
Coolia Species (Dinophyceae) from the Tropical South Atlantic Region: Evidence of Harmfulness of Coolia cf. canariensis Phylogroup II



Abstract
1. Introduction
2. Materials and Methods
2.1. Dinoflagellates Cultures
2.2. Experimental Design
2.2.1. Test Organism
2.2.2. Toxicity Tests
2.3. Data Analysis
3. Results
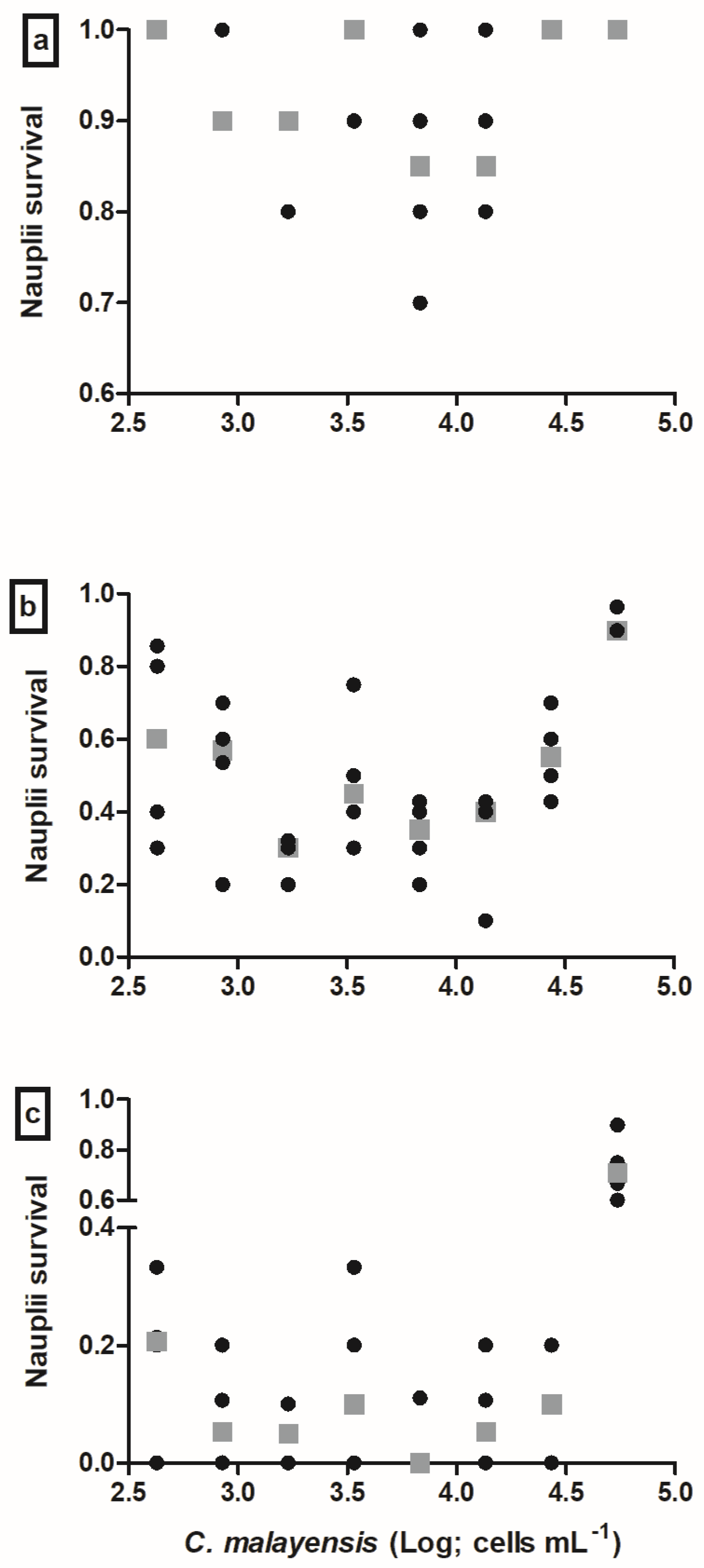
3.1. Nauplii Exposure to C. cf. canariensis PII
3.2. Nauplii Exposure to C. malayensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zingone, A.; Berdalet, E.; Bienfang, P.; Enevoldsen, H.; Evans, J.; Kudela, R.; Testers, P. Harmful Algae in Benthic Systems: A GEOHAB Core Research Program. Cryptogam. Algol. 2012, 33, 225–230. [Google Scholar] [CrossRef]
- Leaw, C.P.; Tan, T.H.; Lim, H.C.; Teng, S.T.; Yong, H.L.; Smith, K.F.; Rhodes, L.; Wolf, M.; Holland, W.C.; Vandersea, M.W.; et al. New scenario for speciation in the benthic dinoflagellate genus Coolia (Dinophyceae). Harmful Algae 2016, 55, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Hoppenrath, M.; Murray, S.A.; Chomerat, N.; Horiguchi, T. Marine Benthic Dinoflagellates—Unveiling Their Worldwide Biodiversity, 1st ed.; Senckenberg, Kleine Senckenberg-Reihe: Frankfurt, Germany, 2014; p. 276. [Google Scholar]
- Neves, R.A.F.; Rodrigues, E.T. Harmful Algal Blooms: Effect on Coastal Marine Ecosystems. In Life Below Water, Encyclopedia of the UN Sustainable Development Goals; Leal Filho, W., Azul, A.M., Brandli, L., Lange Salvia, A., Wall, T., Eds.; Springer Nature: Basel, Switzerland, 2020; pp. 1–31. [Google Scholar] [CrossRef]
- Neves, R.A.F.; Fernandes, T.; Santos, L.N.; Nascimento, S.M. Toxicity of benthic dinoflagellates on grazing, behavior and survival of the brine shrimp Artemia salina. PLoS ONE 2017, 12, e0175168. [Google Scholar] [CrossRef] [PubMed]
- Neves, R.A.F.; Contins, M.; Nascimento, S.M. Effects of the toxic benthic dinoflagellate Ostreopsis cf. ovata on fertilization and early development of the sea urchin Lytechinus Variegatus. Mar. Environ. Res. 2018, 135, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Neves, R.A.F.; Santiago, T.C.; Carvalho, W.F.; Silva, E.S.; da Silva, P.M.; Nascimento, S.M. Impacts of the toxic benthic dinoflagellate Prorocentrum lima on the brown mussel Perna perna: Shell-valve closure response, immunology, and histopathology. Mar. Environ. Res. 2019, 146, 35–45. [Google Scholar] [CrossRef]
- Neves, R.A.F.; Nascimento, S.M.; Santos, L.N. Harmful algal blooms and shellfish in the marine environment: An overview of the main molluscan responses, toxin dynamics, and risks for human health. Environ. Sci. Pollut. Res. 2021, 28, 55846–55868. [Google Scholar] [CrossRef] [PubMed]
