
Bibliometric Analysis Towards Industrial-Scale Use of Marine Algae and Lichens as Soil Amendments and Plant Biofertilizers for Sustainable Agriculture

 , , and
, , and 
Abstract
1. Introduction
2. Bibliometric Analysis of Research on the Use of Seaweeds and Lichens for Agricultural Purposes: Insights from Scopus and Web of Science
2.1. Methods
2.1.1. Descriptive Sources
2.1.2. Methodologies Analysis
2.2. Results of Bibliometric Analysis on the Application of Seaweeds in Agriculture
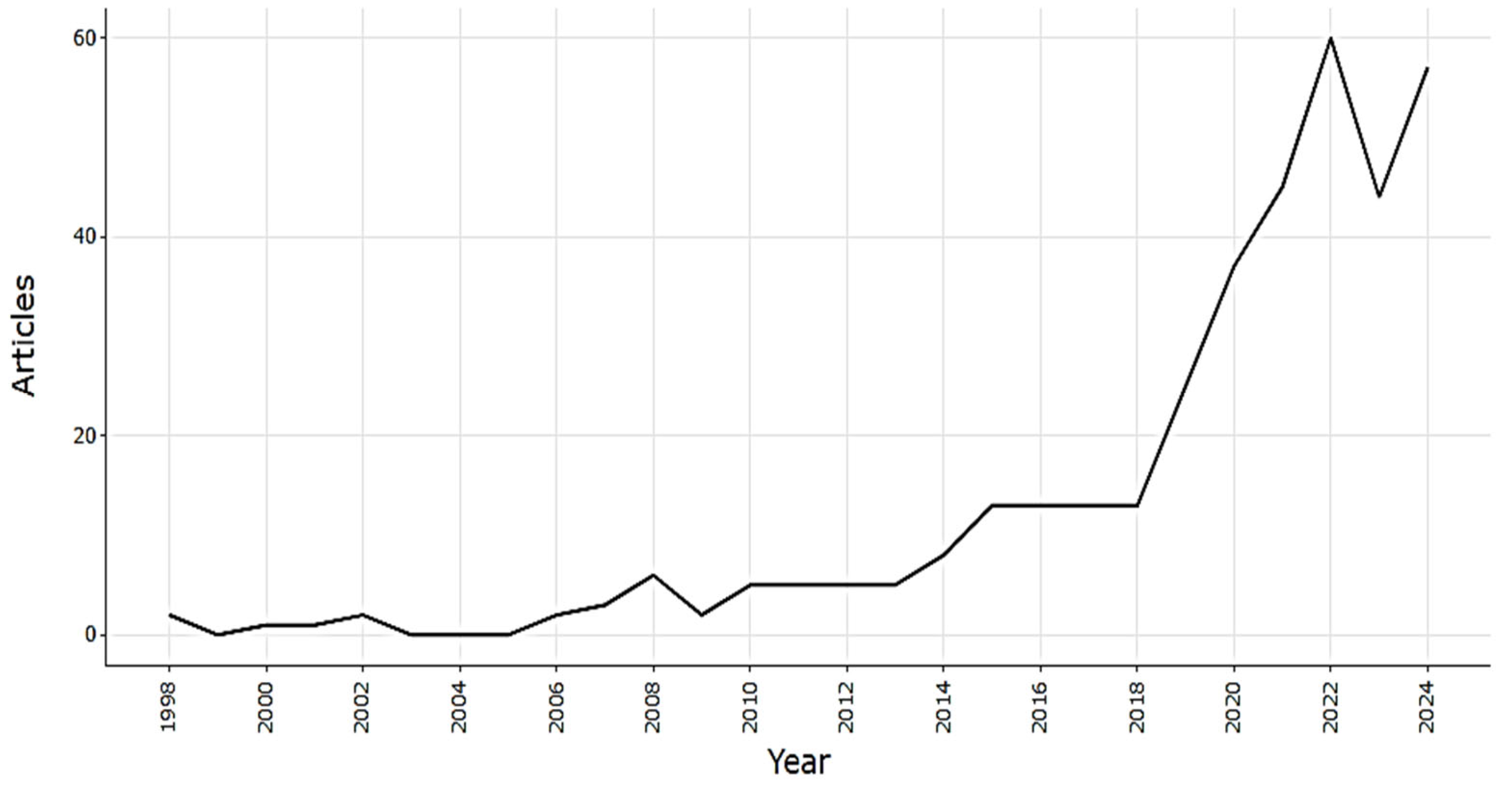
2.2.1. Annual Evolution of the Number of Publications
2.2.2. Keyword Analysis
2.2.3. Three Fields Plot Analysis
2.3. Results of Bibliometric Analysis on the Application of Lichens in Agriculture
2.3.1. Annual Scientific Publication
2.3.2. Keyword Analysis
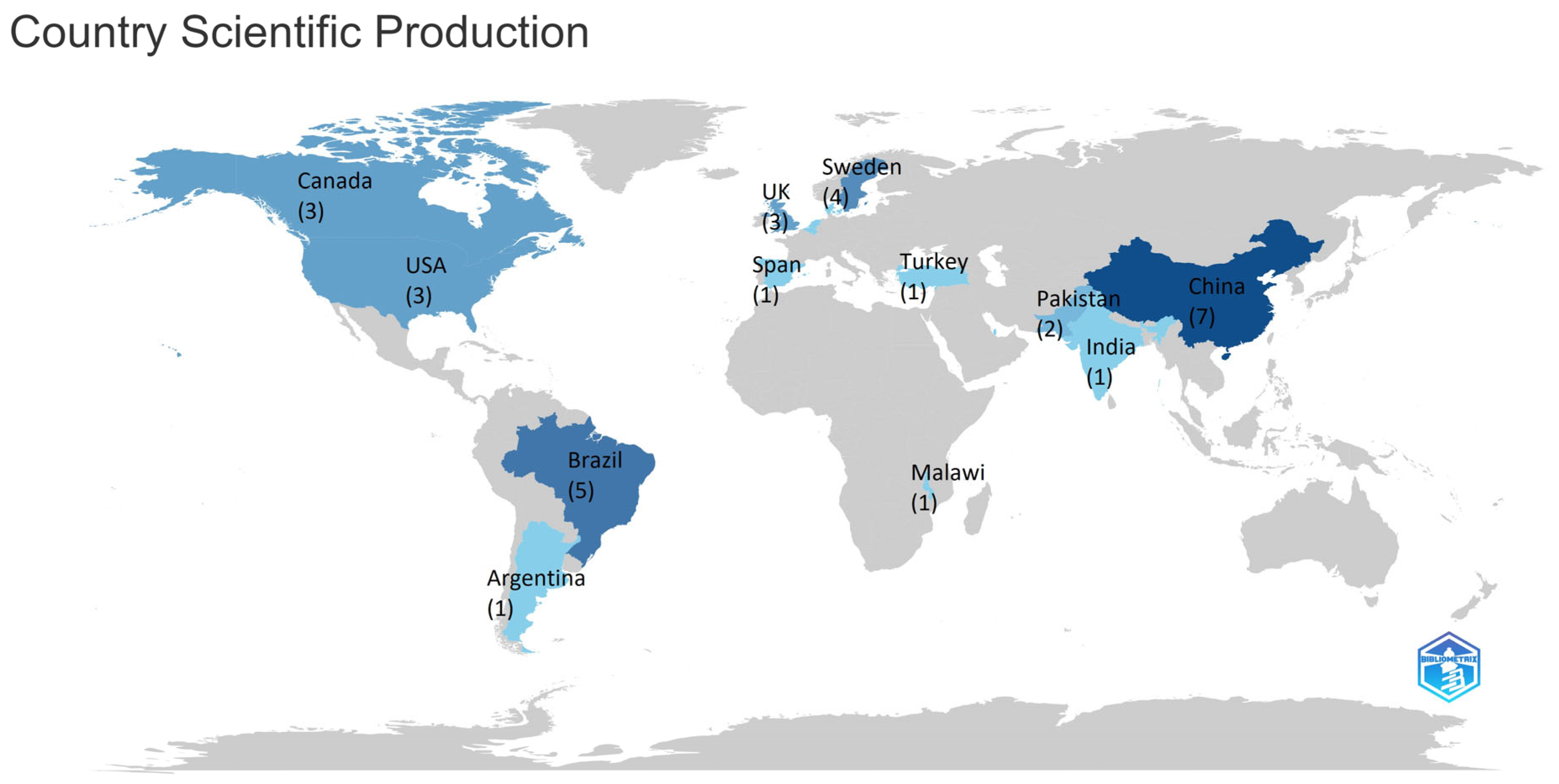
2.3.3. Most Countries Production
2.3.4. Trend Topics Analysis
2.4. Future Orientations Related to the Bibliometric Approach
3. Bibliographical Analysis of the Application of Seaweeds as a Natural Fertilizer to Enhance the Crop Production
3.1. African Seaweed: A Resource for Sustainable Agriculture
3.2. Chemical Composition of Algae
3.2.1. Oligosaccharides
3.2.2. Major Mineral and Trace Elements
3.2.3. Phytohormones
3.3. Extraction of Seaweed Compounds for Agricultural Applications
3.3.1. Aqueous Extract
3.3.2. Organic Extracts
- (a)
- Polysaccharide Extraction
- (b)
- Polyphenol Extraction
- (c)
- Phytohormones Extraction
3.4. Agriculture Application
3.4.1. Effect of Seaweed on Soil Properties
- Chemical properties and availability of nutrients
- Microbial Activity
- (a)
- Bacteria
- (b)
- Fungi
3.4.2. Effect of Seaweed on Plant Growth
- Brown algae
- Green algae
- Red algae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seaweeds | Crops | Method of Application | Effects | Ref. |
|---|---|---|---|---|
| Sargassum wightii Caulerpa chemnitzia | Vigna sinensis | Soaking the seeds for 24 h | The 20% of liquid extracts induced
| [139] |
| Padina gymnospora | Capsicum annum | Soaking the seeds for 24 h | An 8% aqueous extract enhances the germination process of Capsicum annuum, achieving a germination percentage of 70% after 15 days, which is significantly higher than the control group (25%). | [122] |
| Ulva fasciata | Raphanus sativus | Soil incorporation | The application of 10 g dry weight of Ulva fasciata per 500 g of soil resulted in the greatest
| [125] |
| Ecklonia maxima | Eucomis autumnalis | Soil drenching | The application of phenolic compound extract isolated from seaweed at a concentration of 10−6 M improved bulb size and number, fresh weight, root production, and the content of bioactive phytochemicals. | [123] |
| Sargassum wightii | Triticum aestivum | Soaking the seeds for 24 h | The application of a 20% liquid extract resulted in
| [124] |
| Sargassum muticum | Capsicum Annuumm | Soil incorporation | Application of 20 g·kg−1 of seaweed powder improves plant development compared with NPK.
