kristensen, K.; Geitner, K.; Olsen, J.; Saurel, C.; Petersen, J.K. Management of bivalve fisheries in marine protected areas. Mar. Policy 2021, 124, 104357. [Google Scholar] [CrossRef]
- White, T.D.; Ong, T.; Ferretti, F.; Block, B.A.; McCauley, D.J.; Micheli, F.; De Leo, G.A. Tracking the response of industrial fishing fleets to large marine protected areas in the Pacific Ocean. Conserv. Biol. 2020, 34, 1571–1578. [Google Scholar] [CrossRef]
- Turnbull, J.W.; Johnston, E.L.; Clark, G.F. Evaluating the social and ecological effectiveness of partially protected marine areas. Conserv. Biol. 2021, 35, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, G.; Kenchington, R. Guidelines for Establishing Marine Protected Areas. In A Marine Conservation and Development Report; IUCN: Gland, Switzerland, 1992. [Google Scholar]
- Agardy, T.; Bridgewater, P.; Crosby, M.P.; Day, J.; Dayton, P.K.; Kenchington, R.; Laffoley, D.; McConney, P.; Murray, P.A.; Parks, J.E.; et al. Dangerous targets? Unresolved issues and ideological clashes around marine protected areas. Aquat. Conserv. 2003, 13, 353–367. [Google Scholar]
- Castro, N.; Romão, F.; Castro, J.J.; Pereira, T.J.; Pedro, S.; Viegas, V.; Costa, J.L. Catches, sales and discards: Small-scale fisheries in a Portuguese Marine Park. Reg. Stud. Mar. Sci. 2021, 42, 101643. [Google Scholar] [CrossRef]
- Agardy, M.T. Advances in marine conservation: The role of marine protected areas. Trends Ecol. Evol. 1994, 9, 267–270. [Google Scholar] [CrossRef]
- Murray, S.N.; Ambrose, R.F.; Bohnsack, J.A. No-take reserve networks: Sustaining fishery populations and marine ecosystems. Fisheries 1999, 24, 11–25. [Google Scholar] [CrossRef]
- Murawski, S.A.; Brown, R.; Lai, H.L.; Rago, P.J.; Hendrickson, L. Large-scale closed areas as a fishery-management tool in temperate marine systems: The Georges Bank experience. Bull. Mar. Sci. 2000, 66, 775–798. [Google Scholar]
- Sumaila, U.R.; Guenette, S.; Adler, J.; Chuenpagdee, R. Addressing ecosystem effects of fishing using marine protected areas. ICES J. Mar. Sci. 2000, 57, 752–760. [Google Scholar] [CrossRef]
- Roberts, C.M.; Bohnsack, J.A.; Gell, F.; Hawkins, J.P.; Goodridge, R. Effects of marine reserves on adjacent fisheries. Science 2001, 294, 1920–1923. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Charton, J.; Perez-Ruzafa, A.; Marcos, C.; Claudet, J.; Badalamentic, F.; Benedetti-Cecchid, L.; Falcon, J.M.; Milazzo, M.; Schembrig, P.; Stobarth, B.; et al. Effectiveness of European Atlanto-Mediterranean MPAs: Do they accomplish the expected effects on populations, communities and ecosystems? J. Nat. Conserv. 2008, 16, 193–221. [Google Scholar] [CrossRef]
- Fenberg, P.; Caselle, J.; Claudet, J.; Clemence, M.; Gaines, S.; García-Charton, J.; Gonçalves, E.; Grorud-Colvert, K.; Guidetti, P.; Jenkins, S.; et al. The science of European marine reserves: Status, efficacy, and future needs. Mar. Policy 2012, 36, 1012–1021. [Google Scholar] [CrossRef]
- Di Lorenzo, M.; Guidetti, P.; Di Franco, A.; Calò, A.; Claudet, J. Assessing spillover from marine protected areas and its drivers: A meta-analytical approach. Fish Fish 2020, 21, 906–915. [Google Scholar] [CrossRef]
- Palumbi, S.R. The ecology of marine protected areas. In Marine Community Ecology; Bertness, M.D., Gaines, S.D., Hay, M.E., Eds.; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 509–530. [Google Scholar]
- Halpern, B.S. The impact of marine reserves: Do reserves work and does reserve size matter? Ecol. Appl. 2003, 13, 117–137. [Google Scholar] [CrossRef]
- Hilborn, R.; Stokes, K.; Maguire, J.J.; Smith, T.; Botsford, L.W.; Mangel, M.; Orensanz, J.; Parma, A.; Rice, J.; Bell, J.; et al. When can marine reserves improve fisheries management? Ocean. Coast Manag. 2004, 47, 197–205. [Google Scholar] [CrossRef]