- Wakeman, K.C.; Yamaguchi, A.; Roy, M.C.; Jenke-Kodama, H. Morphology, phylogeny and novel chemical compounds from Coolia malayensis (Dinophyceae) from Okinawa, Japan. Harmful Algae 2015, 44, 8–19. [Google Scholar] [CrossRef]
- Meunier, A. Microplankton de la mer Flamande. 3 Les Peridiniens. Mem. Mus. R. Hyst. Nat. Belg. 1919, 8, 3–116. [Google Scholar]
- Faust, M.A. Observations on the morphology and sexual reproduction of Coolia monotis (Dinophyceae). J. Phycol. 1992, 28, 94–104. [Google Scholar] [CrossRef]
- Faust, M.A. Observation of sand-dwelling toxic dinoflagellates (Dinophyceae) from widely differing sites, including two new species. J. Phycol. 1995, 31, 996–1003. [Google Scholar] [CrossRef]
- Ten-Hage, L.; Delaunay, N.; Pichon, V.; Couté, A.; Puiseux-Dao, S.; Turquet, J. Okadaic acid production from the marine benthic dinoflagellate Prorocentrum arenarium Faust (Dinophyceae) isolated from Europa Island Coral Reef Ecosystem (SW Indian Ocean). Toxicon 2000, 38, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Penna, A.; Bianconi, I.; Paz, B.; Zapata, M. Coolia canariensis sp. nov. (Dinophyceae), a new nontoxic epiphytic benthic dinoflagellate from the Canary Islands. J. Phycol. 2008, 44, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Leaw, C.P.; Lim, P.T.; Cheng, K.W.; Ng, B.K.; Usup, G. Morphology and molecular characterization of a new species of thecate benthic dinoflagellate, Coolia malayensis sp. nov. (Dinophyceae). J. Phycol. 2010, 46, 162–171. [Google Scholar] [CrossRef]
- Karafas, S.; York, R.; Tomas, C. Morphological and genetic analysis of the Coolia monotis species complex with the introduction of two new species, Coolia santacroce. Harmful Algae 2015, 46, 18–33. [Google Scholar] [CrossRef]
- David, H.; Laza-Martinez, A.; Rodriguez, F.; Fraga, S.; Orive, E. Coolia guanchica sp. nov. (Dinophyceae) a new epi-benthic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Eur. J. Phycol. 2020, 55, 76–88. [Google Scholar] [CrossRef]
- David, H.; Laza-Martínez, A.; García-Etxebarria, K.; Riobó, P.; Orive, E. Characterization of Prorocentrum elegans and Prorocentrum levis (Dinophyceae) from the Southeastern Bay of Biscay by morphology and molecular phylogeny. J. Phycol. 2014, 50, 718–726. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Silva, R.A.F.; Oliveira, F.; Fraga, S.; Salgueiro, F. Morphology and molecular phylogeny of Coolia tropicalis, Coolia malayensis and a new lineage of the Coolia canariensis species complex (dinophyceae) isolated from Brazil. Eur. J. Phycol. 2019, 54, 484–496. [Google Scholar] [CrossRef]
- Larsson, M.E.; Smith, K.F.; Doblin, M.A. First description of the environmental niche of the epibenthic dinoflagellate species Coolia palmyrensis, C. malayensis, and C. tropicalis (Dinophyceae) from Eastern Australia. J. Phycol. 2019, 55, 565–577. [Google Scholar] [CrossRef]
