
Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Obtention of the Seaweed Liquid Extracts (SLEs)
2.2.1. Preparation of the U. lactuca Aqueous Extract
2.2.2. Preparation of the U. lactuca Ethanolic Extract
2.3. Evaluation of U. lactuca Extracts as Protection Inducers
2.4. Tissue Sampling for Analysis
2.5. Enzyme Assays
2.5.1. The Activity of β-1,3-glucanase
2.5.2. The Activity of Peroxidase Enzyme
2.6. Statistical Analysis
3. Results and Discussion
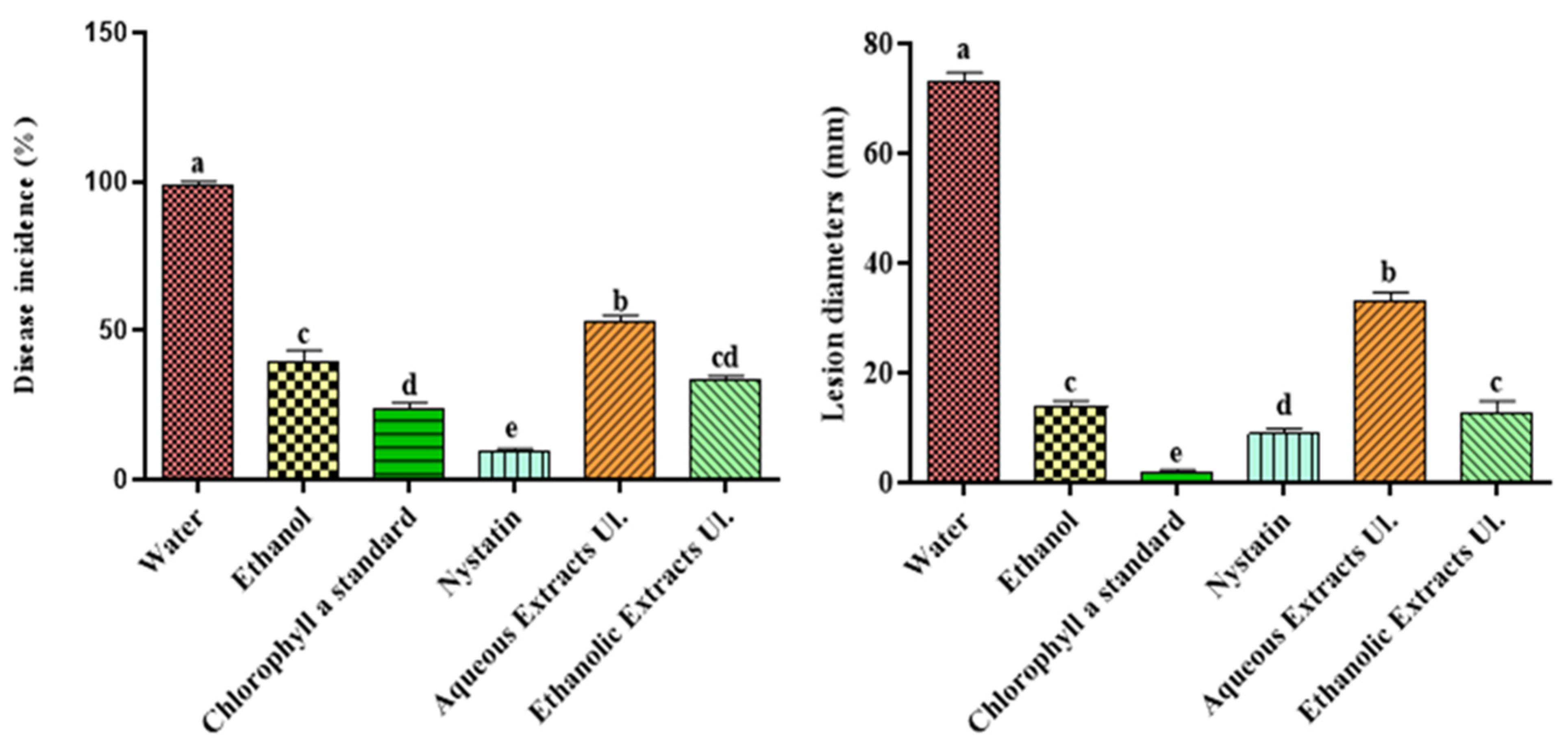
3.1. Effect of Extracts on Green Mold (Artificial Infection)
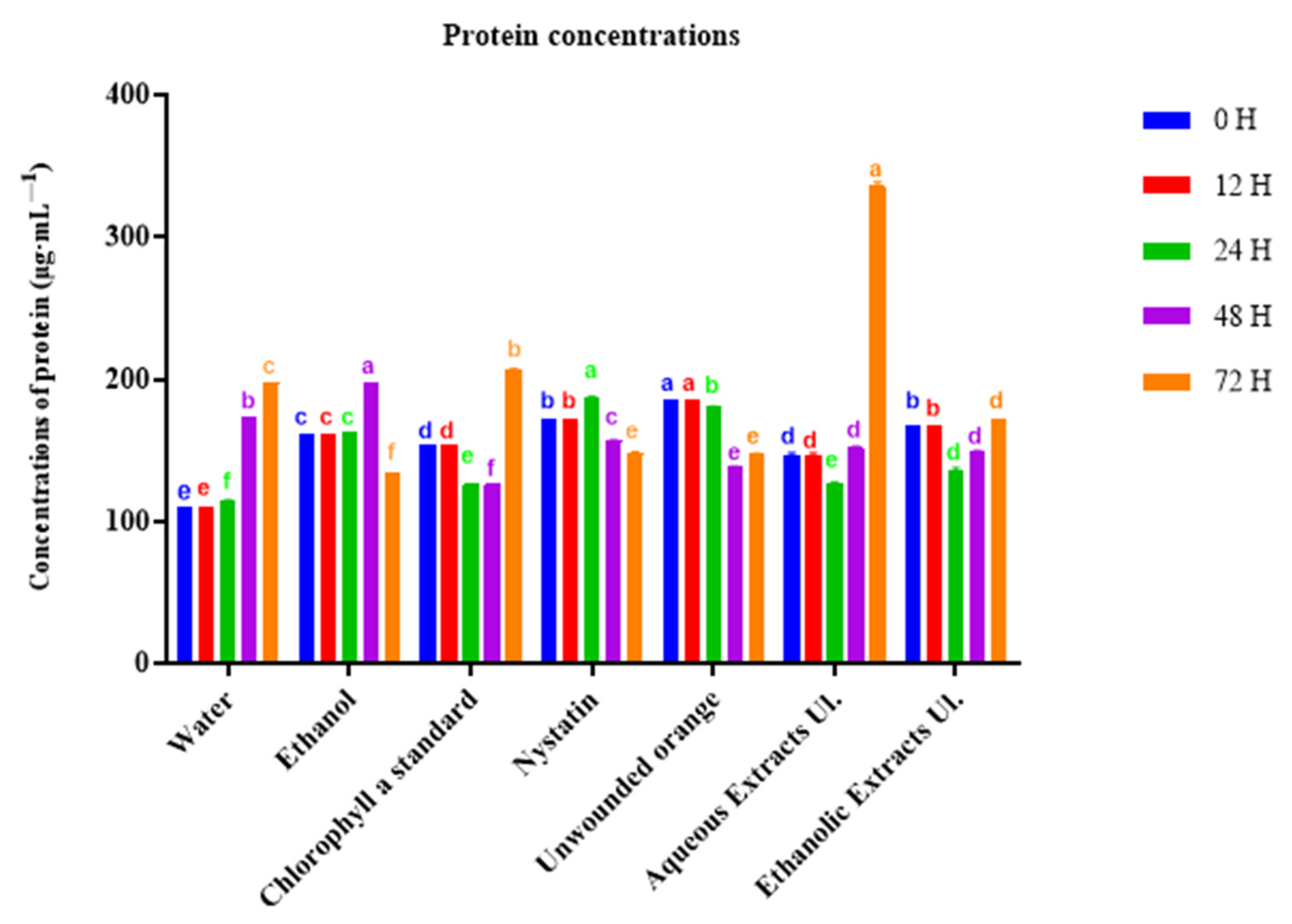
3.2. Protein Quantification
3.3. Enzyme Activity Assays
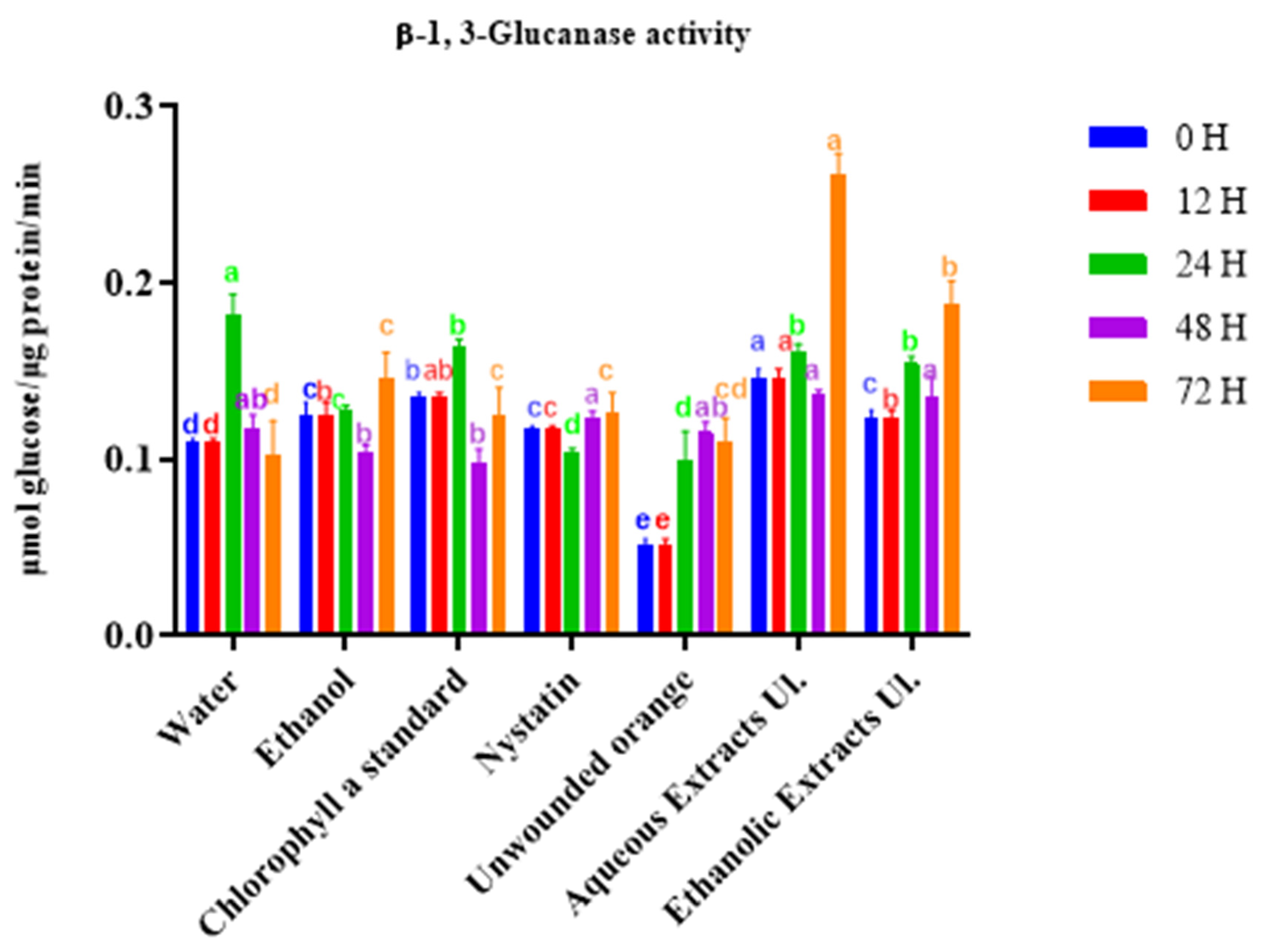
3.3.1. β-1,3-glucanase Activity
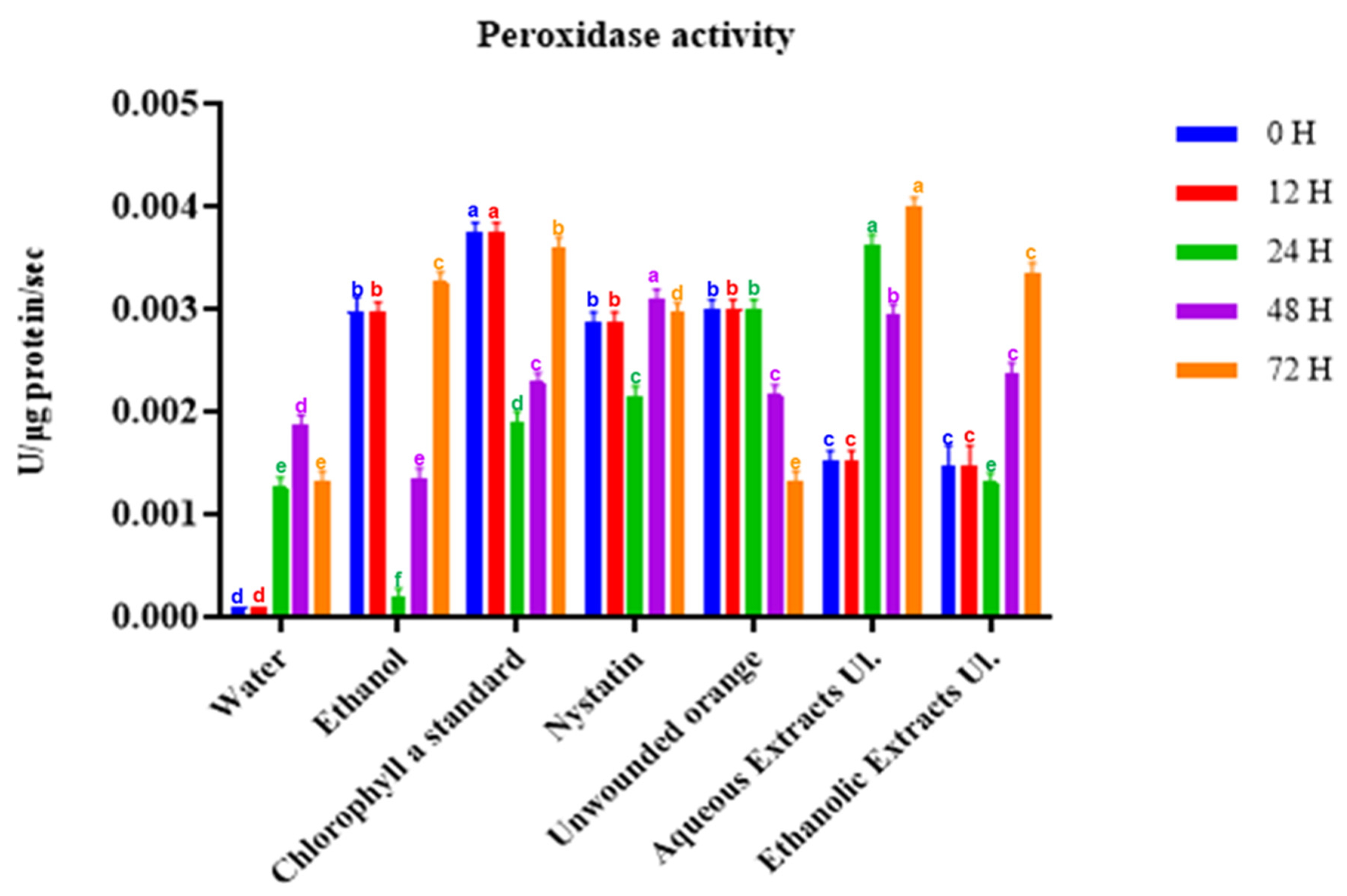
3.3.2. Peroxidase Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caserta, R.; Teixeira-Silva, N.S.; Granato, L.M.; Dorta, S.O.; Rodrigues, C.M.; Mitre, L.K. Citrus biotechnology: What has been done to improve disease resistance in such an important crop? Biotechnol. Res. Innov. 2020, 3, 95–109. [Google Scholar] [CrossRef]
- Dillard, C.J.; German, J.B. Phytochemicals: Nutraceuticals and human health. J. Sci. Food Agric. 2000, 80, 1744–1756. [Google Scholar] [CrossRef]
- Kanetis, L.; Förster, H.; Adaskaveg, J.E. Comparative efficacy of the new postharvest fungicides azoxystrobin, fludioxonil and pyrimethanil for managing citrus green mold. Plant Dis. 2017, 91, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Macarisin, D.; Cohen, L.; Eick, A.; Rafael, G.; Belausov, E.; Wisniewski, M.; Droby, S. Penicillium digitatum suppresses production of hydrogen peroxide in host tissue during infection of citrus fruit. Phytopathology 2007, 97, 1491–1500. [Google Scholar] [CrossRef]
