Abstract
Cannabis sativa L. has garnered attention as a potential source for new antimicrobial agents, particularly due to the increased prevalence of microbial resistance to conventional antimicrobials and the emergence of multidrug-resistant pathogens. This review, conducted according to the PRISMA 2020 statement, systematically analyzed the antimicrobial properties of C. sativa extracts and cannabinoids against various bacteria, fungi, viruses, and parasites. Data were collected from the scientific literature (102 papers) and clinical trials (5 studies) from 2014 to June 2024. Findings revealed that cannabinoids, especially CBD, demonstrate significant antimicrobial activity against Gram-positive bacteria like MRSA, Gram-negative bacteria such as Pseudomonas aeruginosa, various Candida species, SARS-CoV-2, and HIV. Additionally, CBD showed efficacy against parasitic infections like Echinococcus granulosus and Leishmania species. These results suggest that cannabinoids may represent a new class of antimicrobial agents with unique and diverse mechanisms of action, potentially effective in broad-spectrum therapies. This study highlights the urgent need for further research and standardized clinical trials to validate these findings and to develop cannabinoid-based treatments. The antimicrobial properties of C. sativa align with WHO priorities and support global health initiatives, offering promising avenues for addressing antimicrobial resistance and improving public health outcomes.
1. Introduction
Cannabis sativa L., part of the natural products arsenal, has been a rich source for identifying new therapeutic agents. In recent years, there has been a growing interest in using C. sativa and understanding how its bioactive compounds—phytocannabinoids—support the prevention and treatment of various diseases and conditions [1,2]. This interest is particularly relevant given the growing prevalence of microbial resistance to conventional antibiotics and the emergence of multidrug-resistant (MDR) pathogens.
The World Health Organization (WHO) has identified MDR pathogens as a significant threat to global health [3,4]. In 2019, antibiotic-resistant bacteria (ABR) were estimated to be responsible for approximately 1.27 million deaths [5]. Furthermore, the Centers for Disease Control and Prevention (CDC) reported that ABR was linked to an additional 5 million deaths [3,6].
This emphasizes the importance of novel therapeutic strategies as the CDC stated that “the post-antibiotic era has already arrived”. Antibiotic resistance is a part of the broader concept of antimicrobial resistance (AMR), a chronic global public health problem with projections indicating up to 10 million annual deaths by 2050 [7]. Thus, identifying and developing new compounds are crucial to maintain the effectiveness of current and future medical treatments against infectious diseases [8].
It is important to explain that cannabinoids can be produced in the human body (endogenous) or obtained from other sources (exogenous), such as plants like C. sativa, or synthesized. These bioactive compounds interact with the body’s endocannabinoid system (ECS), which consists of endocannabinoids, their receptors, and specific enzyme systems involved in synthesis and degradation [9,10]. This system adapts to changes depending on the type of cannabinoid and its binding to specific cannabinoid receptors, such as CB1 and CB2 (G-protein coupled receptors—GPCRs), as well as other molecular targets like ion channels and enzymes, which are involved in a wide range of physiological functions in different brain regions and tissues [11,12,13].
CB1 receptors, abundant in the human brain, modulate neurotransmitter systems including dopamine, serotonin, glutamate, noradrenaline, opioids, and gamma-aminobutyric acid (GABA), and are thereby chiefly responsible for the effects of endocannabinoids and analogous ligands in the brain and mediate the psychoactive effects of Cannabis [13,14]. They are involved in controlling brain activity, energy metabolism, heart function, the immune system, and reproduction [15]. CB2 receptors are widely distributed throughout the body, particularly in the cells of the immune system and in various brain regions, including the olfactory tubercle, cerebral cortex, striatum, thalamic nuclei, and hippocampus [9]. Both CB1 and CB2 receptors can result in agonist or antagonist effects, triggering numerous biological responses [16].
The ability of cannabinoids to interact with the ECS, primarily through their activity at cannabinoid receptors, influences various physiological functions and modulates neurotransmitter systems in the brain [9]. This interaction offers promising prospects for their therapeutic use in conditions like cancer, ischemia, Parkinson’s disease, Alzheimer’s disease, glaucoma, pain, and multiple sclerosis [9,10,17].
Approximately 120 cannabinoids have been isolated from C. sativa plant extracts. These are terpenophenolic structurally related phytocannabinoids—of which two well-studied examples are Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) [11]. THC acts as a partial agonist of CB1 and CB2 receptors, resulting in effects such as pain relief, muscle relaxation, and reduced nausea, but can also leads to adverse effects such as anxiety, psychosis, and sedation. In contrast, CBD acts as an antagonist to these receptors, demonstrating opposite effects, such as reducing sedation, psychosis, and anxiety, often contributing to mitigating the psychoactive effects of THC. This leads to anticonvulsant, antinociceptive, and antipsychotic effects [11].
In fact, antimicrobial activities of cannabinoids have been described among the many other reported pharmacological properties in recent years. Research conducted in 1976 reported topical antibacterial effects of CBD and THC combined against Gram-positive bacteria [18,19]. Previous studies on five major cannabinoids—THC, CBD, CBC (cannabichromene), CBG (cannabigerol), and CBN (cannabinol)—revealed their potent activity against various clinically relevant methicillin-resistant Staphylococcus aureus (MRSA) strains. Similar results were obtained in 2008 and 2020 for synthetic CBD [19,20].
Studies have shown that synthetic and natural CBD revealed the same pharmacological and therapeutic effects, ensuring that the results of studies with synthetic CBD are applicable to plant-derived CBD [21]. Other clinical and preclinical studies have investigated the efficacy of CBD in different topical applications. For instance, Botanix Pharmaceuticals conducted Phase II clinical trials evaluating synthetic CBD for the elimination of nasal colonization by Staphylococcus aureus (S. aureus), acne, and atopic dermatitis. These studies underscore the potential of CBD as an effective antimicrobial agent, especially against resistant bacterial strains like MRSA [20].
Other cannabinoids may potentially play a significant role in overcoming challenges in combating MDR pathogens, which are of paramount importance to public health due to their high morbidity and mortality profile. The mechanism of action of CBD includes disruption of the bacterial cytoplasmic membrane, which contribute to its antimicrobial activity. Recent investigations suggest that CBD may act synergistically with other antibiotics, such as bacitracin, through inhibition of undecaprenyl pyrophosphate phosphorylation. This ability highlights its potential as a new treatment in clinical settings for antibiotic-resistant Gram-positive bacterial infections [22].
AMR impacts various Sustainable Development Goals (SDGs) due to its broad consequences on health, economy, and society [3,5]. Among the 17 SDGs established under the 2030 Agenda by the United Nations, SDG 3 aims to ensure “Good Health and Well-being”. Specifically, AMR is addressed with indicator 3.d.2 (E. coli), which measures the “Percentage of blood stream infections due to selected organisms resistant to antimicrobials. Indeed, it has been confirmed by the WHO that AMR affects 7 of the 17 SDGs, namely SDGs 1 (No Poverty), 2 (Zero Hunger and Sustainable Agriculture), 3 (Good Health and Well-being), 6 (Clean Water and Sanitation), 8 (Decent Work and Economic Growth), 12 (Responsible Consumption and Production), and 17 (Partnerships for the Goals) [3,5,7]. These connections demonstrate how AMR is not just a health issue but also a challenge that affects various aspects of sustainable development, requiring an integrated and collaborative approach for its mitigation [3].
Based on these data and considering that cannabinoids exhibit broad polypharmacology and are extensively tested for a variety of disease indications, especially antimicrobial activity, they represent a potentially effective alternative, especially when incorporated into innovative dosage forms.
This study aims to explore the therapeutic potential of cannabinoids on infectious diseases, focusing on new strategies and experimental chemical, biological, and toxicological data. It particularly addresses their properties against MDR pathogens and including comparative elements on the biological activity of full-spectrum extracts and isolated cannabinoids to elucidate their full potential in targeting specific human anatomical systems where infections commonly occur.
2. Materials and Methods
This review was conducted according to the criteria described in the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 statement (http://www.prisma-statement.org/; accessed on 8 June 2024). The review protocol was registered in the PROSPERO platform (CRD42024564829).
Quantitative and statistical analysis of publications provides insights into key studies within the field and their interrelationships [23]. Bibliometric analysis serves as an appropriate methodology to understand the key articles, authors, and sources contributing to the field of C. sativa, as well as to uncover the knowledge structure pertaining to potential applications in infectious diseases.
The search strategy is outlined below and further detailed in the PRISMA 2020 Checklist and Table S1 (see Supplementary Materials). Scientific literature and clinical trials dated up to 15 June and 26 June 2024, respectively, were included. Data collection involved two phases:
- Step 1: Literature search on C. sativa and antimicrobial potential;
- Step 2: Clinical trials on C. sativa and applications in infectious diseases.
2.1. Literature Search Strategy
Multiple online databases, including Web of Science (WoS), Scopus, Google Scholar, EMBASE, and PubMed, store bibliographic details and citations of scientific works used. These platforms enable the search and retrieval of relevant information. For this study, Scopus (Elsevier) was selected due to its comprehensive coverage of the literature and its extensive use in bibliometric analysis.
Key search terms related to C. sativa were used for the literature search strategy. The initial search query yielded 70,591 results. Subsequently, the search was then refined using the following search string:
(TITLE-ABS-KEY(Cannabis) AND PUBYEAR > 2013 AND PUBYEAR < 2025 AND (LIMIT-TO(PUBSTAGE, “final”)) AND (LIMIT-TO(LANGUAGE, “English”)) AND (LIMIT-TO(SRCTYPE, “j”).
This refinement resulted in 64,394 records in English. Timespan restrictions were then applied, limiting this study to papers published between 2014 and 2024, reducing the total to 37,848 records.
After identifying the relevant literature, a systematic search was conducted using the terms “Cannabis” OR “Cannabinoids” combined with keywords such as “antimicrobial” “antibacterial,” “antifungal,” “antiparasitic or anthelmintic,” “antiviral,” “infectious diseases” and “multidrug-resistant” with the appropriate Boolean operators (AND, OR, NOT) resulted in 1192 records.
2.2. Criteria for Data Inclusion and Exclusion
The process began with the elimination of duplicate entries using the reference management software EndNote™ v20. After duplicate removal (n = 462), the dataset was scrutinized to identify and exclude irrelevant documents. Inclusion criteria required studies published within the last 10 years, written in English, and focused on the use of cannabinoids in managing bacteria, fungi, parasites, and viruses. This review specifically analyzed studies that investigated prospective interventions regarding the antimicrobial potential of cannabinoids, particularly those addressing broader impacts against MDR pathogens. Exclusion criteria included duplicate publications, studies unrelated to antimicrobials or cannabinoids, and those with irrelevant or incomplete information. Upon completion of the screening process, a total of 158 studies were included in the final dataset, with 102 selected for analysis. Figure 1 summarizes the overall data collection process.
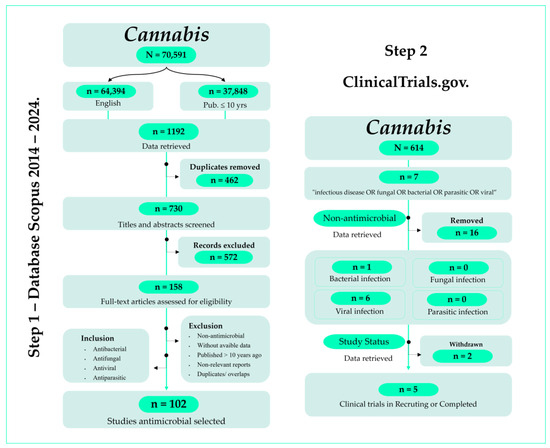
Figure 1.
PRISMA flowchart of the screening process in the different databases.
2.3. Data Cleaning
Data cleaning was performed using OpenRefine™ (version 3.7.2) an open-source tool. This software facilitated the identification of minor spelling variations and the merging of items that represented the same concept within the dataset.
2.4. Clinical Trials on C. sativa and Applications in Infectious Diseases
Building on the initial findings regarding the antimicrobial potential of C. sativa, the second is evaluating ongoing and concluded clinical trials investigating the efficacy and safety of C. sativa in diverse clinical settings related to infectious diseases (Figure 1). Data for this phase were sourced from the ClinicalTrials.gov database (https://clinicaltrials.gov/search?viewType=Table, accessed on 26 June 2024).
A comprehensive search and analysis of relevant clinical trials were conducted to explore the antimicrobial potential of C. sativa. The search parameters included terms such as “infection” OR “infectious disease” OR “viral” OR “bacterial” OR “fungal” OR “parasitic”. In the field “intervention/treatment”, the term used was Cannabis. Included clinical studies took place from 2014 to 2024. Figure 1 details the search results, which include a total of 23 studies. Of which 8 are related to studying the effect of Cannabis in the treatment of infectious diseases. Out of these 8 trials, only one tackled a bacterial infection, while 7 were related to viral infections, mainly Human Immunodeficiency Virus (HIV), which causes AIDS (Acquired Immunodeficiency Syndrome) and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), which causes Coronavirus Disease 2019 (COVID-19). The included clinical trials were also classified as withdrawn (2) or completed/recruiting (6).
3. Results
3.1. Antimicrobial Properties and Potential of C. sativa and Cannabinoids
C. sativa extracts and/or isolated cannabinoids were shown to have promising antimicrobial properties. Table 1, Table 2, Table 3, Table 4 and Table 5 summarize the most reported C. sativa extracts and/or formulation, the corresponding antimicrobial activities, as well as the tests performed to measure this activity. A total of 40 bacterial species (52.6%), 27 fungal species (35.5%), 3 viral species (3.9%), and 6 parasitic species (7.9%) were tested. Interestingly, most of the studied species are pathogens responsible for serious clinical conditions. In addition, most of the studied pathogens, namely bacteria, are known to lose sensitivity to conventional antibiotic therapy. This highlights the serious need to investigate alternative molecules such as the cannabinoids or other extracts of C. sativa.
One can notice that antimicrobial activity is related to the extraction method as well as the solvent. Ethanol was the most deployed solvent, used in 35 studies, followed by hexane, used in 20 studies. Some studies used both solvents (10 studies), while others used methanol and other solvents.
Several parts of the plant of C. sativa were deployed to extract cannabinoids, essential oils or fibers. It was noticed that the flowering parts of the plants gained more attention. This is because the flowering parts contain higher concentrations of cannabinoids, which are responsible for the antimicrobial properties.
The antimicrobial properties can be owed to the phenolic structure of cannabinoids [24]. The chemical structures of the main cannabinoids are shown in Figure 2.
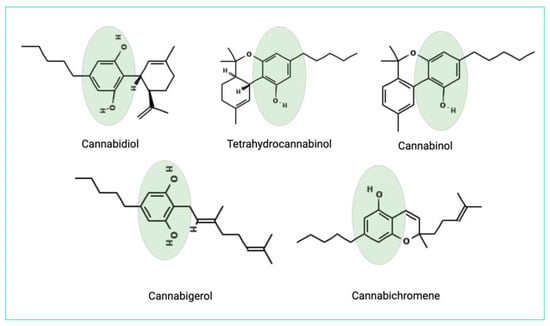
Figure 2.
The chemical structures of the main cannabinoids. Phenolic groups are highlighted.
3.1.1. Antibacterial Activity
C. sativa extracts as well as several pure cannabinoids were reported as potential antibacterial candidates. Table 1 presents the results of the found antibacterial activity for 40 bacterial species. Of those, 22 were Gram-negative and 18 were Gram-positive, including 5 that are multidrug-resistant, namely MRSA and vancomycin-resistant strains (VRE). Extracts were obtained from several parts of the plant of C. sativa. However, from the analyzed studies, the leaf is the most-used part (approximately 31.7%) and is most frequently tested against S. aureus, corresponding to 15 studies.
Bacteria can exist in two forms: planktonic cells and biofilms. The form of life depends on the external conditions. Bacterial cells (planktonic) differ from eucaryotic cells in the absence of membrane-encapsulated organelles such as the nucleus. In addition, they develop complex cell envelopes to preserve their structural integrity and defend against external insults [25]. Furthermore, certain bacterial species exhibits unique structures such as mucous capsules, motile flagella, adhesive pili, and resistance spores [25]. Antibacterials have several mechanisms of action as they can either damage bacterial cellular structures or interfere with the cellular metabolism. Some mechanisms of action include increased membrane permeability, membrane depolarization, inhibition of macromolecules synthesis, disruption of cellular communication, and inhibition of specific metabolic pathways [25,26].
Regarding C. sativa extracts and/or cannabinoids, they potentially have similar mechanisms of action to those of traditional antibiotics. For example, this study [27] showed that CBD was effective against S. aureus leading to the depolarization of the cell membrane, thus perturbating the membrane potential.
Moreover, C. sativa has potential applications in combination with traditional antibacterials. In a study by Abichabki, N. et al., it was demonstrated that the combination of CBD and Polymyxin B (PB) shows additive and/or synergistic effects against Gram-negative bacteria, including Polymyxin B-resistant strains like E. coli, especially those carrying plasmid-mediated colistin resistance (MCR-1)—an enzyme that adds a methyl group to a specific position of colistin, rendering it inactive. This synergistic activity occurs exclusively in the presence of the efflux pump inhibitor: phenylalanine-arginine-β-naphthylamide (PaβN). The addition of PaβN enhances the antibacterial efficacy of CBD + PB likely by increasing outer membrane permeability in Gram-negative bacteria. While the specific mechanisms of action of CBD and CBD + PB against E. coli are not fully elucidated, the study suggests they may operate through multiple pathways, including modulation of bacterial membrane permeability and inhibition of essential metabolic processes [28].
Resistance to colistin is a serious issue in hospitals, as colistin is a last-resort antibiotic for treating severe infections caused by Gram-negative bacteria. PAβN appears to play a crucial role in enhancing CBD penetration into bacterial cells, enabling the CBD + PB combination to effectively combat Gram-negative bacteria, including those resistant to PB [28]. This synergistic effect was also found in Klebsiella pneumoniae (K. pneumoniae). It was found by Abichabki, N. et al., that CB + PB was more effective than PB alone against this G-negative bacteria. The proposed mechanism of action is: PB binding to lipopolysaccharides (LPS) in the bacterial membrane, which leads to structural alterations, increased permeability, and consequently cell death [28].
Structure–activity relationship (SAR) studies were found in some articles. These studies aim at understanding the relation between the chemical structure and the function of molecules extracted from C. sativa. Antibacterial phenolic compounds can reversibly bind to bacterial proteins forming hydrophobic or hydrogen bonds. This, in turn, can lead to inactivating enzymes, microbial adhesins, and membrane transport proteins [29]. An in silico docking study provided information about the molecular interactions of structures of 24 cannabinoid compounds against 3 main bacterial proteins: penicillin-binding protein (PBP), isoleucyl-tRNA synthetase (Iso-TyrRS), and DNA gyrase. These findings helped elucidate the potential mechanisms behind their antibacterial activity, aiding in understanding the potential of optimizing these compounds against MRD bacteria. The cannabinoids showed potential as antimicrobial agents against MRSA, based on their predicted interactions with bacterial proteins. This study highlights the significance of these molecular interactions and their possible applications in developing new antimicrobial treatments [30].