| [11] |
| Lessonia nigrescens Lessonia flavicans | Malus hupehensis | Soil incorporation | Plants treated with 40 g kg−1 of algae showed
| [119] |
3.4.3. Effect Against Biotic Stress
3.4.4. Effect Against Abiotic Stress
3.5. Industrial Development of Seaweed-Based Biofertilizers
4. Bibliographical Analysis of the Potential Impact of Lichens as a Source of Organic Fertilizer
4.1. Diversity of Lichens in Africa
4.2. Biochemical Characterization of Lichen
4.2.1. Polysaccharides
4.2.2. Chlorophyll
4.2.3. Phytohormones
4.2.4. Minerals Elements
4.3. Effect of Lichen on Agricultural Soil
4.3.1. Effect of Lichen on the Physical, Chemical, and Nutritional Characteristics of the Soil
4.3.2. Interactions of Lichen with Soil Microbial Community
4.4. Potential Effect of Lichens on Plant Growth: Future Ecological Opportunity for Sustainable Agriculture
4.5. Agronomic Validation of Lichens: Suggestions for Experimental Axes Towards a New Class of Biofertilizers
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savci, S. Investigation of Effect of Chemical Fertilizers on Environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Nabti, E.; Jha, B.; Hartmann, A. Impact of seaweeds on agricultural crop production as biofertilizer. Int. J. Environ. Sci. Technol. 2017, 14, 1119–1134. [Google Scholar] [CrossRef]
- Gunes, A.; Asian, A.; Turan, M. Biochemical properties of some lichen species as a source of organic fertilizer. Comptes Rendus L’Academie Bulg. Sci. 2016, 69, 539–548. [Google Scholar]
- Pacheco, D.; Cotas, J.; Rocha, C.P.; Araújo, G.S.; Figueirinha, A.; Gonçalves, A.M.M.; Bahcevandziev, K.; Pereira, L. Seaweeds’ carbohydrate polymers as plant growth promoters. Carbohydr. Polym. Technol. Appl. 2021, 2, 100097. [Google Scholar] [CrossRef]
- Pandya, M.; Mehta, S. Effect of Ulva lactuca L. Seaweed Biostimulant on Seed germination, Growth, and some Biochemical properties of Vigna radiata L. Int. J. Environ. Agric. Biotechnol. 2021, 6, 042–053. [Google Scholar] [CrossRef]
- Rao, G.M.N.; Chatterjee, R. Effect of Seaweed Liquid Fertilizer from Gracilaria Textorii and Hypnea Musciformis on Seed Germination and Productivity of Some Vegetable Crops. Univers. J. Plant Sci. 2014, 2, 115–120. [Google Scholar] [CrossRef]
- Ramya, S.S.; Vijayanand, N.; Rathinavel, S. Foliar application of liquid biofertilizer of brown alga Stoechospermum marginatum on growth, biochemical and yield of Solanum melongena. Int. J. Recycl. Org. 2015, 4, 167–173. [Google Scholar] [CrossRef]
- Vinoth, S.; Gurusaravanan, P.; Arun, M.; Saradhadevi, M.; Senthilkumar, N.; Gowtham, P.; Sivakumar, S.R. Biostimulant activity of sulfated polysaccharide extract from red seaweed Halymenia dilatata on yield of Mung bean in greenhouse conditions. J. Appl. Phycol. 2021, 33, 3309–3317. [Google Scholar] [CrossRef]
- Aremu, A.O.; Masondo, N.A.; Rengasamy, K.R.R.; Amoo, S.O.; Gruz, J.; Bíba, O.; Šubrtová, M.; Pěnčík, A.; Novák, O.; Doležal, K.; et al. Physiological role of phenolic biostimulants isolated from brown seaweed Ecklonia maxima on plant growth and development. Planta 2015, 241, 1313–1324. [Google Scholar] [CrossRef]
- Ouala, O.; Essadki, Y.; Redouane, E.M.; Cherifi, O.; El Khalloufi, F.; Oudra, B. Application of Sargassum muticum as a nature-based fertilizer to enhance the growth of Capsicum annuum L. plants. J. Taibah Univ. Sci. 2025, 19, 2484925. [Google Scholar] [CrossRef]
- Omar, H.H.; Abdullatif, B.M.; Al-Kazan, M.M.; El-Gendy, A.M. Various Applications of Seaweed Improves Growth and Biochemical Constituents of Zea mays L. and Helianthus annuus L. J. Plant Nutr. 2015, 38, 28–40. [Google Scholar] [CrossRef]
- Crouch, I.J.; Van Staden, J. Commercial Seaweed Products as Biostimulants in Horticulture. J. Home Consum. Hortic. 1993, 1, 19–76. [Google Scholar] [CrossRef]
- Akila, N.; Jeyadoss, T. The potential of seaweed liquid fertilizer on the growth and antioxidant enhancement of Helianthus annuus L. Orient. J. Chem. 2010, 26, 1353. [Google Scholar]
- Patel, H.D.; Brahmbhatt, N.; Patel, J.; Patel, R.; Thaker, P. Effect of Seaweed Extract on different Vegetables as a Bio Fertilizer in Farming. Int. J. Res. 2019, 7, 2062–2067. [Google Scholar] [CrossRef]
- Pise, N.M.; Sabale, A.B. Effect of seaweed concentrates on the growth and biochemical constituents of Trigonella foenum-graecum L. J. Phytol. 2010, 2010, 50–56. [Google Scholar]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–628. [Google Scholar] [CrossRef]
- Toueileb, S.; Ett, B.L.; Maleinine, D. Effets de l’application foliaire d’ un extrait liquide d’ algue marine (Enteromorophaintestinalis Linnaeus Link.) sur la croissance végétative et la physiologie des jeunes plantes de tomate cultivées sous stress salin. J. New Sci. 2020, 76, 4483–4492. [Google Scholar]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K.Ã. Seaweed extract reduces foliar fungal diseases on carrot. Crop. Prot. 2008, 27, 1360–1366. [Google Scholar] [CrossRef]
- Bano, A.; Waqar, A.; Khan, A.; Tariq, H. Phytostimulants in sustainable agriculture. Front. Sustain. Food Syst. 2022, 6, 801788. [Google Scholar] [CrossRef]
- Aslan, A.; Budak, G.; Karabulut, A. The amounts Fe, Ba, Sr, K, Ca and Ti in some lichens growing in Erzurum province (Turkey). J. Quant. Spectrosc. Radiat. Transf. 2004, 88, 423–431. [Google Scholar] [CrossRef]
- Swamy, C.T.; Gayathri, D.; Devaraja, T.N.; Bandekar, M.; D’Souza, S.E.; Meena, R.M.; Ramaiah, N. Plant growth promoting potential and phylogenetic characteristics of a lichenized nitrogen fixing bacterium, Enterobacter cloacae. J. Basic Microbiol. 2016, 56, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Jannah, M.; A’yun, Q.; Afifah, N.; Prasetya, E.; Hariri, M.R. Usnea in West Java: A potential source of bioactive secondary metabolites. Berk. Penelit. Hayati 2022, 28, 26–31. [Google Scholar] [CrossRef]
- Huneck, S. The significance of lichens and their metabolites. Naturwissenschaften 1999, 86, 559–570. [Google Scholar] [CrossRef]
- Santiago, R.; Silva, N.H.; Silva, F.P.; Martins, M.C.B.; de Vasconcelos, T.L.; Yano-Melo, A.M.; Pereira, E.C. Interactions of the lichen Cladonia salzmannii nyl. With soil, microbiota, mycorrhizae and genipa Americana. J. Soil Sci. Plant Nutr. 2018, 18, 833–850. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Aria, M.; Cuccurullo, C. bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- van Eck, N.J.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M.; Morrison, L.; Rindi, F.; Miranda, S.V.; Mathieson, A.C.; Parker, B.C.; Langangen, A.; John, D.M.; Bárbara, I.; et al. AlgaeBase: An on-line resource for algae. Cryptogam. Algol. 2014, 35, 105–115. [Google Scholar] [CrossRef]
- Bolton, J.J.; De Clerck, O.; John, D.M. Seaweed diversity patterns in sub-Saharan Africa. In Proceedings of the Marine Biodiversity in Sub-Saharan Africa: The Known and the Unknown, Cape Town, South Africa, 24–26 September 2003; pp. 23–26. [Google Scholar]
- Amosu, A.O.; Robertson-Andersson, D.V.; Kean, E.; Maneveldt, G.W.; Cyster, L. Biofiltering and uptake of dissolved nutrients by Ulva armoricana (Chlorophyta) in a land-based aquaculture system. Int. J. Agric. Biol. 2016, 18, 298–304. [Google Scholar] [CrossRef]
