
Sargassum Biomass Movement and Proliferation in the Eastern Tropical Atlantic

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Methods
2.1. Location
2.2. Sargassum Detection and Quantification
2.2.1. Sargassum Detection Methods
2.2.2. Sargassum Biomass Relationship
2.2.3. Spatio-Temporal Variation of Sargassum Biomass
2.3. Assessing Co-Variance
3. Results and Discussion
3.1. Sargassum Detection
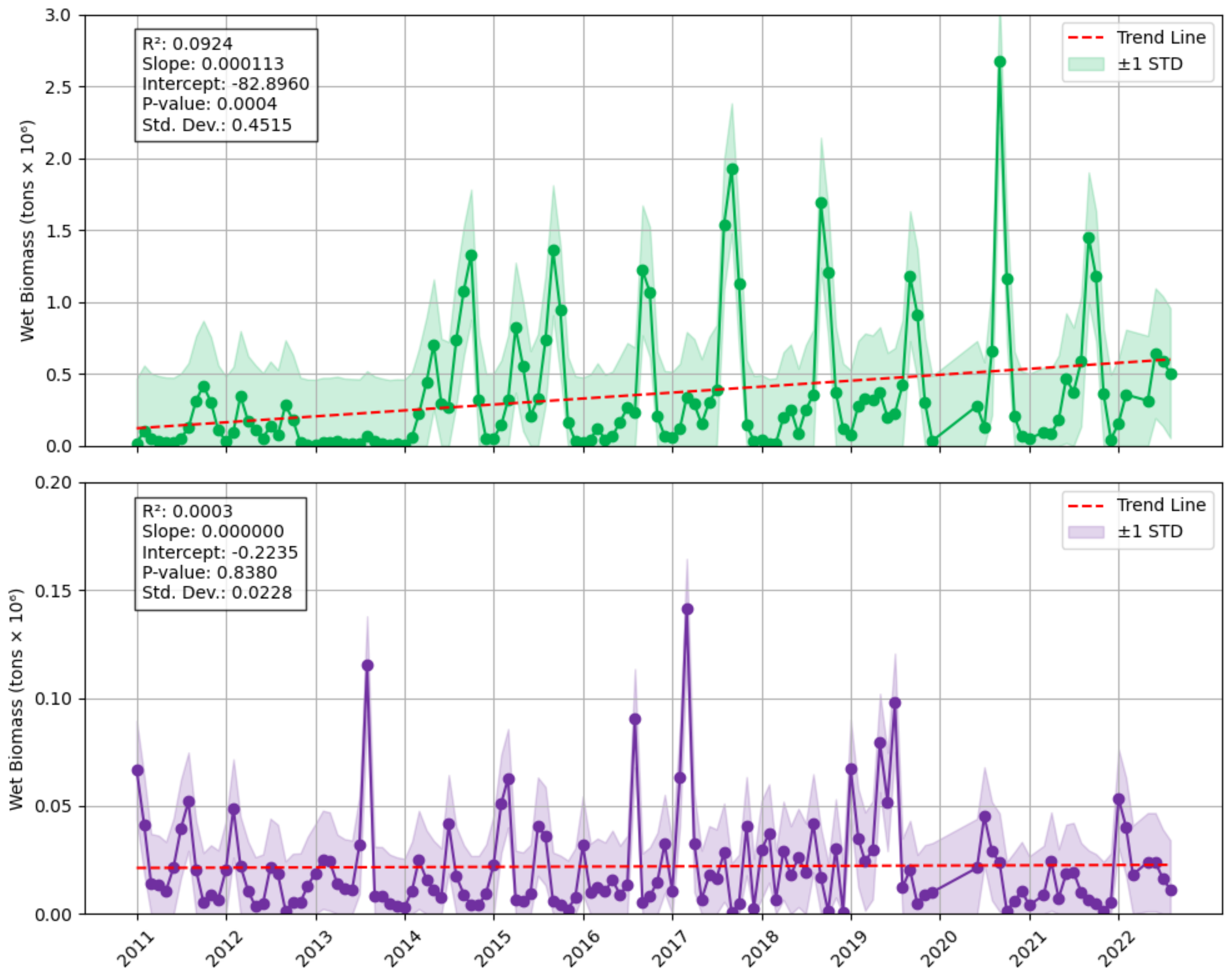
3.2. How Much Floating Sargassum Is There in the Eastern Tropical Atlantic, and What Is the Seasonal and Annual Variability?
