

Manifestation of Heat-Induced Valuable Dietary Nucleotide Salvage in Food Prepared from Aged Fish in Fast Protein and Metabolites Liquid Chromatography, ATP-Bioluminescence Assay, and NMR Spectra

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Methods
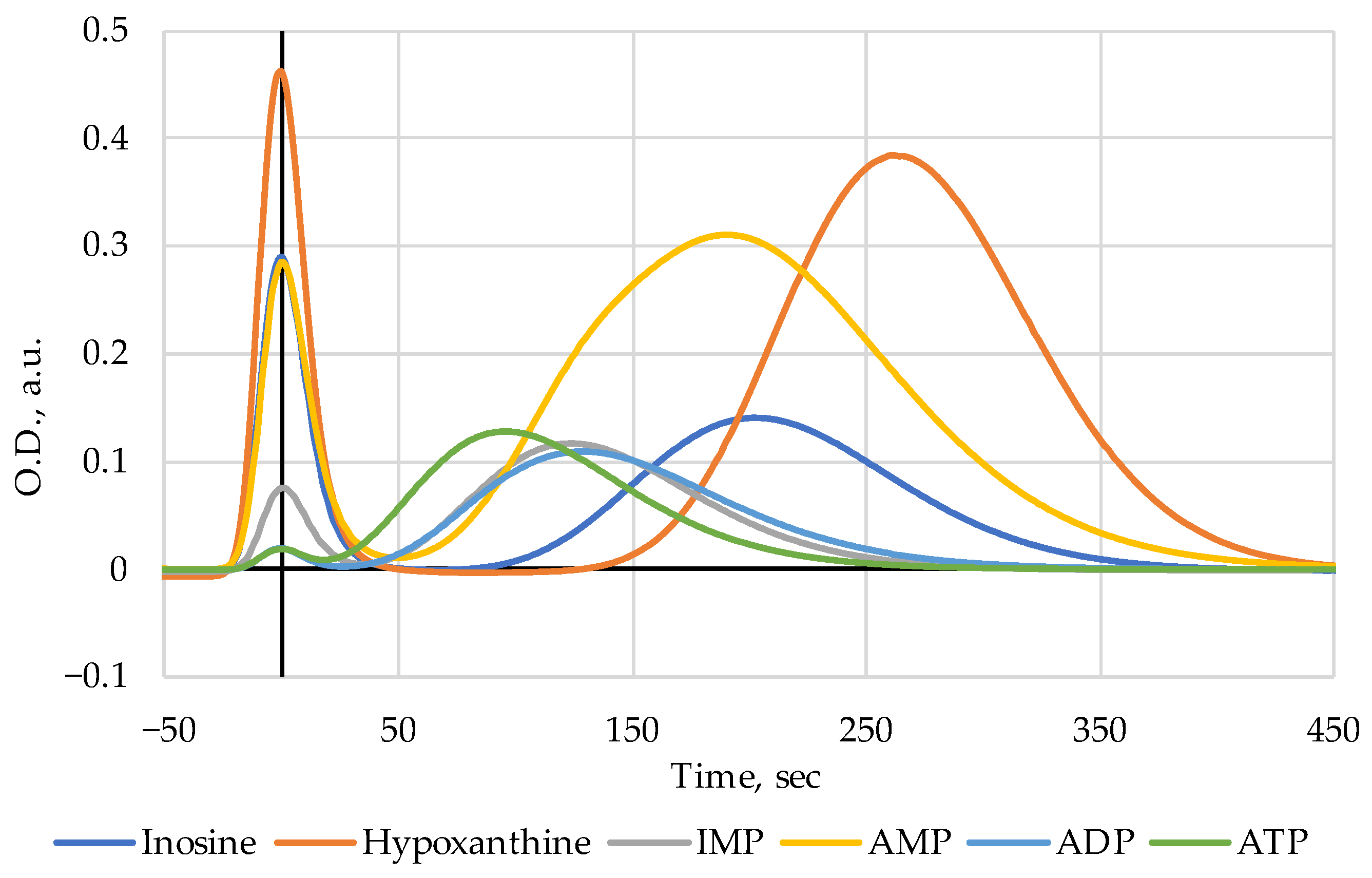
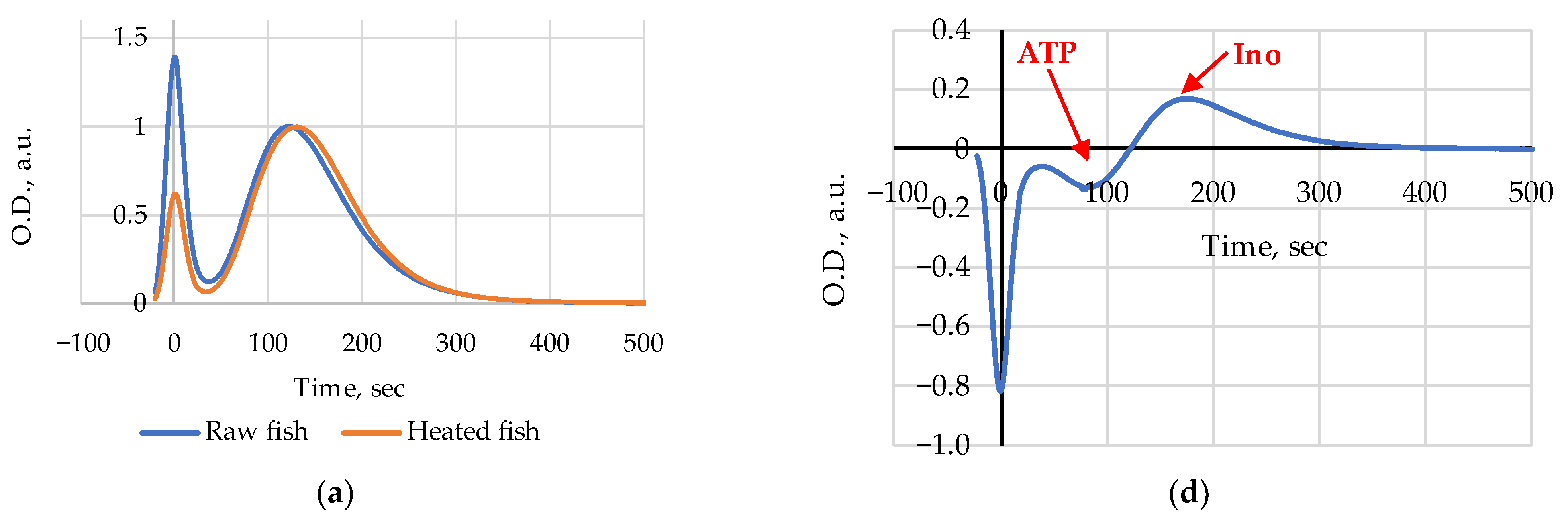
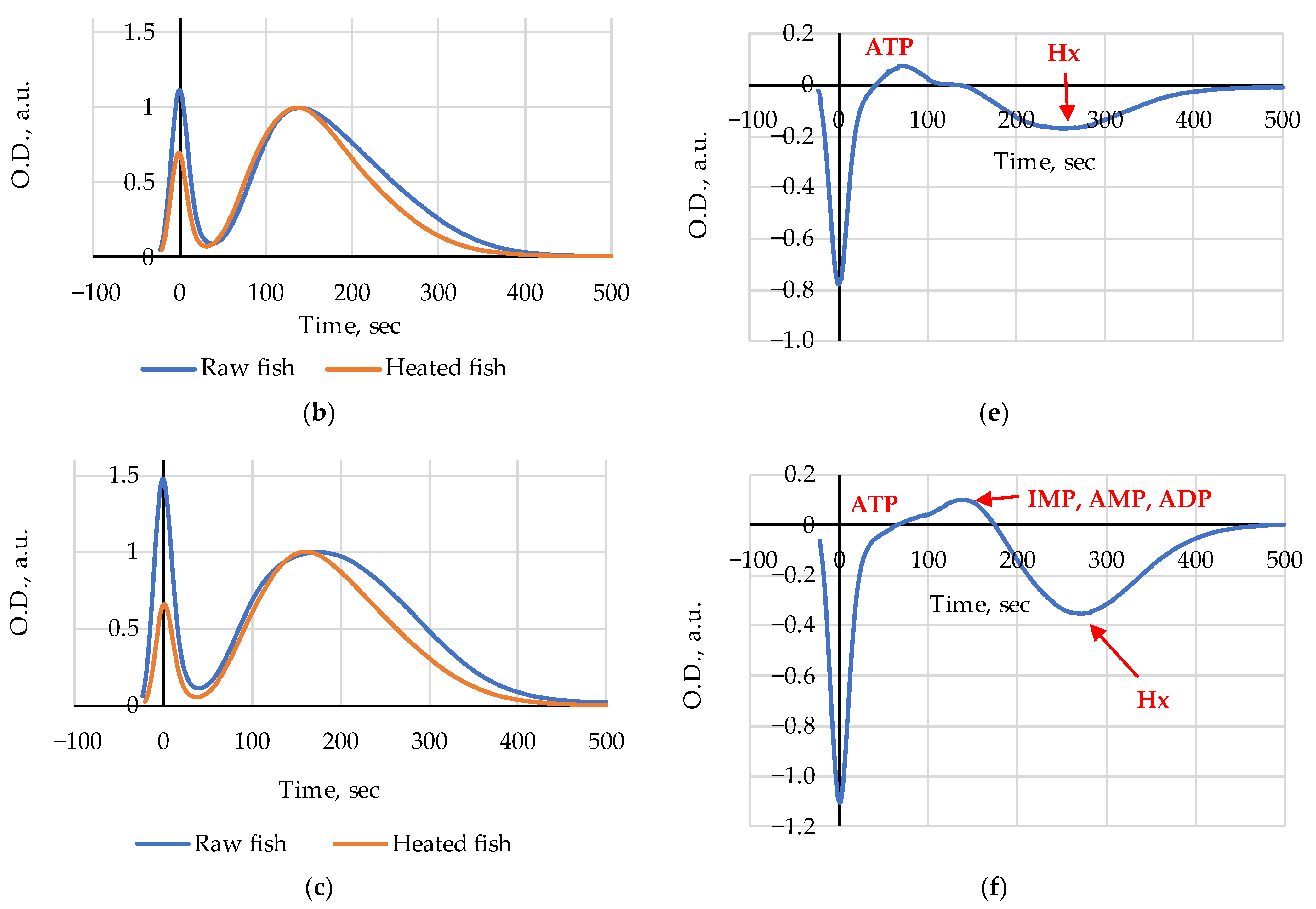
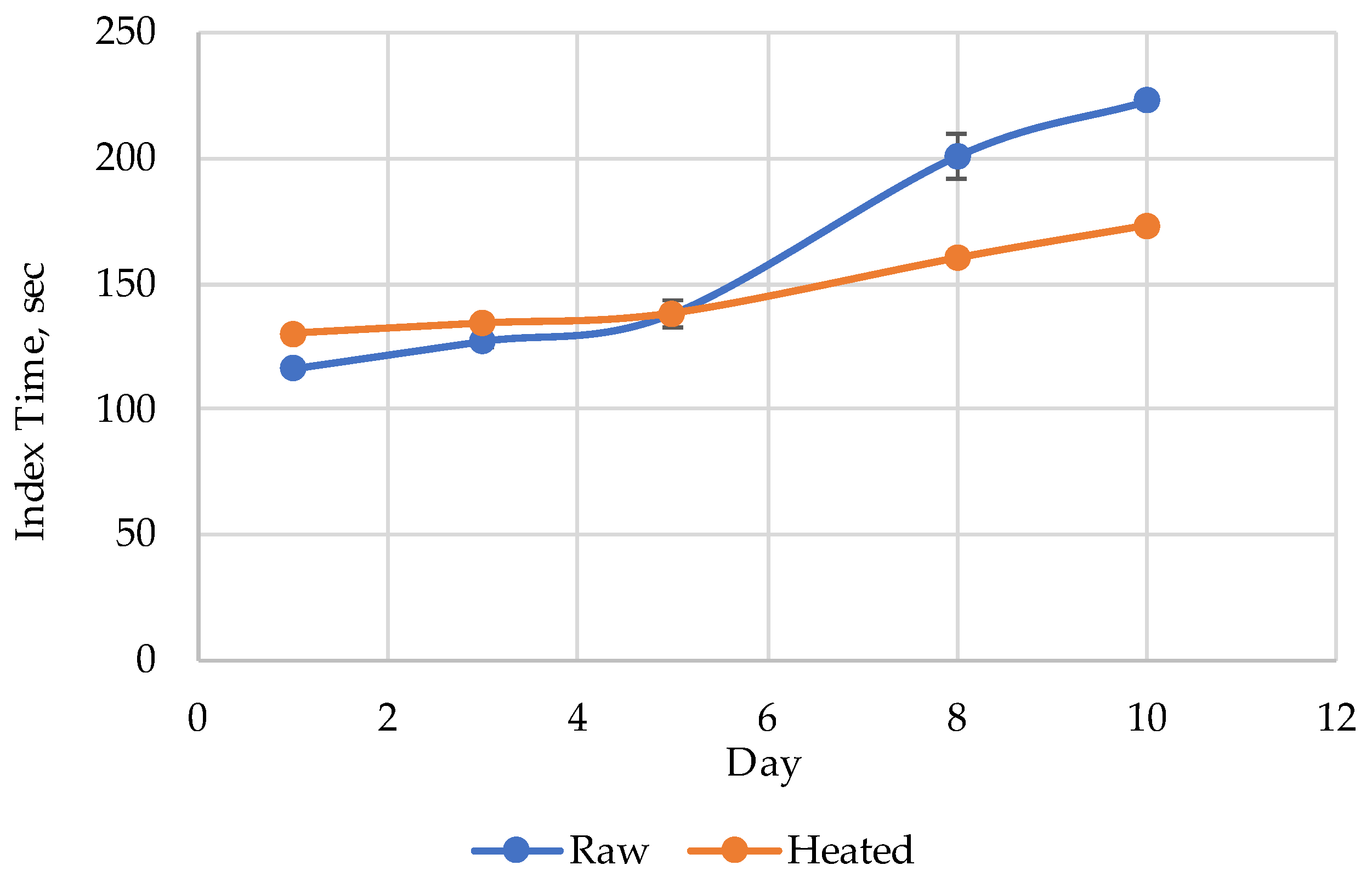
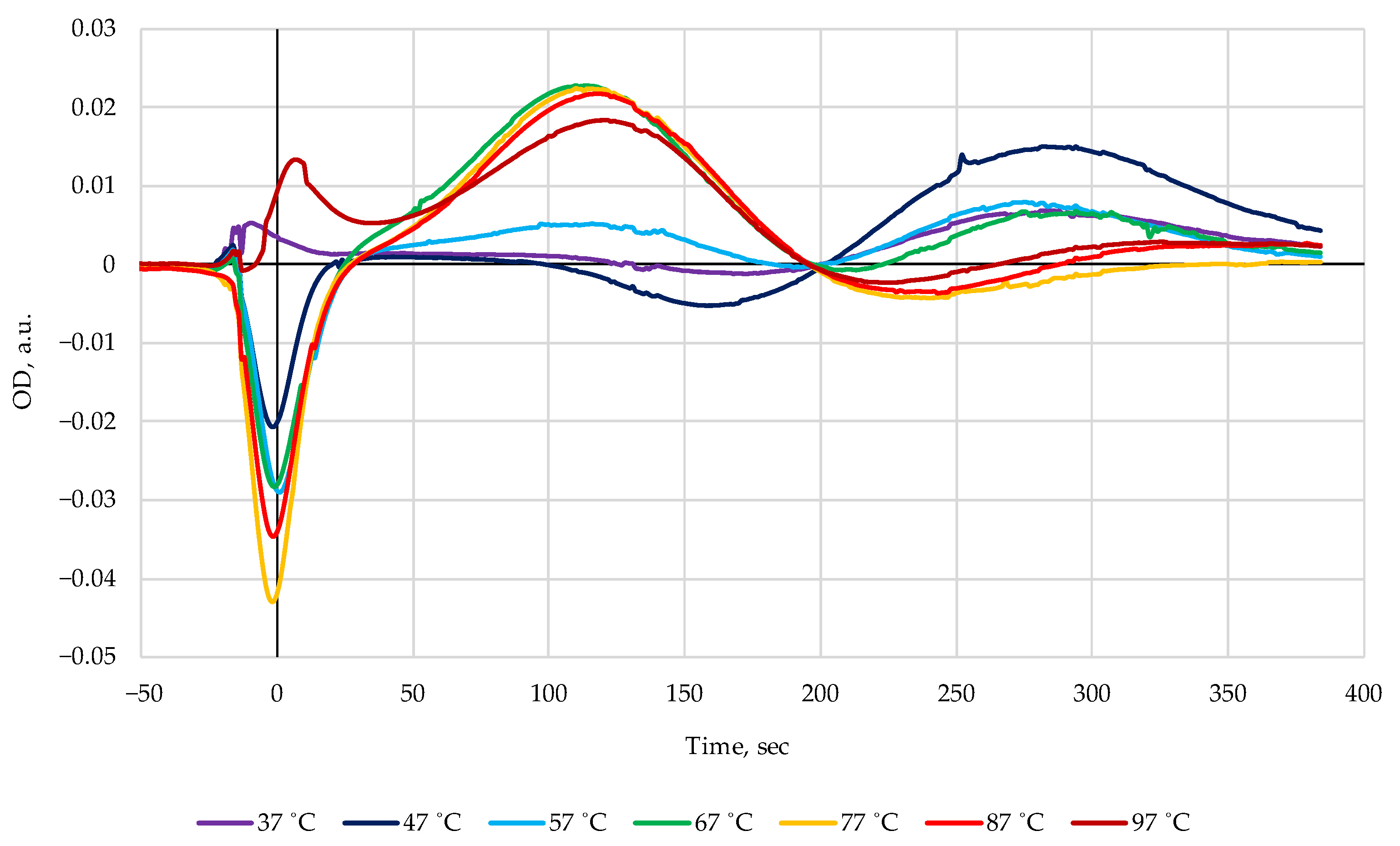
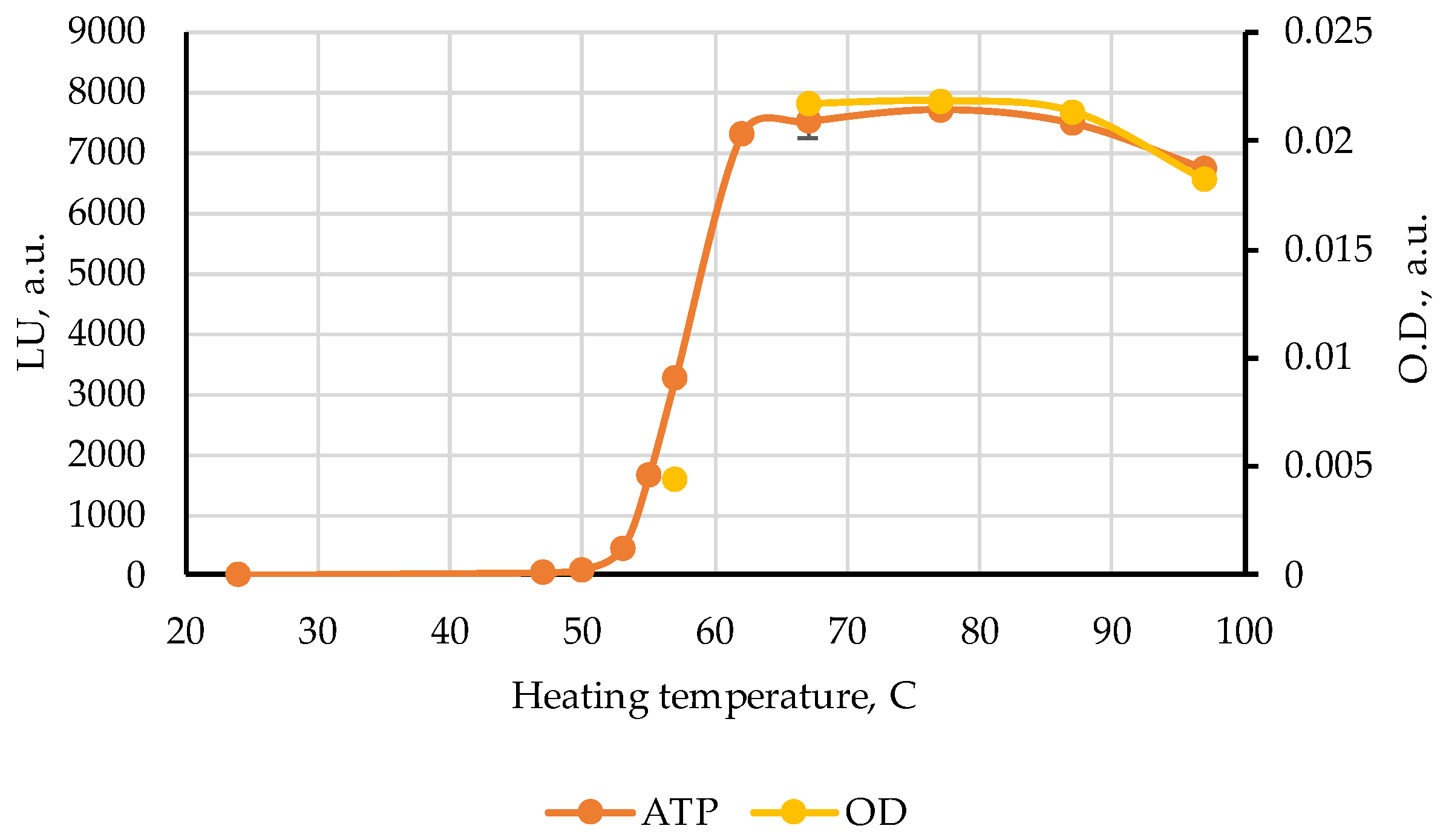
