
Gene-Based Therapy: A New Approach to Feline Induced Sterilization?


Abstract
1. Introduction
2. Hormonal vs. Non-Hormonal Pharmacological Methods for Feline Induced Sterilization
3. Gene-Based Methods for Feline Induced Sterilization
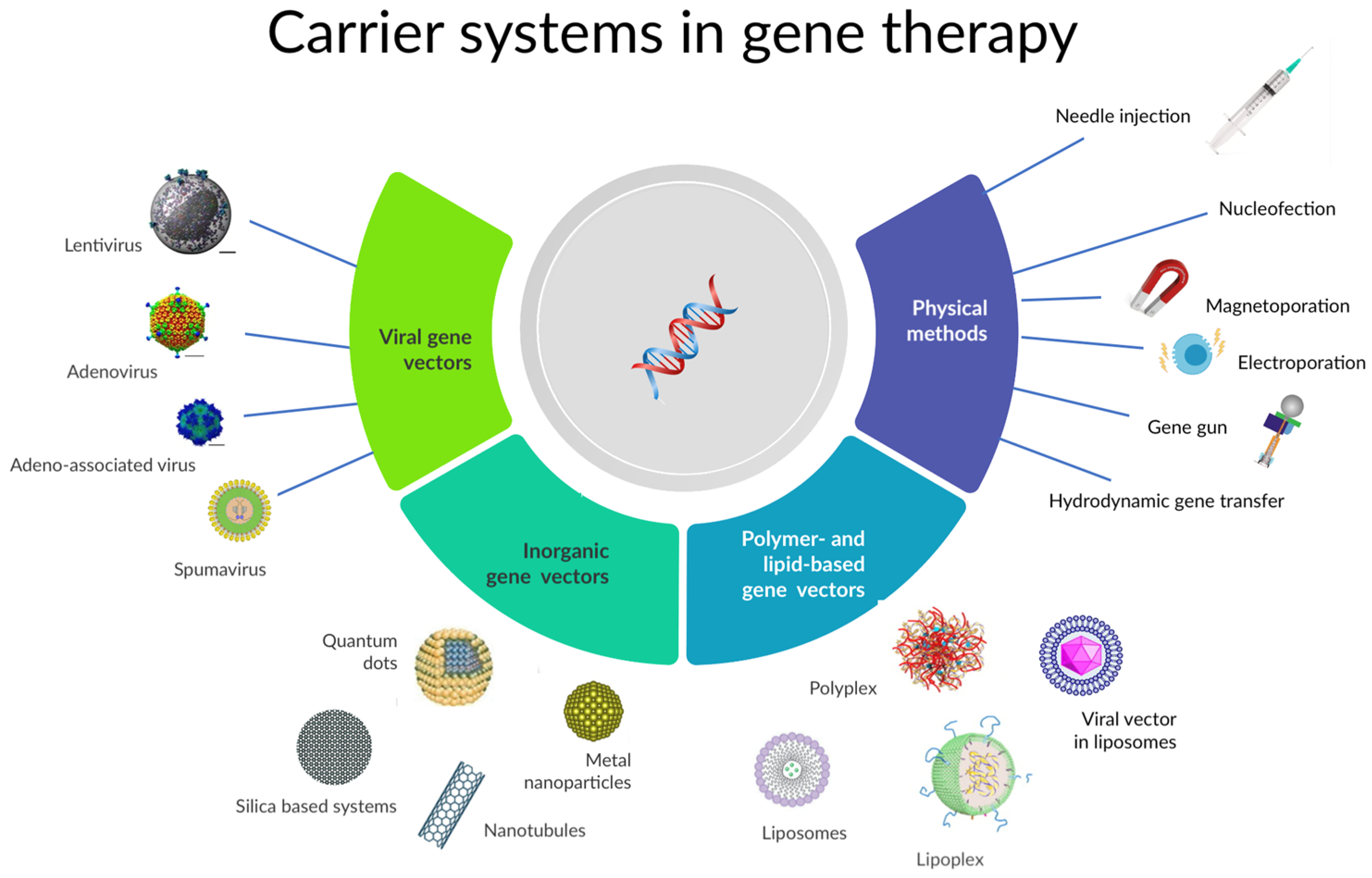
3.1. Mechanistic of Gene-Based Therapy in Brief
3.2. Gene-Based Sterilization
3.3. Risks and Limitations in Gene Therapy
4. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vansandt, L.M.; Meinsohn, M.C.; Godin, P.; Nagykery, N.; Sicher, N.; Kano, M.; Kashiwagi, A.; Chauvin, M.; Saatcioglu, H.D.; Barnes, J.L.; et al. Durable contraception in the female domestic cat using viral-vectored delivery of a feline anti-Müllerian hormone transgene. Nat. Commun. 2023, 14, 3140. [Google Scholar] [CrossRef]
- Hamilton, F. Implementing Nonlethal Solutions for Free-Roaming Cat Management in a County in the Southeastern United States. Front. Vet. Sci. 2019, 6, 259. [Google Scholar] [CrossRef]
- Kennedy, B.P.A.; Cumming, B.; Brown, W.Y. Global Strategies for Population Management of Domestic Cats. Animals 2020, 10, 663. [Google Scholar] [CrossRef]
- Luzardo, O.P.; Zaldívar-Laguía, J.E.; Zumbado, M.; Travieso-Aja, M.D.M. The Role of Veterinarians in Managing Community Cats: A Contextualized, Comprehensive Approach for Biodiversity, Public Health, and Animal Welfare. Animals 2023, 13, 1586. [Google Scholar] [CrossRef]
- Hurley, K.F.; Levy, J.K. Rethinking the Animal Shelter’s Role in Free-Roaming Cat Management. Front. Vet. Sci. 2022, 9, 847081. [Google Scholar] [CrossRef]
- Crawford, H.M.; Calver, M.C.; Fleming, P.A. A Case of Letting the Cat out of The Bag-Why Trap-Neuter-Return Is Not an Ethical Solution for Stray Cat. Animals 2019, 9, 171. [Google Scholar] [CrossRef]
- Calver, M.C.; Crawford, H.M.; Scarff, F.R.; Bradley, J.S.; Dormon, P.; Boston, S.; Fleming, P.A. Intensive Adoption as a Management Strategy for Unowned, Urban Cats: A Case Study of 25 Years of Trap–Assess–Resolve (TAR) in Auckland, New Zealand. Animals 2022, 12, 2301. [Google Scholar] [CrossRef]
- Munson, L. Contraception in felids. Theriogenology 2006, 66, 126–134. [Google Scholar] [CrossRef]
- Massei, G.; Cowan, D. Fertility control to mitigate human–wildlife conflicts: A review. Wildl. Res. 2014, 41, 1–21. [Google Scholar] [CrossRef]
- Kutzler, M.A. Alternative methods for feline fertility control: Use of melatonin to suppress reproduction. J. Feline Med. Surg. 2015, 17, 753–757. [Google Scholar] [CrossRef]
- Massei, G. Fertility Control for Wildlife: A European Perspective. Animals 2023, 13, 428. [Google Scholar] [CrossRef]
- Gunther, I.; Hawlena, H.; Azriel, L.; Gibor, D.; Berke, O.; Klement, E. Reduction of free-roaming cat population requires high-intensity neutering in spatial contiguity to mitigate compensatory effects. Proc. Natl. Acad. Sci. USA 2022, 119, e2119000119. [Google Scholar] [CrossRef]