- Msuya, F.E.; Buriyo, A.; Omar, I.; Pascal, B.; Narrain, K.; Ravina, J.J.M.; Mrabu, E.; Wakibia, J.G. Cultivation and utilisation of red seaweeds in the Western Indian Ocean (WIO) Region. J. Appl. Phycol. 2014, 26, 699–705. [Google Scholar] [CrossRef]
- Msuya, F.E.; Bolton, J.; Pascal, F.; Narrain, K.; Nyonje, B.; Cottier-Cook, E.J. Seaweed farming in Africa: Current status and future potential. J. Appl. Phycol. 2022, 34, 985–1005. [Google Scholar] [CrossRef]
- Kazzaz, M.; Riadi, H. Inventaire préliminaire de la phycoflore benthique du littoral marocain. II. Rhodophyceae. Acta Bot. Barcinonensia 2000, 46, 53–88. [Google Scholar]
- Bahammou, N.; Cherifi, O.; Bouamama, H.; Rezzoum, N.; Sabri, H.; Boundir, Y. Checklist of rhodophyceae and the first report of Aglaothamnion tripinnatum and Gaillona gallica in the Moroccan coastline. Egypt. J. Aquat. Res. 2021, 47, 101–107. [Google Scholar] [CrossRef]
- Bahammou, N.; Sabri, H.; Cherifi, O.; Bouamama, H. Saesonal variation, the ecological index and the potential use of Chlorophyceae and Phaeophyceae of the Moroccan coast Sidi Bouzid Eljadida in the agricultural field. J. Anal. Sci. Appl. Biotechnol. 2021, 3, 56–64. [Google Scholar] [CrossRef]
- Sabri, H.; Haroun, R.; Bahammou, N.; Hasni, M.; Boundir, Y.; Maarouf, A.; Cherifi, O. Intertidal benthic red algae (Rhodophyta) from Essaouira coastline (Morocco). Egypt. J. Aquat. Res. 2021, 47, 277–282. [Google Scholar] [CrossRef]
- Sabri, H.; Cherifi, O.; Maarouf, A.; Bahammou, N.; Boundir, Y. Original Paper the checklist and the ecological index of the brown seaweeds from essaouira coastline (MOROCCO). J. Appl. Sci. Environ. Stud. JASES 2021, 4, 406–417. [Google Scholar]
- Moussa, H.; Hassoun, M.; Salhi, G.; Zbakh, H.; Riadi, H. Checklist of seaweeds of Al-Hoceima National Park of Morocco (Mediterranean Marine Protected Area). Acta Bot. Malacit. 2018, 43, 91–109. [Google Scholar] [CrossRef]
- Benhissoune, S.; Boudouresque, C.F.; Verlaque, M. A checklist of the seaweeds of the Mediterranean and Atlantic coasts of Morocco. II. Phaeophyceae. Bot. Mar. 2002, 45, 217–230. [Google Scholar] [CrossRef]
- Benhissoune, S.; Boudouresque, C.F.; Perret-Boudouresque, M.; Verlaque, M. A checklist of the seaweeds of the Mediterranean and Atlantic coasts of Morocco. IV. Rhodophyceae—Ceramiales. Bot. Mar. 2003, 46, 55–68. [Google Scholar] [CrossRef]
- Marinho-Soriano, E.; Fonseca, P.C.; Carneiro, M.A.A.; Moreira, W.S.C. Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour. Technol. 2006, 97, 2402–2406. [Google Scholar] [CrossRef]
- Peng, Y.; Xie, E.; Zheng, K.; Fredimoses, M.; Yang, X.; Zhou, X.; Wang, Y.; Yang, B.; Lin, X.; Liu, J.; et al. Nutritional and chemical composition and antiviral activity of cultivated seaweed sargassum naozhouense Tseng et Lu. Mar. Drugs 2013, 11, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Yaich, H.; Garna, H.; Besbes, S.; Paquot, M.; Blecker, C.; Attia, H. Chemical composition and functional properties of Ulva lactuca seaweed collected in Tunisia. Food Chem. 2011, 128, 895–901. [Google Scholar] [CrossRef]
- Zheng, L.X.; Liu, Y.; Tang, S.; Zhang, W.; Cheong, K.L. Preparation methods, biological activities, and potential applications of marine algae oligosaccharides: A review. Food Sci. Hum. Wellness 2023, 12, 359–370. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, Z.P.; Wang, L.N.; Peng, J.X.; Wang, Y.N.; Han, Y.T.; Zhao, S.F. Combined enzymatic hydrolysis and selective fermentation for green production of alginate oligosaccharides from Laminaria japonica. Bioresour. Technol. 2019, 281, 84–89. [Google Scholar] [CrossRef]
- Yudiati, E.; Santosa, G.W.; Tontowi, M.R.; Sedjati, S.; Supriyantini, E.; Khakimah, M. Optimization of alginate alkaline extraction technology from Sargassum polycystum and its antioxidant properties. IOP Conf. Ser. Earth Environ. Sci. 2018, 139, 012052. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Y.; Chen, X.Q.; Aweya, J.J.; Cheong, K.L. Catabolism of Saccharina japonica polysaccharides and oligosaccharides by human fecal microbiota. LWT 2020, 130, 109635. [Google Scholar] [CrossRef]
- Ermakova, S.; Sokolova, R.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Fucoidans from brown seaweeds Sargassum hornery, eclonia cava, costaria costata: Structural characteristics and anticancer activity. Appl. Biochem. Biotechnol. 2011, 164, 841–850. [Google Scholar] [CrossRef]
- Rajauria, G.; Ravindran, R.; Garcia-Vaquero, M.; Rai, D.K.; Sweeney, T.; O’Doherty, J. Purification and Molecular Characterization of Fucoidan Isolated from Ascophyllum nodosum Brown Seaweed Grown in Ireland. Mar. Drugs 2023, 21, 315. [Google Scholar] [CrossRef]
- Wang, S.H.; Huang, C.Y.; Chen, C.Y.; Chang, C.C.; Huang, C.Y.; Dong, C.D.; Chang, J.S. Isolation and purification of brown algae fucoidan from Sargassum siliquosum and the analysis of anti-lipogenesis activity. Biochem. Eng. J. 2021, 165, 107798. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Lee, J.S.; Kim, W.S.; Jeon, Y.J. The potential of brown-algae polysaccharides for the development of anticancer agents: An update on anticancer effects reported for fucoidan and laminaran. Carbohydr. Polym. 2017, 177, 451–459. [Google Scholar] [CrossRef]
- Mohd Fauziee, N.A.; Chang, L.S.; Wan Mustapha, W.A.; Md Nor, A.R.; Lim, S.J. Functional polysaccharides of fucoidan, laminaran and alginate from Malaysian brown seaweeds (Sargassum polycystum, Turbinaria ornata and Padina boryana). Int. J. Biol. Macromol. 2021, 167, 1135–1145. [Google Scholar] [CrossRef]
- Tziveleka, L.A.; Ioannou, E.; Roussis, V. Ulvan, a bioactive marine sulphated polysaccharide as a key constituent of hybrid biomaterials: A review. Carbohydr. Polym. 2019, 218, 355–370. [Google Scholar] [CrossRef]
- Lahaye, M.; Ray, B. Cell-wall polysaccharides from the marine green. Carbohydr. Res. 1996, 283, 161–173. [Google Scholar] [CrossRef]
- Cheong, K.L.; Qiu, H.M.; Du, H.; Liu, Y.; Khan, B.M. Oligosaccharides derived from red seaweed: Production, properties, and potential health and cosmetic applications. Molecules 2018, 23, 2451. [Google Scholar] [CrossRef]
- K-da, S.; Peerakietkhajorn, S.; Siringoringo, B.; Muangnil, P.; Wichienchot, S.; Khuituan, P. Oligosaccharides from Gracilaria fisheri ameliorate gastrointestinal dysmotility and gut dysbiosis in colitis mice. J. Funct. Foods 2020, 71, 104021. [Google Scholar] [CrossRef]
- Meinita, M.D.N.; Marhaeni, B.; Hong, Y.K.; Jeong, G.T. Enzymatic saccharification of agar waste from Gracilaria verrucosa and Gelidium latifolium for bioethanol production. J. Appl. Phycol. 2017, 29, 3201–3209. [Google Scholar] [CrossRef]
- Martínez-Sanz, M.; Gómez-Mascaraque, L.G.; Ballester, A.R.; Martínez-Abad, A.; Brodkorb, A.; López-Rubio, A. Production of unpurified agar-based extracts from red seaweed Gelidium sesquipedale by means of simplified extraction protocols. Algal Res. 2019, 38, 101420. [Google Scholar] [CrossRef]
- Al-Alawi, A.A.; Al-Marhubi, I.M.; Al-Belushi, M.S.M.; Soussi, B. Characterization of Carrageenan Extracted from Hypnea bryoides in Oman. Mar. Biotechnol. 2011, 13, 893–899. [Google Scholar] [CrossRef]