- Guidetti, P.; Sala, E. Community-wide effects of marine reserves in the Mediterranean Sea. Mar. Ecol. Prog. Ser. 2007, 335, 43–56. [Google Scholar] [CrossRef]
- Edgar, G.J.; Stuart-Smith, R.D. Ecological effects of marine protected areas on rocky reef communities—A continental-scale analysis. Mar. Ecol. Prog. Ser. 2009, 388, 51–62. [Google Scholar] [CrossRef]
- Pereira, T.J.; Manique, J.; Quintella, B.R.; Castro, N.; Almeida, P.R.; Costa, J.L. Changes in trophic ecology of fish assemblages after no take Marine Protected Area designation in the southwestern coast of Portugal. Ocean Coast Manag. 2017, 137, 144–153. [Google Scholar] [CrossRef]
- Claudet, J.; Osenberg, C.; Benedetti-Cecchi, L.; Domenici, P.; Garcıía-Charton, J.; Perez-Ruzafa, A.; Badalamenti, F.; Bayle-Sempere, J.; Brito, A.; Bulleri, F.; et al. Marine reserves: Size and age do matter. Ecol Lett. 2008, 11, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Jacinto, D.; Cruz, T.; Silva, T.; Castro, J.J. Management of the stalked barnacle (Pollicipes pollicipes) fishery in the Berlengas Nature Reserve (Portugal): Evaluation of bag and size limit regulation measures. Sci. Mar. 2011, 75, 439–445. [Google Scholar] [CrossRef]
- Bertocci, I.; Dominguez, R.; Freitas, C.; Sousa-Pinto, I. Patterns of variation of intertidal species of commercial interest in the Parque Litoral Norte (north Portugal) MPA: Comparison with three reference shores. Mar. Environ. Res. 2012, 77, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.; Mendes, S.; Franco, J.; Castanheira, M.; Castro, N.; Maranhão, P. Fish diversity in the Berlengas Natural Reserve (Portugal), a marine protected area. Ecologia 2011, 3, 35–43. [Google Scholar]
- Sousa, A.; Jacinto, D.; Penteado, N.; Pereira, D.; Silva, T.; Castro, J.J.; Leandro, S.M.; Cruz, T. Temporal variation of the fishers’ perception about the stalked barnacle (Pollicipes pollicipes) fishery at the Berlengas Nature Reserve (Portugal). Reg. Stud. Mar. Sci. 2020, 38, 101378. [Google Scholar] [CrossRef]
- Mendes, R.; Pereira da Silva, C.; Fonseca, C.; Gil, A. Managing and Monitoring the Recreational Use of Coastal Protected Areas: The Case of Berlengas Nature Reserve (Portugal). In Global Coastal Issues of 2020; Malvárez, G., Navas, F., Eds.; Journal of Coastal Research: Coconut Creek, FL, USA, 2020; Special Issue No. 95; pp. 123–127. [Google Scholar]
- Gonçalves, E.; Henriques, M.; Almada, V. Use of Temperate Reef-Fish Community to Identify Priorities in the Establishment of a Marine Area. In Aquatic Protected Areas: What Works Best and How Do We Know? Proceedings of the World Congress on Aquatic Protected Areas, Cairns, Australia; Beumer, J., Grant, A., Smith, D., Eds.; Australian Society for Fish Biology: Cairns, Australia, 2003; pp. 262–272. [Google Scholar]
- Sousa, I. Assessment of Reserve Effect in a Marine Protected Area: The Case Study of the Professor Luiz Saldanha Marine Park (Portugal). Master’s Thesis, Universidade do Algarve, Faro, Portugal, 2011. [Google Scholar]
- Henriques, S.; Pais, M.P.; Costa, M.J.; Cabral, H.N. Seasonal variability of rocky reef fish assemblages: Detecting functional and structural changes due to fishing effects. J. Sea Res. 2013, 79, 50–59. [Google Scholar] [CrossRef]
- Sousa, I.; Gonçalves, J.M.S.; Claudet, J.; Coelho, R.; Gonçalves, E.J.; Erzini, K. Soft-bottom fishes and spatial protection: Findings from a temperate marine protected area. PeerJ 2018, 6, e4653. [Google Scholar] [CrossRef] [PubMed]
- Priester, C.R.; Martínez-Ramírez, L.; Erzini, K.; Abecasis, D. The impact of trammel nets as an MPA soft bottom monitoring method. Ecol. Indic. 2021, 120, 106877. [Google Scholar] [CrossRef]