3.2.1. Magnitude of Peak Biomass of Sargassum in the Eastern Tropical Atlantic
3.2.2. Seasonality of Sargassum in the Eastern Tropical Atlantic
3.2.3. Annual Sargassum Biomass Accumulation in the Eastern Tropical Atlantic
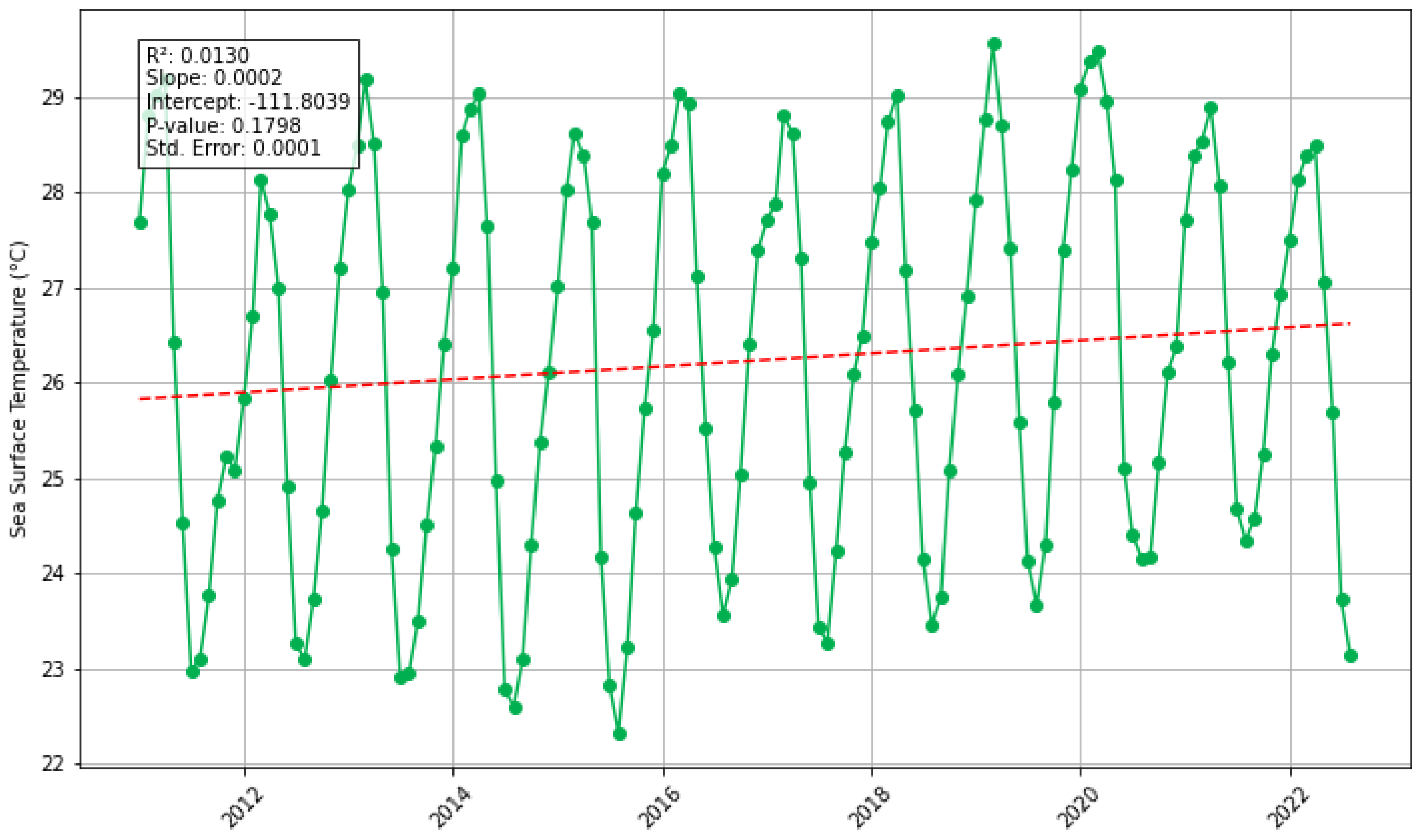
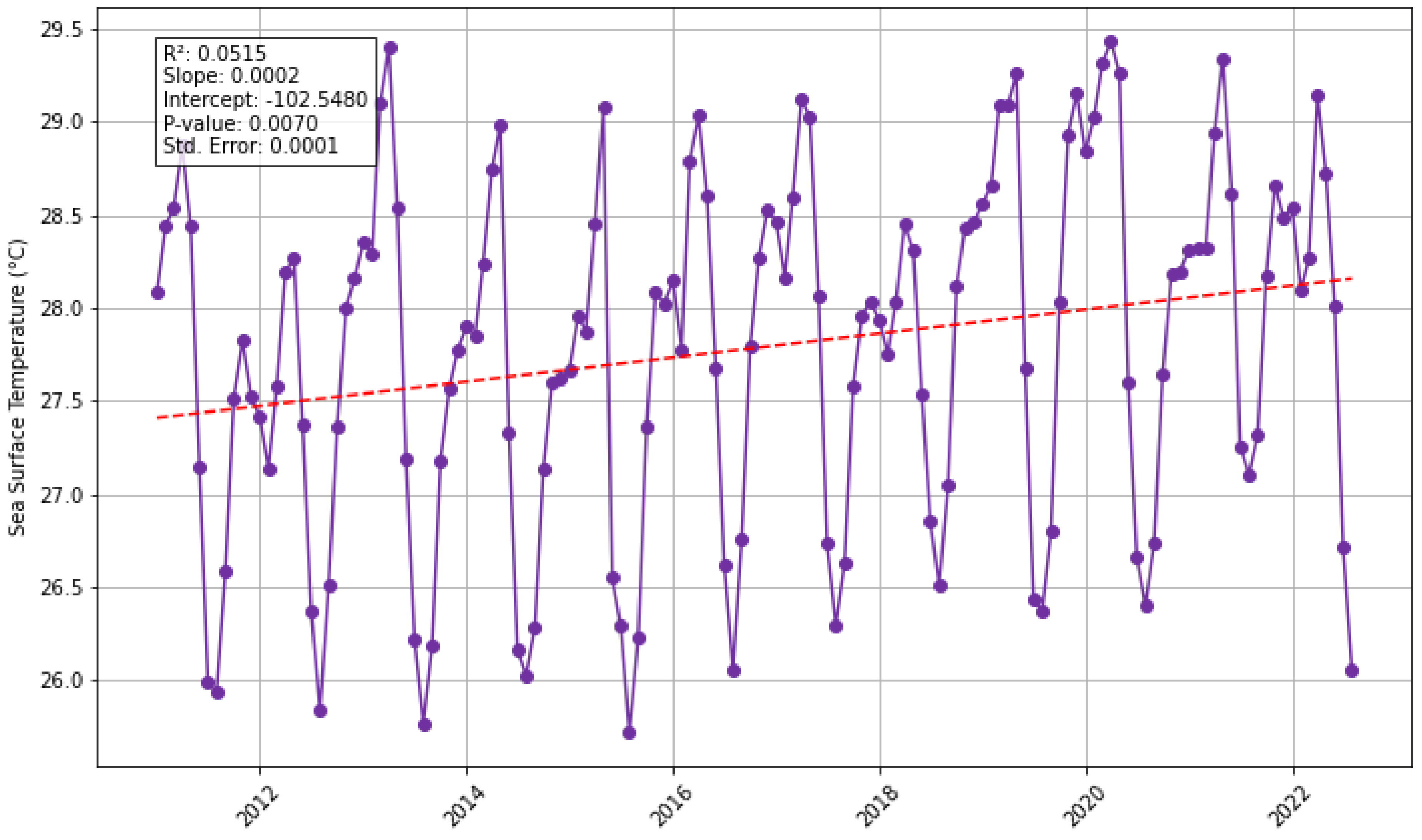
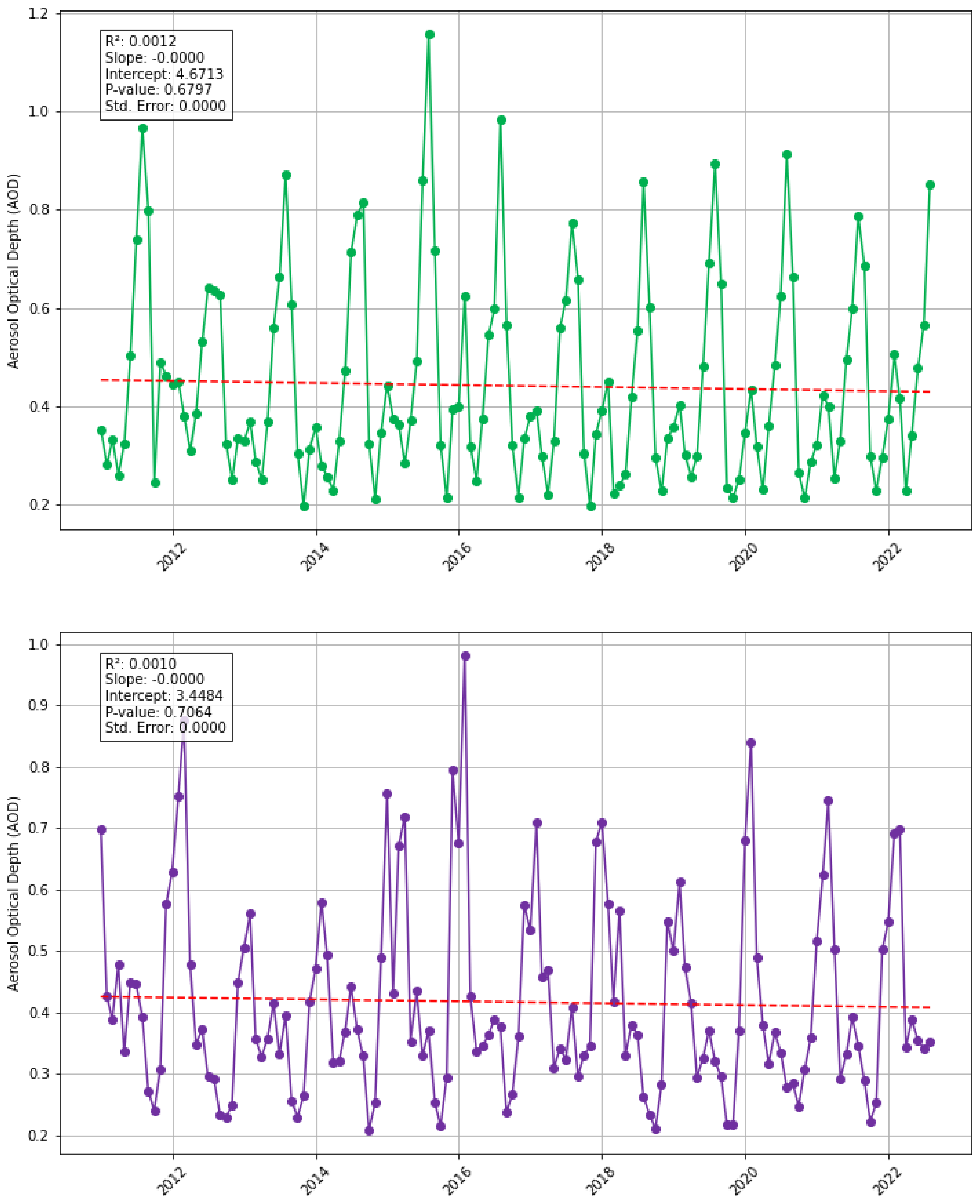
3.3. Does Sea-Surface Temperature, Atmospheric and Riverine/Coastal Nutrient Inputs Co-Vary with Sargassum in the Eastern Tropical Atlantic Region Between 2011 and 2022?