- Palou, L.; Smilanick, J.L.; Droby, S. Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Postharvest Rev. 2008, 2, 1–16. [Google Scholar]
- Moraes Bazioli, J.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; de Moraes Pontes, J.-G.; Kupper, K.C.; Augusto, F.; de Carvalho, J.E.; Fill, T.P. Biological Control of Citrus Postharvest Phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef]
- Kharchoufi, S.; Parafati, L.; Licciardello, F.; Muratore, G.; Hamdi, M.; Cirvilleri, G.; Restuccia, C. Edible coatings incorporating pomegranate peel extract and biocontrol yeast to reduce Penicillium digitatum postharvest decay of oranges. Food Microbiol. 2018, 74, 107–112. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Ji, S.; Chen, T.; Tian, S.; Qin, G. Enhancement of biocontrol efficacy of Cryptococcus laurentii by cinnamic acid against Penicillium italicum in citrus fruit. Postharvest Biol. Technol. 2019, 149, 42–49. [Google Scholar] [CrossRef]
- Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernandez-Montiel, L.G. Biocontrol of postharvest fruit fungal diseases by bacterial antagonists: A review. Agronomy 2019, 9, 121. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. The effect of locust bean gum (LBG)-based edible coatings carrying biocontrol yeasts against Penicillium digitatum and Penicillium italicum causal agents of postharvest decay of mandarin fruit. Food Microbiol. 