- Horta e Costa, B.; Erzini, K.; Caselle, J.E.; Folhas, H.; Gonçalves, E.J. Reserve effect’ within a temperate marine protected area in the north-eastern Atlantic (Arrábida Marine Park, Portugal). Mar. Ecol. Prog. Ser. 2013, 481, 11–24. [Google Scholar] [CrossRef]
- ICN. Turismo de Natureza: Enquadramento Estratégico, Parque Natural Do Sudoeste Alentejano e Costa Vicentina, 2000–2006; Instituto de Conservação da Natureza: Lisbon, Portugal, 2001. [Google Scholar]
- Castro, J.J.; Cruz, T. Marine conservation in a Southwest Portuguese natural park. J. Coast. Res. 2009, 56, 385–389. [Google Scholar]
- Cruz, T. Biologia e Ecologia do Percebe, Pollicipes pollicipes (Gmelin, 1790), no Litoral Sudoeste Português. Ph.D. Thesis, University of Évora, Évora, Portugal, 2000. [Google Scholar]
- Cruz, T.; Jacinto, D.; Sousa, A.; Penteado, N.; Pereira, D.; Fernandes, J.N.; Silva, T.; Castro, J.J. The state of the fishery, conservation and management of the stalked barnacle Pollicipes pollicipes in Portugal. Mar. Environ. Res. 2015, 112, 73–80. [Google Scholar] [CrossRef]
- Silva, J. Alterações na composição e na estrutura trófica das comunidades de peixes das áreas Marinhas Protegidas da Ilha do Pessegueiro e Cabo Sardão após a proibição da pesca. Master’s Thesis, Faculty of Sciences, University of Lisbon, Lisbon, Portugal, 2015. [Google Scholar]
- Belo, A.F.; Pereira, T.J.; Quintella, B.R.; Castro, N.; Costa, J.L.; Almeida, P.R. Movements of Diplodus sargus (Sparidae) within a Portuguese coastal marine protected area: Are they really protected? Mar. Environ. Res. 2016, 114, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.J.; Silva, A.F.; Belo, A.F.; Castro, N.; Costa, J.L.; Almeida, P.R.; Quintella, B.R. Assessing the size adequacy of a small no-take Marine Protected Area for Mediterranean moray and European conger. Mar. Ecol. Prog. Ser. 2017, 584, 213–227. [Google Scholar] [CrossRef]
- Fernández, C.G.; Paulo, D.; Serrão, E.A.; Engelen, A.H. Limited differences in fish and benthic communities and possible cascading effects inside and outside a protected marine area in Sagres (SW Portugal). Mar. Environ. Res. 2016, 114, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.J.; Manique, J.; Quintella, B.R.; Castro, N.; Almeida, P.R.; Costa, J.L. Changes in fish assemblage structure after implementation of Marine Protected Areas in the south western coast of Portugal. Ocean Coast. Manag. 2017, 135, 103–112. [Google Scholar] [CrossRef]
- Whitehead, P.J.; Bauchot, M.-L.; Hureau, J.-C.; Nielsen, J. (Eds.) Fishes of the North-East Atlantic and the Mediterranean; UNESCO: Paris, France, 1989; Volume I–III, p. 1473. [Google Scholar]
- Edgar, G.J.; Stuart-Smith, R.D.; Cooper, A.; Jacques, M.; Valentine, J. New opportunities for conservation of handfishes (Family Brachionichthyidae) and other inconspicuous and threatened marine species through citizen science. Biol. Conserv. 2017, 208, 174–182. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley: Hoboken, NJ, USA, 2014. [Google Scholar]
- Underwood, A.J. Experiments in Ecology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Clarke, K.; Gorley, R. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Di Lorenzo, M.; Claudet, J.; Guidetti, P. Spillover from marine protected areas to adjacent fisheries has an ecological and a fishery component. J. Nat. Conserv. 2016, 32, 62–66. [Google Scholar] [CrossRef]