- Tibiriçá, C.E.J.D.A.; Sibat, M.; Fernandes, L.F.; Bilien, G.; Chomérat, N.; Hess, P.; Mafra, L.L., Jr. Diversity and toxicity of the genus Coolia Meunier in Brazil, and detection of 44-Methyl Gambierone in Coolia tropicalis. Toxins 2020, 12, 327. [Google Scholar] [CrossRef]
- Abdennadher, M.; Zouari, A.B.; Medhioub, W.; Penna, A.; Hamza, A. Characterization of Coolia Spp. (Gonyaucales, Dinophyceae) from Southern Tunisia: First record of Coolia malayensis in the Mediterranean Sea. Algae 2021, 36, 175–193. [Google Scholar] [CrossRef]
- Phua, Y.H.; Roy, M.C.; Lemer, S.; Husnik, F.; Wakeman, K.C. Diversity and Toxicity of pacific strains of the benthic dinoflagellate Coolia (Dinophyceae), with a look at the Coolia canariensis species complex. Harmful Algae 2021, 109, 102120. [Google Scholar] [CrossRef]
- Zhang, H.; Lü, S.; Cen, J.; Li, Y.; Li, Q.; Wu, Z. Morphology and molecular phylogeny of three species of Coolia (Dinophyceae) from Hainan Island, South China Sea. J. Oceanol. Limnol. 2021, 39, 1020–1032. [Google Scholar] [CrossRef]
- Tsikoti, C.; Genitsaris, S. Review of harmful algal blooms in the coastal Mediterranean Sea, with a focus on Greek waters. Diversity 2021, 13, 396. [Google Scholar] [CrossRef]
- Nascimento, S.M. Abundance of Coolia Genus in the Oceanic Trindade Island (South Atlantic Ocean, Brazil); Personal Communication; Federal University of the State of Rio de Janeiro: Rio de Janeiro, Brazil, 2023. [Google Scholar]
- Holmes, M.J.; Lewis, R.J.; Jones, A.; Hoy, A.W.W. Cooliatoxin, the first toxin from Coolia monotis (Dinophyceae). Nat. Toxins 1995, 3, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.L.; Thomas, A.E. Coolia monotis (Dinophyceae): A toxic epiphytic microalgal species found in New Zealand. New Zeal. J. Mar. Freshw. Res. 1997, 31, 139–141. [Google Scholar] [CrossRef]
- Laza-Martinez, A.; Orive, E.; Miguel, I. Morphological and Genetic Characterization of benthic dinoflagellates of the genera Coolia, Ostreopsis and Prorocentrum from the South-Eastern Bay of Biscay. Eur. J. Phycol. 2011, 46, 45–65. [Google Scholar] [CrossRef]
- Murray, J.S.; Nishimura, T.; Finch, S.C.; Rhodes, L.L.; Puddick, J.; Harwood, D.T.; Larsson, M.E.; Doblin, M.A.; Leung, P.; Yan, M.; et al. The role of 44-Methylgambierone in ciguatera fish poisoning: Acute toxicity, production by marine microalgae and its potential as a biomarker for Gambierdiscus Spp. Harmful Algae 2020, 97, 101853. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Leung, P.T.Y.; Gu, J.; Lam, V.T.T.; Murray, J.S.; Harwood, D.T.; Wai, T.-C.; Lam, P.K.S. Hemolysis associated toxicities of benthic dinoflagellates from Hong Kong waters. Mar. Pollut. Bull. 2020, 155, 111114. [Google Scholar] [CrossRef]