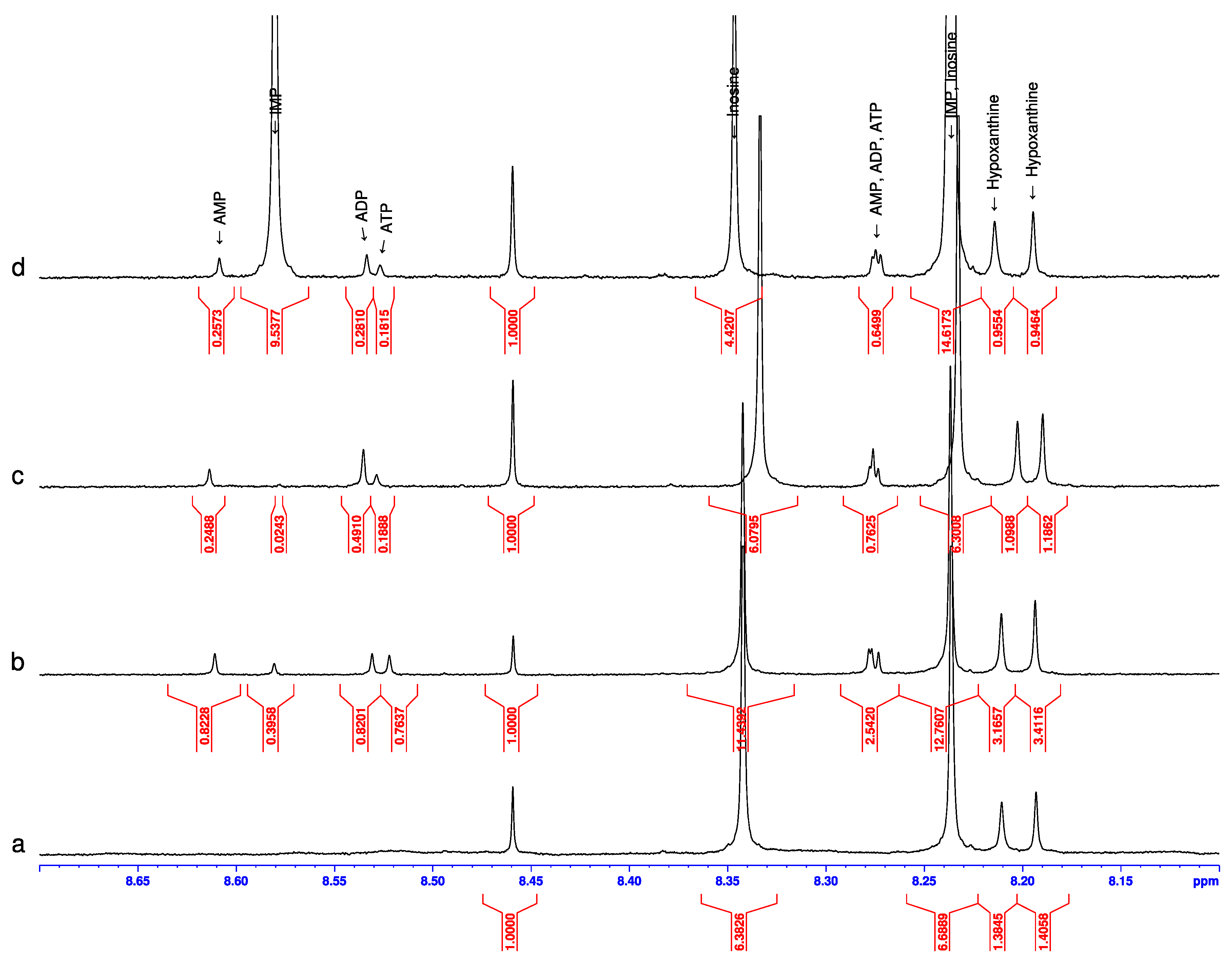
3. Results
4. Discussion
5. Conclusions and Perspectives
- The discovery of the renaissance of adenylate group nucleotides in fish in this work, together with the data on beef [18] and shrimp [20], allows one to conclude with great deal of certainty that this phenomenon is typical and exists in the main food sources: animal meat, fish and, maybe, all shellfish which have undergone conventional heat treatment.
- The fact that we used samples in their macroform for investigations, with no homogenization or protein removal having been carried out, brings this research work as close as possible to reality and enables us to project the results on practice at a 1:1 proportion.
