
Influence of No-Till System with or without Cover Crops on Stomata Sensitivity of Glyphosate-Tolerant Soybeans to Vapor Pressure Deficit


Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Experimental Design
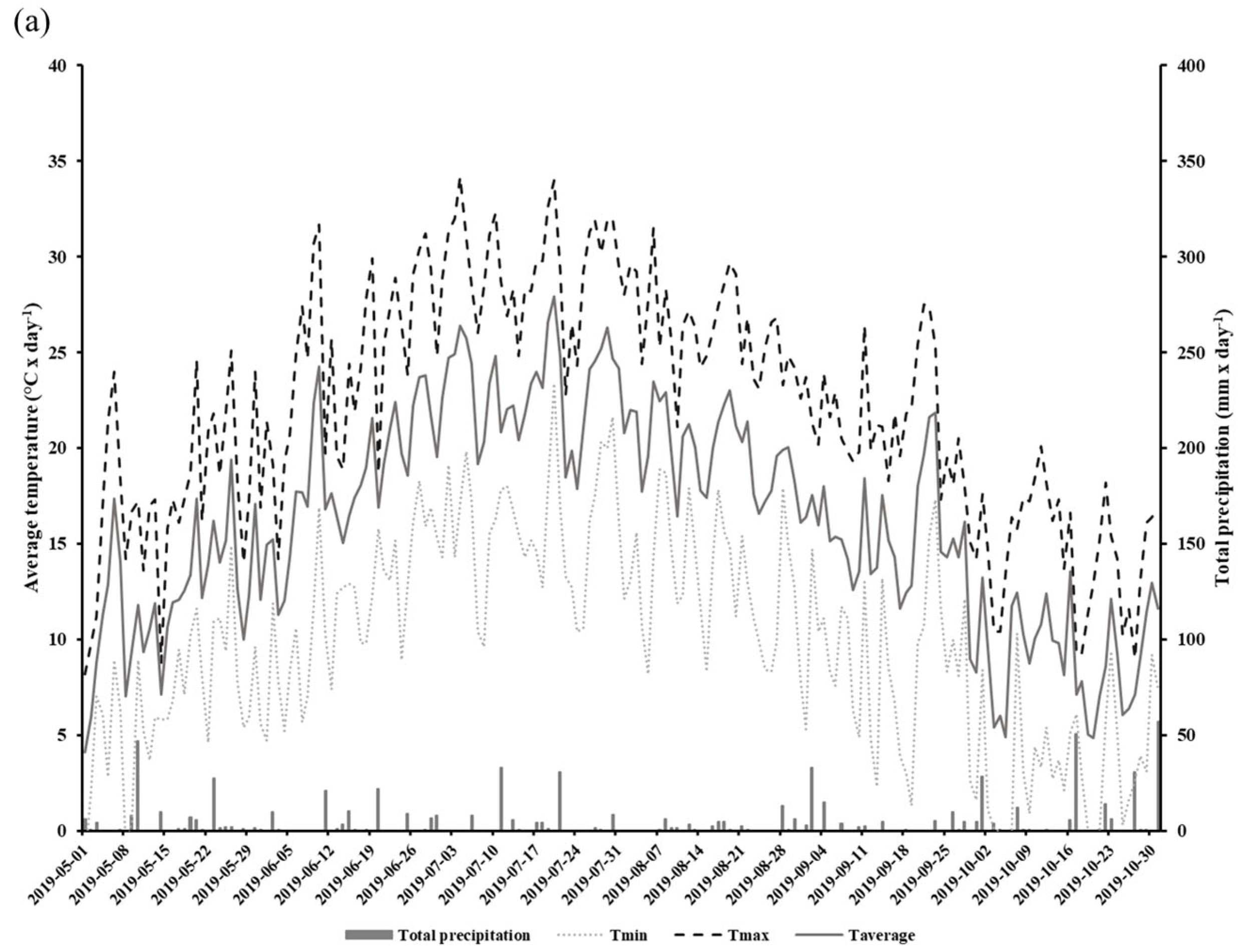
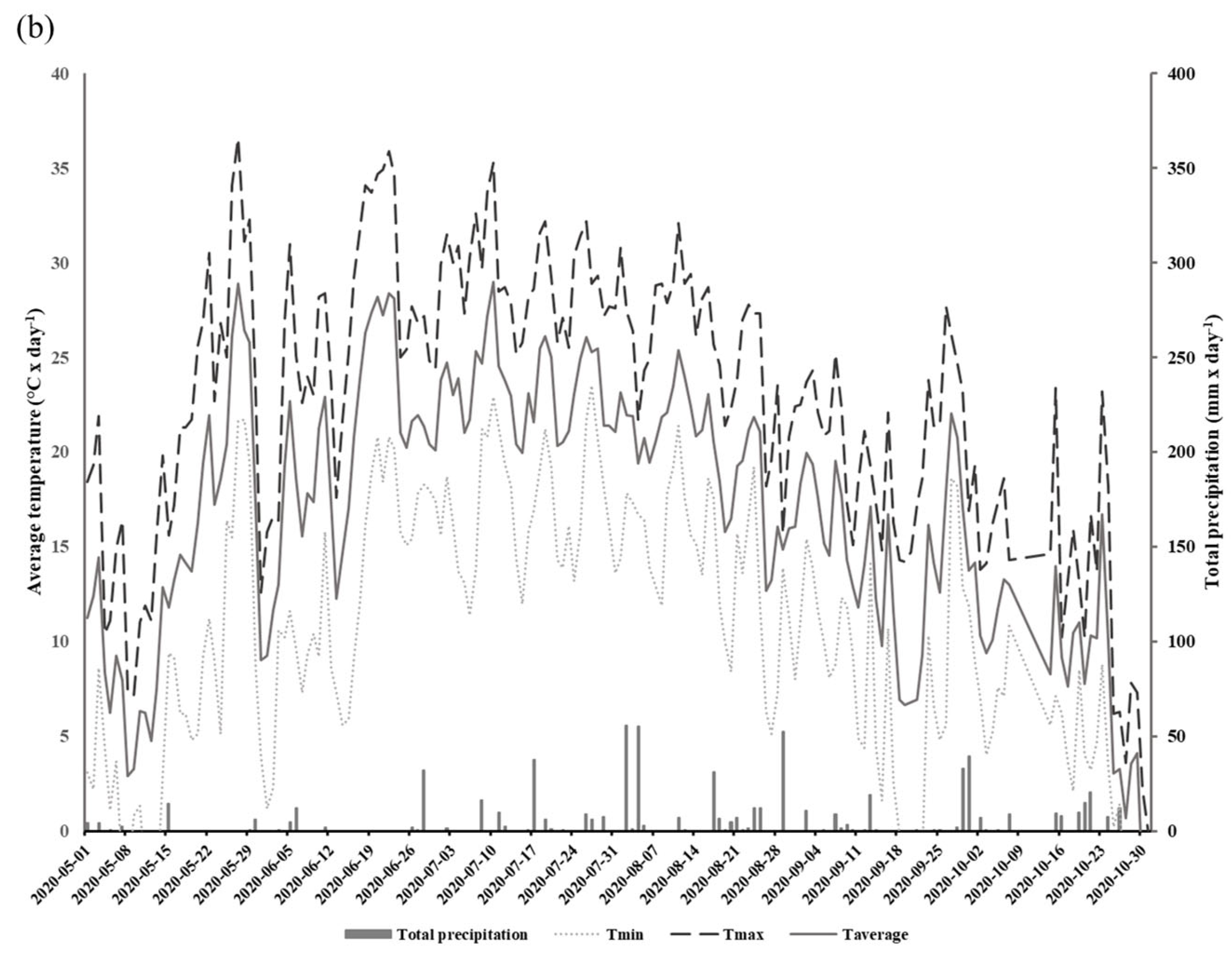
2.2. Sampling and Measurements
Stomatal Conductance and Stomatal Traits
2.3. Foliar Trait
2.4. Statistical Analyses
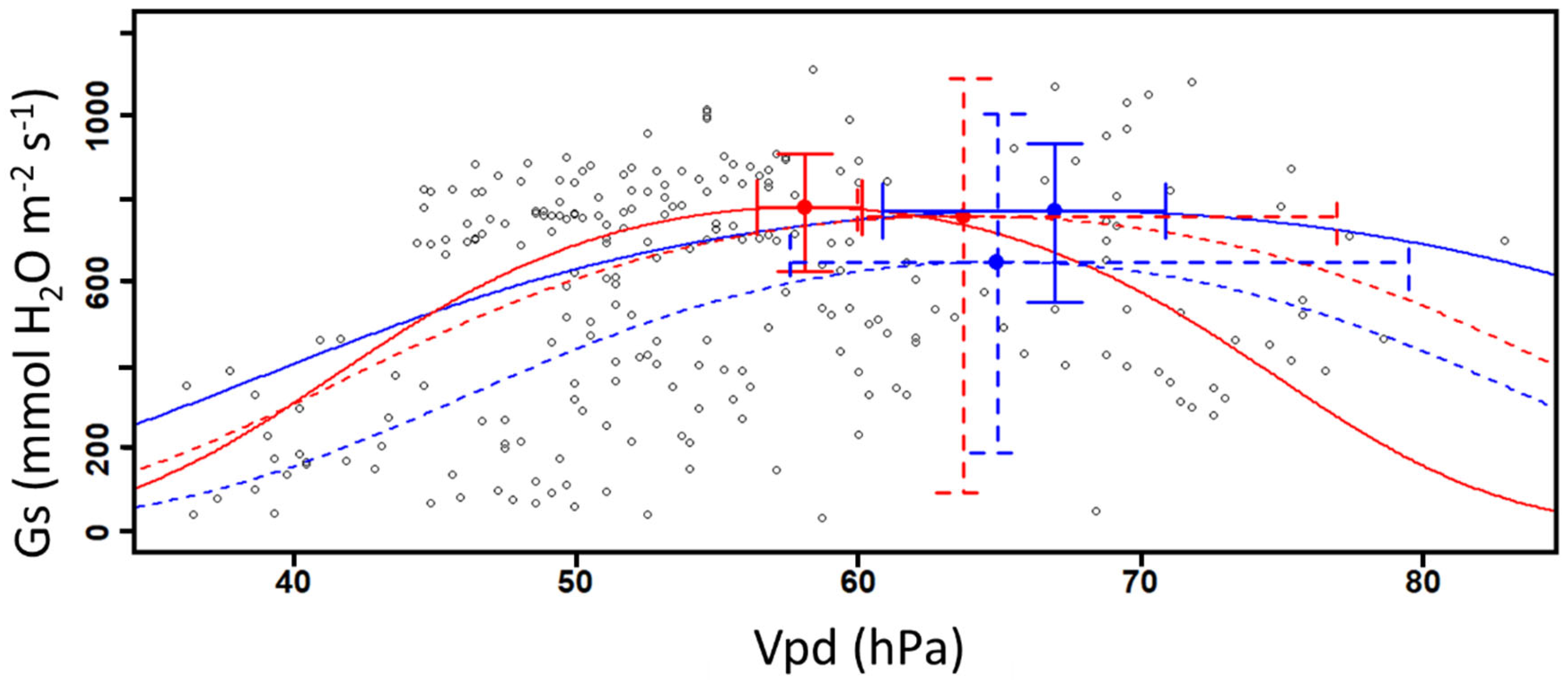
3. Results
3.1. Stomatal Conductance and Vapor Pressure Deficit
3.2. Stomatal Traits
3.3. Foliar Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlson, S.; Stockweel, R. Research priorities for advancing adoption of cover crops in agriculture-intensive regions. J. Agric. Food Syst. Community Dev. 2013, 3, 125–129. [Google Scholar] [CrossRef]
- DeLonge, M.S.; Miles, A.; Carlisle, L. Investing in the transition to sustainable agriculture. Environ. Sci. Policy 2016, 55, 266–273. [Google Scholar] [CrossRef]
- FAO; ITPS. Status of the World’s Soil Ressources; Food and Agricultue Organization of the United Nations: Rome, Italy, 2015; pp. 1–94. [Google Scholar]
- Magdoff, F. Ecological agriculture: Principles, practices, and constraints. Renew. Agric. Food Syst. 2007, 22, 109–117. [Google Scholar] [CrossRef]
- Triplett, G.B.; Dick, W.A. No-tillage crop production: A revolution in agriculture! Agron. J. 2008, 100, 153–166. [Google Scholar] [CrossRef]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Lee, S.; Chua, M.L.; Guzmana, J.A.; Botero-Acostab, A. A comprehensive modeling framework to evaluate soil erosion by water and tillage. J. Environ. Manag. 2021, 279, 111631. [Google Scholar] [CrossRef]
- Kassam, A.; Friedrich, T.; Derpsch, R. Global spread of Conservation Agriculture. Int. J. Environ. Stud. 2019, 76, 29–51. [Google Scholar] [CrossRef]
- Yu, Z.; Lu, C.; Hennessy, D.A.; Feng, H.; Tian, H. Impacts of tillage practices on soil carbon stocks in the US corn-soybean cropping system during 1998 to 2016. Environ. Res. Lett. 2020, 15, 014008. [Google Scholar] [CrossRef]