3.4. Are There Any Other Large-Scale Atmospheric, Oceanic, or Other Events That Co-Vary with Sargassum in the Eastern Tropical Atlantic Region Between 2011 and 2022?
3.5. Co-Variation Discussion Summary
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Hu, C.; Barnes, B.B.; Mitchum, G.; Lapointe, B.; Montoya, J.P. The great Atlantic Sargassum belt. Science 2019, 365, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Ody, A.; Thibaut, T.; Berline, L.; Changeux, T.; Andre, J.M.; Chevalier, C.; Blanfune, A.; Blanchot, J.; Ruitton, S.; Stiger-Pouvreau, V.; et al. From In Situ to satellite observations of pelagic Sargassum distribution and aggregation in the Tropical North Atlantic Ocean. PLoS ONE 2019, 14, e0222584. [Google Scholar] [CrossRef] [PubMed]
- Chávez, V.; Uribe-Martínez, A.; Cuevas, E.; Rodríguez-Martínez, R.E.; Van Tussenbroek, B.I.; Francisco, V.; Estévez, M.; Celis, L.B.; Monroy-Velázquez, L.V.; Leal-Bautista, R.; et al. Massive Influx of Pelagic Sargassum spp. on the Coasts of the Mexican Caribbean 2014–2020: Challenges and Opportunities. Water 2020, 12, 2908. [Google Scholar] [CrossRef]
- Van Tussenbroek, B.I.; Arana, H.A.H.; Rodríguez-Martínez, R.E.; Espinoza-Avalos, J.; Canizales-Flores, H.M.; González-Godoy, C.E.; Barba-Santos, M.G.; Vega-Zepeda, A.; Collado-Vides, L. Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar. Pollut. Bull. 2017, 122, 272–281. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, R.E.; Medina-Valmaseda, A.E.; Blanchon, P.; Monroy-Velázquez, L.V.; Almazán-Becerril, A.; Delgado-Pech, B.; Vásquez-Yeomans, L.; Francisco, V.; García-Rivas, M.C. Faunal mortality associated with massive beaching and decomposition of pelagic Sargassum. Mar. Pollut. Bull. 2019, 146, 201–205. [Google Scholar] [CrossRef]
- Ramlogan, N.R.; McConney, P.; Oxenford, H.A. Socio-Economic Impacts of Sargassum Influx Events on the Fishery Sector of Barbados; Centre for Resource Management and Environmental Studies (CERMES), University of the West Indies: Cave Hill, Barbados, 2017; p. 86. [Google Scholar]
- Solarin, B.B.; Bolaji, D.A.; Fakayode, O.S.; Akinnigbagbe, R.O. Impacts of an invasive seaweed Sargassum hystrix var. fluitans (Børgesen 1914) on the fisheries and other economic implications for the Nigerian coastal waters. IOSR J. Agric. Vet. Sci. 2014, 7, 1–6. [Google Scholar]
- Resiere, D.; Valentino, R.; Nevière, R.; Banydeen, R.; Gueye, P.; Florentin, J.; Cabié, A.; Lebrun, T.; Mégarbane, B.; Guerrier, G.; et al. Sargassum seaweed on Caribbean islands: An international public health concern. Lancet 2018, 392, 2691. [Google Scholar] [CrossRef]
- Atiglo, D.Y.; Jayson-Quashigah, P.N.; Sowah, W.; Tompkins, E.L.; Addo, K.A. Misperception of drivers of risk alters willingness to adapt in the case of sargassum influxes in West Africa. Glob. Environ. Change 2024, 84, 102779. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-value products from macroalgae: The potential uses of the invasive brown seaweed, Sargassum muticum. Rev. Environ. Sci. Bio/Technol. 2016, 15, 67–88. [Google Scholar] [CrossRef]
- Thompson, T.M.; Young, B.R.; Baroutian, S. Pelagic Sargassum for energy and fertiliser production in the Caribbean: A case study on Barbados. Renew. Sustain. Energy Rev. 2020, 118, 109564. [Google Scholar] [CrossRef]
- Gray, L.A.; Bisonó León, A.G.; Rojas, F.E.; Veroneau, S.S.; Slocum, A.H. Caribbean-wide, negative emissions solution to Sargassum spp. low-cost collection device and sustainable disposal method. Phycology 2021, 1, 49–75. [Google Scholar] [CrossRef]
- Oxenford, H.A.; Cox, S.A.; van Tussenbroek, B.I.; Desrochers, A. Challenges of turning the sargassum crisis into gold: Current constraints and implications for the Caribbean. Phycology 2021, 1, 27–48. [Google Scholar] [CrossRef]
- Wang, M.; Hu, C. Predicting Sargassum blooms in the Caribbean Sea from MODIS observations. Geophys. Res. Lett. 2017, 44, 3265–3273. [Google Scholar] [CrossRef]
- Gower, J.F.; King, S.A. Distribution of floating Sargassum in the Gulf of Mexico and the Atlantic Ocean mapped using MERIS. Int. J. Remote Sens. 2011, 32, 1917–1929. [Google Scholar] [CrossRef]
- Marsh, R.; Addo, K.A.; Jayson-Quashigah, P.-N.; Oxenford, H.A.; Maxam, A.; Anderson, R.; Skliris, N.; Dash, J.; Tompkins, E.L. Seasonal predictions of holopelagic sargassum across the tropical atlantic accounting for uncertainty in drivers and processes: The sartrac ensemble forecast system. Front. Mar. Sci. 2021, 8, 1417. [Google Scholar] [CrossRef]
- Maréchal, J.P.; Hellio, C.; Hu, C. A simple, fast, and reliable method to predict Sargassum washing ashore in the Lesser Antilles. Remote. Sens. Appl. Soc. Environ. 2017, 5, 54–63. [Google Scholar] [CrossRef]
- Marsh, R.; Oxenford, H.A.; Cox, S.A.L.; Johnson, D.R.; Bellamy, J. Forecasting seasonal sargassum events across the tropical Atlantic: Overview and challenges. Front. Mar. Sci. 2022, 9, 914501. [Google Scholar] [CrossRef]
- Fidai, Y.A.; Dash, J.; Tompkins, E.L.; Tonon, T. A systematic review of floating and beach landing records of Sargassum beyond the Sargasso Sea. Environ. Res. Commun. 2020, 2, 122001. [Google Scholar] [CrossRef]
- Addico, G.N.D.; deGraft-Johnson, K.A.A. Preliminary investigation into the chemical composition of the invasive brown seaweed Sargassum along the West Coast of Ghana. Afr. J. Biotechnol. 2016, 15, 2184–2191. [Google Scholar]
- Oyesiku, O.O.; Egunyomi, A. Identification and chemical studies of pelagic masses of Sargassum natans (Linnaeus) Gaillon and S. fluitans (Borgessen) Borgesen (brown algae), found offshore in Ondo State, Nigeria. Afr. J. Biotechnol. 2014, 13, 1188–1193. [Google Scholar]
- Adet, L.; Ragatoa, S.D.; Sanou, C.L.; Meminvegni, G.; Sidibe, M. Mapping Sargassum fluorescence distribution on Nigerian sea. In Unlocking Sub-Sahara African Potentials for Sustainable Development in the 21st Century; Nassarawa State University: Keffi, Nigeria, 2018. [Google Scholar]
- Gower, J.; King, S. The distribution of pelagic Sargassum observed with OLCI. Int. J. Remote Sens. 2020, 41, 5669–5679. [Google Scholar] [CrossRef]
- Brooks, M.T.; Coles, V.J.; Hood, R.R.; Gower, J.F. Factors controlling the seasonal distribution of pelagic Sargassum. Mar. Ecol. Prog. Ser. 2018, 599, 1–18. [Google Scholar] [CrossRef]
- Franks, J.S.; Johnson, D.R.; Ko, D.S. Pelagic Sargassum in the tropical North Atlantic. Gulf Caribb. Res. 2016, 27, SC6–SC11. [Google Scholar] [CrossRef]
- Oviatt, C.A.; Huizenga, K.; Rogers, C.S.; Miller, W.J. What nutrient sources support anomalous growth and the recent sargassum mass stranding on Caribbean beaches? A review. Mar. Pollut. Bull. 2019, 145, 517–525. [Google Scholar] [CrossRef]
- Skliris, N.; Marsh, R.; Appeaning Addo, K.; Oxenford, H. Physical drivers of pelagic sargassum bloom interannual variability in the Central West Atlantic over 2010–2020. Ocean. Dyn. 2022, 72, 383–404. [Google Scholar] [CrossRef]
- Djakouré, S.; Araujo, M.; Hounsou-Gbo, A.; Noriega, C.; Bourlès, B. On the potential causes of the recent Pelagic Sargassum blooms events in the tropical North Atlantic Ocean. Biogeosciences Discuss. 2017, 2017, 1–20. [Google Scholar]
- Mendez-Tejeda, R.; Rosado Jiménez, G.A. Influence of climatic factors on Sargassum arrivals to the coasts of the Dominican Republic. J. Oceanogr. Mar. Sci. 2019, 10, 22–32. [Google Scholar] [CrossRef]
- UNEP. Sargassum White Paper: Turning the Crisis into an Opportunity. United Nations Environment Programme (UNEP), & Caribbean Environment Programme. 2021. Available online: https://wedocs.unep.org/20.500.11822/36244 (accessed on 1 July 2023).