Indeed, a synergy test was conducted on MRSA (USA300) between CBD and common antibiotics. A decrease in CFU (colony-forming units) was verified on MRSA [31]. Notably, the MIC values for CBD against both Salmonella typhimurium and Salmonella newington are promising, indicating a strong antibacterial activity. The MIC for S. typhimurium is particularly low (0.0125 µg/mL). The control antibiotic, ampicillin, had an MIC of 0.5 µg/mL for both strains, which is higher compared to the MIC values for CBD. This underscores the effectiveness of CBD as an antibacterial agent [32].
Besides the role of the active substance, formulation plays an important factor in potentiating the antibacterial activity. For example, essential oils (EOs) from C. sativa were studied with chitosan—a marine polymer with known antibacterial activity and found to reduce the cell viability of Aliivibrio fischeri (A. fischeri) to <30%. It is noteworthy that CS can play an important role due to its antibacterial effect. In this same study, it was concluded that film formulations had a stronger impact on cell viability compared to suspensions with increased essential oil concentration [33].
Additionally, the Cannabis-derived nitrogen-doped carbon dots (N@CSDs) can be used as a sustainable exfoliating agent to produce graphene sheets. These graphene sheets (GN@CSDs) exhibited excellent stability under various conditions and showed potential antibacterial properties against E. coli. For example, these findings highlight the potential of using C. sativa as a green carbon source to produce functional graphene materials with applications in antibacterial treatments and other fields [34].
Furthermore, an EO extracted from C. sativa exhibited moderate bactericidal activity against Listeria monocytogenes (L. monocytogenes), with MBC values greater than 2048 µg/mL. It significantly reduced motility, biofilm formation, and cell invasion capabilities of L. monocytogenes. Exposure to this essential oil downregulated key virulence genes, including those responsible for flagellar motility and cell invasion. In vivo studies using Galleria mellonella larvae confirmed the protective effect of this EO, demonstrating increased survival rates of the larvae infected with L. monocytogenes grown in the presence of sublethal concentrations of the essential oil. This highlights its potential application in food safety and public health [35]. The antibacterial activity was attributed to a synergism between several compounds present in the rich phytochemical complex of the EOs. Pure compounds tested, including CBD, α-pinene, β-pinene, and β-myrcene, also exhibited antibacterial activity, especially against Listeria and Enterococcus strains [36]. Another study concluded that the EO of C. sativa demonstrated potential as an anti-virulence agent against L. monocytogenes [37].
Antibacterial effects of cannabinoids were also evaluated on mouthwash products. Cannabinoids infused products were evaluated against the total-culturable aerobic bacterial content from dental plaque samples. Two mouthwash formulations containing CBD and CBG were tested and compared to the standard chlorhexidine digluconate 0.2%. The results demonstrated that both CBD and CBG mouthwashes exhibited antibacterial activity with MIC values of 0.58% compared to 0.24% for chlorhexidine. These findings suggest that further clinical trials are necessary to confirm their effectiveness and safety in vivo [38]. In addition, CBG exhibits potent antibacterial activity against Streptococcus mutans.
As aforementioned, Cannabis has important biofilm-inhibiting properties. A study found that the efficacy of CBG was modulated by quorum sensing (QS) systems, which are intercellular communication mechanisms used by bacteria to coordinate gene expression in response to population density. Two significant QS systems in S. mutans are ComCDE and LuxS. This suggests that CBG could be a promising compound for interfering with quorum sensing signals and combating bacterial infections, particularly those associated with dental biofilms and caries. Moreover, the research highlights the potential of CBG as an anti-quorum sensing agent, which could pave the way for the development of innovative antibacterial treatments [39].
Table 1 shows that among the analyzed cannabinoids and Cannabis extracts, 25.8% of cannabinoids are investigated against Gram-negative bacteria compared to only 17.6% of the found cannabinoids applied in studying Gram-positive bacteria. These results might suggest that there is more interest in testing the effect of cannabinoids (mainly CBD and CBG) against Gram-negative bacteria. This can be related to more resistances developed by these bacteria.

Table 1.
Antibacterial Activity of Extracts and Compounds from C. sativa.
Table 1.
Antibacterial Activity of Extracts and Compounds from C. sativa.
| PU 1 | Extract 2 | Main Compound 3 | Bacterial Strain | Activity (μg/mL) | Ref. | |
|---|---|---|---|---|---|---|
| MIC | Control 4 | |||||
| S | OIL | ∆9-THC THCV | MRSA MST 20654 | 256 | Van: 0.5–1 | [40] |
| CBD | S. aureus ATCC 25923 | 128 | Oxa: 0.125–0.50 | |||
| CBN | C. acnes | 256 | Nor: 0.25–0.25 | |||
| S | EtOH | n.r | S. epidermidis TISTR 517 | 128 | Oxa: 0.125–0.50 | [40] |
| L | DMSO | (1) CBD (2) CBG | S. mutans UA159 | (1) 6.29, (2) 3.16 | Clx: 2.36 | [41] |
| G | Gel | (1) CBD (2) CBG | S. mutans CCM 7409 | (1) 16, (2) 8, | Chx: 1 | [42] |
| (3) CBN (4) CBC | (3) 32, (4) 32 | |||||
| FL | EtOH | THCVA | S. aureus ATCC 29213, 43300 | 32–64 | Oxa: 0.25, >4 | [43] |
| OIL | CBDA CBC CBD | C. difficile ATCC 9689 | 2.5 × 102 to <10CFU/mL | n.r | [44] | |
| L. monocytogenes ATCC 7644 | 3.0 × 102 to <10CFU/mL | |||||
| E. faecalis ATTC 29212 | 1.1 × 102 to <10CFU/mL | |||||
| S. aureus ATCC 25923 | 7.2 × 102 to <10CFU/mL | |||||
| S. pyogenes ATCC 19615 | 2.9 × 102 to <10CFU/mL | |||||
| G | Gel | (1) CBD (2) CBG | P. gingivalis CCM 3985 | (1) 1.5, (2) 4, | Chx: 1 | [42] |
| (3) CBN (4) CBC | (3) 8, (4) 16 | |||||
| S | S | Peptide | E. coli | 33 | Amp: 0.5–2 | [45] |
| S. aureus | 33 | |||||
| S. epidermidis | increased growth | |||||
| S | H2O | Glycerolipids | S. epidermidis O-47 | 100 | 1 | [46] |
| S. epidermidis RP62A | 700–800 | reducing biofilm | ||||
| S | EtOH | (1) CBD, (2) CBG | E. coli O138 | (1) 4, (2) 8, | n.r | [47] |
| (3) CBN, (4) CBC | (3) 16, (4) 32 | |||||
| (1) IF (2) L | EtOH | CBD, CBG | E. coli ATCC 10536 | (1) 4.96, (2) 7.87 | Cip: 31.49 | [48] |
| E. coli PeruMycA 2 | (1) 15.74, (2) 39.68 | Cip: 9.92 | ||||
| E. coli PeruMycA 3 | (1) >200, (2) >200 | Cip: 79.37 | ||||
| P. aeruginosa ATCC 15442 | (1) 39.68 (2) 62.99 | Cip 125.99 | ||||
| S. typhi PeruMycA 7 | (1) >200, (2) >200 | Cip: 79.37 | ||||
| (1) IF (2) L | (1) EtOH (2) H2O | PMBA | B. cereus ATCC 12826 | (1) >200, (2) 125.99 | Cip: 125.99 | [48] |
| B. subtilis PeruMyc 6 | (1) 1.56, (2) 19.84 | Cip: 125.99 | ||||
| S. aureus ATCC 6538 | (1) 15.74, (2) 62.99 | Cip: 200 > 200 | ||||
| L | EtOH DMSO | CBD | S. aureus | 156.25 | Amp: n.r | [49] |
| S. epidermidis | 2.44 | Cip: n.r | ||||
| C. acnes | 2.44 | |||||
| EtOH | Phenolics | S. aureus MRSA | 179.7–718.7 | C+ | [50] | |
| EtOH | CBG CBD/THC | S. mutans | 3.9->250 | n.r | [51] | |
| B. cereus | 5–10 | |||||
| B. thuringiensis | 5–10 | |||||
| EtOH | CBG | S. mutans WT, ∆luxS | 2.5, 2.5 | C+/C− | [52] | |
| S. mutans ∆comC, ∆comE | 1.25, 1.25 | |||||
| S. mutans ∆comCDE | 0.75 | |||||
| EtOH | CBD CBG | P. aeruginosa | 400–1000 | C+ | [53] | |
| E. coli | 400–1000 | C+ | ||||
| L | H2O EtOH | CBD | S. aureus | <30% | n.r | [33] |
| L | EtOH | GEL-ALG–CS | S. aureus | IZ: n.r | n.r | [54] |
| P | (1) EtOH | CBD, CBG | B. subtilis | (1) and (2) >200 | Cip: <0.12 | [52] |
| (2) H2O | CBC, CBN | S. aureus | (1) 39.68, (2) 79.37 | |||
| CO2 | CBD, CBDA | S. aureus | 10.42–66.03 | Cip: 0.781 | [55] | |
| B. subtilis | ||||||
| IF | DME EtOH But | CBC, CBDA, CBG, CBGA, THC, THCA, THCV, Tp | S. aureus ATCC 29213 | 4 | Amp: 1 ChF:8 | [56] |
| S. aureus ATCC 25923 | 4 | Amp: 0.06 ChF:8 | ||||
| S. epidermidis CCM 50 | 8 | Amp: 0.06 ChF:8 | ||||
| S. epidermidis CCM 4418 | 8 | Amp: 0.06 ChF:8 | ||||
| S. lugdunensis CCM 4069 | 16 | Amp: 0.25 ChF:1.6 | ||||
| S. saprophyticus CCM 2727 | 4 | Amp: 0.5 ChF:3.12 | ||||
| S. pyogenes CCM 4425 | 64 | Amp: 16 ChF:3.12 | ||||
| P | (1) EtOH (2) H2O | CBD, CBG CBC, CBN | E. coli ATCC 10536 | (1) 31.49, (2) 39.68 | Cip: <0.12 | [52] |
| P. aeruginosa ATCC 15442 | (1) and (2) >200 | Cip: >0.12 | ||||
| S. typhi clinical isolate | (1) and (2) >200 | Cip: 0.38 | ||||
| CO2 | CBD, CBDA | E. coli P. aeruginosa | 10.42–66.03 | Cip: 0.781 | [55] | |
| L | EtOH | CBD | A. fischeri | 5–10 | Chx: 1–2 | [33] |
| IF | EtOH | CBD | E. coli ATCC 8739 | 0.675 | n.r | [57] |
| IF | EtOH | CBD | S. typhimurium | 0.125 | Amp: 0.5 | [32] |
| S. newington | 0.0125 | Amp: 0.5 | ||||
| IF | EtOH | CBD | S. typhimurium | 0.125 | Amp: 0.5 | [58] |
| S. newington | 0.125 | Amp: 0.5 | ||||
| R | EtOH | n.r | E. coli | 70 | Gent: 0.01 | [59] |
| P. aeruginosa | 75 | |||||
| K. pneumoniae | 60 | |||||
| EtOH | P. aeruginosa ATCC 10145 | 5.0–22.0 | [57] | |||
| S. enteritidis ATCC 13076 | ||||||
| S. typhimurium ATCC 14028 | ||||||
| FL | Hex | CBDA, CBD | S. aureus ATCC 6538 MRSA clinical strains | 4.88–78.1 | Van: 0.32–0.64 | [60] |
| IL, L | H2O | CTp | S. aureus ATCC 29213 | 31.25 | Clinda: 0.12 | [61] |
| S. aureus 101 clinical isolate | 62.5 | Clinda: 8 | ||||
| S. aureus 104 clinical isolate | 125 | Clinda: >128 | ||||
| S. aureus 105 clinical isolate | 15.62 | Clinda: 0.25 | ||||
| L | OIL | CBD, THC, Tp | L. monocytogenes ATCC 7644 L. monocytogenes ATCC 19114 L. monocytogenes LM4 S. aureus STA 32, ST 47 | 1.25–5 | n.r | [37] |
| EtOH | CBG | S. mutans | 2.5 | C+ | [62] | |
| CBD CBDV | S. aureus | 35.47 | Espt: n.r | [63] | ||
| IL, L | H2O | Tp | H. pylori | 7.81 | Clind: 0.06 | [61] |
| n.r | n.r | Carvacrol | Klebsiella pneumoniae | 125–250 | Cip: n.r | [64] |
| n.r | n.r | 2-AG, 1-AG, Anandamide | E. coli | n.r | MeOH: nr | [65] |
| C. rodentium | ||||||
| S. Typhimurium | ||||||
| CBD CBDV | E. coli | 29.1 | Espt: n.r | [63] | ||
| L | EtOH | (1) CBDA (2) CBD | S. aureus ATCC 25923 | (1) 2, (2) 1 | Clind: 0.06->128 | [31] |
| MRSA USA300 | (1) 4, (2) 1 | Tob: 0.06–1 | ||||
| S. epidermidis CA#71 | (1) 4, (2) 2 | Merp: 0.06–16 | ||||
| S. epidermidis ATCC 51625 | (1) 4, (2) 2 | Ofx: 0.5–2 | ||||
| Docking study | Structures 2, 3, 10, 14, 19, 23 | S. aureus SA1199B | 0.125 | Oxac: 0.125 | [30] | |
| Mw | CBD, CBG | Dental plaque bacteria | 0.58% | Chx: 0.24% | [38] | |
| FL | EtOH | PMBA | B. cereus ATCC 12826 B. subtilis PeruMycA 6 S. aureus ATCC 6538 | 4.96–15.74 | Cip: 9.92–125.99 | [2] |
| (1) CO2 | CBD, CBDA | S. aureus | (1) 4.69, (2) 9.38 | n.r | [66] | |
| (2) EtOH | CBN, CBGA, CBG, Tp | |||||
| OIL | CBD, THC | P. aeruginosa | 7.5 mm | n.r | [67] | |
| n.r | OIL | CBDA, CBDVA | B. subtilis | n.r “strong activity” | Amp: n.r | [68] |
| Cellulose | n.r | B. cereus | n.r | Cip: n.r | [69] | |
| n.r | n.r | (1) CBCA, (2) CBDVM | MRSA | (1) 1.41, (2) 4.91 | Vanc: 44.57 | [70] |
| MSSA | (1) 2.83, (2) >78.62 | |||||
| VRE | (1) 2.83, (2) >78.62 | |||||
| L | EtOH | CBDV, CBD, CBN | B. subtilis | 12.5 and 25 | n.r | [29] |
| EtOH | AEA | MRSA ATCC 33592 | >256 | C+ | [29] | |
| M + AEA | MRSA ATCC 33592 | 4 | C+ | |||
| AraS | MRSA ATCC 33592 | 28.4 | C+ | |||
| M + AraS | MRSA ATCC 33592 | 4 | C+ | |||
| n.r | OIL | CBDA, CBDVA | A. fischeri | n.r “strong activity” | Cip: n.r | [68] |
| L | EtOH | (1) CBDA | E. coli ATCC 25922 | (1) >64 (2) >64 | Clind: 0.06–8 | [31] |
| Tob: 0.12–2 | ||||||
| (2) CBD | P. aeruginosa PAA01 | (1) >64, (2) >64 | Merp: 0.06–0.5 | |||
| Ofx: 0.06–1 | ||||||
| Cellulose | n.r | E. coli | 500 | Cip: n.r | [69] | |
| (1) CO2 | CBD, CBDA | E. coli | (1) >37.5 | n.r | [66] | |
| (2) EtOH | CBN, CBGA, CBG | (2) >37.5 | ||||
| (1) CO2 | CBD, CBDA | E. faecalis | 50–100 | Gent: 16 | [67] | |
| (2) EtOH | CBN, CBGA, CBG, Tp | S. pneumonia | 20–50 | |||
| IF | Oil | OTp CBD | S. aureus ATCC 6538, 18As, 386 | 2, 32, 16 | Cip: 8, 16, 16 | [36] |
| S. epidermidis 18Bs | 16 | Cip: 8 | ||||
| L. monocytogenes ATCC 13932 | 32 | Amp: 0.25 | ||||
| L. monocytogenes NCTC 10888 | 4 | Amp: 0.25 | ||||
| L. monocytogenes ATCC 5008 | 8 | Amp: 0.25 | ||||
| L. monocytogenes 70, 139 | 16, 32 | Amp: 2, 0.5 | ||||
| E. faecalis ATCC 29212, V3, EQ19 | 2, 32, 8 | Cip: 4, 0.25, 4 | ||||
| E. faecalis V4, V5, V6 | 32, 16, 16 | Cip: 32, 16, 16 | ||||
| E. hirae ATCC 10541 | 32 | Cip: 8 | ||||
| B. subtilis ATCC 6633 | 16 | Amp: 2 | ||||
| B. cereus EB 362 | 1 | Amp: 16 | ||||
| n.r | Carbon | GN@CSDs | E. coli DH5α | n.r up to 100 | Clin: n.r | [34] |
| L | Hex | Pp | S. aureus ATCC 29213 | 3.8–7.8 | Cip: n.r | [71] |
| B. cereus M 25 | 3.1–9.8 | |||||
| L. lactis MTCC 440 | 2.8–5.1 IC50 | |||||
| B. thuringiensis MTCC 809 | 3.1–7.5 | |||||
| MRSA | 5.8–9.4 | |||||
| L | H2O | Tp | L. monocytogenes clinical isolate S. aureus clinical isolate | >2048 | n.r | [44] |
| S | Hex | Linoleic Acid | P. acnes | 15,000–25,000 | Erm: 3 | [72] |
| L | Hex | Pp | E. coli ATCC 25922 S. typhimurium MTCC 98 K. pneumoniae ATCC 75388 S. boydii NC 09357 P. fluorescens MTCC 103 X. flavus MTCC 132 | Not active | n.r | [71] |
| IF | OIL | OTp | L. monocytogenes 11#strains | >2048 | n.r | [35] |
| FL | PpOH | - | P. aeruginosa | 10,000–20,000 | Cip: 0.25–1 | [73] |
| Hex | E. coli | 10,000–18,000 | Cip: 0.015–0.25 | |||
1 PU—Part used: IF—Inflorescence; FL—Flowers; L—Leaf; R—Root; S—Seed. 2 Extract: BUT—Butane; EtOH—ethanol; DME—Dimethyl ether; IHOil—Industrial Hemp Oil; Hex—Hexane; Pp—Propanolol; TX-100—Triton X-100. 3 Compounds: PMBA—Phenolics mainly benzoic acid; PSP—Prenylspirodinone; Tp—Terpenes. 4 Control: AmB—Amphotericin B; CAS—Caspofungin; CF—Cefixime; Clinda: Clindamycin; Merp: Meropenem; NYS—Nystatin; Ofx: Ofloxacin; Tob: Tobramycin. Abbreviations: ATCC—American type culture collection, Carbon dots (N@CSDs) stabilized graphene (GN@CSDs); n.r—not reported; Q-Δ8-THC—quinone derivative of Δ8-THC. Abbreviation: A. fischeri—Aliivibrio fischeri; B. cereus—Bacillus cereus; B. subtilis—Bacillus subtilis; B. thuringiensis—Bacillus thuringiensis; C. acnes—Cutibacterium acnes; C. difficile—Clostridium difficile; E. coli—Escherichia coli; E. faecalis—Enterococcus faecalis; E. hirae—Enterococcus hirae; H. pylori—Helicobacter pylori; K. pneumoniae—Klebsiella pneumoniae; L. lactis—Lactococcus lactis; L. monocytogenes—Listeria monocytogenes; MRSA—Methicillin-resistant Staphylococcus aureus; P. aeruginosa—Pseudomonas aeruginosa; P. fluorescens—Pseudomonas fluorescens; P. gingivalis—Porphyromonas gingivalis; P. mutans—Propionibacterium mutans; S. albony—Salmonella albony; S. aureus—Staphylococcus aureus; S. boydii—Shigella boydii; S. epidermidis—Staphylococcus epidermidis; S. enteritidis—Salmonella enteritidis; S. lugdunensis—Staphylococcus lugdunensis; S. mutans—Streptococcus mutans; S. newington—Salmonella newington; S. pneumoniae—Streptococcus pneumoniae; S. pyogenes—Streptococcus pyogenes; S. saprophyticus—Staphylococcus saprophyticus; S. typhi—Salmonella typhi; S. typhimurium—Salmonella typhimurium; X. flavus—Xanthomonas flavus.