- It is important for further investigations and practice that a triad of techniques has been successfully tested: the classic NMR, the relatively new and low-cost FPMLC as well as the widely known ATP test used in a new context. Such a combination of techniques makes it easy to verify and apply our results both in scientific laboratories equipped with heavy apparatus (NMR) and in industrial or veterinary laboratories and in the field (FPMLC, ATP test).
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, T.; Song, G.; Liu, X.; Xu, M.; Li, Y. Nucleotides as optimal candidates for essential nutrients in living organisms: A review. J. Funct. Foods 2021, 82, 104498. [Google Scholar] [CrossRef]
- Mouritsen, O.G. Umamification of food facilitates the green transition. Soil Ecol. Lett. 2022, 5, 9. [Google Scholar] [CrossRef]
- Nishimura, T.; Ra Rhue, M.; Okitani, A.; Kato, H. Components Contributing to the Improvement of Meat Taste during Storage. Agric. Biol. Chem. 1988, 52, 2323–2330. [Google Scholar] [CrossRef]
- Macy, R.L.; Naumann, H.D.; Bailey, M.E. Water-Soluble Flavor and Odor Precursors of Meat. 3. Changes in Nucleotides, Total Nusleosides and Bases of Beef, Pork and Lamb During Heating. J. Food Sci. 1970, 35, 78–80. [Google Scholar] [CrossRef]
- Macy, R.L.; Naumann, H.D.; Bailey, M.E. Water-Soluble Flavor and Odor Precursors of Meat. 4. Influence of Cooking on Nucleosides and Bases of Beef Steaks and Roasts and Their Relationship to Flavor, Aroma and Juiciness. J. Food Sci. 1970, 35, 81–83. [Google Scholar] [CrossRef]
- Arya, S.S.; Parihar, D.B.; Vijayaraghavan, P.K. Changes in free nucleotides, nucleosides and bases during preparation of pre-cooked dehdyrated minced meats. Nahrung 1979, 23, 495–499. [Google Scholar] [CrossRef]
- Tikk, M.; Tikk, K.; Tørngren, M.A.; Meinert, L.; Aaslyng, M.D.; Karlsson, A.H.; Andersen, H.J. Development of Inosine Monophosphate and Its Degradation Products during Aging of Pork of Different Qualities in Relation to Basic Taste and Retronasal Flavor Perception of the Meat. J. Agric. Food Chem. 2006, 54, 7769–7777. [Google Scholar] [CrossRef]
- Madruga, M.S.; Elmore, J.S.; Oruna-Concha, M.J.; Balagiannis, D.; Mottram, D.S. Determination of some water-soluble aroma precursors in goat meat and their enrolment on flavour profile of goat meat. Food Chem. 2010, 123, 513–520. [Google Scholar] [CrossRef]
- Piskarev, A.; Dibrasulaev, M.; Korzhenko, V. Changes in the level of free amino acids and nucleotides in fresh-killed and aged meat sterilization. Myas. Ind. SSSR 1972, 2, 34–37. (In Russian) [Google Scholar]
- Rotola-Pukkila, M.K.; Pihlajaviita, S.T.; Kaimainen, M.T.; Hopia, A.I. Concentration of Umami Compounds in Pork Meat and Cooking Juice with Different Cooking Times and Temperatures. J. Food Sci. 2015, 80, C2711–C2716. [Google Scholar] [CrossRef]
- Hattula, T.; Kiesvaara, M. Breakdown Products of Adenosine Triphosphate in Heated Fishery Products as an Indicator of Raw Material Freshness and of Storage Quality. LWT-Food Sci. Technol. 1996, 29, 135–139. [Google Scholar] [CrossRef]
- Sasaki, K.; Motoyama, M.; Mitsumoto, M. Changes in the amounts of water-soluble umami-related substances in porcine longissimus and biceps femoris muscles during moist heat cooking. Meat Sci. 2007, 77, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Howgate, P. A review of the kinetics of degradation of inosine monophosphate in some species of fish during chilled storage. Int. J. Food Sci. Technol. 2006, 41, 341–353. [Google Scholar] [CrossRef]
- Vilas, C.; Alonso, A.A.; Herrera, J.R.; García-Blanco, A.; García, M.R. A model for the biochemical degradation of inosine monophosphate in hake (Merluccius merluccius). J. Food Eng. 2017, 200, 95–101. [Google Scholar] [CrossRef]
- Li, D.; Zhuang, S.; Peng, Y.; Tan, Y.; Hong, H.; Luo, Y. Mechanism of Inosine Monophosphate Degradation by Specific Spoilage Organism from Grass Carp in Fish Juice System. Foods 2022, 11, 2672. [Google Scholar] [CrossRef]
- Boziaris, I.S.; Parlapani, F.F. Specific Spoilage Organisms (SSOs) in Fish. In The Microbiological Quality of Food, 1st ed.; Bevilacqua, A., Corbo, M., Sinigaglia, M., Eds.; Elsevier: Cambridge, UK, 2017; pp. 61–98. ISBN 9780081005026. [Google Scholar]