- Johns, E.M.; Lumpkin, R.; Putman, N.F.; Smith, R.H.; Muller-Karger, F.E.; Rueda-Roa, D.T.; Hu, C.; Wang, M.; Brooks, M.T.; Gramer, L.J.; et al. The establishment of a pelagic Sargassum population in the tropical Atlantic: Biological consequences of a basin-scale long distance dispersal event. Prog. Oceanogr. 2020, 182, 102269. [Google Scholar] [CrossRef]
- Marsh, R.; Skliris, N.; Tompkins, E.L.; Dash, J.; Dominguez Almela, V.; Tonon, T.; Oxenford, H.A.; Webber, M. Climate-sargassum interactions across scales in the tropical Atlantic. PLOS Clim. 2023, 2, e0000253. [Google Scholar] [CrossRef]
- Wang, M.; Hu, C. Mapping and quantifying Sargassum distribution and coverage in the Central West Atlantic using MODIS observations. Remote Sens. Environ. 2016, 183, 350–367. [Google Scholar] [CrossRef]
- Wang, M.; Hu, C.; Cannizzaro, J.; English, D.; Han, X.; Naar, D.; Lapointe, B.; Brewton, R.; Hernandez, F. Remote sensing of Sargassum biomass, nutrients, and pigments. Geophys. Res. Lett. 2018, 45, 12–359. [Google Scholar] [CrossRef]
- Santer, B.D.; Thorne, P.; Haimberger, L.; Taylor, K.E.; Wigley, T.M.; Lanzante, J.; Solomon, S.; Free, M.; Gleckler, P.J.; Jones, P.D.; et al. Consistency of modelled and observed temperature trends in the tropical troposphere. Int. J. Climatol. 2008, 28, 1703–1722. [Google Scholar] [CrossRef]
- Jonsson, B. Brorfred/Ipcc_Oc_Trends: Code Used for the IPPC AR6 Report, version 1.0.0; Zenodo: Genève, Switzerland, 2021. [Google Scholar] [CrossRef]
- Barnston, A.G.; Livezey, R.E. Classification, seasonality and persistence of low-frequency atmospheric circulation patterns. Mon. Weather. Rev. 1987, 115, 1083–1126. [Google Scholar] [CrossRef]
- Behringer, D.W.; Ji, M.; Leetmaa, A. An improved coupled model for ENSO prediction and implications for ocean initialization. Part I: The ocean data assimilation system. Mon. Weather. Rev. 1998, 126, 1013–1021. [Google Scholar] [CrossRef]
- Srokosz, M.A.; Bryden, H.L. Observing the Atlantic Meridional Overturning Circulation yields a decade of inevitable surprises. Science 2015, 348, 1255575. [Google Scholar] [CrossRef]
- Ogbuka, J.C.; Nwanmuoh, E.E.; Ogbo, A.I.; Achoru, F.E. Offshore oil spill response base and management of deepwater/offshore oil resources in the Nigerian marine waters: A review. Int. J. Environ. Impacts 2022, 5, 65–81. [Google Scholar] [CrossRef]
- Fosu, A.K. Oil and Ghana’s Economy. In The Economy of Ghana Sixty Years after Independence; Aryeetey, E., Kanbur, R., Eds.; Oxford Academic: Oxford, UK, 2017. [Google Scholar] [CrossRef]
- Stout, S.A.; Rouhani, S.; Liu, B.; Oehrig, J.; Ricker, R.W.; Baker, G.; Lewis, C. Assessing the footprint and volume of oil deposited in deep-sea sediments following the Deepwater Horizon oil spill. Mar. Pollut. Bull. 2017, 114, 327–342. [Google Scholar] [CrossRef]
- Platnick, S.; Hubanks, P.; Meyer, K.; King, M.D. MODIS Atmosphere L3 Monthly Product (08_L3); NASA MODIS Adaptive Processing System, Goddard Space Flight Center: Washington, DC, USA, 2015. [Google Scholar] [CrossRef]
- Magaña-Gallegos, E.; Villegas-Muñoz, E.; Salas-Acosta, E.R.; Barba-Santos, M.G.; Silva, R.; van Tussenbroek, B.I. The Effect of Temperature on the Growth of Holopelagic Sargassum Species. Phycology 2023, 3, 138–146. [Google Scholar] [CrossRef]
- Leemans, L.; Magaña-Gallegos, E.; van Katwijk, M.M.; Lamers, L.P.; Smolders, A.J.; Bouma, T.J.; Christianen, M.J.; van Tussenbroek, B.I. Iron co-limitation of Sargassum fluitans. Aquat. Bot. 2025, 196, 103807. [Google Scholar] [CrossRef]
