
Influence of Algal Strain on Permeate Flux Rate in Crossflow Microfiltration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Algal Cultures Characteristics
2.2. Filtration Setup
3. Results and Discussion
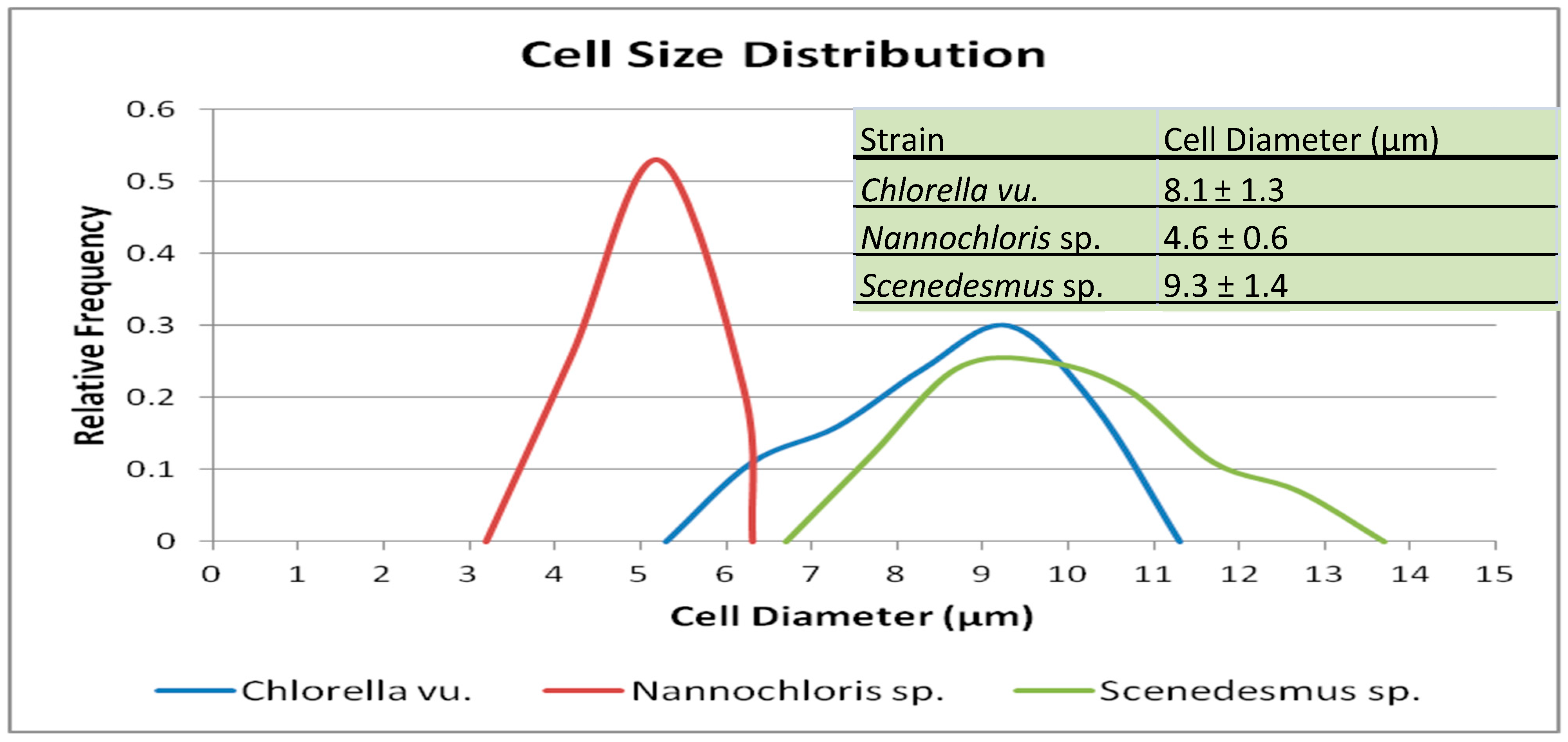
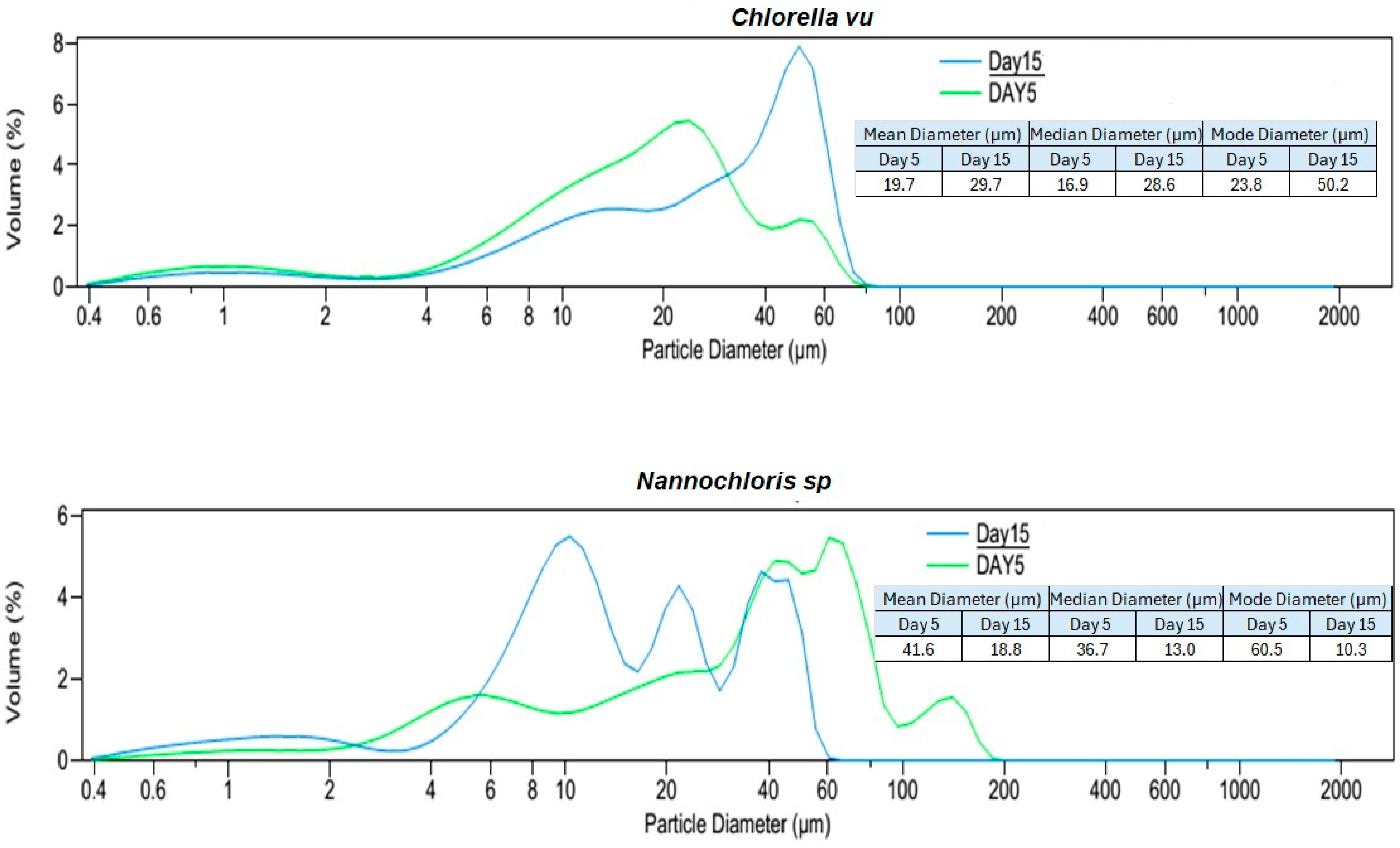
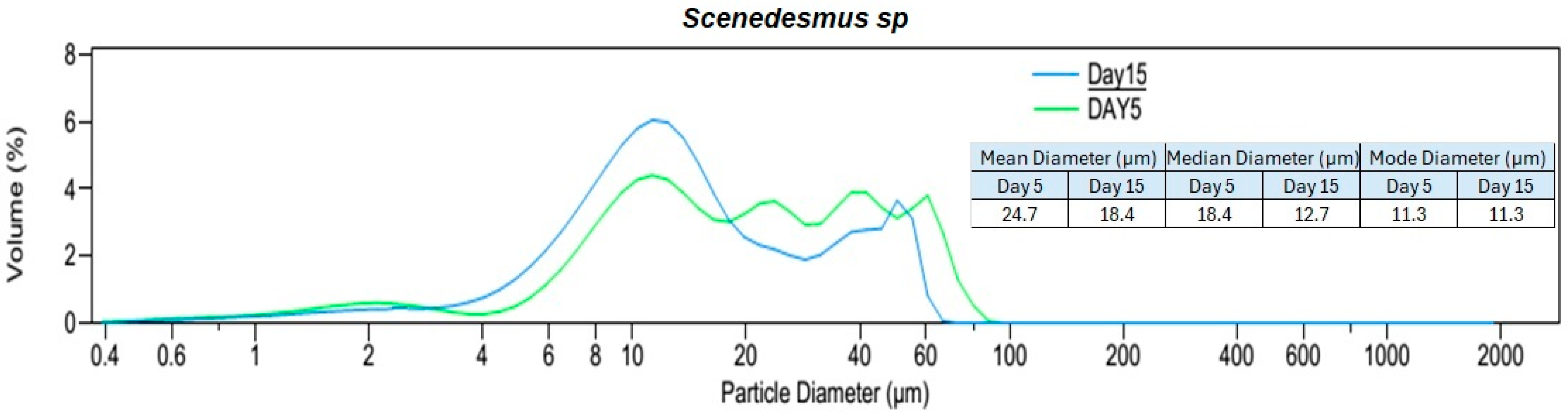
3.1. Algal Suspension Characterization
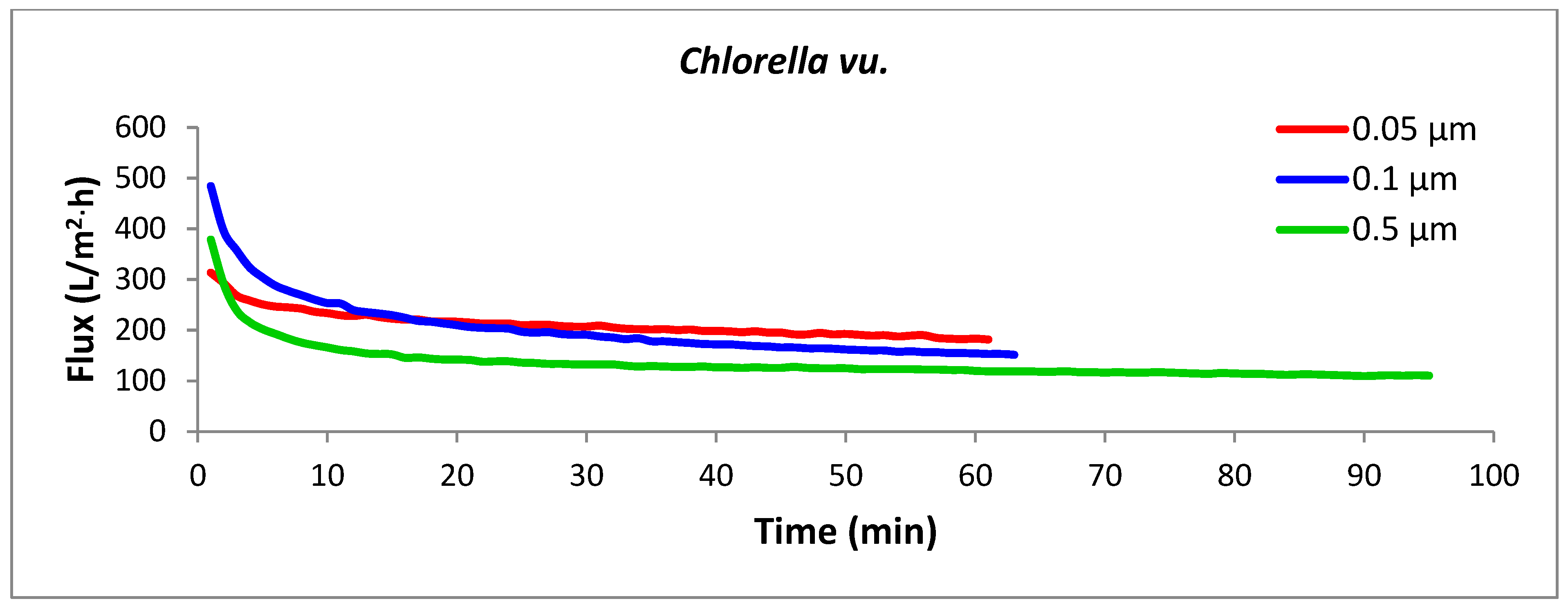