- Corrales, X.; Vilas, D.; Piroddi, C.; Steenbeek, J.; Claudet, J.; Lloret, L.; Calò, A.; Di Franco, A.; Font, T.; Ligas, A.; et al. Multi-zone marine protected areas: Assessment of ecosystem and fisheries benefits using multiple ecosystem models. Ocean Coast. Manag. 2020, 193, 105232. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Ballesteros, E.; Clemente, S.; Gonçalves, E.J.; Estep, A.; Rose, P.; Sala, E. Contrasts in the marine ecosystem of two Macaronesian islands: A comparison between the remote Selvagens Reserve and Madeira Island. PLoS ONE 2017, 12, e0187935. [Google Scholar] [CrossRef]
- Forcada, A.; Valle, C.; Bonhomme, P.; Criquet, G.; Cadiou, G.; Lenfant, P.; Sánchez-Lizaso, J.L. Effects of habitat on spillover from marine protected areas to artisanal fisheries. Mar. Ecol. Prog. Ser. 2009, 379, 197–211. [Google Scholar] [CrossRef]
- Silva, A.F.; Horta e Costa, B.; Costa, J.L.; Pereira, E.; Marques, J.P.; Castro, J.J.; Lino, P.G.; Candeias-Mendes, A.; Pousão-Ferreira, P.; Sousa, I.; et al. Movements of Hatchery-Reared Dusky Groupers Released in a Northeast Atlantic Coastal Marine Protected Area. J. Mar. Sci. Eng. 2022, 10, 904. [Google Scholar] [CrossRef]
- Edgar, G.J.; Bustamante, R.H.; Fariña, J.-M.; Calvopiña, M.; Martínez, C.; Toral-Granda, M.V. Bias in evaluating the effects of marine protected areas: The importance of baseline data for the Galapagos Marine Reserve. Envir. Conserv. J. 2004, 31, 212–218. [Google Scholar] [CrossRef]
- Boubekri, I.; Amara, R.; Djebar, A.B.; Mazurek, H. Baseline data for marine protected areas planning and fisheries monitoring: Potential conflicts between recreational IUU and commercial fisheries in the proposed “Taza” MPA (Algeria, SW Mediterranean). Ocean Coast. Manag. 2021, 201, 105425. [Google Scholar] [CrossRef]
- Claudet, J.; Pelletier, D.; Jouvenel, J.Y.; Bachet, F.; Galzin, R. Assessing the effects of a marine protected area (MPA) on a reef fish assemblage in a northwestern Mediterranean marine reserve: Identifying community-based indicators. Biol. Conserv. 2006, 130, 349–369. [Google Scholar] [CrossRef]
- Moland, E.; Olsen, E.M.; Knutsen, H.; Garrigou, P.; Espeland, S.H.; Kleiven, A.R.; Andre, C.; Knutsen, J.A. Lobster and cod benefit from small-scale northern marine protected areas: Inference from an empirical before– after control-impact study. Proc. R. Soc. B 2013, 280, 20122679. [Google Scholar] [CrossRef]
- Roberts, D.A.; Poore, A.G.B.; Johnston, E.L. MBACI sampling of an episodic disturbance: Stormwater effects on algal epifauna. Mar. Environ. Res. 2007, 64, 514–523. [Google Scholar] [CrossRef]
- Pelletier, D.; Leleu, K.; Mou-Tham, G.; Guillemot, N.; Chabanet, P. Comparison of visual census and high definition video transects for monitoring coral reef fish assemblages. Fish. Res. 2011, 107, 84–93. [Google Scholar] [CrossRef]
- Egerton, J.P.; Johnson, A.F.; Turner, J.; LeVay, L.; Mascareñas-Osorio, I.; Aburto-Oropeza, O. Hydroacoustics as a tool to examine the effects of Marine Protected Areas and habitat type on marine fish communities. Sci. Rep. 2017, 8, 47. [Google Scholar] [CrossRef]
- Rojo, I.; Anadón, J.D.; García-Charton, J.A. Exceptionally high but still growing predatory reef fish biomass after 23 years of protection in a Marine Protected Area. PLoS ONE 2021, 16, e0246335. [Google Scholar] [CrossRef]
- Pais, M.P.; Henriques, S.; Costa, M.J.; Cabral, H.N. Topographic complexity and the power to detect structural and functional changes in temperate reef fish assemblages: The need for habitat-independent sample sizes. Ecol. Indic. 2014, 45, 18–27. [Google Scholar] [CrossRef]
- Henriques, S.; Pais, M.P.; Batista, M.I.; Costa, M.J.; Cabral, H.N. Response of fish-based metrics to anthropogenic pressures in temperate rocky reefs. Ecol. Indic. 2013, 25, 65–76. [Google Scholar] [CrossRef]
- Castro, J.J. Predação Humana No Litoral Rochoso Alentejano: Caracterização, Impacte Ecológico e Conservação. Ph.D. Thesis, University of Évora, Évora, Portugal, 2004. [Google Scholar]