Besides their application as antimicrobial agents, C. sativa can also play a beneficial role in the synthesis of metal nanoparticles (NPs). These NPs (Table 2) are known to possess antimicrobial properties, which are further enhanced by using C. sativa-mediated synthesis.
NPs have numerous applications, especially in medical sciences due to their unique physicochemical properties [74]. Among these, metallic NPs have gained significant attention thanks to their very small sizes and antimicrobial properties. There are several methods applied in the synthesis of NPs, among them there are conventional chemical synthesis and biological methods. Chemical synthesis implies the use of toxic reagents, posing, thus, environmental hazards [74,75]. Biological methods are characterized by their sustainability, making them highly desirable. Recently, the use of plant extracts, such as those from C. sativa, has been shown as a promising green synthesis approach [75].
Many metallic NPs are known for their antimicrobial prosperities. These results from the electrostatic interactions between the positively charged NPs and the negatively charged bacterial cell surfaces owed to amino, carboxyl, and phosphate groups. This interaction leads to the generation of reactive oxygen species (ROS), which penetrate the bacterial cells and damage essential components such as DNA, proteins, and lipids. The greater efficacy of NPs against Gram-negative bacteria is due to their thinner peptidoglycan layer, allowing easier NPs penetration. On the other hand, Gram-positive bacteria, despite having a thicker peptidoglycan layer, possess numerous pores that facilitate NPs ingress, thus supporting their antimicrobial potential [76]. Interestingly, antibacterial properties of PdNPs, are attributed to the slow oxidation and liberation of monovalent Pd ions; PdNPs also interact with thiol proteins on the cell wall and form complexes with electronic donors containing nitrogen, phosphorus, oxygen, or sulfur atoms, further disrupting bacterial cell function [77].
For instance, the CdONPs synthesized using C. sativa, displayed antimicrobial activity against several bacterial strains. This activity varied from one strain to another, being stronger against Bacillus subtilis and Klebsiella pneumoniae, while showing moderate effectiveness against other tested bacterial strains. Also, Cs-CdO-NPs exhibited moderate antibacterial activity against B. subtilis and K. pneumoniae, with inhibition zones of 13 ± 0.8 mm and 12 ± 0.7 mm, respectively. Other strains such as S. aureus, E. coli, P. aeruginosa showed mild to moderate inhibition zones ranging from 7 ± 0.51 mm to 11 ± 1 mm [75].
Moreover, the synthesized C. sativa capped PdNPs exhibited strong antibacterial activity against both normal and MDR strains of E. coli, S. aureus, K. pneumoniae, and S. bony. The MIC values for these PdNPs were found to be 52.04 µg/mL for E. coli, 60.23 µg/mL for S. aureus, 63.1 µg/mL for K. pneumoniae, and 68.3 µg/mL for S. bony. The article showed a substantial and exponential increase in inhibition activity against MDR strains, highlighting the potent antibacterial properties of the synthesized PdNPs [77].
NPs produced using extracts from C. sativa are found in various studies. Examples of those are Ag-based NPs [78] or Au-based NPs [78,79] only, or included such as Ag-Au NPs [59,78,80], Ag-ZnO NPs [81].
Bimetallic nanoparticles (Ag-AuNPs) synthesized using C. sativa waste extract show significant antimicrobial potential against various strains of P. aeruginosa. Interestingly, MIC was consistently 2.0 mg L−1 across all tested strains. However, evaluation of minimal bactericidal concentration (MBC) and evaluation of minimal biofilm inhibitory concentration (MBIC) values varied, reflecting strain-specific responses. These NPs exhibited strong bactericidal effects against P. aeruginosa DBM 3081 and ATCC 10145, with MBC:MIC of 0.1 and 0.4, respectively, while DBM 3777 and 15442 showed bacteriostatic effects. Additionally, the NPs demonstrated high efficacy in inhibiting biofilm formation, with MBIC values ranging from 3.4 to 13.3 mg L−1. P. aeruginosa PAO1 did not display these effects as MBC was not achieved in the concentration range used [80].
Regarding Ag-ZnO doped NPs, these demonstrated excellent antimicrobial activity against various bacterial strains, including P. aeruginosa, K. pneumoniae, S. typhi, and E. coli. Additionally, they exhibited moderate activity against MRSA, with inhibition zones ranging from 10 to 20 mm compared to 5–10 mm for the control (Amp) [81].
ZnO-NPs synthesized from the leaves extract of Cannabis plants exhibited activity against S. aureus (SCC2993), B. cereus (BCC4391) and K. pneumoniae (KCC3812) with a zone of inhibition 11.11 ± 1.0, 09.31 ± 1.4 and 10.26 ± 1.0 mm, respectively [82].
Table 2.
Antibacterial Activity of Nanoparticles Synthesized from C. sativa Extracts.
Table 2.
Antibacterial Activity of Nanoparticles Synthesized from C. sativa Extracts.
| PU 1 | Extract 2 | Nanoparticles 3 | Microorganism/Strain | Zone of Inhibition (mm) | Control 4 (mm) | Ref. |
|---|---|---|---|---|---|---|
| L | EtOH | CdO NPs | S. aureus | 10.00 | Plant extracts alone | [75] |
| B. subtilis | 13.00 | |||||
| E. coli | 8.50 | |||||
| P. aeruginosa | 9.00 | |||||
| K. pneumoniae | 12.00 | |||||
| L | H2O | CeO2 NPs | E. coli ATCC 25922 | 250–500 | Cip:60–1000 | [76] |
| S. enteridis ATCC 13075 | 250–1000 | |||||
| B. subtilis | 500–1000 | |||||
| S. aureus ATCC 25923 | 500–1000 | |||||
| F | H2O | PdNPs | E. coli | 52.04 | C+ and C− | [77] |
| S. aureus | 60.23 | |||||
| K. pneumoniae | 63.10 | |||||
| S. bony | 68.30 | |||||
| L | EtOH | (1) AgNPs (2) Au–AgNPs | S. aureus ATCC 25923 | (1) 0.40, (2) 0.50 | CF: 2.0 | [78] |
| B. subtilis ATCC 6633 | (1) 0.25, (2) 1.25 | CF: 4.0 | ||||
| K. pneumoniae | (1) 0.75, (2) 0.30 | CF: 3.7 | ||||
| P. aeruginosa | (1) 0.50, (2) 1.00 | CF: 4.0 | ||||
| L | EtOH | AuNPs | S. aureus, B. subtilis | 75–71.55 | Ofx: 69.55 | [79] |
| E. coli, S. typhi | 69.55 | |||||
| L | By-P | Ag-AuNPs | P. aeruginosa DBM 3081 | MBC:MICratio 0.1 | MBC: 0.2:0.3 | [80] |
| P. aeruginosa DBM 3777 | MBC:MICratio 5.0 | MBC: 10.0:16.8 | ||||
| P. aeruginosa ATCC 10145 | MBC:MICratio 0.4 | MBC: 0.7:1.1 | ||||
| P. aeruginosa ATCC 15442 | MBC:MICratio 1.0 | MBC: 1.3:2.2 | ||||
| P. aeruginosa PAO1 | MBC:MICratio 0.1 | n.r | ||||
| R | EtOH | Ag-AuNPs | E. coli | 25 | Gent: 10 | [59] |
| S. aureus | 65 | |||||
| P. aeruginosa | 100 | |||||
| K. pneumoniae | 200 | |||||
| L | H2O | (1) Ag-ZnONP (2) ZnONP (3) Ag-ZnONP | MRSA | (1) 10–20 | Amp: 5–10 | [81] |
| P. aeruginosa | (2) no active, (3) 0.48 | Amp: 1.00 | ||||
| K. pneumoniae | (2) no active, (3) 0.52 | Amp: 1.00 | ||||
| S. typhi | (2) no active, (3) 0.70 | Amp: 1.00 | ||||
| E. coli | (2) no active, (3) 1.00 | Amp: 1.00 | ||||
| L | H2O | ZnONPs | S. aureus SCC2993 | 11.11 | Amp: 25 | [82] |
| B. cereus BCC4391 | 9.31 | |||||
| K. pneumoniae KCC3812 | 10.26 |
1 PU—Part used: F—Flowers; L—Leaf; R—Root. 2 Extract: By-P—By-product extract; EtOH—ethanol. 3 Nanoparticles: AgNPs: Silver Nanoparticles; Ag-AuNPs: Silver-Gold Alloy Nanoparticles; CdONPs: Cadmium Oxide Nanoparticles; CeO2NPs: Cerium Oxide Nanoparticles; PdNPs: Palladium Nanoparticles; ZnONPs: Zinc Oxide Nanoparticles. 4 Control: Amp—Ampiciline; AmB—Amphotericin B; Cip—Ciprofloxacin; CF—Cefixime; CLT—Gent-Gentamicine; MBC—Evaluation of minimal bactericidal concentration; Ofx: Ofloxacin. Abbreviations: ATCC—American type culture collection. Abbreviation: B. cereus—Bacillus cereus; B. subtilis—Bacillus subtilis; E. coli—Escherichia coli; K. pneumoniae—Klebsiella pneumoniae; MRSA—Methicillin-resistant Staphylococcus aureus; P. aeruginosa—Pseudomonas aeruginosa; S. bony—Salmonella bony; S. aureus—Staphylococcus aureus; S. enteritidis—Salmonella enteritidis; S. typhi—Salmonella typhi.
NPs have been recognized as a promising alternative to traditional antibiotics due to their ability to combat various infections effectively [74,77]. Studies have shown that NPs synthesized using C. sativa extracts exhibit notable antimicrobial activity against a range of pathogens, including Gram-positive, Gram-negative and MDR-resistant bacteria. This green synthesis method underscores the potential of integrating plant-based resources in nanotechnology for safer and more effective applications.
3.1.2. Antifungal Activity
Antifungal agents function in a manner akin to antibacterials by disrupting essential cellular processes or structures. For example, compromising the integrity of the cell wall leads to the leakage of cytoplasmic contents, membrane disruption, increased osmotic sensitivity, and ultimately, the inhibition of fungal growth [83,84].
Table 3 summarizes the outcomes of antifungal assays using diverse extracts and cannabinoids derived from C. sativa against various fungi and yeast species. The distribution of studies among the fungal species tested is detailed as follows: Candida spp. (43.33%), Aspergillus spp. (16.67%), Trichophyton spp. (13.33%), Malassezia spp. (10.00%), Fusarium spp. (6.67%), Neosartorya spp. (6.67%), Microsporum spp. (3.33%), Cryptococcus neoformans (3.33%), and Rosellinia necatrix (3.33%). These percentages reflect the proportion of each species among the total number of fungal species studied. The dataset encompasses a total of 30 studies, averaging approximately 4.29 studies per year, and includes 33 different fungal species.
Table 3.
Minimal Inhibitory Concentrations of Extracts and Compounds from Cannabis against Fungal and Yeast.
Table 3.
Minimal Inhibitory Concentrations of Extracts and Compounds from Cannabis against Fungal and Yeast.
| PU 1 | Extract 2 | Compound 3 | Microorganism/Strain | MIC/Control 4 (μg/mL) | Ref. | |
|---|---|---|---|---|---|---|
| Aspergillus | ||||||
| FL | EtOH | PMBA | A. fumigatus | 39.68 | GRF: 1.26 | [48] |
| FL-L | EtOH | PMBA | A. currey CCF 5207 | <6.25 | GRF: >8 | [48] |
| FL-L | EtOH | PMBA | A. gypseum CCF 6261 | 125.99–158.74 | GRF: 1.59 | [48] |
| FL-L | EtOH | PMBA | A. quadrifidum CCF 5792 | 31.49–125.99 | GRF: >8 | [48] |
| FL-L | EtOH | PMBA | A. insingulare CCF 5417 | 125.99 | GRF: >8 | [48] |
| IF | BUT | CBDA, CBD, CBGA, CBG, CBN, THCA, THC, THCV | A. insingulare CCF 5943 | 64–256 | CLT: 0.5–1.0 | [56] |
| DME | A. insingulare CCF 5417 | >1024 | TBF: 0.5–2.0 | |||
| EtOH | ||||||
| Candida | ||||||
| L | n.d | CBD isolated | C. albicans | 4312.5 | NYS: n.d | [49] |
| I-L | EtOH | PMBA | C. albicans YEPGA 6379 | >200 | FCZ: 1.0 | [48] |
| I-L | EtOH | PMBA | C. albicans YEPGA 6183 | 15.75 | FCZ: 2.0 | [48] |
| I-L | EtOH | PMBA | C. parapsilosis YEPGA 6551 | <6.25 | FCZ: 4.0 | [48] |
| I-L | EtOH | PMBA | C. tropicalis YEPGA 6184 | 15.75 | FCZ: 2.0 | [48] |
| FL-L | 24 n.d | C. glabrata | 6.54–10.0 | AmB: 2.5 | [85] | |
| FL-L | 24 n.d | C. krusei | 10.0 | AmB: 2.5 | [85] | |
| FL-L | Q-Δ8-THC | C. neoformans | 0.7 | AmB: 2.5 | [85] | |
| IF | TX-100 | CBDVA, CBL, CBD, CBC, CBN, CBG | C. albicans ATCC 10231 | IZ: 3.5 | NYS: 2 | [86] |
| IF | H2O | Phenolics | C. albicans YEPGA 6379 | >200 | FCZ: 1.0 | [52] |
| EtOH | C. albicans YEPGA 6183 | FCZ: 2.0 | ||||
| C. parapsilosis YEPGA 6551 | 62.99 | FCZ: 4.0 | [52] | |||
| C. tropicalis YEPGA 6184 | 79.37 | FCZ: 2.0 | [52] | |||
| IF | Hex | CBDA, CBD, CBGA, CBG, CBN, THCAA | C. albicans ATCC 10231 | Not active | CAS: 0.06 | [60] |
| IF | n.d | CBD | C. albicans, C. dubliniensis | 12.5–100 | NYS: 2 | [87] |
| C. glabrata, C. krusei | ||||||
| C. tropicalis | ||||||
| IF | HOil | n.d | C. albicans, C. glabrata C. krusei, C. tropicalis | 6.25 | AmB: 0.5–1.0 | [61] |
| IF | IHOil | CBD, CBDA, CBG, CBGA, CBN, THC | C. glabrata C. albicans | 16–32 | AmB: 1.0–2.0 NYS: 2 | [66,67] |
| CBD-H | 8–16 | |||||
| Wild-H | 32–64 | |||||
| L | Hex | PSP | C. albicans MTCC 4748 | 11.9–3.57 | NYS: 0.5 | [71] |
| IF | IHOil | CBDA, CBD, CBGA, CBG, CBN, THC | C. albicans CO23 | 195.3 | NYS: 2 | [88] |
| C. glabrata DSY 562 | 97.6 | |||||
| C. krusei 45709 | 390.6 | |||||
| C. tropicalis 47829 | 390.6 | |||||
| F | Hex-Pp | n.d | C. albicans MTCC 3017 | IZ: 20–23 | FCZ: 24 | [73] |
| Epidermophyton | ||||||
| IF | But | CBDA, CBD, CBGA, CBG, CBN, THCA, THC, THCV | E. floccosum | 64–256 | CLT: 0.5–1.0 | [56] |
| DME | ||||||
| EtOH | ||||||
| Malassezia | ||||||
| IF | IHOil | CBDA, CBD, CBGA, CBG, CBN, THC | M. furfur 180 | 390.6 | NYS: 2 Not active | [88] |
| M. furfur 188 | 195.3 | |||||
| M. globosa 130 | 195.3 | |||||
| M. sympodialis 855 | 195.3 | |||||
| L | n.d | CBD isolated | M. furfur | 312.5 | NYS: n.d | [49] |
| IF | But | CBDA, CBD, CBGA, CBG, CBN, THCA, THC, THCV | M. canis CCM 8353 | 128–256 | CLT: 0.5–1.0 TBF: 0.5–2.0 | [56] |
| DME | ||||||
| EtOH | ||||||
| Nylanderia | ||||||
| IF | But | CBDA, CBD, CBGA, CBG, CBN, THCA, THC, THCV | N. fulva CCF 5338, 5782, 6025 N. gypsea CCF 5215 | 64–128 | CLT: 0.5–1.0 TBF: 0.5–2.0 | [56] |
| DME | ||||||
| EtOH | ||||||
| IF | H2O EtOH | Phenolics | N. gypseum CCF 6261 | >200 | GRF: 1.59 | [52] |
| Trichophyton | ||||||
| I-L | EtOH | PMBA | T. erinacei CCF 5930 | 39.68–125.99 | GRF: 3.17 | [48,52] |
| T. mentagrophytes CCF 4823 | 39.68–125.99 | GRF: 2.52 | [48,52] | |||
| T. rubrum CCF 4933 | 62.99–79.37 | GRF: 1.26 | [48,52] | |||
| T. tonsurans CCF 4834 | 62.99–79.37 | GRF: 0.19 | [48,52] | |||
| IF | BUT | CBDA, CBD, CBGA, CBG, CBN, THCA, THC, THCV | T. interdigitale CCM 8337 | 64–128 | TBF: 0.25 | [56] |
| DME | T. tonsurans CCF 4930 | 64–128 | CTZ: 0.13–1 | |||
| EtOH | T. rubrum CCF 4879 | 16–32/128 | CTZ: 0.13–1 | |||
1 PU—Part used: IF—Inflorescence; FL—Flowers; L—Leaf. 2 Compounds: PMBA—Phenolics mainly benzoic acid; PSP—Prenylspirodinone; Tp—Terpenes. 3 Extract: BUT—Butane; EtOH—ethanol; DME—Dimethyl ether; IHOil—Industrial Hemp Oil; Hex—Hexane; Pp—Propanolol; TX-100—Triton X-100. 4 Control: AmB—Amphotericin B; CAS—Caspofungin; CLT—Clotrimazole; FCZ—Fluconazole; GRF—Griseofulvin; NYS—Nystatin; TBF—Terbinafine. Abbreviations: ATCC—American type culture collection, n.r—not reported. Abbreviation: A. currey—Arthroderma currey; A. flavus—Aspergillus flavus; A. fumigatus—Aspergillus fumigatus; A. gypseum—Arthroderma gypseum; A. insingulare—Arthroderma insingulare; A. quadrifidum—Arthroderma quadrifidum; C. albicans—Candida albicans; C. glabrata—Candida glabrata; C. krusei—Candida krusei; C. neoformans—Cryptococcus neoformans; C. parapsilosis—Candida parapsilosis; C. tropicalis—Candida tropicalis; E. floccosum—Epidermophyton floccosum; Fusarium sp.—Fusarium species; M. furfur—Malassezia furfur; M. canis—Microsporum canis; Mucor sp.—Mucor species; N. gypseum—Nannizia gypseum; N. fulva—Neocosmospora fulva; N. gypsea—Neocosmospora gypsea; Penicillium sp.—Penicillium species; Rhodotorula spp.—Rhodotorula species; R. necatrix—Rosellinia necatrix; T. erinacei—Trichophyton erinacei; T. interdigitale—Trichophyton interdigitale; T. mentagrophytes—Trichophyton mentagrophytes; T. rubrum—Trichophyton rubrum; T. tonsurans—Trichophyton tonsurans.