- Zhu, B.; Ni, F.; Xiong, Q.; Yao, Z. Marine oligosaccharides originated from seaweeds: Source, preparation, structure, physiological activity and applications. Crit. Rev. Food Sci. Nutr. 2021, 61, 60–74. [Google Scholar] [CrossRef]
- González, A.; Castro, J.; Vera, J.; Moenne, A. Seaweed Oligosaccharides Stimulate Plant Growth by Enhancing Carbon and Nitrogen Assimilation, Basal Metabolism, and Cell Division. J. Plant Growth Regul. 2013, 32, 443–448. [Google Scholar] [CrossRef]
- El-Mohdy, H.L.A. Radiation-induced degradation of sodium alginate and its plant growth promotion effect. Arab. J. Chem. 2017, 10, S431–S438. [Google Scholar] [CrossRef]
- Yang, J.; Shen, Z.; Sun, Z.; Wang, P.; Jiang, X. Growth Stimulation Activity of Alginate-Derived Oligosaccharides with Different Molecular Weights and Mannuronate/Guluronate Ratio on Hordeum vulgare L. J. Plant Growth Regul. 2021, 40, 91–100. [Google Scholar] [CrossRef]
- Krishna Perumal, P.; Huang, C.Y.; Chen, C.W.; Anisha, G.S.; Singhania, R.R.; Dong, C.D.; Patel, A.K. Advances in oligosaccharides production from brown seaweeds: Extraction, characterization, antimetabolic syndrome, and other potential applications. Bioengineered 2023, 14, 2252659. [Google Scholar] [CrossRef] [PubMed]
- San, P.T.; Khanh, C.M.; Khanh, H.H.N.; Khoa, T.A.; Hoang, N.; Nhung, L.T.; Trinh, N.T.K.; Nguyen, T.D. k-Oligocarrageenan Promoting Growth of Hybrid Maize: Influence of Molecular Weight. Molecules 2020, 25, 3825. [Google Scholar] [CrossRef]
- de Borba, M.C.; de Freitas, M.B.; Stadnik, M.J. Ulvan enhances seedling emergence and reduces Fusarium wilt severity in common bean (Phaseolus vulgaris L.). Crop Prot. 2019, 118, 66–71. [Google Scholar] [CrossRef]
- Salachna, P.; Grzeszczuk, M.; Meller, E.; Soból, M. Oligo-Alginate with low molecular mass improves growth and physiological activity of eucomis autumnalis under salinity stress. Molecules 2018, 23, 812. [Google Scholar] [CrossRef]
- Yi, H.; Xu, G.; Cheng, H.; Wang, J.; Wan, Y.; Chen, H. An Overview of Utilization of Steel Slag. Procedia Environ. Sci. 2012, 16, 791–801. [Google Scholar] [CrossRef]
- Kumar, M.; Kumari, P.; Trivedi, N.; Shukla, M.K.; Gupta, V.; Reddy, C.R.K.; Jha, B. Minerals, PUFAs and antioxidant properties of some tropical seaweeds from Saurashtra coast of India. J. Appl. Phycol. 2011, 23, 797–810. [Google Scholar] [CrossRef]
- Rupérez, P. Mineral content of edible marine seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Sagi, M.; Omarov, R.T.; Lips, S.H. The Mo-hydroxylases xanthine dehydrogenase and aldehyde oxidase in ryegrass as affected by nitrogen and salinity. Plant Sci. 1998, 135, 125–135. [Google Scholar] [CrossRef]
- Shuuluka, D.; Bolton, J.J.; Anderson, R.J. Protein content, amino acid composition and nitrogen-to-protein conversion factors of Ulva rigida and Ulva capensis from natural populations and Ulva lactuca from an aquaculture system, in South Africa. J. Appl. Phycol. 2013, 25, 677–685. [Google Scholar] [CrossRef]
- Mathew, S.; Venkateshwarlu, G.; Ravishankar, C.N. Nutritional profiling of the edible seaweeds Gracilaria edulis, Ulva lactuca and Sargassum sp. Indian J. Fish. 2016, 63, 81–87. [Google Scholar] [CrossRef]
- El-said, G.F.; El-sikaily, A. Chemical composition of some seaweed from Mediterranean. Environ. Monit. Assess. 2013, 185, 6089–6099. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V. High-value products from macroalgae: The potential uses of the invasive brown seaweed, Sargassum muticum. Rev. Environ. Sci. Bio/Technol. 2016, 15, 67–88. [Google Scholar] [CrossRef]
- Taboada, M.C.; Millán, R.; Miguez, M.I. Nutritional value of the marine algae wakame (Undaria pinnatifida) and nori (Porphyra purpurea) as food supplements. J. Appl. Phycol. 2013, 25, 1271–1276. [Google Scholar] [CrossRef]
- Milledge, J.J.; Staple, A.; Harvey, P.J. Slow Pyrolysis as a Method for the Destruction of Japanese Wireweed, Sargassum muticum. Environ. Nat. Resour. Res. 2015, 5, 28–37. [Google Scholar] [CrossRef]
- Balboa, E.M.; Gallego-fábrega, C.; Moure, A.; Domínguez, H. Study of the seasonal variation on proximate composition of oven-dried Sargassum muticum biomass collected in Vigo Ria, Spain. J. Appl. Phycol. 2016, 28, 1943–1953. [Google Scholar] [CrossRef]
- Mawi, S.; Krishnan, S.; Din, M.F.; Arumugam, N.; Chelliapan, S. Bioremediation potential of macroalgae Gracilaria edulis and Gracilaria changii co-cultured with shrimp wastewater in an outdoor water recirculation system. Environ. Technol. Innov. 2019, 17, 100571. [Google Scholar] [CrossRef]
- Saebmehr, H.; Rafiee, F.; Givianrad, M.H.; Mostafavi, G. Determination and quantification of two regulatory phytohormones extracted from Sargassum muticum and Gracilaria corticata seaweeds in the south of Iran. Nova Biol. Reper. 2022, 9, 115–123. [Google Scholar] [CrossRef]
- Saebmehr, H.; Rafiee, F.; Givianrad, M.H.; Mostafavi, G. Extraction of abscisic acid and gibberellin from Sargassum muticum (Phaeophyceae) and Gracilaria corticata (Rhodophyta) harvested from Persian Gulf. Iran. J. Fish. Sci. 2022, 21, 590–604. [Google Scholar] [CrossRef]
- Ben, I.; Karen, A.; Ledezma, D.; Mart, E.; Leyva, S.; Carrera, E.; Ruiz, I.O. Identification and Quantification of Plant Growth Regulators and Antioxidant Compounds in Aqueous Extracts of Padina durvillaei and Ulva lactuca. Agronomy 2020, 10, 866. [Google Scholar] [CrossRef]
- Urbanek Krajnc, A.; Turinek, M.; Ivančič, A. Morphological and physiological changes during adventitious root formation as affected by auxin metabolism: Stimulatory effect of auxin containing seaweed extract treatment. Agricultura 2013, 1, 17–27. [Google Scholar]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.H. Cytokinin-containing seaweed and humic acid extracts associated with creeping bentgrass leaf cytokinins and drought resistance. Crop Sci. 2004, 44, 1737–1745. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.H. Impact of seaweed extract-based cytokinins and zeatin riboside on creeping bentgrass heat tolerance. Crop Sci. 2008, 48, 364–370. [Google Scholar] [CrossRef]
- Mahendran, S.; Maheswari, P.; Sasikala, V. In vitro antioxidant study of polyphenol from red seaweeds dichotomously branched gracilaria Gracilaria edulis and robust sea moss Hypnea valentiae. Toxicol. Rep. 2021, 8, 1404–1411. [Google Scholar] [CrossRef]
- Górka, B.; Wieczorek, P.P. Simultaneous determination of nine phytohormones in seaweed and algae extracts by HPLC-PDA. J. Chromatogr. B 2017, 1057, 32–39. [Google Scholar] [CrossRef]
- Yalçln, S.; Okudan, E.Å.; Karakaş, Ö.; Önem, A.N. Determination of Major Phytohormones in Fourteen Different Seaweeds Utilizing SPE-LC-MS/MS. J. Chromatogr. Sci. 2020, 58, 98–108. [Google Scholar] [CrossRef]
- Godlewska, K.; Michalak, I.; Tuhy, L.; Chojnacka, K. Plant Growth Biostimulants Based on Different Methods of Seaweed Extraction with Water. Biomed Res. Int. 2016, 2016, 5973760. [Google Scholar] [CrossRef]
- Hughes, H.; Mcloughlin, P.; Tan, S.P.; Keeffe, E.O. Antibacterial Activity of Seaweed Extracts against Plant Pathogenic Bacteria Antibacterial Activity of Seaweed Extracts against Plant Pathogenic Bacteria. J. Bacteriol. Mycol. 2019, 6, 1105. [Google Scholar]