- Li, J.; Zhou, G.; Xue, P.; Dong, X.; Xia, Y.; Regenstein, J.; Du, M.; Sun, L. Spoilage microbes’ effect on freshness and IMP degradation in sturgeon fillets during chilled storage. Food Biosci. 2021, 41, 101008. [Google Scholar] [CrossRef]
- Cambero, M.I.; Pereira-Lima, C.I.; Ordoez, J.A.; Garca de Fernando, G.D. Beef broth flavour: Relation of components with the flavour developed at different cooking temperatures. J. Sci. Food Agric. 2000, 80, 1519–1528. [Google Scholar] [CrossRef]
- Cambero, M.I.; Seuss, I.; Honikel, K.O. Flavor Compounds of Beef Broth as Affected by Cooking Temperature. J. Food Sci. 1992, 57, 1285–1290. [Google Scholar] [CrossRef]
- Cambero, M.I.; Jaramillo, C.J.; Ordoñez, J.A.; Cobos, A.; Pereira-Lima, C.I.; García de Fernando, G.D. Effect of cooking conditions on the flavour compounds and composition of shrimp (Parapenaeus longirostris) broth. Z. Lebensm. Forsch. A 1998, 206, 311–322. [Google Scholar] [CrossRef]
- Kuznetsov, A.; Frorip, A.; Sünter, A.; Kasvand, N.; Korsakov, V.; Konoplev, G.; Stepanova, O.; Rusalepp, L.; Anton, D.; Püssa, T.; et al. Fast Protein and Metabolites (Nucleotides and Nucleosides) Liquid Chromatography Technique and Chemical Sensor for the Assessment of Fish and Meat Freshness. Chemosensors 2023, 11, 69. [Google Scholar] [CrossRef]
- Servetnik, G.E. White Amur—Perspective Object for Reservoirs of Agricultural Purpose. Vestn. Rus. Agric. Sci. 2016, 2, 59–61. (In Russian) [Google Scholar]
- Gui, J.-F.; Tang, Q.; Li, Z.; Liu, J.; de Silva, S.S. Aquaculture in China: Success Stories and Modern Trends, 1st ed.; John Wiley & Sons Ltd.: Chichester, UK, 2018; 677p, ISBN 978-1-119-12076-6. [Google Scholar]
- Food and Agriculture Organization of the United Nations. State of World Fisheries and Aquaculture. 2010. Available online: https://www.fao.org/3/i1820e/i1820e.pdf (accessed on 18 April 2023).
- Singapore Statement on Research Integrity. Available online: https://www.wcrif.org/downloads/main-website/singapore-statements/223-singpore-statement-a4size (accessed on 18 April 2023).
- Montreal Statement on Research Integrity in Cross-Boundary Research Collaborations. Available online: https://www.wcrif.org/downloads/main-website/montreal-statement/123-montreal-statement-english (accessed on 18 April 2023).
- GE Healthcare. Instructions 52-1308-00 BB. Available online: http://wwwuser.gwdg.de/~jgrossh/protocols/protein-purification/PD10.pdf (accessed on 18 April 2023).
- Hong, H.; Regenstein, J.M.; Luo, Y. The importance of ATP-related compounds for the freshness and flavor of post-mortem fish and shellfish muscle: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar] [CrossRef]
- UltraSnap Surface ATP Test—Hygiena. Available online: https://www.hygiena.com/aiovg_videos/ultrasnap-surface-atp-test (accessed on 10 April 2023).
- Zhang, Z.; Sun, Y.; Sang, S.; Jia, L.; Ou, C. Emerging Approach for Fish Freshness Evaluation: Principle, Application and Challenges. Foods 2022, 11, 1897. [Google Scholar] [CrossRef] [PubMed]
- Ishiwatari, N.; Fukuoka, M.; Hamada-Sato, N.; Sakai, N. Decomposition kinetics of umami component during meat cooking. J. Food Eng. 2013, 119, 324–331. [Google Scholar] [CrossRef]
- Greiner, J.V.; Glonek, T. Intracellular ATP Concentration and Implication for Cellular Evolution. Biology 2021, 10, 1166. [Google Scholar] [CrossRef]
- Hwang, Y.-H.; Ismail, I.; Joo, S.-T. Identification of Umami Taste in Sous-Vide Beef by Chemical Analyses, Equivalent Umami Concentration, and Electronic Tongue System. Foods 2020, 9, 251. [Google Scholar] [CrossRef]
- Sarower, M.G.; Hasanuzzaman, A.F.M.D.; Biswas, B.; Abe, H. Taste producing components in fish and fisheries products: A review. J. Food. Ferment. Technol. 2012, 2, 113–121. [Google Scholar]
- Hou, C.; Xiao, G.; Amakye, W.K.; Sun, J.; Xu, Z.; Ren, J. Guidelines for purine extraction and determination in foods. Food Front. 2021, 2, 557–573. [Google Scholar] [CrossRef]
- Gordon, J.L. Extracellular ATP: Effects, sources and fate. Biochem. J. 1986, 233, 309–319. [Google Scholar] [CrossRef]
- Veech, R.L.; Todd King, M.; Pawlosky, R.; Kashiwaya, Y.; Bradshaw, P.C.; Curtis, W. The “great” controlling nucleotide coenzymes. IUBMB Life 2019, 71, 565–579. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 238, Triphosadenine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Triphosadenine#section=Synonyms (accessed on 18 April 2023).