- Willis, T.J.; Anderson, M.J. Structure of cryptic reef fish assemblages: Relationships with habitat characteristics and predator density. Mar. Ecol. Prog. Ser. 2003, 257, 209–221. [Google Scholar] [CrossRef]
- Pierpaolo, C.; Gianluca, S.; Gianfranco, M.; Pietro, B.; Teresa, R.; Vincenzo, I.; Franco, A. The effects of protection measures on fish assemblage in the Plemmirio marine reserve (Central Mediterranean Sea, Italy): A first assessment 5 years after its establishment. J. Sea Res. 2013, 79, 20–26. [Google Scholar] [CrossRef]
- de Benito-Abelló, C.; Bentes, L.; Sousa, I.; Pedaccini, M.; Villegas-Ríos, D.; Olsen, E.M.; Gonçalves, J.M.S.; Horta e Costa, B. Among-individual variation in white seabream (Diplodus sargus) spatial behaviour and protection in a coastal no-take area. ICES J. Mar. Sci. 2022, 79, 2265–2276. [Google Scholar] [CrossRef]
- Guidetti, P.; Milazzo, M.; Bussotti, S.; Molinari, A.; Murenu, M.; Pais, A.; Spano, N.; Balzano, R.; Agardy, T.; Boero, F.; et al. Italian marine reserve effectiveness: Does enforcement matter? Biol. Conserv. 2008, 141, 699–709. [Google Scholar] [CrossRef]
- Edgar, G.J.; Barrett, N.S. Effects of the declaration of marine reserves on Tasmanian reef fishes, invertebrates and plants. J. Exp. Mar. Biol. Ecol. 1999, 242, 107–144. [Google Scholar] [CrossRef]
- Halpern, B.S.; Warner, R.R. Marine reserves have rapid and lasting effects. Ecol. Lett. 2002, 5, 361–366. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | MS | Pseudo-F | P(perm) |
|---|---|---|---|---|
| Year | 1 | 9508.6 | 1.3334 | 0.203 |
| Protection | 1 | 7176.8 | 1.1755 | 0.276 |
| Area (Protection) | 6 | 6127.9 | 2.6436 | 0.001 |
| Year × Protection | 1 | 5693.4 | 0.79836 | 0.654 |
| Site (Area (Protection)) | 8 | 2316.9 | 1.3249 | 0.019 |
| Year × Area (Protection) | 6 | 7159.1 | 3.0461 | 0.001 |
| Year × Site (Area (Protection)) | 8 | 2349.1 | 1.3433 | 0.011 |
| Res | 97 | 1748.7 | ||
| Total | 128 |
| Source | df | MS | Pseudo-F | P(perm) |
|---|---|---|---|---|
| Year | 1 | 3734.8 | 1.3893 | 0.261 |
| Protection | 1 | 1123 | 0.40575 | 0.895 |
| Area (Protection) | 6 | 2850.7 | 1.6799 | 0.094 |
| Year × Protection | 1 | 1989.8 | 0.74617 | 0.585 |
| Site (Area (Protection)) | 8 | 1677.9 | 0.85856 | 0.692 |
| Year × Area (Protection) | 5 | 2668.4 | 1.2618 | 0.264 |
| Year × Site (Area (Protection)) | 6 | 2125.3 | 1.0875 | 0.338 |
| Res | 86 | 1954.3 | ||
| Total | 114 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, N.; Pereira, T.J.; Costa, A.; Sertório, J.; Fernandes, A.; Tavares, M.J.; Castro, J.J. Using Fish Assemblages to Assess the Ecological Effects of Marine Protection on Rocky Habitats in a Portuguese Natural Park. Hydrobiology 2023, 2, 181-195. https://doi.org/10.3390/hydrobiology2010012
Castro N, Pereira TJ, Costa A, Sertório J, Fernandes A, Tavares MJ, Castro JJ. Using Fish Assemblages to Assess the Ecological Effects of Marine Protection on Rocky Habitats in a Portuguese Natural Park. Hydrobiology. 2023; 2(1):181-195. https://doi.org/10.3390/hydrobiology2010012
Chicago/Turabian StyleCastro, Nuno, Tadeu J. Pereira, André Costa, Joana Sertório, António Fernandes, Maria J. Tavares, and João J. Castro. 2023. "Using Fish Assemblages to Assess the Ecological Effects of Marine Protection on Rocky Habitats in a Portuguese Natural Park" Hydrobiology 2, no. 1: 181-195. https://doi.org/10.3390/hydrobiology2010012
APA StyleCastro, N., Pereira, T. J., Costa, A., Sertório, J., Fernandes, A., Tavares, M. J., & Castro, J. J. (2023). Using Fish Assemblages to Assess the Ecological Effects of Marine Protection on Rocky Habitats in a Portuguese Natural Park. Hydrobiology, 2(1), 181-195. https://doi.org/10.3390/hydrobiology2010012