- Nakajima, I.; Oshima, Y.; Yasumoto, T. Toxicity of benthic dinoflagellates found in coral reef-II. Toxicity of benthic dinoflagellates in Okinawa. Nippon. Suisan Gakkaishi 1981, 47, 1029–1033. [Google Scholar] [CrossRef]
- Pagliara, P.; Caroppo, C. Toxicity assessment of Amphidinium carterae, Coolia cfr. monotis and Ostreopsis cfr. ovata (Dinophyta) isolated from the Northern Ionian Sea (Mediterranean Sea). Toxicon 2012, 60, 1203–1214. [Google Scholar] [CrossRef]
- Mendes, M.C.Q.; Nunes, J.M.C.; Fraga, S.; Rodríguez, F.; Franco, J.M.; Riobó, P.; Branco, S.; Menezes, M. Morphology, molecular phylogeny and toxinology of Coolia and Prorocentrum strains isolated from the tropical South Western Atlantic Ocean. Bot. Mar. 2019, 62, 125–140. [Google Scholar] [CrossRef]
- Rhodes, L.; Adamson, J.; Suzuki, T.; Briggs, L.; Garthwaite, I. Toxic marine epiphytic dinoflagellates, Ostreopsis siamensis and Coolia monotis (Dinophyceae), in New Zealand. N. Z. J. Mar. Freshw. Res. 2000, 34, 371–383. [Google Scholar] [CrossRef]
- Leung, P.T.Y.; Yan, M.; Yiu, S.K.F.; Lam, V.T.T.; Ip, J.C.H.; Au, M.W.Y.; Chen, C.Y.; Wai, T.C.; Lam, P.K.S. Molecular phylogeny and toxicity of harmful benthic dinoflagellates Coolia (Ostreopsidaceae, Dinophyceae) in a sub-tropical marine ecosystem: The first record from Hong Kong. Mar. Pollut. Bull. 2017, 124, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Yan, M.; Leung, P.T.Y.; Tian, L.; Lam, V.T.T.; Cheng, S.H.; Lam, P.K.S. Toxicity effects of hydrophilic algal lysates from Coolia tropicalis on marine medaka larvae (Oryzias melastigma). Aquat. Toxicol. 2021, 234, 105787. [Google Scholar] [CrossRef]
- Varela, A.T.; Neves, R.A.F.; Nascimento, S.M.; Oliveira, P.J.; Pardal, M.A.; Rodrigues, E.T.; Moreno, A.J. Mitochondrial impairment and cytotoxicity effects induced by the marine epibenthic dinoflagellate Coolia malayensis. Environ. Toxicol. Pharmacol. 2020, 77, 103379. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; Smith, K.; Papiol, G.G.; Adamson, J.; Harwood, T.; Munday, R. Epiphytic dinoflagellates in sub-tropical New Zealand, in particular the genus Coolia Meunier. Harmful Algae 2014, 34, 36–41. [Google Scholar] [CrossRef]
- Guillard, R.R.J. Culture methods. In Manual on Harmful Marine Microalgae; DIOC Manual and Guides; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO: Paris, France, 1995; Volume 33, pp. 45–56. [Google Scholar]
- Nunes, B.S.; Carvalho, F.D.; Guilhermino, L.M.; Van Stappen, G. Use of the genus Artemia in ecotoxicity testing. Environ. Pollut. 2006, 144, 453–462. [Google Scholar] [CrossRef]