- Arts, I.C.; Coolen, E.J.; Bours, M.J.; Huyghebaert, N.; Stuart, M.A.C.; Bast, A.; Dagnelie, P.C. Adenosine 5′-triphosphate (ATP) supplements are not orally bioavailable: A randomized, placebo-controlled cross-over trial in healthy humans. J. Int. Soc. Sport. Nutr. 2012, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Joy, J.M.; Lowery, R.P.; Roberts, M.D.; Lockwood, C.M.; Manninen, A.H.; Fuller, J.C.; De Souza, E.O.; Baier, S.M.; Wilson, S.M.; et al. Effects of oral adenosine-5′-triphosphate supplementation on athletic performance, skeletal muscle hypertrophy and recovery in resistance-trained men. Nutr. Metab. 2013, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Jäger, R.; Purpura, M.; Rathmacher, J.A.; Fuller, J.C.; Pitchford, L.M.; Rossi, F.E.; Kerksick, C.M. Health and ergogenic potential of oral adenosine-5′-triphosphate (ATP) supplementation. J. Funct. Foods 2021, 78, 104357. [Google Scholar] [CrossRef]
- Purpura, M.; Rathmacher, J.A.; Sharp, M.H.; Lowery, R.P.; Shields, K.A.; Partl, J.M.; Wilson, J.M.; Jäger, R. Oral Adenosine-5′-triphosphate (ATP) Administration Increases Postexercise ATP Levels, Muscle Excitability, and Athletic Performance Following a Repeated Sprint Bout. J. Am. Coll. Nutr. 2017, 36, 177–183. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Res[IMP], % | Kind of Heating | Ref | Note | |
|---|---|---|---|---|
| Goat | 67.8 | Cooked | [6] | Given values |
| Sheep | 65.3 | Cooked | [6] | Given Values |
| Goat | 66 | Grilled | [8] | Given Values |
| Pork | 68.2 | Cooked | [7] | Our estimation |
| Beef | 61.2 | Roasted | [4] | Averaged |
| Trout | 66 | Hot smoking | [11] | Our estimation |
| Beef | 33.9 | Roasted | [5] | Our estimation |
| Substance | Molecular Weight, Da | Elution Time, s |
|---|---|---|
| ATP | 507 | 95 |
| ADP | 427 | 127 |
| AMP | 347 | 176 |
| IMP | 348 | 130 |
| Ino | 268 | 200 |
| Hx | 136 | 275 |
| Meat | AMP, µmol/mL | ADP, µmol/mL | ATP, µmol/mL | T, °C | Heating, Min | Ref. |
|---|---|---|---|---|---|---|
| Pollock | 0.036 | 0.039 | 0.025 | 77 | 40 | This work |
| Shrimp | 0.859 | 0.082 | 0.069 | 75 | 60 | [20] |
| Beef | 0.118 | 0.03 | 0.022 | 75 | 60 | [18] |
| Pork | ≈0.5 | ≈0.1 | 100 | 20 | [3] |
| Food Product | Ki | −ΔKi | T, °C | Duration, min | Environment | Ref |
|---|---|---|---|---|---|---|
| Pollock | 1 | 0.189 | 100 | 40 | Water steam | This work |
| Pollock | 1 | 0.161 | Not measured | 1 | Microwave | This work |
| Beef minced | 0.688 | 0.218 (compared to value at room temperature); 0.275 (compared to value at 55 °C) | 95 | 60 | Broth | [13] |
| Shrimp whole | 0.87 | 0.352 (compared to value at 55 °C) | 95 | 60 | Broth | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sünter, A.; Kuznetsov, A.; Raudsepp, P.; Püssa, T.; Toom, L.; Konoplev, G.; Stepanova, O.S.; Stepanova, O.V.; Lyalin, D.; Frorip, A.; et al. Manifestation of Heat-Induced Valuable Dietary Nucleotide Salvage in Food Prepared from Aged Fish in Fast Protein and Metabolites Liquid Chromatography, ATP-Bioluminescence Assay, and NMR Spectra. AppliedChem 2023, 3, 334-349. https://doi.org/10.3390/appliedchem3020021
Sünter A, Kuznetsov A, Raudsepp P, Püssa T, Toom L, Konoplev G, Stepanova OS, Stepanova OV, Lyalin D, Frorip A, et al. Manifestation of Heat-Induced Valuable Dietary Nucleotide Salvage in Food Prepared from Aged Fish in Fast Protein and Metabolites Liquid Chromatography, ATP-Bioluminescence Assay, and NMR Spectra. AppliedChem. 2023; 3(2):334-349. https://doi.org/10.3390/appliedchem3020021
Chicago/Turabian StyleSünter, Alar, Artur Kuznetsov, Piret Raudsepp, Tõnu Püssa, Lauri Toom, Georgii Konoplev, Oksana S. Stepanova, Oksana V. Stepanova, Daniil Lyalin, Aleksandr Frorip, and et al. 2023. "Manifestation of Heat-Induced Valuable Dietary Nucleotide Salvage in Food Prepared from Aged Fish in Fast Protein and Metabolites Liquid Chromatography, ATP-Bioluminescence Assay, and NMR Spectra" AppliedChem 3, no. 2: 334-349. https://doi.org/10.3390/appliedchem3020021
APA StyleSünter, A., Kuznetsov, A., Raudsepp, P., Püssa, T., Toom, L., Konoplev, G., Stepanova, O. S., Stepanova, O. V., Lyalin, D., Frorip, A., & Roasto, M. (2023). Manifestation of Heat-Induced Valuable Dietary Nucleotide Salvage in Food Prepared from Aged Fish in Fast Protein and Metabolites Liquid Chromatography, ATP-Bioluminescence Assay, and NMR Spectra. AppliedChem, 3(2), 334-349. https://doi.org/10.3390/appliedchem3020021