- Massei, G.; Miller, L.A. Nonsurgical fertility control for managing free-roaming dog populations: A review of products and criteria for field applications. Theriogenology 2013, 80, 829–838. [Google Scholar] [CrossRef]
- Kutzler, M.; Wood, A. Non-surgical methods of contraception and sterilization. Theriogenology 2006, 66, 514–525. [Google Scholar] [CrossRef]
- Rhodes, L. New approaches to non-surgical sterilization for dogs and cats: Opportunities and challenges. Reprod. Domest. Anim. 2017, 52 (Suppl. 2), 327–331. [Google Scholar] [CrossRef]
- Goericke-Pesch, S.; Wehrend, A.; Georgiev, P. Suppression of fertility in adult cats. Reprod. Domest. Anim. 2014, 49 (Suppl. 2), 33–40. [Google Scholar] [CrossRef]
- Goericke-Pesch, S. Reproduction control in cats: New developments in non-surgical methods. J. Feline Med. Surg. 2010, 12, 539–546. [Google Scholar] [CrossRef]
- Romagnoli, S.; Ferre-Dolcet, L. Reversible Control of Reproduction In Queens: Mastering the use of reproductive drugs to manipulate cyclicity. J. Feline Med. Surg. 2022, 24, 853–870. [Google Scholar] [CrossRef]
- Romagnoli, S. Progestins to control feline reproduction: Historical abuse of high doses and potentially safe use of low doses. J. Feline Med. Surg. 2015, 17, 743–752. [Google Scholar] [CrossRef]
- Johnston, S.D.; Root Kustritz, M.V.; Olson, P.S. Prevention and termination of feline pregnancy. In Canine and Feline Theriogenology, 1st ed.; Johnston, S.D., Root Kustritz, M.V., Olson, P.S., Eds.; Saunders: Philadelphia, PA, USA, 2001; pp. 447–452. [Google Scholar]
- Graham, L.H.; Swanson, W.F.; Wildt, D.E.; Brown, J.L. Influence of oral melatonin on natural and gonadotropin-induced ovarian function in the domestic cat. Theriogenology 2004, 61, 1061–1076. [Google Scholar] [CrossRef]
- Gimenez, F.; Stornelli, M.C.; Tittarelli, C.M.; Savignone, C.A.; Dorna, I.V.; de la Sota, R.L.; Stornelli, M.A. Suppression of estrus in cats with melatonin implants. Theriogenology 2009, 72, 493–499. [Google Scholar] [CrossRef]
- Schäfer-Somi, S. Effect of melatonin on the reproductive cycle in female cats: A review of clinical experiences and previous studies. J. Feline Med. Surg. 2017, 19, 5–12. [Google Scholar] [CrossRef]
- Furthner, E.; Roos, J.; Niewiadomska, Z.; Maenhoudt, C.; Fontbonne, A. Contraceptive implants used by cat breeders in France: A study of 140 purebred cats. J. Feline Med. Surg. 2020, 22, 984–992. [Google Scholar] [CrossRef]
- Griffin, B.; Heath, A.M.; Young, D.W.; Wright, J.; Rolsma, M.D.; Baker, H.; Ritchey, S. Effects of melatonin implants on ovarian function in the domestic cat. In Proceedings of the 19th American College of Veterinary Internal Medicine, Denver, CO, USA; 2001; p. 843. [Google Scholar]
- Blottner, S.; Jewgenow, K. Moderate seasonality in testis function of domestic cat. Reprod. Domest. Anim. 2007, 42, 536–540. [Google Scholar] [CrossRef]
- Nuñez-Favre, R.; Bonaura, M.C.; Tittarelli, C.M.; Mansilla-Hermann, D.; de la Sota, R.L.; Stornelli, M.A. Effect of natural photoperiod on epididymal sperm quality and testosterone serum concentration in domestic cat (Felis silvestris catus). Reprod. Domest. Anim. 2012, 47 (Suppl. 6), 232–234. [Google Scholar] [CrossRef]