Recent studies have explored various extracts of C. sativa, including those derived from flowers, inflorescences, and leaves, using different solvents such as ethanol (EtOH), propanol, hexane, and water, revealing promising potential in the development of new therapeutic agents. These extracts were extensively evaluated mainly through MIC tests and fungal growth inhibition zones. Notably, C. albicans (16%) was the most studied fungal species, with CBD being the most tested compound against this species, underscoring its relevance in antifungal research. Hydroethanol and aqueous extracts of C. sativa demonstrated significant antifungal activity, with MICs varying from extremely low to moderate values, depending on the fungal species tested [48]. The most relevant MIC observed for Candida was <6.25 μg/mL, compared to the control, fluconazole, with MICs ranging from 1 to 4 μg/mL.
The essential oils (EOs) extracted from the inflorescences of C. sativa demonstrated significant antifungal activity against various species of Candida and Cryptococcus neoformans, with variations in efficacy depending on the type of Cannabis used (industrial, CBD, or wild). The presence of cannabinoids such as CBD, CBDA, CBG, CBGA, CBN, and THC contributed to the observed antifungal activity. Different types of hemp exhibit distinct chemical profiles that influence their antifungal efficacy. For instance, CBD hemp, with its high concentration of CBD, may show greater antifungal activity compared to industrial and wild hemp, which have lower concentrations of bioactive compounds like CBD [67]. Additionally, in another study, the essential oils (EOs) showed significant antifungal activity against various species of Candida and Malassezia, with variations in efficacy depending on the fungal species [88].
Research has highlighted the significant antifungal activity of CBD against various Candida species, with MIC values ranging from <6.25 to 100 μg/L. In addition, CBD was effective in inhibiting biofilm formation, making it a promising candidate for treating Candida infections, especially those involving biofilms that are typically more resistant to treatment. These findings support further exploration and potential use of CBD in antifungal therapies [87].
Studies concluded that several oxygenated cannabinoid derivatives, particularly those derived from Δ8-THC and Δ9-THC, demonstrated significant antifungal activities. For example, O-Δ8-THC was notably effective against Cryptococcus neoformans, while another cannabinoid-derivative exhibited broad-spectrum activity against Candida species. These findings suggest that specific cannabinoid derivatives hold promise as antifungal agents, potentially offering alternatives or adjuncts to conventional antifungal therapies [85].
Analysis concludes that C. sativa extracts, especially when prepared with solvents such as dimethyl ether (DME) and butane (BUT), are effective in inhibiting the growth of microorganisms causing skin diseases. These extracts are rich in cannabinoids, such as CBD and THC, along with other bioactive compounds. The tested dermatophytes included species such as Trichophyton rubrum, T. tonsurans, and Epidermophyton floccosum. MIC values varied, with E. floccosum showing a value of 128 µg/mL for the DME and BUT extracts, and a MIC of 256 µg/mL for the EtOH extracts. These findings suggest that C. sativa extracts, especially those rich in cannabinoids like THCA, have potential as alternative or supportive treatments for skin infections caused by dermatophytes. These extracts have great potential to be developed as natural topical treatments, offering a promising alternative to synthetic antifungal and antibacterial agents [56].
Comparatively, some studies have shown comparable or superior efficacy of C. sativa extracts to conventional antifungal agents like fluconazole, and griseofulvin [48]. MIC values for C. sativa extracts (E1 and E2) are compared against griseofulvin, which is used as a control in the tests. For example, for Arthroderma currey (CCF 5207), both E1 and E2 (<6.25 µg/mL) show greater potency compared to griseofulvin (>8 µg/mL). On the other hand, although C. sativa extracts (E1 and E2) exhibit antifungal activity potential against Candida parapsilosis (YEPGA 6551), E1 and E2 (<6.25 µg/mL) are less effective compared to fluconazole (4 µg/mL) used as a control [48]. Potential mechanisms of action were explored, highlighting phenolic parts of cannabinoids, and terpenes (e.g., α-pinene, β-pinene, myrcene, terpinolene, α-humulene, caryophyllene oxide) as key bioactives compounds responsible for the observed antifungal activity [55]. Among these bioactives, CBD can exert its antifungal effects potentially by disrupting the biosynthesis of ergosterol, a vital component of fungal cell membranes. This disruption could lead to membrane destabilization and increased permeability, ultimately impairing fungal cell viability and growth. These findings significantly contribute to understanding the therapeutic benefits of C. sativa in treating fungal infections, suggesting potential future applications in antifungal medicine.
Further, researchers evaluated also a variety of compounds, including AuNPs [79], ZnONPs, Ag-ZnONPs [81], and Q-Δ8-THC, alongside various cannabinoids [85]. The efficacy of these compounds was tested against a broad spectrum of fungi, demonstrating specific MICs notably low for certain fungi. For instance, studies have demonstrated the antifungal efficacy of AuNPs against A. flavus and A. niger, with zones of inhibition measuring 71.55 mm and 69.55 mm, respectively [79], while testing Ag-doped ZnO nanoparticles revealed MICs for Fusarium spp. and Rosellinia necatrix, with inhibition zones of 14.1 mm and 23.25 mm, respectively. In contrast, the negative controls showed smaller inhibition zones of 41.2 mm for R. necatrix and 38.3 mm for Fusarium spp., indicating that the nanoparticles were more active [81].
However, further research is needed to fully elucidate the molecular mechanisms underlying these effects and to optimize the efficacy and safety of these extracts as potential antifungal treatments. This additional investigation is crucial to validate the therapeutic use of C. sativa and explore its potential in clinical practice as a viable alternative to conventional ant fungal treatments [48,52,73,85,87].
3.1.3. Antiviral Activity
The results of antiviral tests using different extracts and cannabinoids from C. sativa species against several viruses are summarized in Table 4. The antiviral activities of various Cannabis-derived compounds against SARS-CoV-2, HIV, and HCV are highlighted. These viruses include SARS-CoV-2 (75%), HIV (12.5%), and HCV (12.5%). The compounds tested include CBD, Δ9-THC, CBDA, CBGA, CBN, CBNA, THCB, CBN-C3, and Δ8-THCA. The most-used plant part was the leaf, and CBD was the most-tested cannabinoid. The antiviral activities were evaluated using different assays such as in vitro, cell culture, in silico, molecular docking, functional assay, clinical trial, and luciferase assay.
Effective antiviral treatments target various stages of the viral life cycle to prevent viral entry into host cells, inhibit essential enzymes for viral replication, block viral protein processing, prevent viral DNA integration into host DNA, and inhibit the release of new viruses. These mechanisms are crucial for controlling infections while minimizing damage to host cells [89].
The cellular entry of SARS-CoV-2 begins with the interaction between the viral spike protein and the human cell surface receptor known as angiotensin-converting enzyme 2 (ACE2), which is highly expressed on endothelial cells in the pulmonary and cardiovascular systems. The spike protein forms a homotrimeric structure, where each monomer (~150 kDa) consists of an S1 subunit responsible for binding to ACE2, an S2 subunit that facilitates virus-cell membrane fusion, and a transmembrane domain. Following the binding of the S1 subunit to ACE2, the TMPRSS2 protease (transmembrane serine protease 2) located on the human cell membrane cleaves the spike protein at specific S1/S2 and S2 sites, triggering structural changes that enable efficient entry of SARS-CoV-2 into the host cell [90].
Recent studies using affinity selection techniques identified cannabinoids like CBDA, CBGA, and THCA as potential ligands for the SARS-CoV-2 spike protein. CBDA and CBGA effectively blocked viral entry by binding to the S1 protein’s active site, confirmed through competition assays with SBP-1 peptide. CBGA also bound to an allosteric site on the S1 subunit. Functional assays with live SARS-CoV-2 demonstrated that CBDA and CBGA could prevent viral entry and infection, showing strong binding affinity with KD values of 5.6 ± 2.2 μM and 19.8 ± 2.7 μM, respectively. This suggests CBDA and CBGA as promising therapeutic agents against COVID-19, with potential for nanoparticle delivery to enhance efficacy and reduce toxicity [91].
Table 4.
Antiviral Activities of C. sativa-Derived Compounds and Extracts.
Table 4.
Antiviral Activities of C. sativa-Derived Compounds and Extracts.
| PU 1 | Extract 2 | Compound 3 | Results | Assay | Ref. |
|---|---|---|---|---|---|
| L | H2O | CBD | IC50 = 1.86 μM; CBD inhibits SARS-CoV-2 replication in Vero cells (IC50 = 7.91 μM) and human lung carcinoma cells expressing ACE2 (EC50 = 1.24 μM). | In vitro | [92] |
| L | H2O | Δ9-THC | IC50 = 16.23 μM; Δ9-THC inhibits SARS-CoV-2 replication in Vero cells (IC50 = 10.25 μM). | In vitro | [92] |
| L | H2O | CBDA | CBDA binds to the SARS-CoV-2 spike protein, preventing it from attaching to ACE2 and entering the cells. | Cell culture | [92] |
| L | H2O | CBGA | CBGA binds to the SARS-CoV-2 spike protein and inhibits the viral protease 3CLpro (IC50 = 14.40 μM). | Cell culture | [92] |
| L | H2O | CBN | Predicted to inhibit 3CLpro and ACE2 | Cell culture | [92] |
| n.r | n.r | CBNA | Predicted to inhibit viral proteases 3CLpro and PLpro as well as ACE2. | In silico | [92] |
| n.r | n.r | THCB | Found to be a potent inhibitor of 3CLpro (IC50 = 3.62 μM) | In silico | [92] |
| n.r | n.r | CBD | IC50 (ACE2) ranging from 3.96 μM to 0.01 μM; IC50 (3CLpro) ranging from 1.9 μM to 9.4 μM; Superior to remdesivir and nafamostat mesylate. | Molecular Docking | [93] |
| n.r | n.r | CBD | Binding energy range: 2.22–5.30 kcal/mol; Highest binding energy indicating strong inhibitory potential. Effective against main protease, RdRp, PLpro, NSP15, and spike glycoprotein. | Molecular Docking | [94] |
| n.r | n.r | CBDA | Kd = 5.6 ± 2.2 μM; CBDA blocks cell entry and infection of SARS-CoV-2. | Functional Assay | |
| n.r | n.r | CBGA | Kd = 19.8 ± 2.7 μM; CBGA blocks cell entry and infection of SARS-CoV-2. | Functional Assay | [91] |
| n.r | n.r | CBN-C3 | Docking Score: −8.3 kcal/mol; Interactions: 1 hydrogen bond (Phe390), pi-pi stacked (Phe40), pi-alkyl (Leu391, Lys562, Ala99). | Molecular Docking | [95] |
| n.r | n.r | Δ8-THCA | Docking Score: −8.0 kcal/mol; Interactions: Hydrogen bonds (Asp350, Tyr385), pi-sigma (Phe390), pi-alkyl (Trp349, Phe40, Arg393). | Molecular Docking | [95] |
| n.r | n.r | THC | Significant reduction in T-cell activation (CD8+CD38+HLA-DR+); decreased plasma IL-6 and TNF-α; reduced HIV DNA total and integrated; changes in gut microbiome composition. | In vivo Clinical Trial | [96] |
| L | H2O | CBD | Inhibited HCV replication by 86.4% at 10 µM; EC50 = 3.163 µM; IC50 = 15.670 µM; SI = 4.954; Minimal toxicity. | Luciferase Assay | [97] |
1 PU—Part used: L—Leaf; n.r—not reported. 2 Extract: H2O—aqueous. 3 Compounds: CBD—Cannabidiol; Δ9-THC—Δ9-Tetrahydrocannabinol; CBDA—Cannabidiolic Acid; CBGA—Cannabigerolic Acid; TCBN—Cannabinol; CBNA—Cannabinolic Acid; THCB-Δ9—Tetrahydrocannabibutol; CBN—C3—Cannabivarine; Δ8-THCA—Δ8-Tetrahydrocannabinolic Acid; THC—Tetrahydrocannabinol; HCB-Δ9—Tetrahydrocannabibutol. Abbreviations: n.r—not reported.
In a comprehensive docking review, various Cannabis-derived compounds were evaluated for their inhibitory effects on SARS-CoV-2. CBD exhibited the highest binding energy among these compounds, indicating its potential as a drug target, particularly against viral proteins like RNA-dependent RNA polymerase. The study also explored their effects on reducing inflammation associated with SARS-CoV-2 and other viruses like HIV [94].
Overall, These results demonstrate that the inhibitory effects are strongest on the main protease (3CLpro), followed by RNA-dependent RNA polymerase (RdRp), Papain-like protease (PLpro), non-structural protein 15 (NSP15), and spike glycoprotein. mong the tested compounds, Cannabidiol (CBD) showed the highest inhibitory potential, particularly against SARS-CoV-2 and HCV [94].
The synthesis of diethyl and dibutyl alkylated CBD via nucleophilic substitution reaction involves using CBD as a nucleophile to attack the carbon of an alkyl halide, resulting in the formation of alkylated CBD derivatives. This process is significant in modifying the chemical structure of CBD to potentially enhance its biological activity or pharmacokinetic properties [93].
CBD exhibited significant in vitro activity against Hepatitis C virus (HCV) with 86.4% inhibition at 10 µM but showed no activity against Hepatitis B virus (HBV), highlighting its potential as a therapeutic agent for HCV [54].
A study evaluated the safety, tolerability, and effects of oral cannabinoids (THC and CBD) in people living with HIV undergoing effective antiretroviral therapy (ART). A randomized clinical trial was conducted with 26 participants who had an undetectable viral load for at least 3 years, receiving THC and CBD capsules in two ratios (1:1 and 1:9) daily for 12 weeks. Safety and tolerability were assessed through hematological, hepatic, and renal tests, as well as in-person interviews and questionnaires. The results demonstrated that oral cannabinoids were safe and well tolerated, significantly reducing T-cell activation, inflammatory markers, and HIV persistence, while also promoting beneficial changes in gut microbiome composition [95].
These findings underscore the potential of cannabinoids, particularly CBD, in targeting multiple stages of the viral life cycle and reducing viral infectivity. Further research and clinical studies are needed to validate these promising initial findings and explore their therapeutic applications in treating viral infections effectively.
3.1.4. Antiparasitic Activity
The antiparasitic activities of various extracts and cannabinoids derived from C. sativa against different parasite species are summarized in Table 5. The data show that CBD is the most studied compound, involved in five different studies. The distribution of studies across different parasites, namely, Echinococcus granulosus (40%), Leishmania tropica (10%), L. major (30%), L. donovani (10%), and Helminths (10%).
In 2024, CBD was the most tested cannabinoid for its antiparasitic activity, particularly against E. granulosus sensu stricto. This compound demonstrated significant effects, including 0% viability in vitro and reduction in cyst weight, along with complete collapse of the germinal layer in vivo experiments. CBD extracted from decarboxylated inflorescences extract exhibited substantial efficacy, with pronounced morphological changes observed. Complementary in vivo experiments further supported these findings, showing a substantial reduction in cyst weight and complete collapse of the germinal layer when CBD was combined with albendazole. This combination enhances efficacy, suggesting potential for improved therapeutic strategies against cystic echinococcosis. Further research is required to optimize dosing and delivery systems to enhance bioavailability and efficacy [27].
CBGA (4-terpenyl cannabinolate), was evaluated against Leishmania donovani using molecular docking, revealing interactions with cyclophilin and glycerol-3-phosphate dehydrogenase [98]. Additionally, gold-silver bimetallic nanoparticles synthesized with C. sativa aqueous leaf extract demonstrated potent leishmanicidal activity against Leishmania major promastigotes, highlighting their potential as anti-leishmanial agents [78].
Table 5.
In Vitro and In Vivo Antiparasitic Activity of C. sativa.
Table 5.
In Vitro and In Vivo Antiparasitic Activity of C. sativa.
| PU 1/Extract 2 | Compound 3/Assay | Microorganism | Results | Ref. |
|---|---|---|---|---|
| IF/EtOH | In Vitro/CBD | E. granulosus | At 50 µg/mL, 80% decrease in protoscoleces viability at 24 h; 0% viability at 48 h. Significant soma contraction, hook loss, tegumental damage, and rostellar disorganization. Severe structural damage including rostellum loss. | [27] |
| IF/EtOH | In Vitro/CBD | E. granulosus | At 10 µg/mL, 87 ± 23% collapse of germinal layer after 4 days. Extensive germinal layer collapse. | [27] |
| IF/EtOH | In Vivo/CBD | E. granulosus | Significant reduction in cyst weight compared to control. Greater reduction when combined with albendazole. | [27] |
| IF/EtOH | In Vivo/CBD + ABZ | E. granulosus | Enhanced reduction in cyst weight compared to CBD or ABZ alone. Co-administration caused greater ultrastructural alteration of the germinal layer compared to monotherapy. | [27] |
| IF/OIL | In Vivo/CBD, δ9-THC, CBC | L. tropica | Essential oils showed significant efficacy in reducing tissue lesions in infected mice. The reduction in lesion size was comparable to amphotericin B. | [61,67] |
| L/H2O | In Vitro/AuNPs | L. major | Maximum anti-leishmanial activity observed at 250 µg/mL concentration. AuNPs showed significant inhibition of promastigote forms of L. major with an IC50 value of 282.597 µg/mL. | [78] |
| L/H2O | In Vitro/Ag NPs | L. major | AgNPs demonstrated significant anti-leishmanial activity with an IC50 value of 155.824 µg/mL. | [78] |
| L/H2O | In Vitro/Au-Ag NPs | L. major | Au-Ag bimetallic NPs exhibited considerable anti-leishmanial activity with an IC50 value of 227.277 µg/mL. | [78] |
| n.r | In Vitro δ8-THC Derivatives | L. donovani | Compound 14 showed pronounced antileishmanial effect against promastigotes with IC50 = 0.06 µg/mL and IC90 = 0.13 µg/mL, which are significantly lower than the standard compound Amphotericin B (IC50 = 1.0 µg/mL and IC90 = 2.0 µg/mL). | [85] |
| n.r | In Vivo/Δ9-THC | Helminths | Significant negative associations between THCA levels and worm burden. Higher C. sativa use linked to lower helminth infection and reinfection rates. | [85] |
| n.r | In Silico/Cannabinoids | L. spp. | Molecular docking analysis identified several cannabinoids with strong docking energies against various Leishmania protein targets, suggesting potential inhibitory effects. | [98] |
1 PU—Part used: L—Leaf; IF—inflorescence. 2 Extract: H2O—aqueous. 3 Compounds: CBD—Cannabidiol; Δ9-THC—Δ9-Tetrahydrocannabinol; CBDA—Cannabidiolic Acid; CBGA—Cannabigerolic Acid; TCBN—Cannabinol; CBNA—Cannabinolic Acid; THCB-Δ9—Tetrahydrocannabibutol; CBN—C3-Cannabivarine; Δ8-THCA—Δ8-Tetrahydrocannabinolic Acid; THC—Tetrahydrocannabinol; HCB—Δ9—Tetrahydrocannabibutol. n.r—not reported. Abbreviations: E. granulosus—Echinococcus granulosus, L. donovani—Leishmania donovani, L. major—Leishmania major, L. spp.—Leishmania species, L. tropica—Leishmania tropica, and Helminths—Parasitic worms.