- Kassam, A.; Friedrich, T.; Derpsch, R. Successful experiences and lessons from conservation agriculture worldwide. Agronomy 2022, 12, 769. [Google Scholar] [CrossRef]
- Domec, J.-C.; Noormets, A.; Gavazzi, M.J.; Bogg, J.L.; King, J.S.; Sun, G.E.; Treasure, E.A. Decoupling the influence of leaf and root hydraulic conductances on stomatal conductance and its sensitivity to vapour pressure deficit as soil dries in a drained loblolly pine plantation. Plant Cell Environ. 2009, 32, 980–991. [Google Scholar] [CrossRef]
- Driesen, E.; Van den Ende, W.; De Proft, M.; Saeys, W. Influence of Environmental Factors Light, CO2, Temperature, and Relative Humidity on Stomatal Opening and Development: A Review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Tanaka, Y.; Fujii, K.; Shiraiwa, T. Variability of leaf morphology and stomatal conductance in soybean [Glycine max (L.) Merr.] cultivars. Crop Sci. 2010, 50, 2525–2532. [Google Scholar] [CrossRef]
- Zeiger, E.; Farquhar, G.D.; Cowan, I.R. Stomatal Function. In The Evolution of Stomata; Stanford University Press: Redwood City, CA, USA, 1987. [Google Scholar]
- Krober, W.; Bruelheide, H. Transpiration and stomatal control: A cross-species study of leaf traits in 39 evergreen and deciduous broadleaved subtropical tree species. Trees 2014, 28, 901–914. [Google Scholar] [CrossRef]
- Hartwig, N.L.; Ammon, H.U. Cover Crops and Living Mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Woolford, A.R.; Jarvis, P.E. Cover, catch and companion crops: Benefits, challenges and economics for Uk growers. Agricology 2017, 1, 1–28. [Google Scholar]
- Amsili, J.P.; Kaye, J.P. Root traits of cover crops and carbon inputs in an organic grain rotation. Renew. Agric. Food Syst. 2021, 36, 182–191. [Google Scholar] [CrossRef]
- Liu, A.; Ma, B.L.; Bomke, A.A. Effects of cover crops on soil aggregate stability, total organic carbon, and polysaccharides. Soil Sci. Soc. Am. J. 2005, 69, 2041–2048. [Google Scholar] [CrossRef]
- Robertson, G.P.; Gross, K.L.; Hamilton, S.K.; Landis, D.A.; Schmidt, T.M.; Snapp, S.S.; Swinton, S.M. Farming for ecosystem services: An ecological approach to production agriculture. BioScience 2014, 64, 404–415. [Google Scholar] [CrossRef]