- Xu-Yang, Y.; Dessert, C.; Losno, R. Atmospheric Deposition over the Caribbean region: Sea salt and Saharan dust are sources of essential elements on the island of Guadeloupe. J. Geophys. Res. Atmos. 2022, 127, e2022JD037175. [Google Scholar] [CrossRef]
- Xian, P.; Klotzbach, P.J.; Dunion, J.P.; Janiga, M.A.; Reid, J.S.; Colarco, P.R.; Kipling, Z. Revisiting the relationship between Atlantic dust and tropical cyclone activity using aerosol optical depth reanalyses: 2003–2018. Atmos. Chem. Phys. 2020, 20, 15357–15378. [Google Scholar] [CrossRef]
- Kazapoe, R.W.; Amuah, E.E.Y.; Abdiwali, S.A.; Dankwa, P.; Nang, D.B.; Kazapoe, J.P.; Kpiebaya, P. Relationship between small-scale gold mining activities and water use in Ghana: A review of policy documents aimed at protecting water bodies in mining communities. Environ. Chall. 2023, 12, 100727. [Google Scholar] [CrossRef]
- Faseyi, C.A.; Miyittah, M.K.; Sowunmi, A.A.; Yafetto, L. Water quality and health risk assessments of illegal gold mining-impacted estuaries in Ghana. Mar. Pollut. Bull. 2022, 185, 114277. [Google Scholar] [CrossRef]
- Miyittah, M.K.; Tulashie, S.K.; Tsyawo, F.W.; Sarfo, J.K.; Darko, A.A. Assessment of surface water quality status of the Aby Lagoon System in the Western Region of Ghana. Heliyon 2020, 6, e04466. [Google Scholar] [CrossRef]
- Davis, D.; Simister, R.; Campbell, S.; Marston, M.; Bose, S.; McQueen-Mason, S.J.; Gomez, L.D.; Gallimore, W.A.; Tonon, T. Biomass composition of the golden tide pelagic seaweeds Sargassum fluitans and S. natans (morphotypes I and VIII) to inform valorisation pathways. Sci. Total. Environ. 2021, 762, 143134. [Google Scholar] [CrossRef]
- Tonon, T.; Machado, C.B.; Webber, M.; Webber, D.; Smith, J.; Pilsbury, A.; Cicéron, F.; Herrera-Rodriguez, L.; Jimenez, E.M.; Suarez, J.V.; et al. Biochemical and elemental composition of pelagic Sargassum biomass harvested across the caribbean. Phycology 2022, 2, 204–215. [Google Scholar] [CrossRef]
- Nassar, C.A.; Lavrado, H.P.; Yoneshigue-Valentin, Y.O.C.I.E. Effects of iron-ore particles on propagule release, growth and photosynthetic performance of Sargassum vulgare C. Agardh (Phaeophyta, Fucales). Braz. J. Bot. 2002, 25, 459–468. [Google Scholar] [CrossRef]
- Gouvêa, L.P.; Assis, J.; Gurgel, C.F.; Serrão, E.A.; Silveira, T.C.; Santos, R.; Duarte, C.M.; Peres, L.M.; Carvalho, V.F.; Batista, M.; et al. Golden carbon of Sargassum forests revealed as an opportunity for climate change mitigation. Sci. Total Environ. 2020, 729, 138745. [Google Scholar] [CrossRef]
- Heyes, A.; Powers, L.; Leonzo, K.; Alvarez, J.P.; Gonsior, M. Mercury and Carbon Linkages in Macroalgae Sargassum Mats: Coastal Stranding Leads to Degradation, Shifts in Dissolved Organic Matter Composition and Mercury Methylation. In Proceedings of the AGU Fall Meeting Abstracts, Washington, DC, USA, 10–14 December 2018; Volume 2018, p. B51K-2101. [Google Scholar]
- Hanisak, M.D.; Samuel, M.A. Growth rates in culture of several species of Sargassum from Florida, USA. In Twelfth International Seaweed Symposium, Proceedings of the Twelfth International Seaweed Symposium, Sao Paulo, Brazil, 27 July–1 August 1986; Springer: Dordrecht, The Netherlands, 1987; pp. 399–404. [Google Scholar]
- Machado, C.B.; Maddix, G.M.; Francis, P.; Thomas, S.L.; Burton, J.A.; Langer, S.; Larson, T.R.; Marsh, R.; Webber, M.; Tonon, T. Pelagic Sargassum events in Jamaica: Provenance, morphotype abundance, and influence of sample processing on biochemical composition of the biomass. Sci. Total Environ. 2022, 817, 152761. [Google Scholar] [CrossRef]