- Ferré-Dolcet, L.; Romagnoli, S. Reversible control of reproduction in tom cats: Medical options for manipulating libido and fertility. J. Feline Med. Surg. 2023, 25, 1098612X231171406. [Google Scholar] [CrossRef]
- Nuñez Favre, R.; Bonaura, M.C.; Tittarelli, C.M.; Stornelli, M.C.; de la Sota, R.L.; Stornelli, M.A. Effect of refractoriness to long photoperiod on sperm production and quality in tomcats. Reprod. Domest. Anim. 2012, 47 (Suppl. 6), 235–237. [Google Scholar] [CrossRef]
- Favre, R.N.; Bonaura, M.C.; Praderio, R.; Stornelli, M.C.; de la Sota, R.L.; Stornelli, M.A. Effect of melatonin implants on spermatogenesis in the domestic cat (Felis silvestris catus). Theriogenology 2014, 82, 851–856. [Google Scholar] [CrossRef]
- Padula, A.M. GnRH analogues—Agonists and antagonists. Anim. Reprod. Sci. 2005, 88, 115–126. [Google Scholar] [CrossRef]
- Fontaine, C. Long-term contraception in a small implant: A review of Suprelorin (deslorelin) studies in cats. J. Feline Med. Surg. 2015, 17, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Borges, P.; Fontaine, E.; Maenhoudt, C.; Payan-Carreira, R.; Santos, N.; Leblond, E.; Fontaine, C.; Fontbonne, A. Fertility in Adult Bitches Previously Treated with a 4.7 mg Subcutaneous Deslorelin Implant. Reprod. Domest. Anim. 2015, 50, 965–971. [Google Scholar] [CrossRef]
- Romagnoli, S.; Baldan, A.; Ferro, S.; Righetti, C.; Scenna, L.; Gabai, G.; Badon, T.; Fontaine, C.; Mollo, A.; Stelletta, C.; et al. Length of efficacy and effect of implant location in adult tom cats treated with a 9.4 mg deslorelin subcutaneous implant. J. Feline Med. Surg. 2019, 21, 507–519. [Google Scholar] [CrossRef]
- Levy, J.K. Contraceptive vaccines for the humane control of community cat populations. Am. J. Reprod. Immunol. 2011, 66, 63–70. [Google Scholar] [CrossRef]
- Munson, L.; Harrenstien, L.A.; Acton, A.E.; Graham, P.A.; Chassy, L.M.; Kirkpatrick, J.F. Immunologic responses and adverse reactions to Freund’s-adjuvanted porcine zona pellucida immuno-contraceptives in domestic cats. Vaccine 2005, 23, 5646–5654. [Google Scholar] [CrossRef]
- Lee, Y.J.; Jo, E.J.; Lee, H.W.; Hwang, B.R.; Kim, Y.H.; Park, B.J.; Cho, Y.J.; Lee, Y.A.; Choi, I.S.; Han, J.S. Evaluation of infertility efficacy of the E. coli expressed STF2-GnRH vaccine in male cats. J. Vet. Sci. 2019, 20, e30. [Google Scholar] [CrossRef]
- Ochoa, J.S.; Favre, R.N.; García, M.F.; Stornelli, M.C.; Sangache, W.C.; Rearte, R.; de la Sota, L.; Stornelli, M.A. Immunocontraception of male domestic cats using GnRH vaccine Improvac. Theriogenology 2023, 198, 211–216. [Google Scholar] [CrossRef]
- Samoylov, A.; Napier, I.; Morrison, N.; Cochran, A.; Schemera, B.; Wright, J.; Cattley, R.; Samoylova, T. DNA Vaccine Targeting Gonadotropin-Releasing Hormone Receptor and Its Application in Animal Contraception. Mol. Biotechnol. 2019, 61, 73–83. [Google Scholar] [CrossRef]