In 2021, essential oil from C. sativa inflorescence achieved a remarkable 96.012% cure rate for L. tropica (KWH23) in mice, comparable to amphotericin [37,61]. Similarly, a study conducted in 2020 showed that essential oil extracted by hydrodistillation showed significant inhibitory zones against Candida spp., as previously reported. Chemical synthesis in 2020 involving various compounds showed promising IC50 values against P. falciparum and L. donovani, highlighting their potential as antiparasitic agents [61].
Several symptoms of parasitic diseases stem from parasites residing in various tissues of the host. The drug’s ability to penetrate these specific tissues, where the infection is localized, is crucial for successful treatment. Therefore, to enhance established dosing regimens, it is essential to consider not only plasma pharmacokinetics but also how the drug distributes in tissues affected by parasitic infection [99]. Helminths’ outer surfaces act as multifunctional shields crucial for protecting the organism from its external environment. The tegumental syncytium in flatworms fulfills essential roles in nutrient uptake, immune defense, osmoregulation, and structural support. Lipophilic anthelmintic compounds exploit the permeability of this external barrier, accumulating within the parasite via passive diffusion. CBD, characterized by its lipophilicity and ability to modulate membrane permeability, interacts uniquely with the surface structure of helminths. This interaction likely facilitates CBD’s entry into protoscoleces and cysts, thereby enhancing its efficacy as an anthelmintic agent [27].
Essential oils derived from industrial hemp inflorescences, containing cannabinoids such as CBD, δ9-THC, and CBC, demonstrate significant antiparasitic activity against Leishmania tropica in an in vivo model [100]. The effectiveness of these essential oils in reducing tissue lesions suggests their potential application as alternative or complementary therapies for leishmaniasis. Further research is warranted to elucidate the specific contributions of these cannabinoids to the observed antiparasitic effects and to optimize their therapeutic use in clinical settings [61,67].
Gold (Au), silver (Ag), and gold-silver bimetallic (Au-Ag) nanoparticles synthesized using C. sativa leaf extract show antiparasitic activity against Leishmania major. The Ag nanoparticles displayed the highest activity, followed by the bimetallic Au-Ag nanoparticles, and then the Au nanoparticles. These findings suggest that nanoparticles synthesized using C. sativa can be potent anti-leishmanial agents, warranting further investigation into their therapeutic potential and mechanisms of action [78].
The δ8-THC derivative (Compound 14) exhibits highly potent antileishmanial activity against L. donovani promastigotes, surpassing Amphotericin B. This suggests δ8-THC derivatives are promising therapeutic agents for leishmaniasis [100].
The study suggests that C. sativa use among Aka foragers, farmers of the central African republic, is associated with reduced helminth infection and reinfection rates [85]. This supports the hypothesis that C. sativa may be used as a form of self-medication against parasitic infections, although the Aka do not explicitly use C. sativa for this purpose. Further research is needed to explore the potential antiparasitic properties of C. sativa and its active compounds in more detail [85].
Cannabinoids exhibit significant potential as antileishmanial agents based on in silico molecular docking studies. These findings warrant further investigation through in vitro and in vivo studies to validate the efficacy and safety of these compounds for potential therapeutic applications against Leishmania infections [98].
3.1.5. Comprehensive Analysis of the Main Pathogens Species
A broad spectrum of antimicrobial activity against various pathogens was reported in the studies summarized in Table 1, Table 2, Table 3, Table 4 and Table 5. Figure 3 highlights the specific pathogens within each group and the relative emphasis given to each one of them, by percentages of main pathogen species found in the literature targeting C. sativa. Key highlights include
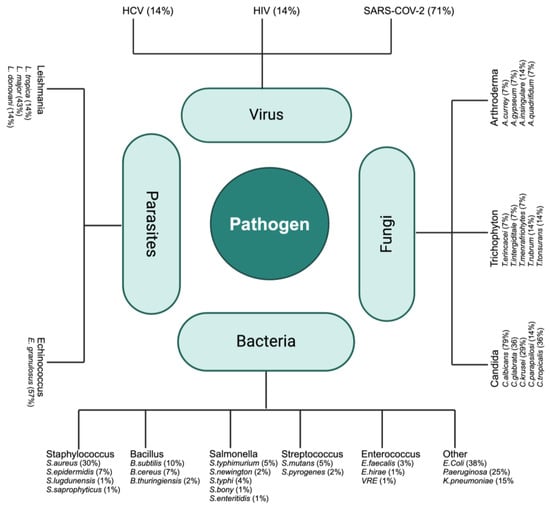
Figure 3.
Main pathogen species found in the literature targets of C. sativa.
- Clinical Data: Emphasis on high-impact viruses, including SARS-CoV-2, HIV, and HCV.
- Microbiological Data: Extensive research focus on bacterial pathogens such as E. coli and S. aureus (MRSA including), as well as fungal pathogens like C. albicans.
- Experimental Data: Significant studies on parasitic infections, particularly those caused by Leishmania and Echinococcus species (responsible for hydatid disease, which poses significant health challenges, especially in endemic regions).
3.2. Cannabinoids Against Multidrug-Resistant Pathogens
According to the WHO, the AMR occurs when bacteria, viruses, fungi, and parasites no longer respond to medicines, making people sicker and increasing the risk of spread of infections that are difficult to treat, illness and deaths. AMR is driven largely by the misuse and overuse of antimicrobials, yet, at the same time, many people around the world do not have access to essential antimicrobial medicines [101].
In this context, it has been demonstrated that cannabinoids exhibit antimicrobial properties that may be potent against AMR; Bahman et al. (2016) outlined four main mechanisms through which bacteria acquire antibiotic resistance. First, bacteria reduce the permeability of their outer membrane, hindering antibiotic penetration. Second, they alter their internal biochemical pathways to diminish or neutralize the antibiotic’s effects once inside the cell. Third, bacteria produce enzymes that specifically break down antibiotic compounds. Lastly, they undergo structural changes that modify the sites where antibiotics typically exert their action [102]. Furthermore, repeated use of conventional antibiotics can induce genetic mutations in microorganisms, leading to the emergence of resistant strains [103]. Among the 15 families of antibiotic-resistant bacterial pathogens, particular attention is given to Gram-negative bacteria resistant to last-resort antibiotics, drug-resistant Mycobacterium tuberculosis, and other high-burden resistant pathogens such as Salmonella, Shigella, Neisseria gonorrhoeae, P. aeruginosa, and S. aureus [3].
Clinical Trials and Clinical Research on C. sativa and Cannabinoids
Eight clinical trials addressing the safety and efficacy of Cannabis in infectious diseases were found from 2014 to 2024. Most of the reported trials investigated the effect of Cannabis on viral infections. Only two viral infections were studied: SARS-CoV-2 and HIV. Among these, three are completed, two are recruiting, and one is withdrawn. Regarding the trials on COVID-19 (n = 2), they focused on the effect of Cannabis in relieving symptoms such as inflammation and pain. Interestingly, one trial investigated the role of CBD in managing long COVID-19 symptoms. Another trial examined the rate of infection in Cannabis users versus non-users.
Regarding trials on HIV (n = 4), most investigated the effect of Cannabis, medical or recreational, on the severity of symptoms. One study evaluated the difference in effects based on the content of CBD or THC. However, this study was withdrawn—stopped before recruiting its first participant.
Only one trial addressed a bacterial infection. This trial investigated the efficacy of Cannabis oil in the treatment of hidradenitis suppurativa, a skin condition characterized by inflammation of the hair follicles, most commonly in the inguinal, anogenital, and axillary regions. This study was withdrawn in phase II.
From the retrieved data, it is noticeable that a significant portion of clinical trials investigating the role of Cannabis in managing infectious diseases focused on its role in controlling symptoms such as pain and inflammation. Interestingly, one article considered the antiviral effect by evaluating antiretroviral adherence and HIV viral load suppression. The same was found in the literature search, where the mechanisms of action of cannabinoids were elucidated. Nonetheless, it has been shown that Cannabis and related products can have significant potential as antimicrobials (Table 1, Table 2, Table 3 and Table 4).
Therefore, it is important to consider the antimicrobial properties for future clinical trials. Regarding fungal and parasitic infections, no clinical trials were found. Comparing this to the data obtained from the literature search, there is significant potential for Cannabis to be investigated in treating many conditions related to infections, such as those found in the table of fungal and parasitic infections.
3.3. Potential Pharmacological Effects of Antimicrobial Cannabinoids
There is a wide variety of conditions caused by infectious agents. These conditions vary in severity and scope depending on several factors and can be classified according to the pathogenic agent. Having that said there are four main classes of infectious diseases: antibacterial, antifungal, antiviral, and antiparasitic. Furthermore, pathological conditions can be divided according to the main body sites or systems affected by infection. This section aims to study the pharmacological potentials of cannabinoids especially with the increased concern over antibiotic resistance and potential global public health impact.
3.3.1. The Respiratory System
Cannabinoids were found to have significant antimicrobial activity. For example, CBD has demonstrated significant antibacterial activity against high-priority pathogens, including antibiotic-resistant ones, which pose global concerns particularly in healthcare settings. This suggests potential for treating healthcare-associated respiratory infections caused by Klebsiella pneumoniae, Staphylococcus pneumoniae, Pseudomonas aeruginosa, and S. aureus, which can lead to nosocomial pneumonia [21,28,33,37,54].
These infections pose significant challenges, particularly in children, the elderly, and immunocompromised individuals, who exhibit resistance to multiple antibiotics, including strains resistant to carbapenems, cephalosporins, methicillin, and macrolides [3]. Of particular concern in pediatric patients is the occurrence of drug-resistant Enterobacterales infections, such as extended-spectrum β-lactamase-producing K. pneumoniae, which complicate initial antibiotic therapies [3].
Furthermore, CBD has demonstrated antibacterial activity against susceptible, isoniazid-resistant and rifampicin-resistant M. tuberculosis (RR-TB) [28]. RR-TB is classified as critically important due to its virulence, ability to transfer resistance genes, and its significant global burden, especially in developing countries [3].
Regarding viral infections, COVID-19 quickly became a leading cause of death, ranking third globally in 2020 and second in 2021 [5]. Cannabinoids, especially CBD, show potential in managing COVID-19 through their immune-modulating, anti-inflammatory, and possible antiviral properties [91]. Cannabis compounds have shown potential in inhibiting SARS-CoV-2 by acting on multiple steps of the viral life cycle. These compounds exhibit anti-inflammatory properties, which are beneficial in reducing inflammation associated with SARS-CoV-2 and HIV [96]. CBD can inhibit pro-inflammatory cytokines, modulate ACE2 expression, and potentially reduce viral replication and inflammation. The CB2 receptor also plays a significant role in immune response modulation, making it a key target for cannabinoid-based therapies in these diseases [96].
3.3.2. The Digestive System
Studies indicate that CBD exhibits anti-inflammatory and antibacterial effects in the gastrointestinal tract, showing promising applications in the treatment of Helicobacter pylori infection [104]. Furthermore, another study identified CBD as an effective inhibitor of extracellular vesicle release in both human cancer cells and the intestinal parasite Giardia intestinalis [105].
3.3.3. The Genitourinary System
CBD exhibits significant antibacterial activity against pathogens causing urinary tract infections (UTIs), such as E. coli [28], which has recently been moved from the 4th to the 2nd position in the 2024 WHO Priority Pathogen List. This is due to its resistance to third-generation cephalosporins and carbapenems [3]. While E. coli remains the most prevalent cause of cystitis and pyelonephritis, K. pneumoniae, which can also cause these infections and presents a high mortality rate, has been moved from the 5th to the 1st position on the same list due to its carbapenem resistance [3]. The inclusion of these pathogens underscores the need for enhanced surveillance and stricter control measures to address these emerging bacterial strains.
Further, CDB showed promising antibacterial activities in combination with other traditional antibiotics such as colistin and PB. Colistin is a peptide antibiotic, applied in the eliminations with Gram-negative bacilli mainly P. aeruginosa. Resistance to colistin mediated by MCR-1 pose a significant challenge in medicine [106]. Therefore, combining the two molecules might lead to promising clinical results. Also, while CBD alone shows limited efficacy against Neisseria gonorrhoeae, with a relatively high MIC of 256 µg/mL, its combination with PB could enhance its antibacterial activity [28].
3.3.4. Skin and Soft Tissue
CBD has shown significant efficacy against skin-infecting bacteria such as resistant S. aureus, P. acnes, and C. acnes [49], making it relevant for treating skin and soft tissue infections such as cellulitis, abscesses and acne.
The skin’s endocannabinoid system (ECS), involving CB1R and CB2R receptors, play a pivotal role in regulating cutaneous functions. Therefore, the dysregulation of the ECS can contribute to various skin disorders, including acne [49]. While conventional therapies for acne are effective, they are associated with adverse effects and the risk of antibiotic resistance. Cannabis-derived compounds, particularly CBD, demonstrate anti-inflammatory, antiproliferative, and antimicrobial properties, positioning them as promising candidates for acne management. CBD is particularly effective against P. acnes and C. acnes, which are the main contributors to acne infection and exacerbation [40,72].
Preclinical studies highlight cannabinoids’ capacity to normalize lipogenesis, reduce inflammation, and inhibit bacterial growth, all critical factors in acne pathogenesis. Ongoing clinical trials are assessing the safety and efficacy of topical cannabinoid formulations. Brands like Nordic Oil and cbdMD have already introduced CBD-based skincare products targeting acne treatment [107].
3.3.5. Systemic Infections
Bacterial priorities from the list by the WHO, such as sepsis and septicemia, pose significant challenges due to resistant strains like MRSA. The ability of CBD to combat resistant bacterial strains, including S. aureus, suggests its potential in the treatment of severe systemic infections such as septicemia and endocarditis. Table 1 shows that CBD exhibits MIC values as low as 4 µg/mL against certain strains of S. aureus, indicating its strong antibacterial properties. Also, the promising results of CBD in reducing bacterial load and combating resistance is highlighted in the literature [2,40,43].
3.3.6. The Central Nervous System
CBD can be investigated for use against vector-borne pathogens, such as those responsible for malaria, dengue, and Lyme disease, due to its anti-inflammatory and immunomodulatory properties. Although specific data for vector-borne pathogens are not provided, the ability of CBD to combat Gram-negative bacteria like Neisseria meningitidis suggests promising future applications. The document reports a MIC of 128 µg/mL for CBD against N. meningitidis, with enhanced efficacy when combined with Polymyxin B, indicating its potential as an antibacterial agent in central nervous system infections [28].
3.3.7. Oral and Dental Infectious
The spectra of oral and dental infections include chronic tooth decay, oral cancers, gingivitis, periodontal diseases such as periodontitis, noma, oro-dental trauma, and oral manifestations of HIV, along with issues like sensitive, cracked, or chipped teeth. Furthermore, congenital anomalies like cleft lip and palate are widespread, presenting significant public health challenges in many countries [108]. Therefore, preventing and controlling the spread of this health issue is a global challenge, requiring increased efforts and potentially innovative approaches to address this problem [4,109]. A clinical trial on a gel containing C. sativa compounds highlights the potential of cannabinoids in managing oral and dental infections [41,42]. Due to their antimicrobial, anti-inflammatory, and analgesic properties, CBD can be considered an excellent candidate for treating periodontal infections [10]. The same study highlights the distribution of cannabinoid receptors in the oral mucosa, indicating that these compounds have regenerative effects and can influence the expression of COX-2, which is crucial for inflammation modulation [41].
3.3.8. Ophthalmic Infections
The relationship between bacterial infections and other diseases, including cancer, is an active area of research [110]. Effective primary prevention strategies are required to prevent future impacts on health systems [110,111]. Table 1 shows that CBD exhibits significant activity against key bacterial pathogens such as S. aureus and P. aeruginosa. These findings indicate that CBD could be explored for managing conditions like conjunctivitis and uveitis [40,41,42,80].
3.4. Future Directions and Therapeutic Potential
Cannabinoids have shown promising antimicrobial properties that could be leveraged to combat AMR [19,20,29,70]. Future research should focus on elucidating the mechanisms through which cannabinoids inhibit microbial growth, especially against high-priority pathogens such as Gram-negative bacteria, M. tuberculosis, and other resistant strains.
According to Joseph et al. currently, scientists are working on developing NPs, with the main task of fighting various infections [74]. These findings highlight the potential of C. sativa-mediated NPs as powerful antimicrobial agents. Future research should aim to standardize methodologies and conduct comprehensive clinical trials to fully realize and optimize their therapeutic potential. Clinical trials should be designed to assess the efficacy of cannabinoids in treating infections caused by these resistant pathogens.
3.4.1. Clinical Trials and Applications
Our review highlights the need for more clinical trials investigating the therapeutic potential of cannabinoids in infectious diseases. As shown in Figure 1 above, current data are predominantly focused on viral infections, with limited trials exploring bacterial, fungal, and parasitic infections. Future clinical research should aim to evaluate the effectiveness of cannabinoids in managing a broader range of infections, particularly those involving multidrug-resistant organisms. Additionally, trials should consider the role of cannabinoids in combination therapies, potentially enhancing the efficacy of existing antimicrobial agents.
3.4.2. Mechanisms of Action and Pharmacological Effects
Understanding the pharmacological effects of cannabinoids is crucial for their therapeutic application. Cannabinoids have demonstrated significant antimicrobial activity against pathogens affecting various body systems, including the respiratory [21,28,33,37,54,105,112,113], digestive, genitourinary, and central nervous systems. Future research should aim to further delineate the mechanisms of action, including the modulation of immune responses, inhibition of microbial growth, and potential synergy with other antimicrobial agents. Investigating these mechanisms will provide insights into optimizing cannabinoid-based therapies for clinical use.