2016, 58, 87–94. [Google Scholar] [CrossRef]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials—A review. Plants 2016, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Hammet, S.M.; El-Rhman, A.A.; Abdel-Raouf, N.; Ibraheem, I.B.M. Role of marine macroalgae in plant protection & improvement for sustainable agriculture technology. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar] [CrossRef]
- Suthin Raj, T.; Vignesh, S.; Nishanthi, P.; Hane graff, K.; Ann Suji, H. Induction of defence enzymes activities in grape plant treated by seaweed algae against Plasmopara viticola and Uncinula necator causing downy and powdery mildews of grapes. Nov. Res. Microbiol. J. 2018, 2, 122–137. [Google Scholar] [CrossRef]
- Salim, D.; de Caro, P.; Merah, O.; Chbani, A. Control of post-harvest citrus green mold using Ulva lactuca extracts as a source of active substances. Int. J. Biores. Stress Manag. 2020, 11, 287–296. [Google Scholar] [CrossRef]
- Salim, D.; de Caro, P.; Chasseray, X.; Sing, A.S.C. Development of biobased emulsions for postharvest citrus fruit preservation. Sustain. Chem. Pharm. 2022, 25, 100583. [Google Scholar] [CrossRef]
- Romanazzi, G.; Feliziani, E.; Santini, M.; Landi, L. Effectiveness of postharvest treatment with chitosan and other resistance inducers in the control of storage decay of strawberry. Postharvest Biol. Technol. 2013, 75, 24–27. [Google Scholar] [CrossRef]