- Kasim, W.A.; Hamada, E.A.M.; El-din, N.G.S.; Eskander, S.; Agri, I.J.; Agri, R. Influence of seaweed extracts on the growth, some metabolic activities and yield of wheat grown under drought stress. Int. J. Agron. Agric. Res. 2015, 7, 173–189. [Google Scholar]
- He, J.; Xu, Y.; Chen, H.; Sun, P. Extraction, Structural Characterization, and Potential Antioxidant Activity of the Polysaccharides from Four Seaweeds. Int. J. Mol. Sci. 2016, 17, 1988. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Wang, W.; Zhao, X.; Zhang, J.; Ewart, S.H. Properties of polysaccharides in several seaweeds from Atlantic Canada and their potential anti-influenza viral activities. J. Ocean Univ. China 2012, 11, 205–212. [Google Scholar] [CrossRef]
- Dobrinčić, A.; Dobroslavić, E.; Pedisić, S.; Balbino, S.; Elez Garofulić, I.; Čož-Rakovac, R.; Dragović-Uzelac, V. The effectiveness of the Fucus virsoides and Cystoseira barbata fucoidan isolation as a function of applied pre-treatment and extraction conditions. Algal Res. 2021, 56, 102286. [Google Scholar] [CrossRef]
- Mzibra, A.; Aasfar, A.; El Arroussi, H.; Khouloud, M.; Dhiba, D.; Kadmiri, I.M.; Bamouh, A. Polysaccharides extracted from Moroccan seaweed: A promising source of tomato plant growth promoters. J. Appl. Phycol. 2018, 30, 2953–2962. [Google Scholar] [CrossRef]
- Hamouda, R.A.; Hussein, M.H.; El-Naggar, N.E.A.; Karim-Eldeen, M.A.; Alamer, K.H.; Saleh, M.A.; Al Masoudi, L.M.; Sharaf, E.M.; El-Azeem, R.M.A. Promoting Effect of Soluble Polysaccharides Extracted from Ulva spp. on Zea mays L. Growth. Molecules 2022, 27, 1394. [Google Scholar] [CrossRef]
- Paulert, R.; Talamini, V.; Cassolato, J.E.F.; Duarte, M.E.R.; Noseda, M.D.; Smania, A.; Stadnik, M.J. Effects of sulfated polysaccharide and alcoholic extracts from green seaweed Ulva fasciata on anthracnose severity and growth of common bean (Phaseolus vulgaris L.). J. Plant Dis. Prot. 2009, 116, 263–270. [Google Scholar] [CrossRef]
- Ben Salah, I.; Aghrouss, S.; Douira, A.; Aissam, S.; El, Z.; Filali-maltouf, A.; El Modafar, C. Seaweed polysaccharides as bio-elicitors of natural defenses in olive trees against verticillium wilt of olive. J. Plant Interact. 2018, 13, 248–255. [Google Scholar] [CrossRef]
- Kisiriko, M.; Anastasiadi, M.; Terry, L.A.; Yasri, A.; Beale, M.H.; Ward, J.L. Phenolics from medicinal and aromatic plants: Characterisation and potential as biostimulants and bioprotectants. Molecules 2021, 26, 6343. [Google Scholar] [CrossRef]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Extraction of phenolic compounds: A review. Curr. Res. Food Sci. 2021, 4, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Mori, I.C.; Ikeda, Y.; Matsuura, T.; Hirayama, T.; Mikami, K. Phytohormones in red seaweeds: A technical review of methods for analysis and a consideration of genomic data. Bot. Mar. 2017, 60, 153–170. [Google Scholar] [CrossRef]
- Sanderson, K.J.; Jameson, P.E.; Zabkiewicz, J.A. Auxin in a Seaweed Extract: Identification and Quantitation of Indole-3-acetic acid by Gas Chromatography-Mass Spectrometry. J. Plant Physiol. 1987, 129, 363–367. [Google Scholar] [CrossRef]
- Tay, S.A.B.; Macleod, J.K.; Palni, L.M.S.; Letham, D.S. Detection of cytokinins in a seaweed extract. Phytochemistry 1985, 24, 2611–2614. [Google Scholar] [CrossRef]
- Sulakhudin, S.; Hatta, M.; Suryadi, U.E. Application of coastal sediments and foliar seaweed extract and its influence to soil properties, growth and yield of shallot in peatland. AGRIVITA J. Agric. Sci. 2019, 41, 450–460. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; EL-Shershaby, N.A. Algae as Bio-fertilizers: Between current situation and future prospective: The role of Algae as a Bio-fertilizer in serving of ecosystem. Saudi J. Biol. Sci. 2022, 29, 3083–3096. [Google Scholar] [CrossRef]
- Kumari, R.; Kaur, I.; Bhatnagar, A.K. Enhancing soil health and productivity of Lycopersicon esculentum Mill. using Sargassum johnstonii Setchell Gardner as a soil conditioner and fertilizer. J. Appl. Phycol. 2013, 25, 1225–1235. [Google Scholar] [CrossRef]
- Arthur, G.D.; Aremu, A.O.; Moyo, M.; Stirk, W.A.; Van Staden, J. Growth-promoting effects of a seaweed concentrate at various pH and water hardness conditions. S. Afr. J. Sci. 2013, 109, 6. [Google Scholar] [CrossRef]
- Adderley, A.; Wallace, S.; Stubbs, D.; Connor, C.B.O.; Ferguson, J. Sargassum sp. as a biofertilizer: Is it really a key towards sustainable agriculture for The Bahamas? Bull. Natl. Res. Cent. 2023, 47, 112. [Google Scholar] [CrossRef]
- Msimbira, L.A.; Smith, D.L. The Roles of Plant Growth Promoting Microbes in Enhancing Plant Tolerance to Acidity and Alkalinity Stresses. Front. Sustain. Food Syst. 2020, 4, 106. [Google Scholar] [CrossRef]
- Hussain, H.I.; Kasinadhuni, N.; Arioli, T. The effect of seaweed extract on tomato plant growth, productivity and soil. J. Appl. Phycol. 2021, 33, 1305–1314. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Huang, Z.; Su, G.; Li, X.; Sun, Z.; Qin, Y. Impact of short-term application of seaweed fertilizer on bacterial diversity and community structure, soil nitrogen contents, and plant growth in maize rhizosphere soil. Folia Microbiol. 2020, 65, 591–603. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Li, Y.; Chen, L.; Liu, Z.; Wang, X. Responses of soil microbial communities to a short-term application of seaweed fertilizer revealed by deep amplicon sequencing. Appl. Soil Ecol. 2018, 125, 288–296. [Google Scholar] [CrossRef]
- Zhou, G.; Qiu, X.; Zhang, J.; Tao, C. Bioresource Technology E ffects of seaweed fertilizer on enzyme activities, metabolic characteristics, and bacterial communities during maize straw composting. Bioresour. Technol. 2019, 286, 121375. [Google Scholar] [CrossRef]
- Qiqin, L.; Huaguang, Z.; Minxiu, S.; Qian, L.; Haijun, F.; Haimin, C.; Rui, Y. Improvement of Soil Structure and Bacterial Composition by Long-Term Application of Seaweed Fertilizer. J. Soil Sci. Plant Nutr. 2023, 23, 5122–5132. [Google Scholar] [CrossRef]
- Espinosa-Antón, A.A.; Zamora-Natera, J.F.; Zarazúa-Villaseñor, P.; Santacruz-Ruvalcaba, F.; Sánchez-Hernández, C.V.; Águila Alcántara, E.; Torres-Morán, M.I.; Velasco-Ramírez, A.P.; Hernández-Herrera, R.M. Application of Seaweed Generates Changes in the Substrate and Stimulates the Growth of Tomato Plants. Plants 2023, 12, 1520. [Google Scholar] [CrossRef]
- Wang, Y.; Xiang, L.; Wang, S.; Wang, X.; Chen, X.; Mao, Z. Effects of seaweed fertilizer on the Malus hupehensis Rehd. seedlings growth and soil microbial numbers under continue cropping. Acta Ecol. Sin. 2017, 37, 180–186. [Google Scholar] [CrossRef]
- Mahmoud, S.H.; Salama, D.M.; El-Tanahy, A.M.M.; Abd El-Samad, E.H. Utilization of seaweed (Sargassum vulgare) extract to enhance growth, yield and nutritional quality of red radish plants. Ann. Agric. Sci. 2019, 64, 167–175. [Google Scholar] [CrossRef]
- Ramarajan, S.; Ganthi, S. Effect of Seaweed Liquid Fertilizer on the Germination and Pigment Concentration of Soybean. J. Crop Sci. Technol. 2012, 1, 1–15. [Google Scholar]