- Dunbar, C.E.; High, K.A.; Joung, J.K.; Kohn, D.B.; Ozawa, K.; Sadelain, M. Gene therapy comes of age. Science 2018, 359, eaan4672. [Google Scholar] [CrossRef]
- Alnasser, S.M. Review on mechanistic strategy of gene therapy in the treatment of disease. Gene 2021, 769, 145246. [Google Scholar] [CrossRef]
- Li, J.; Olvera, A.I.; Akbari, O.S.; Moradian, A.; Sweredoski, M.J.; Hess, S.; Hay, B.A. Vectored antibody gene delivery mediates long-term contraception. Curr. Biol. 2015, 25, R820–R822. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kamimura, K.; Suda, T.; Zhang, G.; Liu, D. Advances in Gene Delivery Systems. Pharm. Med. 2011, 25, 293–306. [Google Scholar] [CrossRef]
- Butt, M.H.; Zaman, M.; Ahmad, A.; Khan, R.; Mallhi, T.H.; Hasan, M.M.; Khan, Y.H.; Hafeez, S.; Massoud, E.E.S.; Rahman, M.H.; et al. Appraisal for the Potential of Viral and Nonviral Vectors in Gene Therapy: A Review. Genes 2022, 13, 1370. [Google Scholar] [CrossRef]
- Guo, X.; Huang, L. Recent advances in nonviral vectors for gene delivery. Acc. Chem. Res. 2012, 45, 971–979. [Google Scholar] [CrossRef]
- Wang, C.; Pan, C.; Yong, H.; Wang, F.; Bo, T.; Zhao, Y.; Ma, B.; He, W.; Li, M. Emerging non-viral vectors for gene delivery. J. Nanobiotechnol. 2023, 21, 272. [Google Scholar] [CrossRef]
- Huang, S.; Kamihira, M. Development of hybrid viral vectors for gene therapy. Biotechnol. Adv. 2013, 31, 208–223. [Google Scholar] [CrossRef]
- Mahato, M.; Jayandharan, G.R.; Vemula, P.K. Viral- and Non-viral-Based Hybrid Vectors for Gene Therapy. In Gene and Cell Therapy: Biology and Applications; Jayandharan, G.R., Ed.; Springer: Singapore, 2018; pp. 111–130. [Google Scholar]
- Zhang, J.; Jia, Y.; Li, X.; Hu, Y. Facile engineering of biocompatible materials with pH-modulated degradability. Adv. Mater. 2011, 23, 3035–3040. [Google Scholar] [CrossRef]
- Chen, H.; Liu, X.; Dou, Y.; He, B.; Liu, L.; Wei, Z.; Li, J.; Wang, C.; Mao, C.; Zhang, J.; et al. A pH-responsive cyclodextrin-based hybrid nanosystem as a nonviral vector for gene delivery. Biomaterials 2013, 34, 4159–4172. [Google Scholar] [CrossRef]
- Stribley, J.M.; Rehman, K.S.; Niu, H.; Christman, G.M. Gene therapy and reproductive medicine. Fertil. Steril. 2002, 77, 645–657. [Google Scholar] [CrossRef]
- Anguela, X.M.; High, K.A. Entering the Modern Era of Gene Therapy. Annu. Rev. Med. 2019, 70, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. RNA Viruses as Tools in Gene Therapy and Vaccine Development. Genes 2019, 10, 189. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.K.; Kim, S.W. Recent advances in the development of gene delivery systems. Biomater. Res. 2019, 23, 8. [Google Scholar] [CrossRef]
- Ledley, F.D. Nonviral gene therapy: The promise of genes as pharmaceutical products. Hum. Gene Ther. 1995, 6, 1129–1144. [Google Scholar] [CrossRef]
- Áyen, Á.; Jiménez Martínez, Y.; Marchal, J.A.; Boulaiz, H. Recent Progress in Gene Therapy for Ovarian Cancer. Int. J. Mol. Sci. 2018, 19, 1930. [Google Scholar] [CrossRef]