3.4.3. Challenges and Regulatory Considerations
Despite the promising potential of cannabinoids, there are several challenges that need to be addressed [114]. These include the variability in cannabinoid content and potency, the need for standardized formulations, and the regulatory landscape governing the use of Cannabis-derived products [114,115]. Future research should focus on developing standardized and reproducible cannabinoid formulations with consistent therapeutic effects. Additionally, engaging with regulatory bodies to streamline the approval process for cannabinoid-based therapies will be essential for their successful integration into clinical practice.
3.5. Limitations of This Study and Future Research
While our systematic review provides valuable insights into the antimicrobial potential of cannabinoids, several limitations must be acknowledged. First, the heterogeneity of the included studies, in terms of methodologies, cannabinoid formulations, and microbial strains tested, may introduce variability that complicates direct comparisons. Second, many studies were preclinical, with limited clinical data available, thus restricting the generalizability of the findings to human populations. Third, publication bias could influence the results, as studies showing significant effects are more likely to be published. Finally, the regulatory status and legal variations regarding Cannabis use across different regions may impact the feasibility of research and implementation of cannabinoid-based therapies.
Future research should expand beyond these criteria to include older studies, non-English publications, and data from multiple databases. This broader approach will help to uncover a more comprehensive understanding of the antimicrobial potential of cannabinoids and their applications. Additionally, future systematic reviews should aim to include a wider variety of methodologies, cannabinoid formulations, and microbial strains to provide a more holistic view of the field and pave the way for the effective integration of cannabinoids into antimicrobial therapy.
4. Discussion
The WHO Bacterial Priority Pathogens List (BPPL) for 2024 outlines critical, high, and medium-priority bacterial pathogens that pose significant threats to public health due to their resistance to antibiotics. The WHO BPPL categorizes bacterial pathogens based on their resistance profiles and public health impact. The critical priority group includes bacteria such as: carbapenem-resistant A. baumannii, carbapenem-resistant P. aeruginosa, carbapenem-resistant Enterobacterales, vancomycin-resistant E. faecium, and methicillin-resistant MRSA. This emphasizes the urgent need for new antimicrobial agents, highlighting the potential role of Cannabis-based treatments in addressing these resistant bacterial infections.
From the current review of the scientific literature published from 2014 to 2024 (Table 1, Table 2, Table 3 and Table 4), the antibacterial activity of cannabinoids against several microorganisms is demonstrated. Cannabinoids (CBD, CBG, Δ9-THC, THCV, CBN) tested against Gram-positive bacteria have shown effectiveness against MRSA, which is a high-priority pathogen on the WHO list. Additionally, CBD and CBG were tested against E. faecium, showing potential effectiveness against VRE.
Concerning Gram-negative bacteria, P. aeruginosa, a critical priority pathogen, has shown susceptibility to cannabinoids like CBD and CBG. Furthermore, both cannabinoids (CBD and CBG) have demonstrated activity against E. coli, including multidrug-resistant strains, which are the second biggest global problem on the WHO BPPL.
Thus, cannabinoids may represent a new class of antibacterial agents, potentially effective due to their unique in diverse mechanisms of action compared to traditional antibiotics. This can be crucial in the fight against AMR. The effectiveness of cannabinoids against both Gram-positive and Gram-negative bacteria suggests promising applications in broad-spectrum antibacterial therapies.
Despite the results being encouraging, this analysis highlights several limitations, such as incomplete data. For example, some studies lack detailed information on the specific parts of the C. sativa plant used, extraction methods, the exact compositions of cannabinoid extracts or the positive control. Additionally, standardized methodologies in future studies are necessary to better compare the efficacy of cannabinoids with existing antibiotics. Also, comprehensive studies and clinical trials are required to fully understand the potential of cannabinoids in treating bacterial infections, especially those caused by high-priority pathogens.
Concerning the potential of cannabinoids as antifungal agents, they demonstrate activity against several priority fungi listed by the WHO BPPL-2022. The list includes 19 fungal entities in three priority groups (critical, high, and medium) to prioritize research efforts, especially those in the critical group, such as C. neoformans [116]. Tests with Q-Δ8-THC showed a MIC of 0.7 µg/mL, comparable to Amphotericin B (2.5 µg/mL), important due to the high mortality in immunocompromised patients. For A. fumigatus, tests with PMBA showed a MIC of 39.68 µg/mL. Other cannabinoids presented varied results. For C. albicans, tests with CBD showed MICs ranging from 12.5 to 100 µg/mL. Furthermore, CBD was effective in inhibiting biofilm formation, suggesting significant therapeutic potential, as this fungus is included in the critical group with high priority, with concerns about resistance and biofilms.
Additionally, tests with CBD and other cannabinoids showed activity with MICs approximately 6.25–10.0 µg/mL against C. glabrata. Furthermore, extracts including cannabinoids showed activity against C. tropicalis (tests with PMBA showed MIC approximately 15.75 µg/mL; other cannabinoids presented MICs up to 390.6 µg/mL) and C. parapsilosis (tests with PMBA showed MIC less than 6.25 µg/mL; other cannabinoids presented MICs up to 62.99 µg/mL), notably important due to hospital infections and fluconazole resistance. Against Fusarium spp., ZnO nanoparticles showed inhibition. However, more inhibition was observed in Ag-doped ZnO nanoparticles. Surprisingly, no specific data are available in the analyzed studies on the efficacy of cannabinoids against C. auris, Mucorales, or Histoplasma spp.
Table 3 and Table 4 demonstrated the potential efficacy of cannabinoids against priority viral and parasitic pathogens. Regarding HIV, in vivo clinical trials showed a significant reduction in T-cell activation (CD8+CD38+HLA-DR+), decreased plasma IL-6 and TNF-α levels, reduced total and integrated HIV DNA, and changes in gut microbiome composition. Cannabinoids were found effective against a set of parasitic infections such as leishmaniasis and echinococcosis. For leishmaniasis, cannabinoids significantly inhibited parasitic replication in both in vitro and in vivo studies. Additionally, for echinococcosis, CBD significantly reduced cyst weight and altered the germinal layer, indicating potential future applications. These findings highlight the promising role of cannabinoids in the treatment of these critical infections.
Nano-sized pharmaceutical dosage forms, such as NPs for targeted transport, offer enhanced or additional functionality of Cannabis-based products [77,79,82]. These features include improved efficacy, bioavailability, and reduced side effects, thereby minimizing toxicity. The process of nanoencapsulation forms drug-loaded particles ranging from 1 to 1000 nm, allowing drugs to be absorbed directly from the surface of the NPs or enclosed within the particle [74]. This technology enhances the antimicrobial properties of the NPs, providing a novel approach to combat resistant bacterial strains and other pathogens [74].
Overall, as demonstrated in Figure 3, S. aureus is the most prominent, accounting for 30% of the bacterial targets. This includes MRSA, a significant concern in clinical settings due to its resistance to multiple antibiotics. Among fungi, Candida species are the most frequently targeted, with C. albicans representing 16%. This is crucial given the prevalence of Candida infections and the increasing resistance to conventional antifungal therapies. Notably, the most significant antiviral activity is against SARS-CoV-2, comprising 71% of the viral targets. This highlights the potential role of cannabinoids in managing COVID-19. In the category of parasites, E. granulosus is notably targeted by cannabinoids, representing 57% of the parasitic pathogens. This parasite is responsible for hydatid disease, which poses significant health challenges, especially in endemic regions, which struggle against neglected diseases.
While cannabinoids offer promising potential as novel antimicrobial agents, further research and standardized approaches are essential to fully harness their capabilities. Standardization of study methodologies and the conduction of clinical trials are necessary to validate these findings and develop cannabinoid-based treatments for priority infections. Therefore, the antimicrobial properties of C. sativa should be considered in future clinical trials, investigating its potential in treating various infection-related conditions, including those found in these studies and other emerging infections.
5. Conclusions
Cannabinoids derived from Cannabis sativa L. exhibit promise in the realm of infectious diseases, particularly in combating multidrug-resistant pathogens. The properties and applications of Cannabis align closely with the priorities established by the WHO and support the United Nations Millennium Development Goals and the Sustainable Development Goals, especially Goal 3: Ensure healthy lives and promote well-being for all at all ages. The ability of Cannabis to combat antimicrobial-resistant infections, potentially in combination with traditional antimicrobials, could substantially contribute to global health by providing novel treatment avenues and reducing the burden of infectious diseases worldwide.
Cannabis extracts and cannabinoids have demonstrated the capacity to inhibit the growth of certain bacterial strains at concentrations comparable to traditional antimicrobials. These findings represent a significant advancement in the battle against antimicrobial resistance, offering a perspective for future treatments. Notably, eight clinical trials addressing the safety and efficacy of Cannabis in infectious diseases were identified between 2014 and 2024. Most trials focused on viral infections, specifically SARS-CoV-2 and HIV. Among these, three are completed, two are recruiting, and one is withdrawn. Trials on COVID-19 primarily explored Cannabis’s role in alleviating symptoms such as inflammation and pain, with one investigating CBD’s efficacy in managing long COVID-19 symptoms.
Regarding HIV, most trials examined Cannabis’s impact on symptom severity, with one study evaluating the differential effects of CBD and THC content, although this study was withdrawn before recruitment. A single trial addressed a bacterial infection, investigating Cannabis oil’s efficacy in treating hidradenitis suppurativa, which was also withdrawn in phase II. These data suggest a significant portion of clinical trials on Cannabis for infectious diseases focused on symptom management, such as pain and inflammation, rather than direct antimicrobial effects. Nonetheless, the literature suggests that Cannabis and related products have substantial potential as antimicrobials.
Future clinical trials should prioritize investigating the antimicrobial properties of Cannabis, including its potential efficacy against fungal and parasitic infections, which have yet to be explored. Data from preclinical studies indicate a need for comprehensive clinical investigations to evaluate Cannabis’s full potential in treating a wide range of infectious diseases. Emphasizing these antimicrobial properties in future research could pave the way for innovative treatments and contribute significantly to global efforts against antimicrobial resistance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/futurepharmacol4030033/s1, Figure S1: PRISMA 2020 Checklist; Table S1: Search Strategies and Keywords According to PRISMA Guidelines.
Author Contributions
Conceptualization, A.R. and H.C.-M.; investigation, methodology, and data collection and data analysis, A.R., R.A., D.O., and R.L., software, A.R. and R.A., validation, visualization, A.R., R.A., and H.C.-M., writing—original draft preparation, A.R., and R.A.; writing—review and editing, A.R., R.A., D.O., R.L. and H.C.-M.; supervision, H.C.-M.; project administration, H.C.-M.; funding acquisition, H.C.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by FCT—Foundation for Science and Technology, I.P., through National Funds, under the projects UID/DTP/04138/2021.
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are unavailable due to privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Russo, E.B. Taming THC: Potential Cannabis Synergy and Phytocannabinoid-terpenoid Entourage Effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Peters, M.; Murillo-Rodriguez, E.; Hanuš, L.O. Cannabidiol—Recent Advances. Chem. Biodivers. 2007, 4, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2024; Volume 898, pp. 1–72. [Google Scholar]
- Ribeiro, A.; Serrano, R.; da Silva, I.B.M.; Gomes, E.T.; Pinto, J.F.; Silva, O. The Genus Diospyros: A Review of Novel Insights into the Biological Activity and Species of Mozambican Flora. Plants 2023, 12, 2833. [Google Scholar] [CrossRef]