- Thriunavukkarasu, R.; Joseph, J.; Aruni, W. Effect of seaweed on seed germination and biochemical constituents of Capsicum annuum. Biocatal. Agric. Biotechnol. 2020, 29, 101761. [Google Scholar]
- Ahmed, D.A.E. Efficacy of two seaweeds dry mass in bioremediation of heavy metal polluted soil and growth of radish (Raphanus sativus L.) plant. Environ. Sci. Pollut. Res. 2020, 28, 12831–12846. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from Red Seaweeds As Promoters of Growth and Elicitors of Defense Response in Plants. Front. Mar. Sci. 2016, 3, 81. [Google Scholar] [CrossRef]
- Kumar, G.; Sahoo, D. Effect of seaweed liquid extract on growth and yield of Triticum aestivum var. Pusa Gold. J. Appl. Phycol. 2011, 23, 251–255. [Google Scholar] [CrossRef]
- Paul, J. Influence of seaweed liquid fertilizer of Gracilaria dura (AG.) J. AG.(red seaweed) on Vigna radiata (l.) r. Wilczek., in Thoothukudi, Tamil Nadu, India. World J. Pharm. Res. 2014, 3, 968–978. [Google Scholar]
- Yamamoto, M.; Fukushima, M.; Kiso, E.; Kato, T.; Shibuya, M.; Horiya, S.; Nishida, A.; Otsuka, K.; Komai, T. Application of Iron Humates to Barren Ground in a Coastal Area forRestoring Seaweed Beds. J. Chem. Eng. Jpn. 2010, 43, 627–634. [Google Scholar] [CrossRef]
- El-Din, S.M. Utilization of seaweed extracts as bio-fertilizers to stimulate the growth of wheat seedlings. Egypt. J. Exp. Biol. 2015, 11, 31–39. [Google Scholar]
- Jayasinghe, P.S.; Pahalawattaarachchi, V.; Kkds, R. Fertilizer on plant growth of Capsicum annum. Discovery 2016, 52, 723–734. [Google Scholar]
- Patel, R.V.; Pandya, K.Y.; Brahmbhatt, N. Significance of Green And Brown Seaweed Liquid Fertilizer on Seed Germination of Solanum Melongena, Solanum Lycopersicum. Int. J. Recent Sci. Res. 2018, 2, 24065–24072. [Google Scholar] [CrossRef]
- Satish, L.; Rameshkumar, R.; Rathinapriya, P.; Pandian, S.; Rency, A.S.; Sunitha, T.; Ramesh, M. Effect of seaweed liquid extracts and plant growth regulators on in vitro mass propagation of brinjal (Solanum melongena L.) through hypocotyl and leaf disc explants. J. Appl. Phycol. 2015, 27, 993–1002. [Google Scholar] [CrossRef]
- Hidangmayum, A.; Sharma, R. Effect of different concentrations of commercial seaweed liquid extract of Ascophyllum nodosum as a plant bio stimulant on growth, yield and biochemical constituents of onion (Allium cepa L.). J. Pharmacogn. Phytochem. 2017, 6, 658–663. [Google Scholar]
- Abbas, M.; Anwar, J.; Zafar-ul-hye, M.; Khan, R.I. horticulturae Effect of Seaweed Extract on Productivity and Quality Attributes of Four Onion Cultivars. Horticulturae 2020, 6, 28. [Google Scholar] [CrossRef]
- Latique, S.; Chernane, H.; Mansori, M.; Kaoua, E. Seaweed Liquid Fertilizer Effect On Physiological And Biochemical Parameters Of Bean Plant (Phaesolus Vulgaris Variety Paulista) Under Hydroponic System. Eur. Sci. J. 2013, 9, 30. [Google Scholar]
- Castellanos-Barriga, L.G.; Santacruz-Ruvalcaba, F.; Hernández-Carmona, G.; Ramírez-Briones, E.; Hernández-Herrera, R.M. Effect of seaweed liquid extracts from Ulva lactuca on seedling growth of mung bean (Vigna radiata). J. Appl. Phycol. 2017, 29, 2479–2488. [Google Scholar] [CrossRef]
- Karthik, T.; Sarkar, G.; Babu, S.; Amalraj, L.D.; Jayasri, M.A. Preparation and evaluation of liquid fertilizer from Turbinaria ornata and Ulva reticulata. Biocatal. Agric. Biotechnol. 2020, 28, 101712. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Durgadevi, S.; Arulmozhi, P.; Rajalakshmi, S.; Gopalakrishnan, T.; Parameswari, N. Acta Ecologica Sinica Effect of seaweed liquid fertilizer on yield and quality of Capsicum. Acta Ecol. Sin. 2019, 39, 406–410. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Zañudo-Hernández, J.; Hernández-Carmona, G. Activity of seaweed extracts and polysaccharide-enriched extracts from Ulva lactuca and Padina gymnospora as growth promoters of tomato and mung bean plants. J. Appl. Phycol. 2016, 28, 2549–2560. [Google Scholar] [CrossRef]
- Sivasankari, S.; Venkatesalu, V.; Anantharaj, M.; Chandrasekaran, M. Effect of seaweed extracts on the growth and biochemical constituents of Vigna sinensis. Bioresour. Technol. 2006, 97, 1745–1751. [Google Scholar] [CrossRef]
- Patel, A.M.J.S. Seaweed extract: Biostimulator of plant defense and plant productivity. Int. J. Environ. Sci. Technol. 2020, 17, 553–558. [Google Scholar] [CrossRef]
- Arumugam, R.; Anantharaman, P. Antibacterial activity of Seaweeds of Pudumadam coast Antibacterial Activity of Some Selected Seaweeds from Pudumadam Coastal Regions. Glob. J. Pharmacol. 2016, 3, 50–52. [Google Scholar]
- Čmiková, N.; Galovičová, L.; Miškeje, M.; Borotová, P.; Kluz, M.; Kačániová, M. Determination of Antioxidant, Antimicrobial Activity, Heavy Metals and Elements Content of Seaweed Extracts. Plants 2022, 11, 1493. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Venkatesalu, V.; Raj, G.A. Antibacterial activity of Ulva fasciata against Multidrug Resistant Bacterial Strains. Int. Lett. Nat. Sci. 2014, 14, 40–51. [Google Scholar] [CrossRef]
- Nagayama, K.; Iwamura, Y.; Shibata, T.; Hirayama, I.; Nakamura, T. Bactericidal activity of phlorotannins from the brown alga Ecklonia kurome. J. Antimicrob. Chemother. 2002, 50, 889–893. [Google Scholar] [CrossRef]
- Aftabuddin, S.; Abdul, M.; Siddique, M.; Habib, A.; Akter, S.; Hossen, S.; Tanchangya, P.; Al, M.A. Effects of seaweeds extract on growth, survival, antibacterial activities, and immune responses of Penaeus monodon against Vibrio parahaemolyticus. Ital. J. Anim. Sci. 2021, 20, 243–255. [Google Scholar] [CrossRef]
- Fayzi, L.; Askarne, L.; Cherifi, O.; Boufous, E.H.; Cherifi, K. Comparative Antibacterial Activity of Some Selected Seaweed Extracts from Agadir Coastal Regions in Morocco. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 390–399. [Google Scholar] [CrossRef]
- Indira, K.; Balakrishnan, S.; Srinivasan, M.; Bragadeeswaran, S.; Balasubramanian, T. Evaluation of in vitro antimicrobial property of seaweed (Halimeda tuna) from Tuticorin coast, Tamil Nadu, Southeast coast of India. Afr. J. Biotechnol. 2013, 12, 284–289. [Google Scholar] [CrossRef]
- Ara, J.; Sultana, V.; Qasim, R.; Ehteshamul-haque, S.; Ahmad, V.U. Biological Activity of Spatoglossum asperum: A Brown Alga. Phytotherapy Res. 2005, 623, 618–623. [Google Scholar] [CrossRef] [PubMed]
- El Boukhari, M.E.M.; Barakate, M.; Drissi, B.; Bouhia, Y.; Lyamlouli, K. Seaweed Extract Biostimulants Differentially act in Mitigating Drought Stress on Faba Bean (Vicia faba L.). J. Plant Growth Regul. 2023, 42, 5642–5652. [Google Scholar] [CrossRef]
- Xu, C.; Leskovar, D.I. Effects of A. nodosum seaweed extracts on spinach growth, physiology and nutrition value under drought stress. Sci. Hortic. 2015, 183, 39–47. [Google Scholar] [CrossRef]
- Yarsi, G. Effects of mycorrhiza, seaweed and bionutrient applied to reduce the salt stress on nutrient content, plant growth, malondialdehyde (MDA) and proline in pepper. J. Elem. 2023, 28. [Google Scholar] [CrossRef]