- Beck, B.; Chen, Y.-F.; Dere, W.; Devanarayan, V.; Eastwood, B.J.; Farmen, M.W.; Iturria, S.J.; Iversen, P.W.; Kahl, S.D.; Moore, R.A.; et al. Assay Operations for SAR Support; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004.
- Calabrese, E.J. Evidence that hormesis represents an “overcompensation” response to a disruption in homeostasis. Ecotoxicol. Environ. Saf. 1999, 42, 135–137. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. Hormesis: A generalizable and unifying hypothesis. Crit. Rev. Toxicol. 2001, 31, 353–424. [Google Scholar] [CrossRef]
- Naveira, C.; Rodrigues, N.; Santos, F.S.; Santos, L.N.; Neves, R.A.F. Acute toxicity of bisphenol a (bpa) to tropical marine and estuarine species from different trophic groups. Environ. Pollut. 2021, 268, 115911. [Google Scholar] [CrossRef]
- Neves, R.A.F.; Nascimento, S.M.; Santos, L.N. Sublethal fish responses to short-term food chain transfer of DSP toxins: The role of somatic condition. J. Exp. Mar. Bio. Ecol. 2020, 524, 151317. [Google Scholar] [CrossRef]
- Hansen, P.J.; Cembella, A.D.; Moestrup, Ø. The marine dinoflagellate Alexandrium ostenfeldii: Paralytic shellfish toxin concentration, composition, and toxicity to a tintinnid ciliate. J. Phycol. 1992, 28, 597–603. [Google Scholar] [CrossRef]
- Fulco, V.K. Harmful effects of the toxic dinoflagellate Alexandrium tamarense on the tintinnids Favella taraikaensis and Eutintinnus sp. J. Mar. Biol. Assoc. UK 2007, 87, 1085–1088. [Google Scholar] [CrossRef]
- Sulkin, S.; Hinz, S.; Rodriguez, M. Effects of exposure to toxic and non-toxic dinoflagellates on oxygen consumption and locomotion, in stage 1 larvae of the crabs Cancer oregonensis and C. magister. Mar. Biol. 2003, 142, 205–211. [Google Scholar] [CrossRef]
- Silva, N.J.; Tang, K.W.; Lopes, R.M. Effects of microalgal exudates and intact cells on subtropical marine zooplankton. J. Plankton Res. 2013, 35, 855–865. [Google Scholar] [CrossRef]
- Borcier, E.; Morvezen, R.; Boudry, P.; Miner, P.; Charrier, G.; Laroche, J.; Hegaret, H. Effects of bioactive extracellular compounds and paralytic shellfish toxins produced by Alexandrium minutum on growth and behaviour of juvenile great scallops Pecten maximus. Aquat. Toxicol. 2017, 184, 142–154. [Google Scholar] [CrossRef]
- Pan, L.; Chen, J.; Shen, H.; He, X.; Li, G.; Song, X.; Zhou, D.; Sun, C. Profiling of extracellular toxins associated with diarrhetic shellfish poison in Prorocentrum lima culture medium by high-performance liquid chromatography coupled with mass spectrometry. Toxins 2017, 9, 308. [Google Scholar] [CrossRef] [PubMed]
- Castrec, J.; Soudant, P.; Payton, L.; Tran, D.; Miner, P.; Lambert, C.; Le Goïc, N.; Huvet, A.; Quillien, V.; Boullot, F.; et al. Bioactive extracellular compounds produced by the dinoflagellate Alexandrium minutum are highly detrimental for oysters. Aquat. Toxicol. 2018, 199, 188–198. [Google Scholar] [CrossRef]