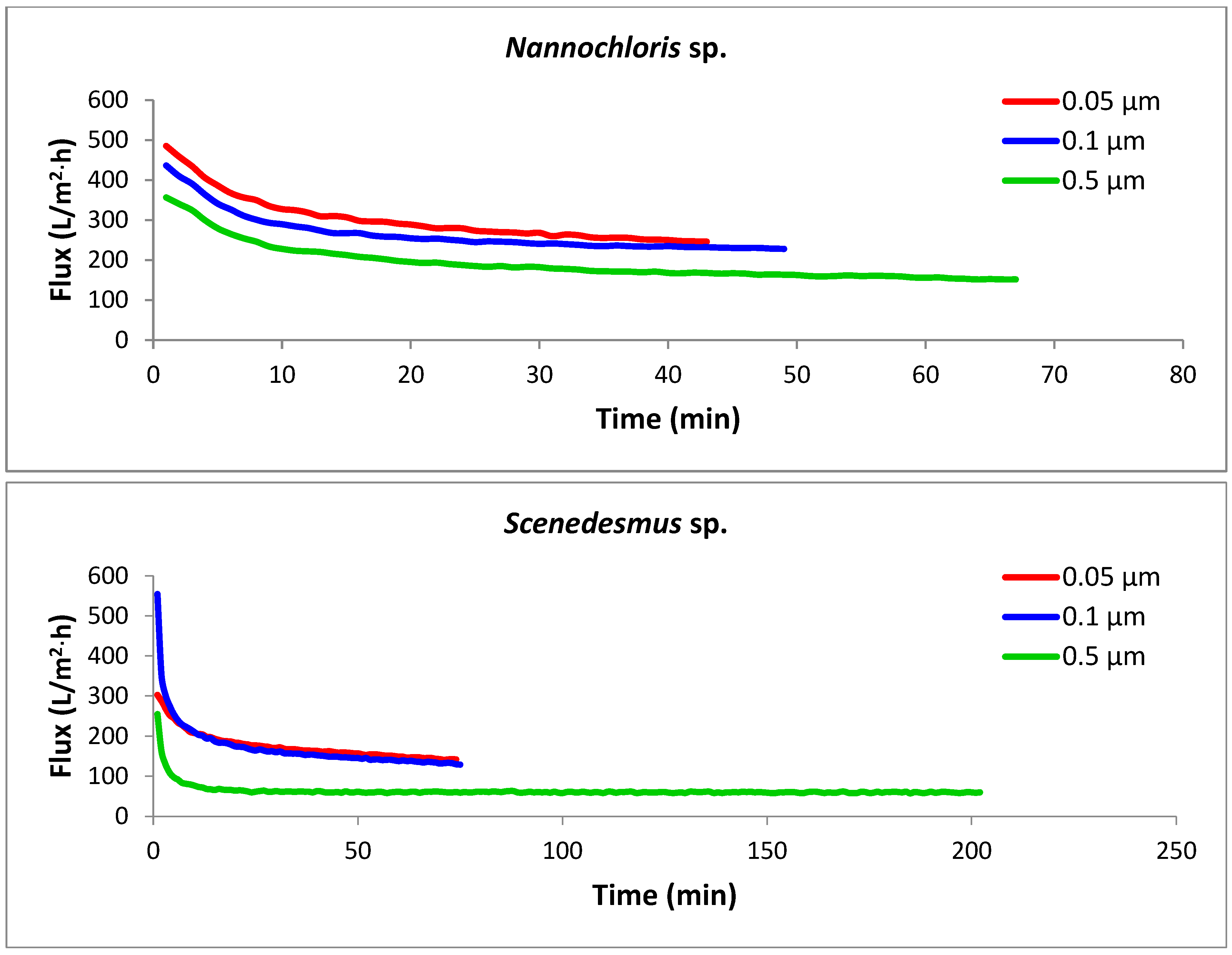
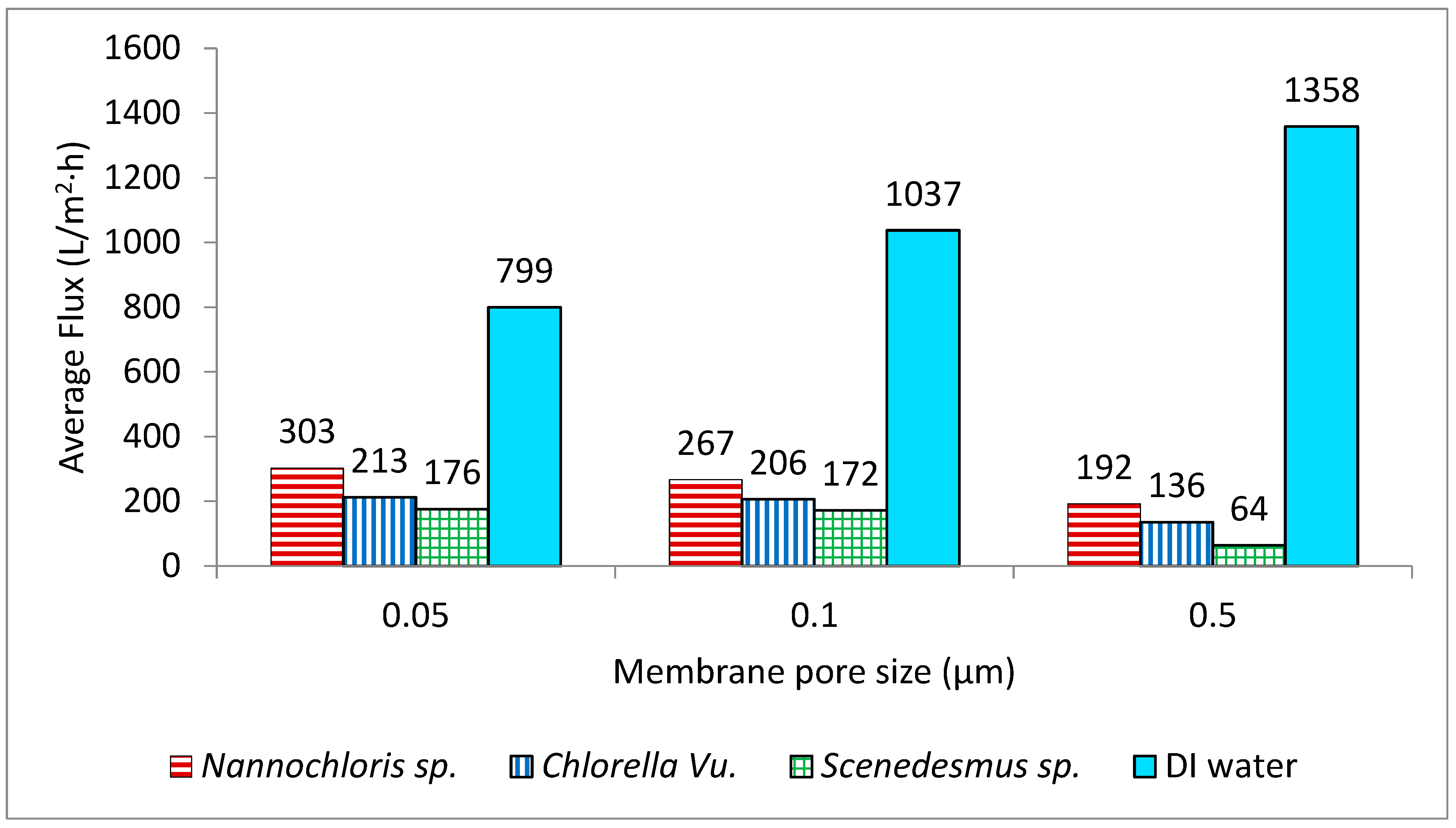
3.2. Filtration
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, C.; Zhao, Z.; Ma, S.; Rasool, M.A.; Wang, L.; Zhang, J. Optimization of ultrasonic-assisted extraction, refinement and characterization of water-soluble polysaccharide from Dictyosphaerium sp. and evaluation of antioxidant activity in vitro. J. Food Meas. Charact. 2020, 14, 963–977. [Google Scholar] [CrossRef]