- World Health Organization. World Health Statistics 2024: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: Geneva, Switzerland, 2024. [Google Scholar]
- Butler, M.S.; Henderson, I.R.; Capon, R.J.; Blaskovich, M.A.T. Antibiotics in the Clinical Pipeline as of December 2022. J. Antibiot. 2023, 76, 431–473. [Google Scholar] [CrossRef]
- Tang, K.W.K.; Millar, B.C.; Moore, J.E. Antimicrobial Resistance (AMR). Br. J. Biomed. Sci. 2023, 28, 11387. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Kavanagh, A.M.; Elliott, A.G.; Zhang, B.; Ramu, S.; Amado, M.; Lowe, G.J.; Hinton, A.O.; Pham, D.M.T.; Zuegg, J.; et al. The Antimicrobial Potential of Cannabidiol. Commun. Biol. 2021, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Caltana, L.R.; Brusco, A. Cannabinoids: A Group of Promising Neuroprotective Agents. In Neuroprotective Natural Products; Wiley: Hoboken, NY, USA, 2017; pp. 321–339. [Google Scholar]
- Pacher, P.; Bátkai, S.; Kunos, G. The Endocannabinoid System as an Emerging Target of Pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef]
- Oz, M.; Yang, K.-H.S.; Mahgoub, M.; Petroianu, G.; Lorke, D.E. Effects of Cannabidiol and Other Phytocannabinoids on Voltage-and Ligand-Gated Ion Channels. In Cannabis Use, Neurobiology, Psychology, and Treatment; Elsevier: Amsterdam, The Nederlands, 2023; pp. 445–456. [Google Scholar]
- Shahbazi, F.; Grandi, V.; Banerjee, A.; Trant, J.F. Cannabinoids and Cannabinoid Receptors: The Story so Far. iScience 2020, 23, 101301. [Google Scholar] [CrossRef]
- Hunter, M.R.; Finlay, D.B.; Glass, M. Signaling and Regulation of the Cannabinoid CB1 Receptor. In Handbook of Cannabis and Related Pathologies; Elsevier: Amsterdam, The Nederlands, 2017; pp. 564–572. [Google Scholar]
- Brunt, T.M.; Bossong, M.G. The Neuropharmacology of Cannabinoid Receptor Ligands in Central Signaling Pathways. Eur. J. Neurosci. 2022, 55, 909–921. [Google Scholar] [CrossRef]
- Marzo, V.D. Cannabinoids; Endocannabinoid Research Group, Ed.; John Wiley & Sons: Pozzuoli, Italy, 2014; ISBN 1118451279/9781118451274. [Google Scholar]
- Dhopeshwarkar, A.; Murataeva, N.; Makriyannis, A.; Straiker, A.; Mackie, K. Two Janus Cannabinoids That Are Both CB2 Agonists and CB1 Antagonists. J. Pharmacol. Exp. Ther. 2017, 360, 300–311. [Google Scholar] [CrossRef]
- Kaszyńska, A.A. The Role of the Endocannabinoid System in the Pathogenesis of Degenerative Diseases. Neuropsychiatr. I Neuropsychol. 2022, 17, 140–144. [Google Scholar] [CrossRef]
- Karas, J.A.; Wong, L.J.M.; Paulin, O.K.A.; Mazeh, A.C.; Hussein, M.H.; Li, J.; Velkov, T. The Antimicrobial Activity of Cannabinoids. Antibiotics 2020, 9, 406. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Khan, T.A.; Ali, I.; Hassan, A.; Ali, H.; Din, Z.U.; Hassan, Z.; Tabassum, S.; Saqib; Majid, A.; et al. In Vitro Antibacterial Activity of Cannabis sativa Leaf Extracts to Some Selective Pathogenicbacterial Strains. Int. J. Biosci. (IJB) 2014, 4, 65–70. [Google Scholar]
- Kaur, S.; Sharma, N.; Roy, A. Role of Cannabinoids in Various Diseases: A Review. Curr. Pharm. Biotechnol. 2022, 23, 1346–1358. [Google Scholar] [PubMed]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial Cannabinoids from Cannabis sativa: A Structure−Activity Study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef]
- Wassmann, C.S.; Højrup, P.; Klitgaard, J.K. Cannabidiol Is an Effective Helper Compound in Combination with Bacitracin to Kill Gram-Positive Bacteria. Sci. Rep. 2020, 10, 4112. [Google Scholar] [CrossRef]
- Donthu, N.; Kumar, S.; Mukherjee, D.; Pandey, N.; Lim, W.M. How to Conduct a Bibliometric Analysis: An Overview and Guidelines. J. Bus. Res. 2021, 133, 285–296. [Google Scholar] [CrossRef]
- Blebea, N.M.; Pricopie, A.I.; Vlad, R.-A.; Hancu, G. Phytocannabinoids: Exploring Pharmacological Profiles and Their Impact on Therapeutical Use. Int. J. Mol. Sci. 2024, 25, 4204. [Google Scholar] [CrossRef]
- Silhavy, T.J.; Kahne, D.; Walker, S. The Bacterial Cell Envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef]
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and Resistance Mechanisms of Antibiotics: A Guide for Clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300. [Google Scholar] [CrossRef]
- Albani, C.M.; Fuentes, G.; Ramírez, C.L.; Pensel, P.E.; Gatti, F.; Albanese, A.; Nutter, D.; Aguirre, M.E.; Di Iorio, Y.D.; Elissondo, M.C. Anthelmintic Effect of Cannabidiol against Echinococcus granulosus Sensu Stricto. Trop. Med. Infect. Dis. 2024, 9, 35. [Google Scholar] [CrossRef]
- Abichabki, N.; Zacharias, L.V.; Moreira, N.C.; Bellissimo-Rodrigues, F.; Moreira, F.L.; Benzi, J.R.L.; Ogasawara, T.M.C.; Ferreira, J.C.; Ribeiro, C.M.; Pavan, F.R.; et al. Potential Cannabidiol (CBD) Repurposing as Antibacterial and Promising Therapy of CBD plus Polymyxin B (PB) against PB-Resistant Gram-Negative Bacilli. Sci. Rep. 2022, 12, 6454. [Google Scholar] [CrossRef]
- Li, H.; Zhao, Q.; Chang, S.; Wang, L.; Zhao, B. Phytochemical Analysis and Bioactivity of Different Ethanolic Extracts from Cannabidiol Full-Spectrum Oil. J. Mol. Liq. 2023, 372, 121173. [Google Scholar] [CrossRef]
- Cortes, E.; Mora, J.; Márquez, E. Modelling the Anti-Methicillin-Resistant Staphylococcus aureus (MRSA) Activity of Cannabinoids: A QSAR and Docking Study. Crystals 2020, 10, 692. [Google Scholar] [CrossRef]
- Martinenghi, L.D.; Jønsson, R.; Lund, T.; Jenssen, H. Isolation, Purification, and Antimicrobial Characterization of Cannabidiolic Acid and Cannabidiol from Cannabis sativa L. Biomolecules 2020, 10, 900. [Google Scholar] [CrossRef] [PubMed]
- Gildea, L.; Ayariga, J.A.; Ajayi, O.S.; Xu, J.; Villafane, R.; Samuel-Foo, M. Cannabis sativa CBD Extract Shows Promising Antibacterial Activity against Salmonella Typhimurium and S. Newington. Molecules 2022, 27, 2669. [Google Scholar] [CrossRef]
- Chelminiak-Dudkiewicz, D.; Smolarkiewicz-Wyczachowski, A.; Mylkie, K.; Wujak, M.; Mlynarczyk, D.T.; Nowak, P.; Bocian, S.; Goslinski, T.; Ziegler-Borowska, M. Chitosan-Based Films with Cannabis Oil as a Base Material for Wound Dressing Application. Sci. Rep. 2022, 12, 18658. [Google Scholar] [CrossRef]
- Tiwari, P.; Sharma, V.; Kaur, N.; Ahmad, K.; Mobin, S.M. Sustainable Graphene Production: New Insights into Cannabis sativa Engineered Carbon Dots Based Exfoliating Agent for Facile Production of Graphene. ACS Sustain. Chem. Eng. 2019, 7, 11500–11510. [Google Scholar] [CrossRef]
- Marini, E.; Magi, G.; Ferretti, G.; Bacchetti, T.; Giuliani, A.; Pugnaloni, A.; Rippo, M.R.; Facinelli, B. Attenuation of Listeria Monocytogenes Virulence by Cannabis sativa L. Essential Oil. Front. Cell. Infect. Microbiol. 2018, 8, 293. [Google Scholar] [CrossRef]
- Iseppi, R.; Brighenti, V.; Licata, M.; Lambertini, A.; Sabia, C.; Messi, P.; Pellati, F.; Benvenuti, S. Chemical Characterization and Evaluation of the Antibacterial Activity of Essential Oils from Fibre-Type Cannabis sativa L. (Hemp). Molecules 2019, 24, 2302. [Google Scholar] [CrossRef]
- Palmieri, S.; Maggio, F.; Pellegrini, M.; Ricci, A.; Serio, A.; Paparella, A.; Lo Sterzo, C. Effect of the Distillation Time on the Chemical Composition, Antioxidant Potential and Antimicrobial Activity of Essential Oils from Different Cannabis sativa L. Cultivars. Molecules 2021, 26, 4770. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, K.; Stahl, V. Cannabinoids Infused Mouthwash Products Are as Effective as Chlorhexidine on Inhibition of Total-Culturable Bacterial Content in Dental Plaque Samples. J. Cannabis Res. 2020, 2, 20. [Google Scholar] [CrossRef]
- Aqawi, M.; Sionov, R.V.; Friedman, M.; Steinberg, D. The Antibacterial Effect of Cannabigerol toward Streptococcus Mutans Is Influenced by the Autoinducers 21-CSP and AI-2. Biomedicines 2023, 11, 668. [Google Scholar] [CrossRef] [PubMed]
- Sangkanu, S.; Pitakbut, T.; Phoopha, S.; Khanansuk, J.; Chandarajoti, K.; Dej-adisai, S. A Comparative Study of Chemical Profiling and Bioactivities between Thai and Foreign Hemp Seed Species (Cannabis sativa L.) Plus an In-Silico Investigation. Foods 2023, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Garzón, H.S.; Loaiza-Oliva, M.; Martínez-Pabón, M.C.; Puerta-Suárez, J.; Téllez Corral, M.A.; Bueno-Silva, B.; Suárez, D.R.; Díaz-Báez, D.; Suárez, L.J. Antibiofilm and Immune-Modulatory Activity of Cannabidiol and Cannabigerol in Oral Environments—In Vitro Study. Antibiotics 2024, 13, 342. [Google Scholar] [CrossRef]
- Jirasek, P.; Jusku, A.; Frankova, J.; Urbankova, M.; Diabelko, D.; Ruzicka, F.; Papouskova, B.; Chytilova, K.; Vrba, J.; Havlasek, J.; et al. Phytocannabinoids and Gingival Inflammation: Preclinical Findings and a Placebo-controlled Double-blind Randomized Clinical Trial with Cannabidiol. J. Periodontal Res. 2024, 59, 468–479. [Google Scholar] [CrossRef]
- Malikova, L.; Malik, M.; Pavlik, J.; Ulman, M.; Pechouckova, E.; Skrivan, M.; Kokoska, L.; Tlustos, P. Anti-Staphylococcal Activity of Soilless Cultivated Cannabis across the Whole Vegetation Cycle under Various Nutritional Treatments in Relation to Cannabinoid Content. Sci. Rep. 2024, 14, 4343. [Google Scholar] [CrossRef]
- Stasiłowicz-Krzemień, A.; Szymanowska, D.; Szulc, P.; Cielecka-Piontek, J. Antimicrobial, Probiotic, and Immunomodulatory Potential of Cannabis sativa Extract and Delivery Systems. Antibiotics 2024, 13, 369. [Google Scholar] [CrossRef]
- Kim, E.; Jang, J.; Seo, H.H.; Lee, J.H.; Moh, S.H. Cannabis sativa (Hemp) Seed-Derived Peptides WVYY and PSLPA Modulate the Nrf2 Signaling Pathway in Human Keratinocytes. PLoS ONE 2024, 19, e0298487. [Google Scholar] [CrossRef]
- Casillo, A.; D’Angelo, C.; Imbimbo, P.; Monti, D.M.; Parrilli, E.; Lanzetta, R.; Gomez d’Ayala, G.; Mallardo, S.; Corsaro, M.M.; Duraccio, D. Aqueous Extracts from Hemp Seeds as a New Weapon against Staphylococcus Epidermidis Biofilms. Int. J. Mol. Sci. 2023, 24, 16026. [Google Scholar] [CrossRef]
- Frazzini, S.; Torresani, M.C.; Roda, G.; Dell’Anno, M.; Ruffo, G.; Rossi, L. Chemical and Functional Characterization of the Main Bioactive Molecules Contained in Hulled Cannabis sativa L. Seeds for Use as Functional Ingredients. J. Agric. Food Res. 2024, 16, 101084. [Google Scholar] [CrossRef]
- Serventi, L.; Flores, G.A.; Cusumano, G.; Barbaro, D.; Tirillini, B.; Venanzoni, R.; Angelini, P.; Acquaviva, A.; Di Simone, S.C.; Orlando, G.; et al. Comparative Investigation of Antimicrobial and Antioxidant Effects of the Extracts from the Inflorescences and Leaves of the Cannabis sativa L. Cv. Strawberry. Antioxidants 2023, 12, 219. [Google Scholar] [CrossRef]
- Kesavan Pillai, S.; Hassan Kera, N.; Kleyi, P.; de Beer, M.; Magwaza, M.; Ray, S.S. Stability, Biofunctional, and Antimicrobial Characteristics of Cannabidiol Isolate for the Design of Topical Formulations. Soft Matter 2024, 20, 2348–2360. [Google Scholar] [CrossRef] [PubMed]
- Barbălată-Mândru, M.; Serbezeanu, D.; Butnaru, M.; Rîmbu, C.M.; Enache, A.A.; Aflori, M. Poly(Vinyl Alcohol)/Plant Extracts Films: Preparation, Surface Characterization and Antibacterial Studies against Gram Positive and Gram Negative Bacteria. Materials 2022, 15, 2493. [Google Scholar] [CrossRef] [PubMed]
- Lelario, F.; Scrano, L.; De Franchi, S.; Bonomo, M.G.; Salzano, G.; Milan, S.; Milella, L.; Bufo, S.A. Identification and Antimicrobial Activity of Most Representative Secondary Metabolites from Different Plant Species. Chem. Biol. Technol. Agric. 2018, 5, 13. [Google Scholar] [CrossRef]
- Acquaviva, A.; Cristina Di Simone, S.; Canini, A.; Braglia, R.; Di Marco, G.; Campana, C.; Angelini, P.; Angeles Flores, G.; Venanzoni, R.; Loreta Libero, M.; et al. Phytochemical and Biological Investigations on the Pollen from Industrial Hemp Male Inflorescences. Food Res. Int. 2022, 161, 111883. [Google Scholar] [CrossRef]
- Luz-Veiga, M.; Amorim, M.; Pinto-Ribeiro, I.; Oliveira, A.L.S.; Silva, S.; Pimentel, L.L.; Rodríguez-Alcalá, L.M.; Madureira, R.; Pintado, M.; Azevedo-Silva, J.; et al. Cannabidiol and Cannabigerol Exert Antimicrobial Activity without Compromising Skin Microbiota. Int. J. Mol. Sci. 2023, 24, 2389. [Google Scholar] [CrossRef]
- Antezana, P.E.; Municoy, S.; Orive, G.; Desimone, M.F. Design of a New 3D Gelatin—Alginate Scaffold Loaded with Cannabis sativa Oil. Polymers 2022, 14, 4506. [Google Scholar] [CrossRef]
- Jokić, S.; Jerković, I.; Pavić, V.; Aladić, K.; Molnar, M.; Kovač, M.J.; Vladimir-Knežević, S. Terpenes and Cannabinoids in Supercritical CO2 Extracts of Industrial Hemp Inflorescences: Optimization of Extraction, Antiradical and Antibacterial Activity. Pharmaceuticals 2022, 15, 1117. [Google Scholar] [CrossRef]
- Skala, T.; Kahánková, Z.; Tauchen, J.; Janatová, A.; Klouček, P.; Hubka, V.; Fraňková, A. Medical Cannabis Dimethyl Ether, Ethanol and Butane Extracts Inhibit the in Vitro Growth of Bacteria and Dermatophytes Causing Common Skin Diseases. Front. Microbiol. 2022, 13, 953092. [Google Scholar] [CrossRef]
- Tyagi, P.; Gutierrez, J.N.; Lucia, L.A.; Hubbe, M.A.; Pal, L. Evidence for Antimicrobial Activity in Hemp Hurds and Lignin-Containing Nanofibrillated Cellulose Materials. Cellulose 2022, 29, 5151–5162. [Google Scholar] [CrossRef]
- Sun, Y.; Li, D.; Yu, Y.; Chen, J.; Fan, W. Separation and Characterization of Cellulose Fibers from Cannabis Bast Using Foamed Nickel by Cathodic Electro-Fenton Oxidation Strategy. Polymers 2022, 14, 380. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.; Loveleen, L.; Bhandari, M.; Syed, A.; Bahkali, A.H.; Manchanda, R.; Nimesh, S. Antibacterial, Antioxidant, and Haemolytic Potential of Silver Nanoparticles Biosynthesized Using Roots Extract of Cannabis sativa Plant. Artif. Cells Nanomed. Biotechnol. 2022, 50, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Muscarà, C.; Smeriglio, A.; Trombetta, D.; Mandalari, G.; La Camera, E.; Grassi, G.; Circosta, C. Phytochemical Characterization and Biological Properties of Two Standardized Extracts from a Non-psychotropic Cannabis sativa L. Cannabidiol-chemotype. Phytother. Res. 2021, 35, 5269–5281. [Google Scholar] [CrossRef] [PubMed]