- Steen, H. Effects of reduced salinity on reproduction and germling development in Sargassum muticum (Phaeophyceae, Fucales). Eur. J. Phycol. 2004, 39, 293–299. [Google Scholar] [CrossRef]
- Langmann, B.; Zakšek, K.; Hort, M.; Duggen, S. Volcanic ash as fertiliser for the surface ocean. Atmos. Chem. Phys. 2010, 10, 3891–3899. [Google Scholar] [CrossRef]
- Natsuike, M.; Endo, Y.; Ito, H.; Miyamoto, M.; Yoshimura, C.; Fujii, M. Iron uptake kinetics by coastal micro- and macro-algae in relation to riverine and coastal organic matter. Estuar. Coast. Shelf Sci. 2020, 235, 106580. [Google Scholar] [CrossRef]
- Wu, J.; Boyle, E. Iron in the Sargasso Sea: Implications for the processes controlling dissolved Fe distribution in the ocean. Glob. Biogeochem. Cycles 2002, 16, 33-1–33-8. [Google Scholar] [CrossRef]
- Achterberg, E.P.; Steigenberger, S.; Marsay, C.M.; LeMoigne, F.A.; Painter, S.C.; Baker, A.R.; Connelly, D.P.; Moore, C.M.; Tagliabue, A.; Tanhua, T. Iron biogeochemistry in the high latitude North Atlantic Ocean. Sci. Rep. 2018, 8, 1283. [Google Scholar] [CrossRef]
- Sanchez-Rubio, G.; Perry, H.; Franks, J.S.; Johnson, D.R. Occurrence of pelagic Sargassum in waters of the US Gulf of Mexico in response to weather-related hydrographic regimes associated with decadal and interannual variability in global climate. Fish. Bull. 2018, 116, 93–106. [Google Scholar]
- Joly, M.; Voldoire, A. Role of the Gulf of Guinea in the inter-annual variability of the West African monsoon: What do we learn from CMIP3 coupled simulations? Int. J. Clim. 2010, 30, 1843–1856. [Google Scholar] [CrossRef]
- Almeda, R.; Cosgrove, S.; Buskey, E.J. Oil spills and dispersants can cause the initiation of potentially harmful dinoflagellate blooms (“Red Tides”). Environ. Sci. Technol. 2018, 52, 5718–5724. [Google Scholar] [CrossRef]
- Powers, S.P.; Hernandez, F.J.; Condon, R.H.; Drymon, J.M.; Free, C.M. Novel pathways for injury from offshore oil spills: Direct, sublethal and indirect effects of the Deepwater Horizon oil spill on pelagic Sargassum communities. PLoS ONE 2013, 8, e74802. [Google Scholar] [CrossRef]
- Najoui, Z.; Amoussou, N.; Riazanoff, S.; Aurela, G.; Frappartd, F. Oil slicks in the Gulf of Guinea–10 years of Envisat ASAR observations. Earth Syst. Sci. Data Discuss. 2022, 14, 4569–4588. [Google Scholar] [CrossRef]
- Hu, C. A novel ocean color index to detect floating algae in the global oceans. Remote Sens. Environ. 2009, 113, 2118–2129. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Chlus, A.; Russell, B. Hyperspectral discrimination of floating mats of seagrass wrack and the macroalgae Sargassum in coastal waters of Greater Florida Bay using airborne remote sensing. Remote Sens. Environ. 2015, 167, 247–258. [Google Scholar] [CrossRef]
- Hu, C.; Feng, L.; Hardy, R.F.; Hochberg, E.J. Spectral and spatial requirements of remote measurements of pelagic Sargassum macroalgae. Remote Sens. Environ. 2015, 167, 229–246. [Google Scholar] [CrossRef]
- Gower, J.; Hu, C.; Borstad, G.; King, S. Ocean color satellites show extensive lines of floating Sargassum in the Gulf of Mexico. IEEE Trans. Geosci. Remote Sens. 2006, 44, 3619–3625. [Google Scholar] [CrossRef]
- Gower, J.; King, S. Satellite images show the movement of floating Sargassum in the Gulf of Mexico and Atlantic Ocean. Nat. Preced. 2008, 1–13. [Google Scholar] [CrossRef]
- Gower, J.; Young, E.; King, S. Satellite images suggest a new Sargassum source region in 2011. Remote Sens. Lett. 2013, 4, 764–777. [Google Scholar] [CrossRef]