- Drakopoulou, E.; Anagnou, N.P.; Pappa, K.I. Gene Therapy for Malignant and Benign Gynaecological Disorders: A Systematic Review of an Emerging Success Story. Cancers 2022, 14, 3238. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Visser, J.A.; Themmen, A.P. Regulation of ovarian function: The role of anti-Müllerian hormone. Reproduction 2002, 124, 601–609. [Google Scholar] [CrossRef]
- Sriraman, V.; Niu, E.; Matias, J.R.; Donahoe, P.K.; Maclaughlin, D.T.; Hardy, M.P.; Lee, M.M. Müllerian Inhibiting Substance Inhibits Testosterone Synthesis in Adult Rats. J. Androl. 2001, 22, 750–758. [Google Scholar] [CrossRef]
- Pépin, D.; Sosulski, A.; Zhang, L.; Wang, D.; Vathipadiekal, V.; Hendren, K.; Coletti, C.M.; Yu, A.; Castro, C.M.; Birrer, M.J.; et al. AAV9 delivering a modified human Mullerian inhibiting substance as a gene therapy in patient-derived xenografts of ovarian cancer. Proc. Natl. Acad. Sci. USA 2015, 112, E4418–E4427. [Google Scholar] [CrossRef]
- Kano, M.; Sosulski, A.E.; Zhang, L.; Saatcioglu, H.D.; Wang, D.; Nagykery, N.; Sabatini, M.E.; Gao, G.; Donahoe, P.K.; Pépin, D. AMH/MIS as a contraceptive that protects the ovarian reserve during chemotherapy. Proc. Natl. Acad. Sci. USA 2017, 114, E1688–E1697. [Google Scholar] [CrossRef] [PubMed]
- Hay, B.A.; Li, J.; Guo, M. Vectored gene delivery for lifetime animal contraception: Overview and hurdles to implementation. Theriogenology 2018, 112, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.; Johnson, A.; Wilborn, R.; Braden, T.; Kutzler, M.; Roberts, S.; Van Kampen, K.; Trumble, J.; Baker, H. Adenoviral-vectored gonadotropin releasing hormone vaccine for estrus suppression in mares. Clin. Theriogenol. 2020, 12, 124–133. [Google Scholar]
- Wu, X.; Yang, Y.; Kling, C.; Seigler, L.; Gallardo-Romero, N.F.; Martin, B.E.; Smith, T.G.; Olson, V.A. Inactivated Rabies Virus-Vectored Immunocontraceptive Vaccine in a Thermo-Responsive Hydrogel Induces High and Persistent Antibodies against Rabies, but Insufficient Antibodies against Gonadotropin-Releasing Hormone for Contraception. Vaccines 2019, 7, 73. [Google Scholar] [CrossRef]
- Shahryari, A.; Burtscher, I.; Nazari, Z.; Lickert, H. Engineering Gene Therapy: Advances and Barriers. Adv. Ther. 2021, 4, 2100040. [Google Scholar] [CrossRef]
- Brown, A.M.; Blind, J.; Campbell, K.; Ghosh, S. Safeguards for Using Viral Vector Systems in Human Gene Therapy: A Resource for Biosafety Professionals Mitigating Risks in Health Care Settings. Appl. Biosaf. 2020, 25, 184–193. [Google Scholar] [CrossRef]
- Ghosh, S.; Brown, A.M.; Jenkins, C.; Campbell, K. Viral Vector Systems for Gene Therapy: A Comprehensive Literature Review of Progress and Biosafety Challenges. Appl. Biosaf. 2020, 25, 7–18. [Google Scholar] [CrossRef]
- Bakker, J.M.; Bleeker, W.K.; Parren, P.W. Therapeutic antibody gene transfer: An active approach to passive immunity. Mol. Ther. 2004, 10, 411–416. [Google Scholar] [CrossRef]