- Latique, S.; Aymen, E.M.; Halima, C.; Chérif, H.; Mimoun, E.K. Alleviation of Salt Stress in Durum Wheat (Triticum durum L.) Seedlings Through the Application of Liquid Seaweed Extracts of Fucus spiralis. Commun. Soil Sci. Plant Anal. 2017, 48, 2582–2593. [Google Scholar] [CrossRef]
- Mireya, R.; Vanessa, C.; Andrea, P.; Ocampo-alvarez, H.; Santacruz-ruvalcaba, F. Seaweed Extract Improves Growth and Productivity of Tomato Plants under Salinity Stress. Agronomy 2022, 12, 2495. [Google Scholar] [CrossRef]
- Stirk, W.A.; Tarkowská, D.; Turečová, V.; Strnad, M.; van Staden, J. Abscisic acid, gibberellins and brassinosteroids in Kelpak®, a commercial seaweed extract made from Ecklonia maxima. J. Appl. Phycol. 2014, 26, 561–567. [Google Scholar] [CrossRef]
- Pereira, L.; Morrison, L.; Shukla, P.S.; Critchley, A.T. A concise review of the brown macroalga Ascophyllum nodosum (Linnaeus) Le Jolis. J. Appl. Phycol. 2020, 32, 3561–3584. [Google Scholar] [CrossRef]
- Ugarte, R.; Sharp, G. Management and production of the brown algae Ascophyllum nodosum in the Canadian maritimes. J. Appl. Phycol. 2012, 24, 409–416. [Google Scholar] [CrossRef]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a biostimulant derived from the brown seaweed Ascophyllum nodosum on ripening dynamics and fruit quality of grapevines. Sci. Hortic. 2018, 232, 97–106. [Google Scholar] [CrossRef]
- Szczepanek, M.; Wszelaczynska, E.; Poberezny, J. Effect of Seaweed Biostimulant Application in Sprng Wheat. AgroLife Sci. J. 2018, 7, 131–136. [Google Scholar]
- Ashour, M.; Al-Souti, A.S.; Hassan, S.M.; Ammar, G.A.G.; Goda, A.M.A.S.; El-Shenody, R.; Abomohra, A.E.F.; El-Haroun, E.; Elshobary, M.E. Commercial Seaweed Liquid Extract as Strawberry Biostimulants and Bioethanol Production. Life 2023, 13, 85. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulatory activities of Ascophyllum nodosum extract in tomato and sweet pepper crops in a tropical environment. PLoS ONE 2019, 14, e0216710. [Google Scholar] [CrossRef]
- Ahmed, M.; Ullah, H.; Himanshu, S.K.; García-Caparrós, P.; Tisarum, R.; Cha-um, S.; Datta, A. Ascophyllum nodosum seaweed extract and potassium alleviate drought damage in tomato by improving plant water relations, photosynthetic performance, and stomatal function. J. Appl. Phycol. 2024, 36, 2255–2268. [Google Scholar] [CrossRef]
- Belnap, J.; Lange, O.L. (Eds.) Biological Soil Crusts: Structure, Function, and Management; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 150. [Google Scholar]
- Ward, D.A.; Adhikari, S.; Struwig, M.; Skikne, S.; Fryday, A.; Smith, D.; Rajakaruna, N. Lichen morphospecies diversity and community composition across the Tswalu Kalahari Reserve, South Africa. S. Afr. J. Bot. 2024, 174, 978–987. [Google Scholar] [CrossRef]
- Aoussar, N.; Manzali, R.; Nattah, I.; Rhallabi, N.; Vasiljevic, P.; Bouksaim, M.; Douira, A.; Manojlović, N.; Mellouki, F. Chemical composition and antioxidant activity of two lichens species (Pseudevernia furfuracea L. and Evernia prunastri L.) collected from Morocco. J. Mater. Environ. Sci. 2017, 8, 1968–1976. [Google Scholar]
- Ajaj, A.; Ouazzani Touhami, A.; Benkirane, R. Contribution to the update catalogue of lichenized and lichenicolous fungi in Morocco. J. Anim. Plant Sci. 2013, 19, 2961–3025. [Google Scholar]
- Seaward, M.R.D.; Amrani, S. Checklist of lichens and lichenicolous fungi of Morocco. Herzogia 2023, 35, 564–612. [Google Scholar] [CrossRef]
- Badridze, G.; Chkhubianishvili, E.; Rapava, L.; Kikvidze, M.; Chigladze, L.; Tsiklauri, N.; Tsilosani, K.; Kupradze, I.; Chanishvili, S. Active metabolites of some lichens growing in Georgia. J. Biodivers. Environ. Sci. 2019, 15, 1–15. [Google Scholar]
- Akbulut, G.; Yildiz, A.; Kodu, D.; Code, R. An Overview to Lichens: The Nutrient Composition of Some Species Lichens, composed of fungi (microbiont) and algae (photobiont), are chemically and structurally very different from vascular plants. They represent a type of symbiosis, generally reg. Kafkas Üniversitesi Fen Bilim. Enstitüsü Derg. 2010, 3, 79–86. [Google Scholar]
- Ergün, N. Auxin (Indole-3-acetic acid), Gibberellic acid (GA 3), Abscisic Acid (ABA) and Cytokinin (Zeatin) Production by Some Species of Mosses and Lichens. Turk. J. Bot. 2002, 26, 13–18. [Google Scholar]
- Silva, M.d.L.C.d.; Iacomini, M.; Jablonski, E.; Gorin, P.A. Carbohydrate, glycopeptide and protein components of the lichen Sticta sp. and effect of storage. Phytochemistry 1993, 33, 547–552. [Google Scholar] [CrossRef]
- Yildiz, A.; Aksoy, A.; Akbulut, G.; Demirezen, D.; Islek, C.; Altuner, E.M.; Duman, F. Kayseri İlinde Pseudevernia furfuracea Türünde Klorofil Yikimi ile Aǧir Metal Miktari Arasindaki İlişki. Ekoloji 2011, 20, 82–88. [Google Scholar] [CrossRef]
- Kranner, I.; Cram, W.J.; Zorn, M.; Wornik, S.; Yoshimura, I.; Stabentheiner, E.; Pfeifhofer, H.W. Antioxidants and photoprotection in a lichen as compared with its isolated symbiotic partners. Proc. Natl. Acad. Sci. USA 2005, 102, 3141–3146. [Google Scholar] [CrossRef]
- Karakaş, V.E.; Öztürk, Ş.; Oran, S. Original Research Article Comparison of Photosynthetic Pigment Contents in Lichen Samples were Collected from Different Localities in Bursa. J. Biol. Environ. Sci. 2017, 11, 121–127. [Google Scholar]
- Pichler, G.; Stöggl, W.; Trippel, D.; Candotto Carniel, F.; Muggia, L.; Ametrano, C.G.; Çimen, T.; Holzinger, A.; Tretiach, M.; Kranner, I. Phytohormone release by three isolated lichen mycobionts and the effects of indole-3-acetic acid on their compatible photobionts. Symbiosis 2020, 82, 95–108. [Google Scholar] [CrossRef]
- Demirbas, A. Trace element concentrations in ashes from various types of Lichen biomass species. Energy Sources 2004, 26, 499–506. [Google Scholar] [CrossRef]
- Adamo, P.; Giordano, S.; Vingiani, S.; Cobianchi, R.C.; Violante, P. Trace element accumulation by moss and lichen exposed in bags in the city of Naples (Italy). Environ. Pollut. 2003, 122, 91–103. [Google Scholar] [CrossRef]
- Sorbo, S.; Aprile, G.; Strumia, S.; Castaldo Cobianchi, R.; Leone, A.; Basile, A. Trace element accumulation in Pseudevernia furfuracea (L.) Zopf exposed in Italy’s so called Triangle of Death. Sci. Total Environ. 2008, 407, 647–654. [Google Scholar] [CrossRef]
- Malaspina, P.; Giordani, P.; Modenesi, P.; Abelmoschi, M.L.; Magi, E.; Soggia, F. Bioaccumulation capacity of two chemical varieties of the lichen Pseudevernia furfuracea. Ecol. Indic. 2014, 45, 605–610. [Google Scholar] [CrossRef]
- Palmqvist, K. Carbon economy in lichens. New Phytol. 2000, 148, 11–36. [Google Scholar] [CrossRef]
- Clark, B.M.; Clair, L.L.S.; Mangelson, N.F.; Rees, L.B.; Grant, P.G.; Bench, G.S. Characterization of mycobiont adaptations in the foliose lichen Xanthoparmelia chlorochroa (Parmeliaceae). Am. J. Bot. 2001, 88, 1742–1749. [Google Scholar] [CrossRef]
- Pike, L.H. The Importance of Epiphytic Lichens in Mineral Cycling. Bryologist 1978, 81, 247–257. [Google Scholar] [CrossRef]
- Dahlman, L.; Persson, J.; Näsholm, T.; Palmqvist, K. Carbon and nitrogen distribution in the green algal lichens Hypogymnia physodes and Platismatia glauca in relation to nutrient supply. Planta 2003, 217, 41–48. [Google Scholar] [CrossRef]