- Wagg, C.; van Erk, A.; Fava, E.; Comeau, L.-P.; Mitterboeck, T.F.; Goyer, C.; Li, S.; McKenzie-Gopsill, A.; Mills, A. Full-season cover crops and their traits that promote agroecosystem services. Agriculture 2021, 11, 830. [Google Scholar] [CrossRef]
- Roche, D. Stomatal conductance is essential for higher yield potential of C3 crops. Crit. Rev. Plant Sci. 2015, 34, 429–453. [Google Scholar] [CrossRef]
- Seager, R.; Hooks, A.; Parkwilliams, A.; Cook, B.; Nakamura, J.; Henderson, N. Climatology, variability, and trends in the U.S. vapor pressure deficit, an important fire-related meteorological quantity. J. Appl. Meteorol. Climatol. 2015, 54, 1121–1141. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Roberts, M.J.; Schlenker, W.; Braun, N.; Little, B.B.; Rejesus, R.M.; Hammer, G.L. Greater sensitivity to drought accompanies maize yield Increase in the U.S. midwest. Science 2014, 344, 516–519. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Novick, K.A.; Poulter, B.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [PubMed]
- Ocheltree, T.W.; Nippert, J.B.; Prasad, P.V.V. Stomatal responses to changes in vapor pressure deficit reflect tissue-specific differences in hydraulic conductance. Plant Cell Environ. 2014, 37, 132–139. [Google Scholar] [CrossRef]
- Bernier Brillon, J.; Moingt, M.; Lucotte, M. Direct seeding under cover crops: A solution to optimize the potential for adaptation of transgenic field crops to water stress in a context of glyphosate exposure. J. Agric. Crop Res. 2022, 10, 85–97. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Sims, B.; Corsi, S.; Gbehounou, G.; Kienzle, J.; Taguchi, M.; Friedrich, T. Sustainable weed management for conservation agriculture: Options for smallholder farmers. Agriculture 2018, 8, 118. [Google Scholar] [CrossRef]
- Alduchov, O.A.; Eskridge, R.E. Improved Magnus form approximation of saturation vapor pressure. J. Appl. Meteorol. 1996, 35, 601–609. [Google Scholar] [CrossRef]