- Asimakis, E.; Shehata, A.A.; Eisenreich, W.; Acheuk, F. Algae and their metabolites as potential bio-pesticides. Microorganisms 2022, 10, 307. [Google Scholar] [CrossRef]
- Costa, J.H.; Bazioli, J.M.; de Moraes Pontes, J.G.; Fill, T.P. Penicillium digitatum infection mechanisms in citrus: What do we know so far? Fungal. Biol. 2019, 123, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Lafuente, M.T.; González-Candelas, L. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit–Penicillium digitatum interaction. Postharvest Biol. Technol. 2010, 56, 31–38. [Google Scholar] [CrossRef]
- Youssef, K.; Sanzani, S.M.; Ligorio, A.; Ippolito, A.; Terry, L.A. Sodium carbonate and bicarbonate treatments induce resistance to postharvest green mould on citrus fruit. Postharvest Biol. Technol. 2014, 87, 61–69. [Google Scholar] [CrossRef]
- Fallanaj, F.; Ippolito, A.; Ligorio, A.; Garganese, F.; Zavanella, C.; Sanzani, S.M. Electrolyzed sodium bicarbonate inhibits Penicillium digitatum and induces defence responses against green mould in citrus fruit. Postharvest Biol. Technol. 2016, 115, 18–29. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, F.; Lu, Y.; Deng, J. Combination of chitosan and salicylic acid to control postharvest green mold caused by Penicillium digitatum in grapefruit fruit. Sci. Hortic. 2018, 233, 54–60. [Google Scholar] [CrossRef]