- Ahmad, A.L.; Yasin, N.M.; Derek, C.J.C.; Lim, J.K. Microalgae as a sustainable energy source for biodiesel production: A review. Renew. Sust. Energy Rev. 2011, 15, 584–593. [Google Scholar] [CrossRef]
- Kang, A.; Lee, T.S. Converting sugars to biofuels: Ethanol and beyond. Bioengineering 2015, 2, 184–203. [Google Scholar] [CrossRef] [PubMed]
- Castro-Muñoz, R.; García-Depraect, O. Membrane-based harvesting processes for microalgae and their valuable-related molecules: A review. Membranes 2021, 11, 585. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, S.; Kalra, A.; Simsek, H.; Kumar, G.; Prajapati, S.K. Current trends and prospects in microalgae-based bioenergy production. J. Environ. Chem. Eng. 2020, 8, 104025. [Google Scholar] [CrossRef]
- Hu, Q. Progress and perspectives on microalgal mass culture. Algal Res. 2014, 4, 1–122. [Google Scholar] [CrossRef]
- Markets, R.M. $4.6 Billion Worldwide Microalgae Industry to 2027—Impact of COVID-19 on the Market. 2020. Available online: https://www.globenewswire.com/news-release/2020/09/18/2095854/28124/en/4-6-Billion-Worldwide-Microalgae-Industry-to-2027-Impact-of-COVID-19-on-the-Market.html (accessed on 28 April 2024).