- Kim, L.; Balani, S.; Edelberg, M.; Macke, N. Effects of various environmental factors on stomatal density, area, and potential conductance index. J. Emerg. Investig. 2021, 4, 1–8. [Google Scholar] [CrossRef]
- Uhl, D.; Mosbrugger, V. Leaf venation density as a climate and environmental proxy: A critical review and new data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1999, 149, 15–26. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Devi, J.; Shekoofa, A.; Choudhary, S.; Sadok, W.; Vadez, V.; Riar, M.; Rufty, T. Limited-transpiration response to high vapor pressure deficit in crop species. Plant Sci. 2017, 260, 109–118. [Google Scholar] [CrossRef]
- Carins Murphy, M.R.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Environ. 2014, 37, 124–131. [Google Scholar] [CrossRef]
- Franks, P.J.; Drake, P.L.; Beerling, D.J. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: An analysis using Eucalyptus globulus. Plant Cell Environ. 2009, 32, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Flexas, J.; Yu, T.; Peng, S.; Huang, J. Leaf anatomy mediates coordination of leaf hydraulic conductance and mesophyll conductance to CO2 in Oryza. New Phytol. 2017, 213, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Puglielli, G.; Catoni, R.; Spoletini, A.; Varone, L.; Gratani, L. Short-term physiological plasticity: Trade-off between drought and recovery responses in three Mediterranean Cistus species. Ecol. Evol. 2017, 7, 10880–10889. [Google Scholar] [CrossRef] [PubMed]
- Scoffoni, C.; Rawls, M.; McKown, A.; Cochard, H.; Lawren Sack, L. Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture. Plant Physiol. 2011, 156, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, H.; Zou, Y.; Wen, Y. Stomatal behaviors reflect enantioselective phytotoxicity of chiral herbicide dichlorprop in Arabidopsis thaliana. Sci. Total Environ. 2016, 562, 73–80. [Google Scholar] [CrossRef]
- Gaskell, M.L.; Pearce, R.B. Stomatal frequency and stomatal resistance of maize hybrids differing in photosynthetic capability. Crop Sci. 1983, 23, 176–177. [Google Scholar] [CrossRef]