- Abouraïcha, E.; El Alaoui-Talibi, Z.; El Boutachfaiti, R.; Petit, E.; Courtois, B.; Courtois, J.; El Modafar, C. Induction of natural defense and protection against Penicillium expansum and Botrytis cinerea in apple fruit in response to bioelicitors isolated from green algae. Sci. Hortic. 2015, 181, 121–128. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In The Protein Protocols Handbook, 2nd ed.; Walker, J.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2002; pp. 15–21. [Google Scholar]
- Abeles, F.B.; Forrence, L.E. Temporal and hormonal control of β-1,3-glucanase in Phaseolus Vulgaris L. Plant Physiol. 1979, 45, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Hammer-Schmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Bhagavathy, S.; Sumathi, P.; Jancy Sherene Bell, I. Green algae Chlorococcum humicola-a new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop. Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Li Destri Nicosia, G.M.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of postharvest fungal rots on citrus fruit and sweet cherries using a pomegranate peel extract. Postharvest Biol. Technol. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Martínez, P.G.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Papoutsis, K.; Mathioudakis, M.M.; Hasperué, J.H.; Ziogas, V. Non-chemical treatments for preventing the postharvest fungal rotting of citrus caused by Penicillium digitatum (green mold) and Penicillium italicum (blue mold). Trends Food Sci. Technol. 2019, 86, 479–491. [Google Scholar] [CrossRef]