- Zhao, Z.; Muylaert, K.; Vankelecom, I.F. Combining patterned membrane filtration and flocculation for economical microalgae harvesting. Water Res. 2021, 198, 117181. [Google Scholar] [CrossRef]
- Zhao, Z.; Ilyas, A.; Muylaert, K.; Vankelecom, I.F. Optimization of patterned polysulfone membranes for microalgae harvesting. Bioresour. Technol. 2020, 309, 123367. [Google Scholar] [CrossRef]
- Bilad, M.R.; Arafat, H.A.; Vankelecom, I.F. Membrane technology in microalgae cultivation and harvesting: A review. Biotechnol. Adv. 2014, 32, 1283–1300. [Google Scholar] [CrossRef]
- Fasaei, F.; Bitter, J.H.; Slegers, P.M.; van Boxtel, A.J.B. Technoeconomic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018, 31, 347–362. [Google Scholar] [CrossRef]
- Li, S.; Hu, T.; Xu, Y.; Wang, J.; Chu, R.; Yin, Z.; Mo, F.; Zhu, L. A review on flocculation as an efficient method to harvest energy microalgae: Mechanisms, performances, influencing factors and perspectives. Renew. Sustain. Energy Rev. 2020, 131, 110005. [Google Scholar] [CrossRef]
- Muhammad, G.; Alam, A.; Mofjur, M.; Jahirul, M.I.; Lv, Y.; Xiong, W.; Ong, H.C.; Xu, J. Modern developmental aspects in the field of economical harvesting and biodiesel production from microalgae biomass. Renew. Sustain. Energy Rev. 2021, 135, 110209. [Google Scholar] [CrossRef]
- Bansfield, D.; Spilling, K.; Mikola, A.; Piiparinen, J. Bioflocculation of Euglena gracilis via direct application of fungal filaments: A rapid harvesting method. J. Appl. Phycol. 2022, 34, 321–334. [Google Scholar] [CrossRef]
- Drexler, I.L.; Yeh, D.H. Membrane applications for microalgae cultivation and harvesting: A review. Rev. Environ. Sci. Bio-Technol. 2014, 13, 487–504. [Google Scholar] [CrossRef]
- Zhao, F.; Chu, H.; Yu, Z.; Jiang, S.; Zhao, X.; Zhou, X.; Zhang, Y. The filtration and fouling performance of membranes with different pore sizes in algae harvesting. Sci. Total Environ. 2017, 587, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Mohanty, K.A. Comprehensive review on microalgal harvesting strategies: Current status and future prospects. Algal Res. 2019, 44, 101683. [Google Scholar] [CrossRef]
- Mo, W.; Soh, L.; Werber, J.R.; Elimelech, M.; Zimmerman, J.B. Application of membrane dewatering for algal biofuel. Algal Res. 2015, 11, 1–12. [Google Scholar] [CrossRef]
- Landsman, M.R.; Sujanani, R.; Brodfuehrer, S.H.; Cooper, C.M.; Darr, A.G.; Davis, R.J.; Kim, K.; Kum, S.; Nalley, L.K.; Nomaan, S.M.; et al. Water treatment: Are membranes the panacea? Annu. Rev. Chem. Biomol. Eng. 2020, 11, 559–585. [Google Scholar] [CrossRef]
- Yazdanabad, S.K.; Samimi, A.; Shokrollahzadeh, S.; Kalhori, D.M.; Moazami, N.; González, M.J.; Sobczuk, T.M.; Grima, E.M. Microalgae biomass dewatering by forward osmosis: Review and critical challenges. Algal Res. 2021, 56, 102323. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, Q. Algal fouling of microfiltration and ultrafiltration membranes and control strategies: A review. Sep. Purif. Technol. 2018, 203, 193–208. [Google Scholar] [CrossRef]