- Hu, C.; Hardy, R.; Ruder, E.; Geggel, A.; Feng, L.; Powers, S.; Hernandez, F.; Graettinger, G.; Bodnar, J.; McDonald, T. Sargassum coverage in the northeastern Gulf of Mexico during 2010 from Landsat and airborne observations: Implications for the Deepwater Horizon oil spill impact assessment. Mar. Pollut. Bull. 2016, 107, 15–21. [Google Scholar] [CrossRef]
- McCarthy, S.; Gallegos, S.C.; Armstrong, D. Automated Sargassum Detection for Landsat Imagery. In Proceedings of the American Geophysical Union, The Ocean Sciences Meeting 2016, New Orleans, LA, USA, 21–26 February 2016; p. OD34A-2501. [Google Scholar]
- Ped, J.; Scaduto, E.; Accorsi, E.; Torres-Pérez, J. Caribbean Oceans: Utilizing NASA Earth Observations to Detect, Monitor, and Respond to Unprecedented Levels of Sargassum in the Caribbean Sea; NTRS—NASA Technical Reports, report no. NF1676L-24244; NASA: Washington, DC, USA, 2016. [Google Scholar]
- Wang, M.; Hu, C. Automatic extraction of Sargassum features from sentinel-2 msi images. IEEE Trans. Geosci. Remote Sens. 2020, 59, 2579–2597. [Google Scholar] [CrossRef]
- Wang, M.; Hu, C. On the continuity of quantifying floating algae of the Central West Atlantic between MODIS and VIIRS. Int. J. Remote Sens. 2018, 39, 3852–3869. [Google Scholar] [CrossRef]
- Minghelli, A.; Chevalier, C.; Descloitres, J.; Berline, L.; Blanc, P.; Chami, M. Synergy between Low Earth Orbit (LEO)—MODIS and Geostationary Earth Orbit (GEO)—GOES Sensors for Sargassum Monitoring in the Atlantic Ocean. Remote Sens. 2021, 13, 1444. [Google Scholar] [CrossRef]
- Sutton, M.; Stum, J.; Hajduch, G.; Dufau, C.; Maréchal, J.P.; Lucas, M. Monitoring a new type of pollution in the Atlantic Ocean: The sargassum algae. In Proceedings of the OCEANS 2019-Marseille 2019, Marseille, France, 17–20 June 2019; IEEE: Marseille, France, 2019; pp. 1–4. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Description/Data Type and Processing | Data Source |
|---|---|---|
| Ocean | ||
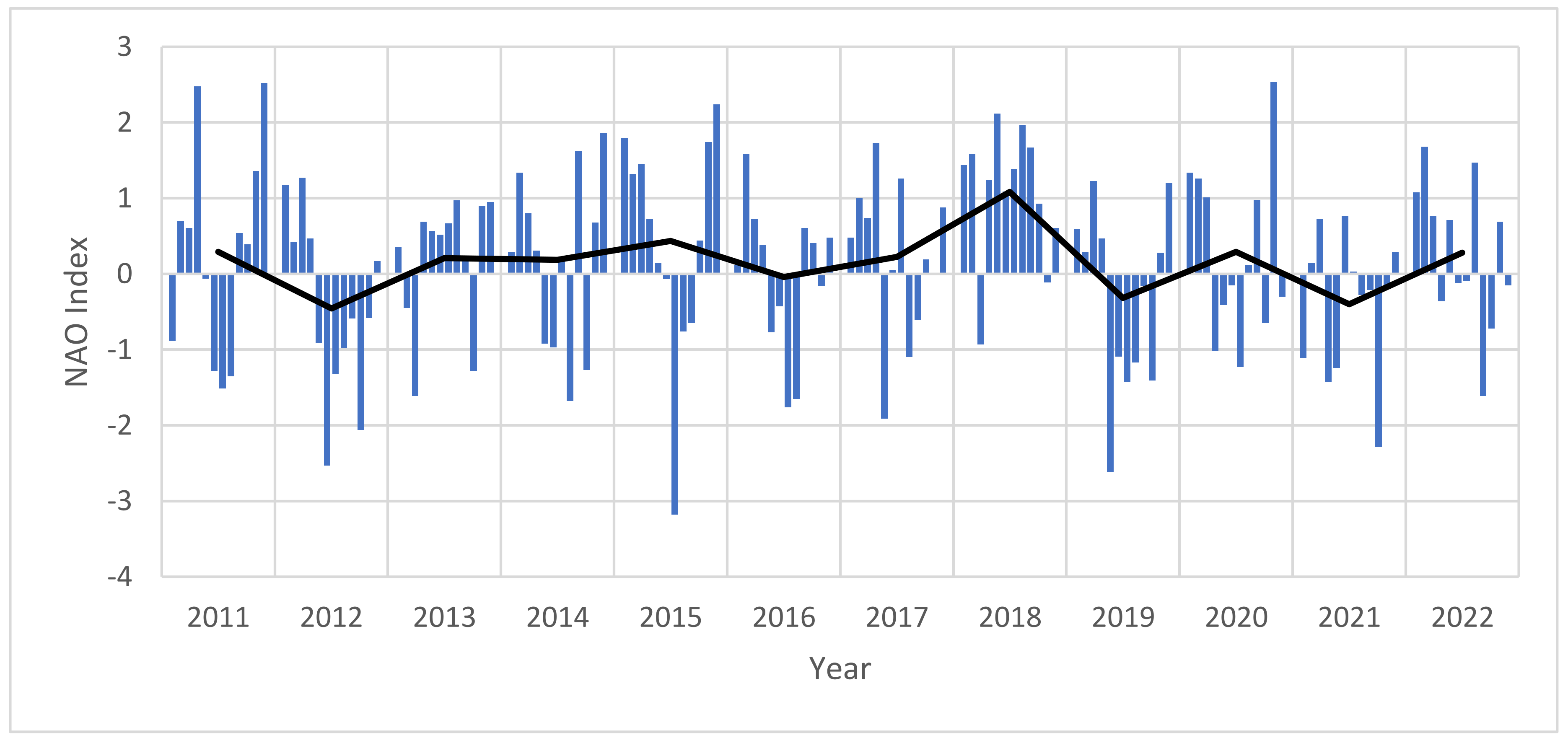
| North Atlantic Oscillation (NAO) | Monthly NAO index (calculation based on the Rotated Principle Component Analysis by Barnston and Livezey 1987 [37]). Downloaded in ASCII format and processed in MS Excel. | National Oceanic and Atmospheric Administration (NOAA) URL: https://www.cpc.ncep.noaa.gov/products/precip/CWlink/pna/norm.nao.monthly.b5001.current.ascii.table (accessed on 1 July 2023) |
| Sea surface temperature | Daytime Sea Surface Temperature (measured in Celsius) derived from MODIS Aqua Satellite data. Downloaded in csv format and processed in MS Excel and Python. | Giovanni NASA Earth Data, Aqua MODIS Global Mapped 11 µm Daytime Sea Surface Temperature (Data Product: MODISA_L3 m_SST_ Monthly_9 km_vR2019.0) URL: https://giovanni.gsfc.nasa.gov/ (accessed on 1 July 2023) |
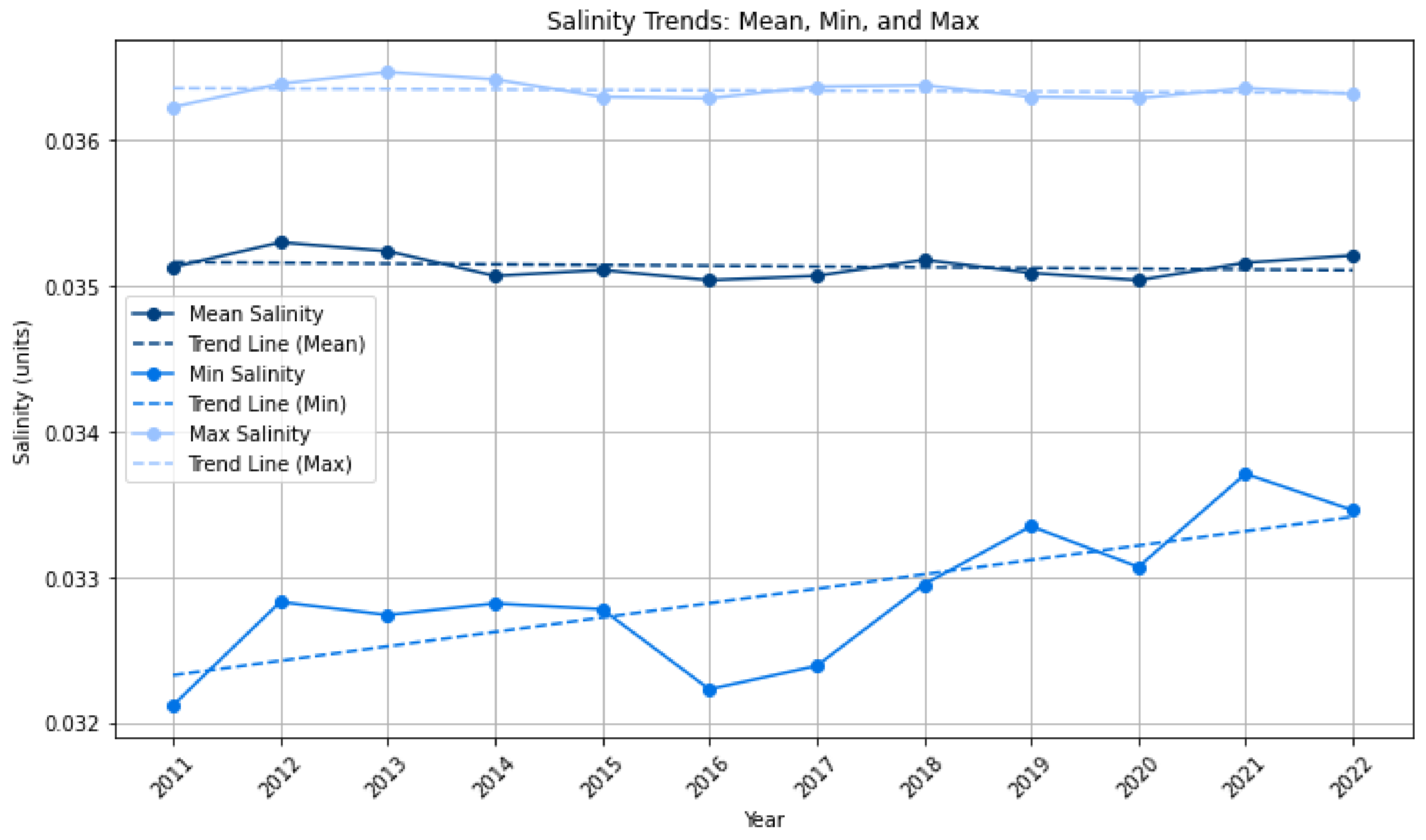
| Sea salinity | Salinity, measured as monthly mean, measured in kilograms, for sea surface up to 5.0 mdeep. Downloaded in netCDF format and processed in ArcGIS Pro and Python. | Behringer and Leetmaa (1998) [38] from NCEP Global Ocean Data Assimilation System (GODAS) data provided by the NOAA Physical Sciences Laboratory, Boulder, Colorado, USA, URL: https://psl.noaa.gov (accessed on 1 July 2023) |
| Atlantic Meridional Overturning Circulation (AMOC) | The AMOC system of ocean currents circulates water within the Atlantic Ocean, moving warm water and nutrients across the globe; it is important for heat transport. Here, trends and anomalies trends are noted and discussed. Information/data from empirical publications. | Srokosz and Bryden (2015) [39] |
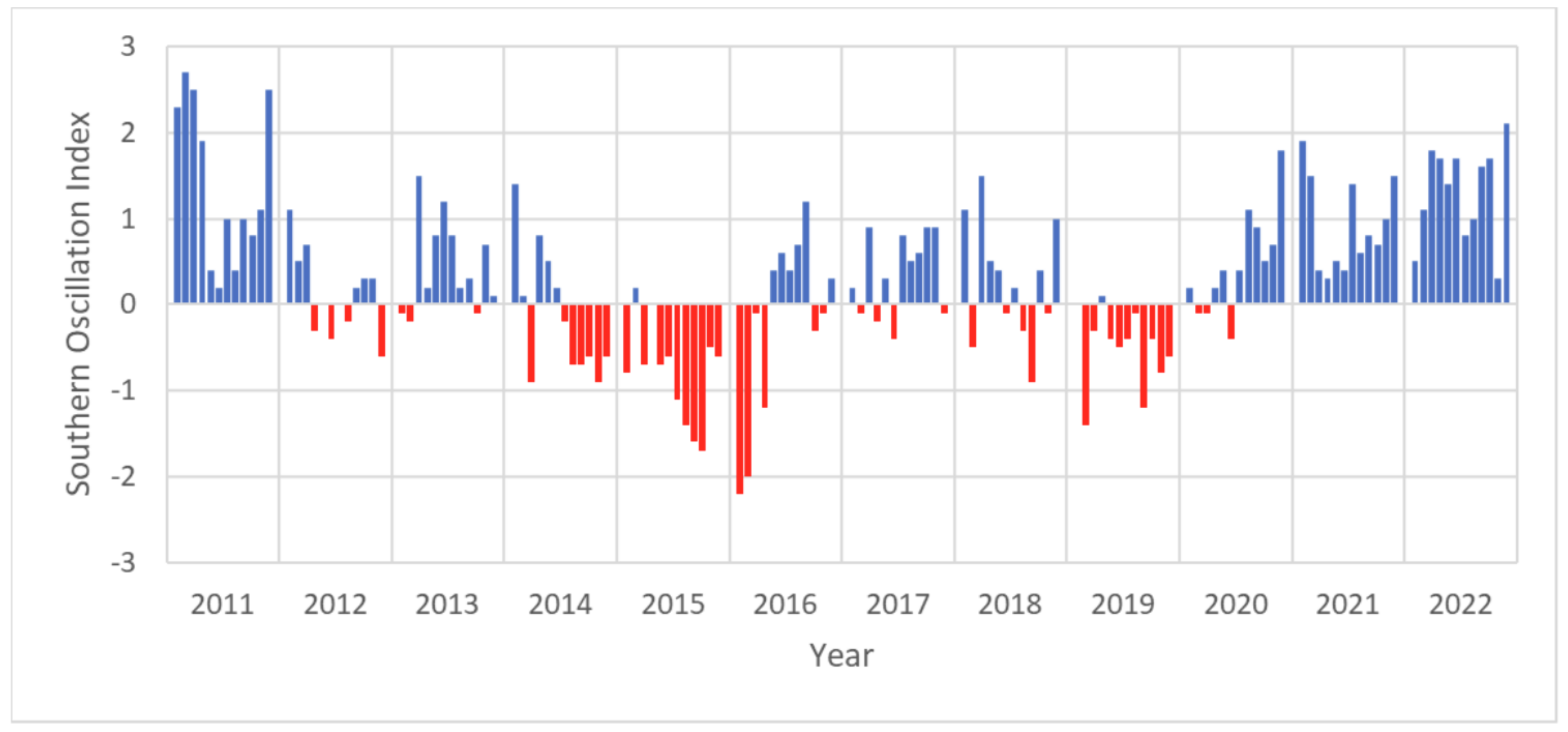