- Long, M.; Tallec, K.; Soudant, P.; Lambert, C.; Le Grand, F.; Sarthou, G.; Jolley, D.; Hégaret, H. A rapid quantitative fluorescence-based bioassay to study allelochemical interactions from Alexandrium minutum. Environ. Pollut. 2018, 242, 1598–1605. [Google Scholar] [CrossRef]
- Neely, T.; Campbell, L. A modified assay to determine hemolytic toxin variability among Karenia clones isolated from the Gulf of Mexico. Harmful Algae 2006, 5, 592–598. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| C. cf. canariensis PII Concentration (Cells mL−1) | Cumulative Mortality (%) | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| 0 | 1.67 | 4.17 | 5.83 |
| 330 | 12.5 * | 35.0 * | 75.0 |
| 660 | 15.0 * | 40.0 * | 77.5 |
| 1320 | 12.5 | 50.0 * | 87.5 |
| 2641 | 5.0 | 37.5 * | 87.5 |
| 5283 | 5.0 | 45.0 * | 80.0 |
| 10,567 | 12.5 | 45.0 * | 95.0 * |
| 21,135 | 35.0 * | 75.0 * | 95.0 * |
| 42,270 | 50.0 * | 97.5 * | 100 * |
| Exposure Time (h) | LC50 (Cells mL−1) | 95% CI (Cells mL−1) | R2 | FE (%) |
|---|---|---|---|---|
| 24 | 18,064 | 10,815–30,171 | 0.853 | 23.3 |
| 48 | 19,968 | 11,343–35,150 | 0.829 | 25.70 |
| C. malayensis Concentration (Cells mL−1) | Cumulative Mortality (%) | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| 0 | 0 | 1.6 | 3.3 |
| 426 | 0 | 42.5 | 82.5 |
| 852 | 7.5 | 50.0 | 92.5 * |
| 1704 | 12.5 | 72.5 * | 95.0 * |
| 3408 | 2.5 | 52.5 | 87.5 |
| 6816 | 15.0 | 67.5 * | 97.5 * |
| 13,633 | 12.5 | 67.5 * | 92.5 * |
| 27,266 | 0 | 45.0 | 90.0 |
| 54,531 | 0 | 10.0 | 30.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miralha, A.; Nascimento, S.M.; Neves, R.A.F. Coolia Species (Dinophyceae) from the Tropical South Atlantic Region: Evidence of Harmfulness of Coolia cf. canariensis Phylogroup II. Phycology 2023, 3, 242-254. https://doi.org/10.3390/phycology3020015
Miralha A, Nascimento SM, Neves RAF. Coolia Species (Dinophyceae) from the Tropical South Atlantic Region: Evidence of Harmfulness of Coolia cf. canariensis Phylogroup II. Phycology. 2023; 3(2):242-254. https://doi.org/10.3390/phycology3020015
Chicago/Turabian StyleMiralha, Agatha, Silvia M. Nascimento, and Raquel A. F. Neves. 2023. "Coolia Species (Dinophyceae) from the Tropical South Atlantic Region: Evidence of Harmfulness of Coolia cf. canariensis Phylogroup II" Phycology 3, no. 2: 242-254. https://doi.org/10.3390/phycology3020015
APA StyleMiralha, A., Nascimento, S. M., & Neves, R. A. F. (2023). Coolia Species (Dinophyceae) from the Tropical South Atlantic Region: Evidence of Harmfulness of Coolia cf. canariensis Phylogroup II. Phycology, 3(2), 242-254. https://doi.org/10.3390/phycology3020015