- Menghini, L.; Ferrante, C.; Carradori, S.; D’Antonio, M.; Orlando, G.; Cairone, F.; Cesa, S.; Filippi, A.; Fraschetti, C.; Zengin, G.; et al. Chemical and Bioinformatics Analyses of the Anti-Leishmanial and Anti-Oxidant Activities of Hemp Essential Oil. Biomolecules 2021, 11, 272. [Google Scholar] [CrossRef]
- Aqawi, M.; Sionov, R.V.; Gallily, R.; Friedman, M.; Steinberg, D. Anti-Bacterial Properties of Cannabigerol Toward Streptococcus Mutans. Front. Microbiol. 2021, 12, 656471. [Google Scholar] [CrossRef]
- Russo, C.; Lavorgna, M.; Nugnes, R.; Orlo, E.; Isidori, M. Comparative Assessment of Antimicrobial, Antiradical and Cytotoxic Activities of Cannabidiol and Its Propyl Analogue Cannabidivarin. Sci. Rep. 2021, 11, 22494. [Google Scholar] [CrossRef]
- Pormohammad, A.; Hansen, D.; Turner, R.J. Antibacterial, Antibiofilm, and Antioxidant Activity of 15 Different Plant-Based Natural Compounds in Comparison with Ciprofloxacin and Gentamicin. Antibiotics 2022, 11, 1099. [Google Scholar] [CrossRef]
- Ellermann, M.; Pacheco, A.R.; Jimenez, A.G.; Russell, R.M.; Cuesta, S.; Kumar, A.; Zhu, W.; Vale, G.; Martin, S.A.; Raj, P.; et al. Endocannabinoids Inhibit the Induction of Virulence in Enteric Pathogens. Cell 2020, 183, 650–665.e15. [Google Scholar] [CrossRef]
- Žitek, T.; Leitgeb, M.; Golle, A.; Dariš, B.; Knez, Ž.; Knez Hrnčič, M. The Influence of Hemp Extract in Combination with Ginger on the Metabolic Activity of Metastatic Cells and Microorganisms. Molecules 2020, 25, 4992. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Sikora, V.; Dincheva, I.; Kačániová, M.; Astatkie, T.; Semerdjieva, I.B.; Latkovic, D. Industrial, CBD, and Wild Hemp: How Different Are Their Essential Oil Profile and Antimicrobial Activity? Molecules 2020, 25, 4631. [Google Scholar] [CrossRef] [PubMed]
- Corni, G.; Brighenti, V.; Pellati, F.; Morlock, G.E. Effect-Directed Analysis of Bioactive Compounds in Cannabis sativa L. by High-Performance Thin-Layer Chromatography. J. Chromatogr. A 2020, 1629, 461511. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.S.; Gupta, M.K. Isolation of Nanocellulose from Hemp Fibers by Chemo-mechanical Method and Its Characterization. Polym. Compos. 2020, 41, 5257–5268. [Google Scholar] [CrossRef]
- Galletta, M.; Reekie, T.A.; Nagalingam, G.; Bottomley, A.L.; Harry, E.J.; Kassiou, M.; Triccas, J.A. Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 523. [Google Scholar] [CrossRef] [PubMed]
- Nalli, Y.; Arora, P.; Riyaz-Ul-Hassan, S.; Ali, A. Chemical Investigation of Cannabis sativa Leading to the Discovery of a Prenylspirodinone with Anti-Microbial Potential. Tetrahedron Lett. 2018, 59, 2470–2472. [Google Scholar] [CrossRef]
- Jin, S.; Lee, M.-Y. The Ameliorative Effect of Hemp Seed Hexane Extracts on the Propionibacterium Acnes-Induced Inflammation and Lipogenesis in Sebocytes. PLoS ONE 2018, 13, e0202933. [Google Scholar] [CrossRef]
- Khatak, S.; Satyender; Malik, D.K. Antimicrobial Activity of Withania Somnifera, Gymnema Sylvestre and Cannabis sativa against Pathogenic Bacteria. Int. J. Chem. Sci. 2014, 12, 286–292. [Google Scholar]
- Joseph, T.; Kar Mahapatra, D.; Esmaeili, A.; Piszczyk, Ł.; Hasanin, M.; Kattali, M.; Haponiuk, J.; Thomas, S. Nanoparticles: Taking a Unique Position in Medicine. Nanomaterials 2023, 13, 574. [Google Scholar] [CrossRef]
- Nasrullah, M.; Gul, F.Z.; Hanif, S.; Mannan, A.; Naz, S.; Ali, J.S.; Zia, M. Green and Chemical Syntheses of CdO NPs: A Comparative Study for Yield Attributes, Biological Characteristics, and Toxicity Concerns. ACS Omega 2020, 5, 5739–5747. [Google Scholar] [CrossRef]
- Korkmaz, N.; Kısa, D.; Ceylan, Y.; Güçlü, E.; Şen, F.; Karadağ, A. Synthesis of CeO2 Nanoparticles from Hemp Leaf Extract: Evaluation of Antibacterial, Anticancer and Enzymatic Activities. Inorg. Chem. Commun. 2024, 159, 111797. [Google Scholar] [CrossRef]
- Yaseen, B.; Gangwar, C.; Nayak, R.; Kumar, I.; Sarkar, J.; Baker, A.; Prasad, S.; Naik, R.M. Cannabis sativa Mediated Palladium Nanoparticles as an Effective Nanodrug against Multi-Drug Resistant Bacteria and A549 Lung Cancer Cells. Inorg. Chem. Commun. 2023, 157, 111254. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Zaka, M.; Hashmi, S.S.; Khan, Z. Biogenic Synthesis of Au, Ag and Au–Ag Alloy Nanoparticles Using Cannabis sativa Leaf Extract. IET Nanobiotechnol. 2018, 12, 277–284. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, S.P.; Sharma, U.R. Phytofabrication, Characterisation and Antimicrobial Studies of Gold Nanoparticles Using Cannabis sativa (Bhang), Leaf Extract. Int. J. Nanoparticles 2017, 9, 143. [Google Scholar] [CrossRef]
- Michailidu, J.; Miškovská, A.; Jarošová, I.; Čejková, A.; Mat’átková, O. Bimetallic Nanoparticle Production Using Cannabis sativa and Vitis Vinifera Waste Extracts. RSC Adv. 2024, 14, 5309–5318. [Google Scholar] [CrossRef]
- Chauhan, A.; Verma, R.; Kumari, S.; Sharma, A.; Shandilya, P.; Li, X.; Batoo, K.M.; Imran, A.; Kulshrestha, S.; Kumar, R. Photocatalytic Dye Degradation and Antimicrobial Activities of Pure and Ag-Doped ZnO Using Cannabis sativa Leaf Extract. Sci. Rep. 2020, 10, 7881. [Google Scholar] [CrossRef]
- Thakur, S.; Shandilya, M.; Guleria, G. Appraisement of Antimicrobial Zinc Oxide Nanoparticles through Cannabis Jatropha Curcasa Alovera and Tinosporacordifolia Leaves by Green Synthesis Process. J. Environ. Chem. Eng. 2021, 9, 104882. [Google Scholar] [CrossRef]
- Hasim, S.; Coleman, J.J. Targeting the Fungal Cell Wall: Current Therapies and Implications for Development of Alternative Antifungal Agents. Future Med. Chem. 2019, 11, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Galal Osman, A.; Elokely, K.M.; Yadav, V.K.; Carvalho, P.; Radwan, M.; Slade, D.; Gul, W.; Khan, S.; Dale, O.R.; Husni, A.S.; et al. Bioactive Products from Singlet Oxygen Photooxygenation of Cannabinoids. Eur. J. Med. Chem. 2018, 143, 983–996. [Google Scholar] [CrossRef]
- Motiejauskaitė, D.; Ullah, S.; Kundrotaitė, A.; Žvirdauskienė, R.; Bakšinskaitė, A.; Barčauskaitė, K. Isolation of Biologically Active Compounds from Cannabis sativa L. Inflorescences by Using Different Extraction Solvents and Evaluation of Antimicrobial Activity. Antioxidants 2023, 12, 998. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Ożarowski, M.; Seremak-Mrozikiewicz, A.; Wolski, H.; Adamczak, A. Plant Preparations and Compounds with Activities against Biofilms Formed by Candida Spp. J. Fungi 2021, 7, 360. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic Analyses, In Vitro Biological Activities, and Cytotoxicity of Cannabis sativa L. Essential Oil: A Multidisciplinary Study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef]
- Lu, L.; Su, S.; Yang, H.; Jiang, S. Antivirals with Common Targets against Highly Pathogenic Viruses. Cell 2021, 184, 1604–1620. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Muchiri, R.N. Affinity Selection-mass Spectrometry in the Discovery of Anti-SARS-CoV-2 Compounds. Mass Spectrom. Rev. 2024, 43, 39–46. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Simchuk, D. Antiviral Activities of Hemp Cannabinoids. Clin. Sci. 2023, 137, 633–643. [Google Scholar] [CrossRef] [PubMed]
- de Lima Marques, G.V.; de Almeida Marques, D.P.; Marques, G.V.; Clarindo, F.A.; Avendaño-Villarreal, J.A.; Guerra, F.S.; Fernandes, P.D.; dos Santos, E.N.; Gusevskaya, E.V.; Kohlhoff, M.; et al. Synthesis of Cannabidiol-Based Compounds as ACE2 Inhibitors with Potential Application in the Treatment of COVID-19. Eur. J. Med. Chem. 2023, 260, 115760. [Google Scholar] [CrossRef]
- Zargari, F.; Mohammadi, M.; Nowroozi, A.; Morowvat, M.H.; Nakhaei, E.; Rezagholi, F. The Inhibitory Effects of the Herbals Secondary Metabolites (7α-Acetoxyroyleanone, Curzerene, Incensole, Harmaline, and Cannabidiol) on COVID-19: A Molecular Docking Study. Recent Pat. Biotechnol. 2024, 18, 316–331. [Google Scholar] [CrossRef]
- El Ouafi, Z.; Rhalem, W.; Habib, N.; Idrissi Azami, A.; Sehli, S.; Al Idrissi, N.; Bakkali, F.; Abderrazak, R.; Merzouki, M.; Allali, I.; et al. Molecular Modeling Targeting the ACE2 Receptor with Cannabis sativa ’s Active Ingredients for Antiviral Drug Discovery against SARS-CoV-2 Infections. Bioinform. Biol. Insights 2022, 16, 117793222211453. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Saneei, Z.; Routy, J.-P.; Margolese, S.; Mandarino, E.; Singer, J.; Lebouché, B.; Cox, J.; Szabo, J.; Brouillette, M.-J.; et al. Oral Cannabinoids in People Living with HIV on Effective Antiretroviral Therapy: CTN PT028—Study Protocol for a Pilot Randomised Trial to Assess Safety, Tolerability and Effect on Immune Activation. BMJ Open 2019, 9, e024793. [Google Scholar] [CrossRef]
- Lowe, H.I.C.; Toyang, N.; McLaughlin, W. Potential of Cannabidiol for the Treatment of Viral Hepatitis. Pharmacogn. Res. 2017, 9, 116. [Google Scholar]
- Ogungbe, I.V.; Erwin, W.R.; Setzer, W.N. Antileishmanial Phytochemical Phenolics: Molecular Docking to Potential Protein Targets. J. Mol. Graph. Model. 2014, 48, 105–117. [Google Scholar] [CrossRef]
- Roulette, C.J.; Kazanji, M.; Breurec, S.; Hagen, E.H. High Prevalence of Cannabis Use among Aka Foragers of the Congo Basin and Its Possible Relationship to Helminthiasis. Am. J. Hum. Biol. 2016, 28, 5–15. [Google Scholar] [CrossRef]
- Al Jalali, V.; Zeitlinger, M. Systemic and Target-Site Pharmacokinetics of Antiparasitic Agents. Clin. Pharmacokinet. 2020, 59, 827–847. [Google Scholar] [CrossRef]
- World Health Organization. WHO Releases Report on State of Development of Antibacterials; World Health Organization: Geneva, Switzerland, 2024. [Google Scholar]
- Khameneh, B.; Diab, R.; Ghazvini, K.; Fazly Bazzaz, B.S. Breakthroughs in Bacterial Resistance Mechanisms and the Potential Ways to Combat Them. Microb. Pathog. 2016, 95, 32–42. [Google Scholar] [CrossRef]
- Qiu, H.; Si, Z.; Luo, Y.; Feng, P.; Wu, X.; Hou, W.; Zhu, Y.; Chan-Park, M.B.; Xu, L.; Huang, D. The Mechanisms and the Applications of Antibacterial Polymers in Surface Modification on Medical Devices. Front. Bioeng. Biotechnol. 2020, 8, 910. [Google Scholar] [CrossRef]
- Gong, X.; Liu, L.; Li, X.; Xiong, J.; Xu, J.; Mao, D.; Liu, L. Neuroprotection of Cannabidiol in Epileptic Rats: Gut Microbiome and Metabolome Sequencing. Front. Nutr. 2022, 9, 1028459. [Google Scholar] [CrossRef] [PubMed]
- Kosgodage, U.S.; Matewele, P.; Awamaria, B.; Kraev, I.; Warde, P.; Mastroianni, G.; Nunn, A.V.; Guy, G.W.; Bell, J.D.; Inal, J.M.; et al. Cannabidiol Is a Novel Modulator of Bacterial Membrane Vesicles. Front. Cell. Infect. Microbiol. 2019, 9, 324. [Google Scholar] [CrossRef]
- MacNair, C.R.; Stokes, J.M.; Carfrae, L.A.; Fiebig-Comyn, A.A.; Coombes, B.K.; Mulvey, M.R.; Brown, E.D. Overcoming Mcr-1 Mediated Colistin Resistance with Colistin in Combination with Other Antibiotics. Nat. Commun. 2018, 9, 458. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.; Lopes, C.M.; Amaral, M.H. Treatment Advances for Acne Vulgaris: The Scientific Role of Cannabinoids. Cosmetics 2024, 11, 22. [Google Scholar] [CrossRef]
- Ribeiro, A.; Serrano, R.; da Silva, I.B.M.; Pinto, J.F.; Silva, O. Development of an Adhesive Tablet for the Delivery of Diospyros Villosa Root for the Treatment of Oral Cavity Diseases. J. Drug Deliv. Sci. Technol. 2023, 90, 105148. [Google Scholar] [CrossRef]
- Ribeiro, A.; Serrano, R.; Silva, I.B.M.d.; Gomes, E.T.; Pinto, J.F.; Silva, O. Diospyros Villosa Root Monographic Quality Studies. Plants 2022, 11, 3506. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.; Ribeiro, A.; Diniz, S.; Cabral-Marques, H.; Sousa-Martins, D. Incidence of Malignant Eyelid Tumors: A 6-Year Period Review (2015–2021). Pan-Am. J. Ophthalmol. 2024, 1, 62. [Google Scholar] [CrossRef]
- Oliveira, D.; Ribeiro, A.; Bacalhau, C.; Almada, S.; Diniz, S.; Saramago, A.R.; Cabral-Marques, H.; Sousa-Martins, D. Are We Capable of Contributing to the Reduction of Eyelid Basal Cell Carcinoma? Building a Healthcare Team. J. Ophthalmol. Res. Rev. Rep. 2023, 146, 1–8. [Google Scholar]
- Zhao, T.; Li, X.; Gong, Y.; Guo, Y.; Quan, F.; Shi, Q. Study on Polysaccharide Polyelectrolyte Complex and Fabrication of Alginate/Chitosan Derivative Composite Fibers. Int. J. Biol. Macromol. 2021, 184, 181–187. [Google Scholar] [CrossRef]
- Yang, D.; Gong, L.; Li, Q.; Fan, B.; Ma, C.; He, Y.-C. Preparation of a Biobased Polyelectrolyte Complex from Chitosan and Sodium Carboxymethyl Cellulose and Its Antibacterial Characteristics. Int. J. Biol. Macromol. 2023, 227, 524–534. [Google Scholar] [CrossRef]
- Persia, D.; Mangiavacchi, F.; Marcotullio, M.C.; Rosati, O. Cannabinoids as Multifaceted Compounds. Phytochemistry 2023, 212, 113718. [Google Scholar] [CrossRef]
- de Brito Siqueira, A.L.G.; Cremasco, P.V.V.; Bahú, J.O.; Pioli da Silva, A.; Melo de Andrade, L.R.; González, P.G.A.; Crivellin, S.; Cárdenas Concha, V.O.; Krambeck, K.; Lodi, L.; et al. Phytocannabinoids: Pharmacological Effects, Biomedical Applications, and Worldwide Prospection. J. Tradit. Complement. Med. 2023, 13, 575–587. [Google Scholar] [CrossRef]
- Casalini, G.; Giacomelli, A.; Antinori, S. The WHO Fungal Priority Pathogens List: A Crucial Reappraisal to Review the Prioritisation. Lancet Microbe 2024, 5, 717–724. [Google Scholar] [CrossRef]
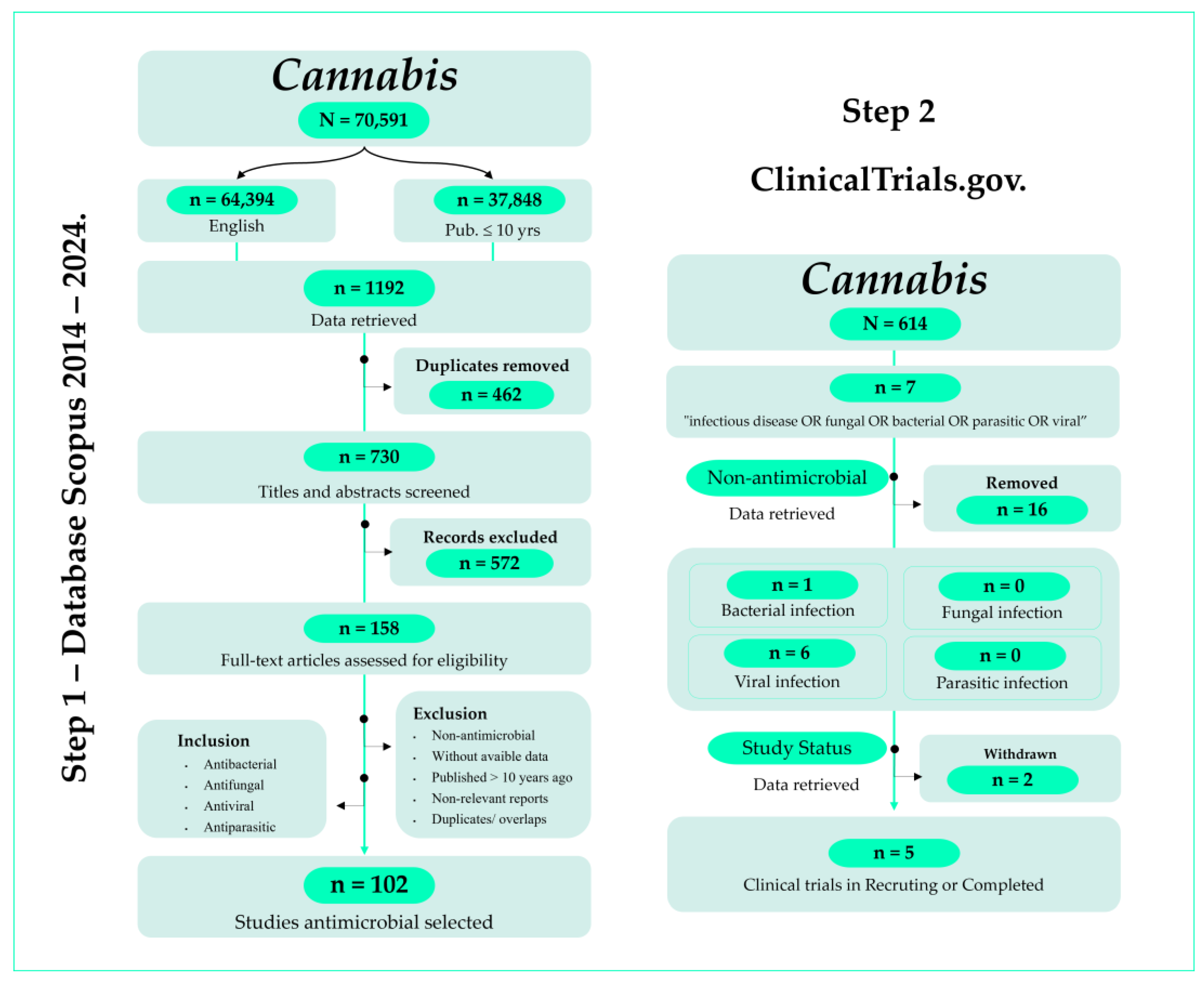
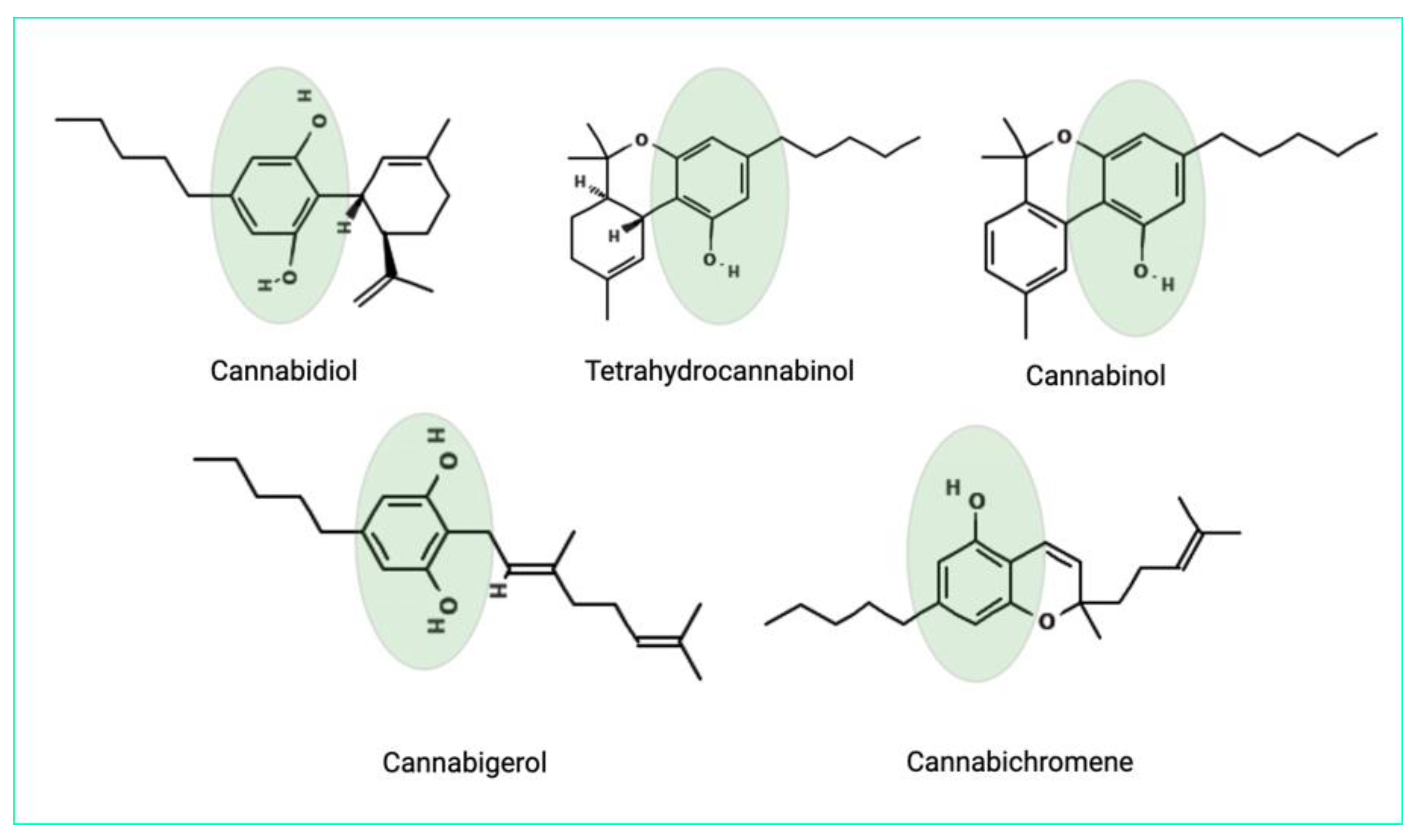
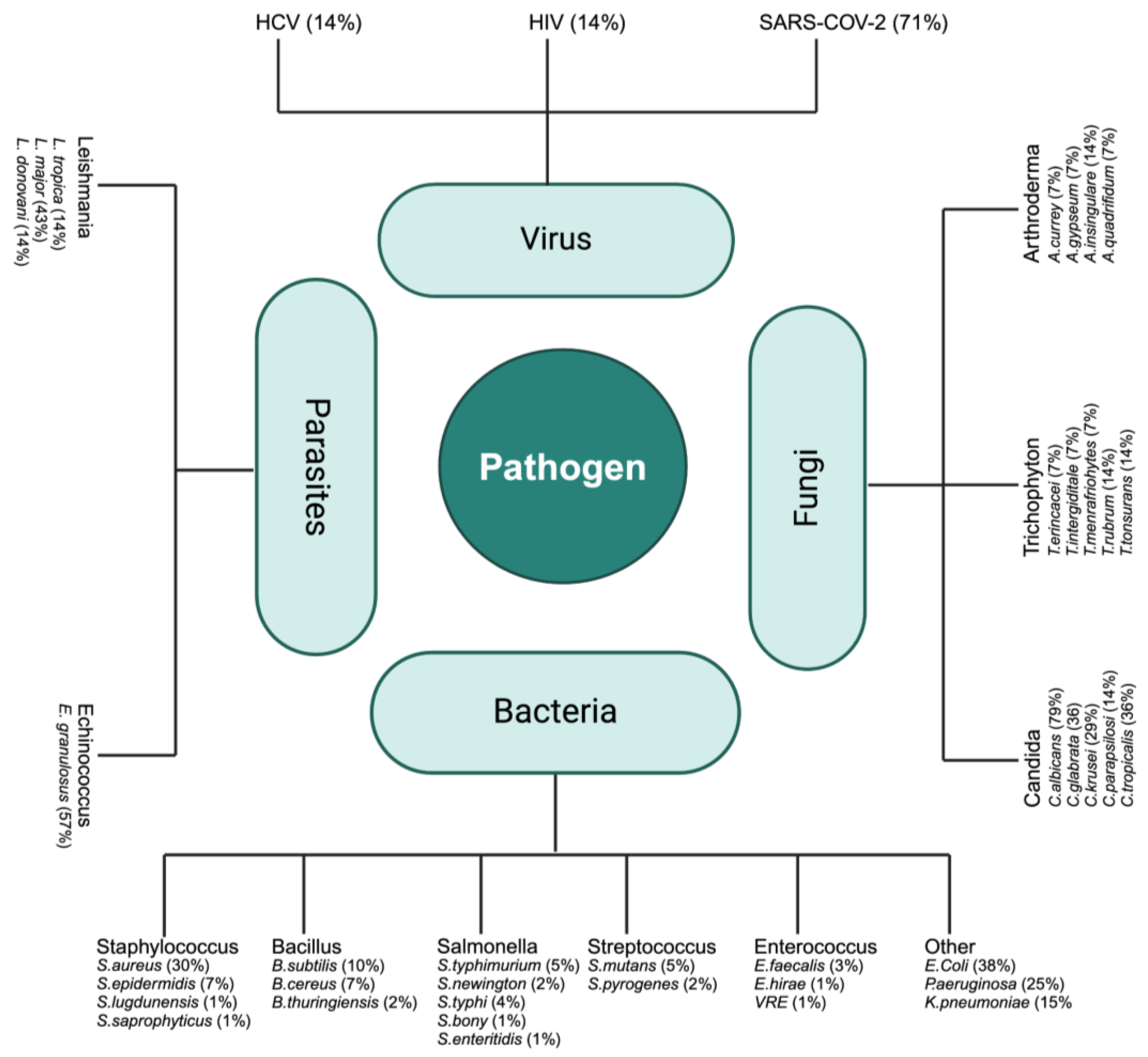
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).