| El Niño Southern Oscillation (ENSO) | Measured by the Southern Oscillation Index (standardised index based on sea level pressure differences between Tahiti (French Polynesia) and Darwin, (Australia)). Downloaded in csv format and processed in MS Excel and Python. | National Oceanic and Atmospheric Administration (NOAA) URL: https://www.ncei.noaa.gov/access/monitoring/enso/soi (accessed on 1 July 2023) |
| Oil spills | Major events in the Atlantic and West Africa. Information/data from empirical publications. | Ogbuka et al. (2022) [40]; Fosu (2017) [41]; Stout et al. (2017) [42] |
| Atmospheric | ||
| Saharan Dust | Aerosol Optic Depth (a measure of the aerosols, including dust, distributed within a column of air from the instrument to the earth’s surface) derived from MODIS Terra data. Downloaded in csv format and processed in MS Excel and Python. | Platnick et al. (2015) [43] from Giovanni NASA Earth Data, (Data Product: MYD08_M3 v6.1) URL: https://giovanni.gsfc.nasa.gov/ (accessed on 1 July 2023) |
| Volcanic eruptions | Volcanic eruptions in the Atlantic region, which were also associated with ash, gas clouds or material production. | Smithsonian Institution, Global Volcanism Program URL: https://volcano.si.edu/ (accessed on 1 July 2023) |
| Nutrient inputs | ||
| Volcanic eruptions | (see above) | |
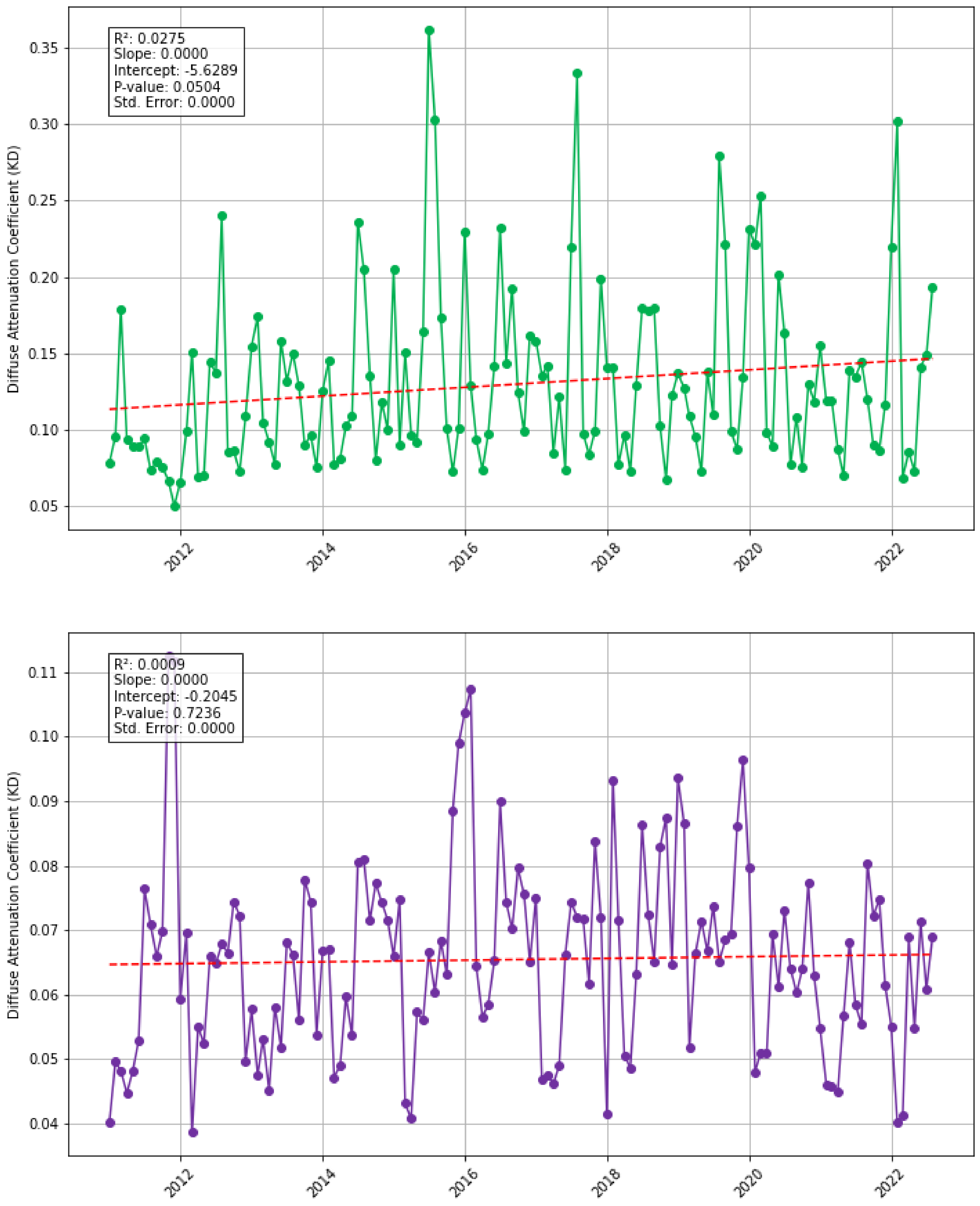
| Suspended sediment | Diffuse attenuation coefficient for downwelling irradiance (derived from MODIS Aqua data) measures how light dissipates with depth in water. Additionally, any notable impacts/events on water pollution quality or policies which came into effect 2009 onwards in the West African region were considered. Downloaded in csv format and processed in MS Excel and Python. | DOI: 10.5067/AQUA/MODIS/L3M/KD/2022 from Giovanni NASA Earth Data URL: https://giovanni.gsfc.nasa.gov/ (accessed on 1 July 2023) |
| Driver | Co-Variance | Causal Pathway | |||||
|---|---|---|---|---|---|---|---|
| Temporal Co-Variance | Temporal Delay | None Observed/Unknown | Persistence/Proliferation | Transfer to New | Transport Within | None Determined/Unknown | |
| Sea surface temperature | H | H | N.D/U | ||||
| Salinity | M | H | |||||
| NAO | N.O/U | N.D/U | |||||
| Saharan Dust | L | H | |||||
| Volcanic eruptions | L | H | |||||
| AMOC | H | L | L | L | |||
| ENSO | N.O/U | L | L | ||||
| Oil Spills | N.O/U | H | |||||
| Water quality/river pollution | L | L | L | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fidai, Y.A.; Dash, J.; Tompkins, E.; Atiglo, D.Y.; Jayson-Quashigah, P.-N.; Sowah, W.N.A.; Addo, K.A. Sargassum Biomass Movement and Proliferation in the Eastern Tropical Atlantic. Phycology 2025, 5, 17. https://doi.org/10.3390/phycology5020017
Fidai YA, Dash J, Tompkins E, Atiglo DY, Jayson-Quashigah P-N, Sowah WNA, Addo KA. Sargassum Biomass Movement and Proliferation in the Eastern Tropical Atlantic. Phycology. 2025; 5(2):17. https://doi.org/10.3390/phycology5020017
Chicago/Turabian StyleFidai, Yanna Alexia, Jadu Dash, Emma Tompkins, Donatus Yaw Atiglo, Philip-Neri Jayson-Quashigah, Winnie Naa Adjorkor Sowah, and Kwasi Appeaning Addo. 2025. "Sargassum Biomass Movement and Proliferation in the Eastern Tropical Atlantic" Phycology 5, no. 2: 17. https://doi.org/10.3390/phycology5020017
APA StyleFidai, Y. A., Dash, J., Tompkins, E., Atiglo, D. Y., Jayson-Quashigah, P.-N., Sowah, W. N. A., & Addo, K. A. (2025). Sargassum Biomass Movement and Proliferation in the Eastern Tropical Atlantic. Phycology, 5(2), 17. https://doi.org/10.3390/phycology5020017