- Qi, X.; Torii, K.U. Hormonal and environmental signals guiding stomatal development. BMC Biol. 2018, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Shiraiwa, T. Stem growth habit affects leaf morphology and gas exchange traits in soybean. Ann. Bot. 2009, 104, 1293–1299. [Google Scholar] [CrossRef]
- Tanaka, Y.; Shiraiwa, T.; Nakajima, A.; Sato, J.; Nakazaki, T. Leaf gas exchange activity in soybean as related to leaf traits and stem growth habit. Crop Sci. 2008, 48, 1925–1932. [Google Scholar] [CrossRef]
- Bernier Brillon, J.; Lucotte, M.; Tremblay, G.; Smedbol, E.; Paquet, S. Impacts of glyphosate-based herbicide on leaf stomatal density and biomass production of transgenic soybean (Glycine max [L.] Merr.) and corn (Zea mays L.). Acta Physiol. Plant. 2023, 45, 68. [Google Scholar] [CrossRef]
- Aasama, K.; Sober, A.; Rahi, M. Leaf anatomical characteristics associated with shoot hydraulic conductance and stomatal sensitivity to changes of leaf water status in temperate deciduous trees. Aust. J. Plant Physiol. 2001, 28, 765–774. [Google Scholar] [CrossRef]
- Correia, O.; Ascensão, L. Summer semi-deciduous species of the Mediterranean landscape: A winning strategy of Cistus species to face the predicted changes of the Mediterranean climate. Plant Biodivers. Monit. Assess. Conserv. 2017, 195–217. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mineral Elements | Content Values |
|---|---|
| P | 12.87 ± 2.51 mg kg−1 |
| K | 313.50 ± 20.84 mg kg−1 |
| C | 2.94 ± 0.22 g kg−1 |
| Mg | 803.17 ± 48.27 mg kg−1 |
| Al | 1056.71 ± 19.32 mg kg−1 |
| Cu | 11.00 ± 0.47 mg kg−1 |
| Fe | 24.92 ± 5.72 mg kg−1 |
| Mn | 2.33 ± 0.18 mg kg−1 |
| Na | 47.60 ± 3.06 mg kg−1 |
| 2019 | 2020 | |||
|---|---|---|---|---|
| DSCC | DS | DSCC | DS | |
| Gs (mmol m−2 s−1) | ||||
| mean | 772.45 | 780.00 | 649.15 | 759.07 |
| 95% CI [LL] | 553.56 | 627.52 | 191.77 | 95.61 |
| 95% CI [UL] | 932.89 | 909.13 | 1004.51 | 1088.46 |
| Vpd (hPa) | ||||
| mean | 66.94 | 58.10 | 64.92 | 63.73 |
| 95% CI [LL] | 60.87 | 56.41 | 57.58 | 59.95 |
| 95% CI [UL] | 70.86 | 60.14 | 79.49 | 76.95 |
| 2019 | 2020 | |||||
|---|---|---|---|---|---|---|
| Stomatal Traits | DSCC | DS | p Value | DSCC | DS | p Value |