- Johansson, O.; Palmqvist, K.; Olofsson, J. Nitrogen deposition drives lichen community changes through differential species responses. Glob. Chang. Biol. 2012, 18, 2626–2635. [Google Scholar] [CrossRef]
- Balarinová, K.; Barták, M.; Hazdrová, J.; Hájek, J.; Jílková, J. Changes in photosynthesis, pigment composition and glutathione contents in two Antarctic lichens during a light stress and recovery. Photosynthetica 2014, 52, 538–547. [Google Scholar] [CrossRef]
- Haugwitz, M.S.; Michelsen, A. Long-term addition of fertilizer, labile carbon, and fungicide alters the biomass of plant functional groups in a subarctic-alpine community. Plant Ecol. 2011, 212, 715–726. [Google Scholar] [CrossRef]
- Tunç, E.; Doğan, B. The Effect of Lichens on Soil Agregate Stability. Int. J. Energy Eng. Sci. 2022, 7, 40–51. [Google Scholar]
- Ghiloufi, W.; Yun, J.; Kim, J.; Lee, J.; Kang, H. The influences of lichens on soil physico-chemical properties, enzymes and microbes are species specific: Insights from South Mediterranean arid ecosystem. Appl. Soil Ecol. 2023, 181, 104656. [Google Scholar] [CrossRef]
- Ji, C.J.; Yang, Y.H.; Han, W.X.; He, Y.F.; Smith, J.; Smith, P. Climatic and Edaphic Controls on Soil pH in Alpine Grasslands on the Tibetan Plateau, China: A Quantitative Analysis. Pedosphere 2014, 24, 39–44. [Google Scholar] [CrossRef]
- Asta, J.; Orry, F.; Toutain, F.; Souchier, B.; Villemin, G. Micromorphological and ultrastructural investigations of the lichen—Soil interface. Soil Biol. Biochem. 2001, 33, 323–337. [Google Scholar] [CrossRef]
- Sauze, J.; Ogée, J.; Maron, P.A.; Crouzet, O.; Nowak, V.; Wohl, S.; Kaisermann, A.; Jones, S.P.; Wingate, L. The interaction of soil phototrophs and fungi with pH and their impact on soil CO2, CO18O and OCS exchange. Soil Biol. Biochem. 2017, 115, 371–382. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Liu, M.; Sui, X.; Hu, Y.; Feng, F. Microbial community structure and the relationship with soil carbon and nitrogen in an original Korean pine forest of Changbai Mountain, China. BMC Microbiol. 2019, 19, 218. [Google Scholar] [CrossRef]
- Stark, S.; Hyvärinen, M. Are phenolics leaching from the lichen Cladina stellaris sources of energy rather than allelopathic agents for soil microorganisms? Soil Biol. Biochem. 2003, 35, 1381–1385. [Google Scholar] [CrossRef]
- Sedia, E.G.; Ehrenfeld, J.G. Differential effects of lichens, mosses and grasses on respiration and nitrogen mineralization in soils of the New Jersey Pinelands. Oecologia 2005, 144, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Ohtonen, R.; Väre, H. Vegetation composition determines microbial activities in a boreal forest soil. Microb. Ecol. 1998, 36, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Mendili, M.; Essghaier, B.; Seaward, M.R.D.; Khadhri, A. In vitro evaluation of lysozyme activity and antimicrobial effect of extracts from four Tunisian lichens: Diploschistes ocellatus, Flavoparmelia caperata, Squamarina cartilaginea and Xanthoria parietina. Arch. Microbiol. 2021, 203, 1461–1469. [Google Scholar] [CrossRef]
- Galanty, A.; Paśko, P.; Podolak, I. Enantioselective activity of usnic acid: A comprehensive review and future perspectives. Phytochem. Rev. 2019, 18, 527–548. [Google Scholar] [CrossRef]
- Basile, A.; Rigano, D.; Loppi, S.; Di Santi, A.; Nebbioso, A.; Sorbo, S.; Conte, B.; Paoli, L.; De Ruberto, F.; Molinari, A.M.; et al. Antiproliferative, antibacterial and antifungal activity of the lichen Xanthoria parietina and its secondary metabolite parietin. Int. J. Mol. Sci. 2015, 16, 7861–7875. [Google Scholar] [CrossRef]
- Castillo-Monroy, A.P.; Bowker, M.A.; García-Palacios, P.; Maestre, F.T. Aspects of soil lichen biodiversity and aggregation interact to influence subsurface microbial function. Plant Soil 2014, 386, 303–316. [Google Scholar] [CrossRef]
- Medison, R.G.; Jiang, J.; Medison, M.B.; Tan, L.T.; Kayange, C.D.M.; Sun, Z.; Zhou, Y. Evaluating the potential of Bacillus licheniformis YZCUO202005 isolated from lichens in maize growth promotion and biocontrol. Heliyon 2023, 9, e20204. [Google Scholar] [CrossRef]






| Species | N | P | K | Na | Ca | Mg | Fe | Zn | Mn | Cu | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ulva Lactuca | 2900 | 93 | 209.0 | 351.6 | 180.6 | - | 34.4 | 1780 | 4.8 | 1.8 | [44,73,74] |
| Codium tomentosum | - | - | 429 | 1179 | 550 | - | 10,017 | - | - | - | [75] |
| Enteromorpha intestinalis | - | 155.2 | 865 | 2212 | 277 | - | 10,189 | - | - | - | [75] |
| Fucus vesiculosus | 2530 | 193.5 | 4322 | 5469 | 938 | 994 | 4.2 | 3.7 | 5.5 | <0.5 | [71,76] |
| Laminaria digitata | 900 | - | 11.5 | 3818 | 1005 | 659 | 3.2 | 1.7 | <0.5 | <0.5 | [71,76] |
| Undaria pinnatifida | - | 1070 | 8699 | 7064 | 931 | 118 | 7.5 | 1.7 | 3.2 | <0.5 | [77] |
| Sargassum naozhouense | - | 120 | 4170 | 3250 | 66.9 | - | 147 | 9.0 | 5.8 | 0.3 | [43] |
| Sargassum muticum | 4890 | - | 1370 | 740 | 960 | 1510 | 160 | 4.0 | 4.7 | - | [76,78,79] |
| Chondrus crispus | - | - | 3184 | 4270 | 420 | 732 | 3.9 | 7.1 | 1.3 | <0.5 | [71] |
| Porphyra tenera | - | - | 3500 | 3627 | 390 | 565 | 10.3 | 2.7 | 2.7 | <0.5 | [71] |
| Gracilaria edulis | 1500 | - | 282.5 | 423.3 | 223.3 | - | 65.2 | 1.7 | 3.9 | 1.7 | [74,80] |
| Species | Cytokinin (mg·100 g−1) | Gibberellin (mg·g−1) | Indole 3-Acetic Acid (mg·g−1) | Abscisic Acid (mg·g−1) | Ref. |
|---|---|---|---|---|---|
| Halimeda opuntiam | 2264.1 | 0.6561 | 0.6561 | 0.1133 | [12] |
| Ulva lactuca | - | 1.51× 10−6 | 4.93 × 10−5 | 1.78 × 10−5 | [83] |
| Cladophora glomerata | - | - | 0.02391 | - | [90] |
| Cladostephus verticillatum | - | - | - | 1.23 × 10−4 | [91] |
| Cladostephus sinuosa | - | - | 5.36 × 10−5 | - | [91] |
| Cladostephus barbata | - | 3.57 × 10−6 | - | - | [91] |
| Turbinaria triquetra | 2675.4 | 0.7705 | 0.01391 | 0.073 | [12] |
| Padina Durvillaei | - | 3.7 × 10−7 | 3.9 × 10−5 | 1.5 × 10−6 | [83] |
| Gracilaria corticate | 2400.4 | 24.004 | 0.1481 | 0.1986 | [12] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouala, O.; Essadki, Y.; Oudra, B.; El Khalloufi, F.; Martins, R. Bibliometric Analysis Towards Industrial-Scale Use of Marine Algae and Lichens as Soil Amendments and Plant Biofertilizers for Sustainable Agriculture. Phycology 2025, 5, 29. https://doi.org/10.3390/phycology5030029
Ouala O, Essadki Y, Oudra B, El Khalloufi F, Martins R. Bibliometric Analysis Towards Industrial-Scale Use of Marine Algae and Lichens as Soil Amendments and Plant Biofertilizers for Sustainable Agriculture. Phycology. 2025; 5(3):29. https://doi.org/10.3390/phycology5030029
Chicago/Turabian StyleOuala, Oumaima, Yasser Essadki, Brahim Oudra, Fatima El Khalloufi, and Rosario Martins. 2025. "Bibliometric Analysis Towards Industrial-Scale Use of Marine Algae and Lichens as Soil Amendments and Plant Biofertilizers for Sustainable Agriculture" Phycology 5, no. 3: 29. https://doi.org/10.3390/phycology5030029
APA StyleOuala, O., Essadki, Y., Oudra, B., El Khalloufi, F., & Martins, R. (2025). Bibliometric Analysis Towards Industrial-Scale Use of Marine Algae and Lichens as Soil Amendments and Plant Biofertilizers for Sustainable Agriculture. Phycology, 5(3), 29. https://doi.org/10.3390/phycology5030029