- Llorens, E.; García-Agustín, P.; Lapeña, I. Advances in induced resistance by natural compounds: Towards new options for woody plant protection. Sci. Agric. 2018, 74, 90–100. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Y.; Luo, W.; Deng, L.; Yao, S.; Zeng, K. Primary metabolites analysis of induced citrus fruit disease resistance upon treatment with oligochitosan, salicylic acid and Pichia membranaefaciens. Biol. Control. 2020, 148, 104289. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (Liquid Extracts) | Total Polyphenol Content (mg GAE.g–1 Dry Extract) * | |
|---|---|---|
| Chlorophyll a standard | - | 0.5 mg·mL–1 ethanol |
| U. l. aqueous extract: 12 mg DM·mL–1 | 0.45 ± 0.01 | 0.5 ± 0.1 mg·g–1 DE |
| U. l. ethanolic extract: 2.9 mg DM·mL–1 | 1.66 ± 0.05 | 1.1 ± 0.1 mg·g–1 DE |
| Unwounded fruit: untreated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salim, D.; Al-Alam, J.; Merah, O.; Chbani, A.; de Caro, P. Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges. Phycology 2023, 3, 202-210. https://doi.org/10.3390/phycology3010013
Salim D, Al-Alam J, Merah O, Chbani A, de Caro P. Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges. Phycology. 2023; 3(1):202-210. https://doi.org/10.3390/phycology3010013
Chicago/Turabian StyleSalim, Douaa, Josephine Al-Alam, Othmane Merah, Asma Chbani, and Pascale de Caro. 2023. "Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges" Phycology 3, no. 1: 202-210. https://doi.org/10.3390/phycology3010013
APA StyleSalim, D., Al-Alam, J., Merah, O., Chbani, A., & de Caro, P. (2023). Extracts of Ulva lactuca Induce Responses against Penicillium digitatum on Oranges. Phycology, 3(1), 202-210. https://doi.org/10.3390/phycology3010013