- Miller, D.J.; Dreyer, D.R.; Bielawski, C.W.; Paul, D.R.; Freeman, B.D. Surface modification of water purification membranes. Angew. Chem. Int. Ed. 2017, 56, 4662–4711. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhao, Q.; Wang, P.; Chen, M.; Wang, Z.; Ma, J. Inhibition of algae-induced membrane fouling by in-situ formed hydrophilic micropillars on ultrafiltration membrane surface. J. Membr. Sci. 2021, 638, 119648. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Y.; He, M.; Su, Y.; Zhao, X.; Elimelech, M.; Jiang, Z. Antifouling membranes for sustainable water purification: Strategies and mechanisms. Chem. Soc. Rev. 2016, 45, 5888–5924. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.E.; Leite, G.B.; Hallenbeck, P.C. Addressing the challenges for sustainable production of algal biofuels: II. Harvesting and conversion to biofuels. Environ. Technol. 2013, 34, 1807–1836. [Google Scholar] [CrossRef]
- Dassey, A.J.; Theegala, C.S. Harvesting economics and strategies using centrifugation for cost effective separation of microalgae cells for biodiesel applications. Bioresour. Technol. 2013, 128, 241–245. [Google Scholar] [CrossRef]
- Danquah, M.K.; Gladman, B.; Moheimani, N.; Forde, G.M. Microalgal growth characteristics and subsequent influence on dewatering efficiency. Chem. Eng. J. 2009, 151, 73–78. [Google Scholar] [CrossRef]
- Baerdemaeker, T.; Lemmens, B.; Dotremont, C.; Fret, J.; Roef, L.; Goiris, K.; Diels, L. Benchmark study on algae harvesting with backwashable submerged flat panel membranes. Bioresour. Technol. 2013, 129, 582–591. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Nakao, S.; Kimura, S. Effects of size and compressibility of suspended particles and surface pore size of membrane on flux in crossflow filtration. J. Membr. Sci. 1993, 81, 173–190. [Google Scholar] [CrossRef]
- Connell, H.; Zhu, J.; Bassi, A. Effect of particle shape on crossflow filtration flux. J. Membr. Sci. 1999, 153, 121–139. [Google Scholar] [CrossRef]
- Lu, W.M.; Ju, S.C. Selective Particle Deposition in Crossflow Filtration. Sep. Sci. Technol. 1989, 24, 517–540. [Google Scholar] [CrossRef]
- Foley, G.; MacLoughlin, P.F.; Malone, D.M. Preferential deposition of smaller cells during crossflow microfiltration of a yeast suspension. Biotechnol. Tech. 1992, 6, 115–120. [Google Scholar] [CrossRef]
- McCarthy, A.; Walsh, P.; Foley, G. Experimental techniques for quantifying the cake mass, the cake and membrane resistances and the specific cake resistance during crossflow filtration of microbial suspensions. J. Membr. Sci. 2002, 201, 31–45. [Google Scholar] [CrossRef]
- Mota, M.; Teixeira, J.; Yelshin, A. Influence of cell-shape on the cake resistance in dead-end and crossflow filtrations. Sep. Purif. Technol. 2002, 27, 137–144. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khademi, M.; Theegala, C. Influence of Algal Strain on Permeate Flux Rate in Crossflow Microfiltration. Phycology 2025, 5, 18. https://doi.org/10.3390/phycology5020018
Khademi M, Theegala C. Influence of Algal Strain on Permeate Flux Rate in Crossflow Microfiltration. Phycology. 2025; 5(2):18. https://doi.org/10.3390/phycology5020018
Chicago/Turabian StyleKhademi, Mostafa, and Chandra Theegala. 2025. "Influence of Algal Strain on Permeate Flux Rate in Crossflow Microfiltration" Phycology 5, no. 2: 18. https://doi.org/10.3390/phycology5020018
APA StyleKhademi, M., & Theegala, C. (2025). Influence of Algal Strain on Permeate Flux Rate in Crossflow Microfiltration. Phycology, 5(2), 18. https://doi.org/10.3390/phycology5020018