- Alton, E.W.F.W.; Boyd, A.C.; Davies, J.C.; Gill, D.R.; Griesenbach, U.; Harman, T.E.; Hyde, S.; McLachlan, G. Gene Therapy for Respiratory Diseases: Progress and a Changing Context. Hum. Gene Ther. 2020, 31, 911–916. [Google Scholar] [CrossRef]
- Iwasaki, A. Immune Regulation of Antibody Access to Neuronal Tissues. Trends Mol. Med. 2017, 23, 227–245. [Google Scholar] [CrossRef]
- Sabatini, M.T.; Chalmers, M. The Cost of Biotech Innovation: Exploring Research and Development Costs of Cell and Gene Therapies. Pharm. Med. 2023, 37, 365–375. [Google Scholar] [CrossRef]
- Editorial, N. The gene-therapy revolution risks stalling if we don’t talk about drug pricing. Nature 2023, 616, 629–630. [Google Scholar] [CrossRef]


{kind=link}
| Treatment | Disadvantages | Reported Side-Effects | Comments |
|---|---|---|---|
| Melatonin implants (18 mg) |
|
|
|
| Slow-release GnRH implants (4.7 mg or 9.4 mg) |
|
|
|
| Immunocontraception |
|
|
|
|
|
|
| Features | Retroviruses | Lentiviruses | Spumaviruses | Adenovirus | Adeno-Associated Adenoviruses |
|---|---|---|---|---|---|
| Viral genome | Single-stranded RNA | Single-stranded RNA | Single-stranded RNA | DNA | DNA |
| Cell division requirements in target cells | Dividing cells | G1 phase | No preferences | No | No |
| Gene loading limitation | 8 kb | 8 kb | 9.2 kb | 8–37 kb | 4–5 kb |
| Immune responses to the vector | Low | Low | Low | Extensiveat the inoculation local | Low |
| Genome integration | Yes | Yes | No preferences | No | Some integration ability |
| Main disadvantages | Random integration Low titers | Random integration Potential for pathogenic vector mutation | Random integration | Transient expression Requests repeated administration | Later onset of expression |
| Main advantages | Persistent gene transfer in dividing cells Long term expression | Broad host range High transduction efficiency Persistent gene transfer in transduced tissues | Highly effective in dividing cells No expression of viral proteins | Highly effective in transducing various tissues Large gene loading capacity | Elicits few inflammatory responses Sustained gene expression Non-pathogenic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Payan-Carreira, R. Gene-Based Therapy: A New Approach to Feline Induced Sterilization? Future Pharmacol. 2023, 3, 938-950. https://doi.org/10.3390/futurepharmacol3040057
Payan-Carreira R. Gene-Based Therapy: A New Approach to Feline Induced Sterilization? Future Pharmacology. 2023; 3(4):938-950. https://doi.org/10.3390/futurepharmacol3040057
Chicago/Turabian StylePayan-Carreira, Rita. 2023. "Gene-Based Therapy: A New Approach to Feline Induced Sterilization?" Future Pharmacology 3, no. 4: 938-950. https://doi.org/10.3390/futurepharmacol3040057
APA StylePayan-Carreira, R. (2023). Gene-Based Therapy: A New Approach to Feline Induced Sterilization? Future Pharmacology, 3(4), 938-950. https://doi.org/10.3390/futurepharmacol3040057