| StoLength | 15.91 ± 0.23 bB | 16.78 ± 0.23 aA | 0.0068 * | 16.18 ± 0.23 aAB | 16.12 ± 0.24 aAB | 0.8379 |
| StoWidth | 6.75 ± 0.15 bB | 7.60 ± 0.13 aA | <0.0001 * | 6.93 ± 0.10 aB | 6.97 ± 0.09 aB | 0.7506 |
| StoSize | 109.34 ± 3.54 bB | 128.72 ± 3.22 aA | <0.0001 * | 112.60 ± 2.35 aB | 113.43 ± 2.88 aB | 0.8230 |
| StoDen | 307.0 ± 8.3 aA | 282.6 ± 9.5 bB | 0.0481 * | 265.3 ± 9.2 aBC | 247.0 ± 7.9 bC | 0.0247 * |
| StoIndex | 0.0343 ± 0.0023 aAB | 0.0364 ± 0.0017 aA | 0.4481 | 0.0297 ± 0.0011 aBC | 0.0277 ± 0.0010 aC | 0.2129 |
| 2019 | 2020 | |||||
|---|---|---|---|---|---|---|
| DSCC | DS | p Value | DSCC | DS | p Value | |
| Leaf size (cm2) | 11.83 ± 0.48 aA | 11.39 ± 0.43 aA | 0.4912 | 11.77 ± 0.32 aA | 12.47 ± 0.47 aA | 0.2166 |
| Midrib Width (mm) | 9.13 ± 0.28 aA | 7.88 ± 0.22 bAB | 0.001 * | 7.25 ± 0.35 aB | 6.88 ± 0.29 aC | 0.4201 |
| DistVein [34] | 0.63 ± 0.02 bC | 0.80 ± 0.02 aA | 0.001 * | 0.69 ± 0.01 bB | 0.76 ± 0.02 aAB | 0.0442 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernier Brillon, J.; Moingt, M.; Lucotte, M. Influence of No-Till System with or without Cover Crops on Stomata Sensitivity of Glyphosate-Tolerant Soybeans to Vapor Pressure Deficit. Physiologia 2023, 3, 531-541. https://doi.org/10.3390/physiologia3040039
Bernier Brillon J, Moingt M, Lucotte M. Influence of No-Till System with or without Cover Crops on Stomata Sensitivity of Glyphosate-Tolerant Soybeans to Vapor Pressure Deficit. Physiologia. 2023; 3(4):531-541. https://doi.org/10.3390/physiologia3040039
Chicago/Turabian StyleBernier Brillon, Jérôme, Matthieu Moingt, and Marc Lucotte. 2023. "Influence of No-Till System with or without Cover Crops on Stomata Sensitivity of Glyphosate-Tolerant Soybeans to Vapor Pressure Deficit" Physiologia 3, no. 4: 531-541. https://doi.org/10.3390/physiologia3040039
APA StyleBernier Brillon, J., Moingt, M., & Lucotte, M. (2023). Influence of No-Till System with or without Cover Crops on Stomata Sensitivity of Glyphosate-Tolerant Soybeans to Vapor Pressure Deficit. Physiologia, 3(4), 531-541. https://doi.org/10.3390/physiologia3040039