Abstract
The histopathological effects of microplastics (MPs) in the gastrointestinal tracts of fish following long-term exposure and depuration are relatively understudied. This study investigated histomorphological damage in the small intestine of Oreochromis urolepis larvae following 65 d exposure to 38–45 μm of polyethylene microspheres (PE MPs) and after a recovery period of 60 d. Larval fish were assigned to each treatment group (control, 1, 10 and 100 PE MPs), where ingestion and degenerative changes in the small intestine were examined using a routine hematoxylin and eosin staining technique. The results highlighted significant PE MPs ingestion and retention proportional to exposure dose (χ2 = 49.54; df = 2). Villi height and width and epithelial cell height were significantly affected and differed between treatment groups. Indices of damage to the small intestine organ (χ2 = 47.37; df = 2; p < 0.05) and reaction patterns of villi, epithelial, goblet and cryptic glandular cells, leucocytic infiltration and blood congestion revealed significant occurrence of alteration as PE MPs exposure dose increased. After the recovery period, no PE MPs were observed, and villi height, width and epithelial cells showed recovery with no significant difference between treatment groups. Organ indices declined (χ2 = 12; df = 2; p < 0.05) but remained significantly different between treatment groups, largely due to leucocytic infiltration (χ2 = 9.08; df = 2; p < 0.05). The study demonstrated that microplastics induced small intestinal wall degeneration, but recovery in young fish occurred slowly. The damage likely compromised its digestive function, which may affect growth and reproduction. This requires further research.
1. Introduction
The study of the presence, fate and effects of microplastics (MPs, <5mm in size) in aquatic systems in Africa remains limited in comparison to other regions of the world [1,2]. This is despite Africa having many of the most notable freshwater bodies in the world including the African Great Lakes (Lakes Victoria, Malawi and Tanganyika), and the Rivers Nile and Congo, which are known to have high levels of species diversity and richness [3]. In Africa, MPs have been recovered from the gastrointestinal tracts of a number of ecologically and economically important fish species, including Nile perch (Lates niloticus) and Nile tilapia (Orechromis niloticus) from Lake Victoria in Tanzania [4], Catfish (Bagrus bajad) from the Nile River in Egypt [5], and Nile tilapia (Orechromis niloticus), Catfish (Clarias gariepinus), Common carp (Cyprinus carpio) and Crucian carp (Carassius carassius) from Lake Ziway in Ethiopia [6]. However, there is limited knowledge on the fate of MPs following ingestion (i.e., retention or depuration) and the localized toxicological damage this may cause in the gastrointestinal tracts of fish in general, and especially from Africa.
Elsewhere, it is reported that the ingestion of MPs has caused significant gastrointestinal (GI) tract damage in fish [7,8]. Physical abrasion of MPs passing through the GI tract can result in damages to the small intestines including the loss of cryptic cells, villi cracking, hyperemia and luecocytic infiltration, as reported in Dicentrarchus labrax [9], Girella laevifrons [10] and D. rerio [11] in a dose-dependent manner. Unlike wild fish, the majority of exposure experiments are acute and use marine fish which either were acclimatized and fed non-MPs feed for a couple of days or lengthily starved prior to the commencement of the experiment, contrary to natural environmental setting, where exposure to fish essentially begins at the larval stage. Investigations into the possibility that damaged gastrointestinal tracts are able to recover post exposure to MPs are scant, particularly in Africa. The existing depuration studies conducted so far are few, relatively short-term and focus on physiological status [12] such as oxidative stress [13] or microbiota composition [14], rather than on the gastrointestinal tract, which directly interacts with MP pollutants. However, the affected aquatic organisms are expected to recover following a period without MP exposure. Such insight has encouraged the development of plastic pollution-curbing strategies at national, regional and global levels [15], recognizing plastic-induced fatality and their ubiquity as a global concern [16]. Various legal frameworks [17] and bilateral agreements [16,18] have been ratified and some already implemented, including usage and disposal restrictions [19] to ensure zero plastic in aquatic habitats. Therefore, a broad knowledge of the effects of plastic pollution on fish recovery after gut voidance is critical for ecological conservation and waste management.
With this in mind, this study aimed to investigate intestinal histomorphological impairment of Wami tilapia (Oreochromis urolepis) after long-term exposure to polyethylene microspheres (PE MPs) and recovery after an equally long depuration period. The exposure period was 65 d, starting at larvae and continuing to juveniles, followed by 60 d of recovery. Long term studies such as this are rare and in the case of African freshwater fish this study is novel. O. urolepis has economic potential in both aquaculture and wild catch [20,21] and is characterized by high fecundity and a tolerance to a broad range of salinities and pH [20,21]. O. urolepis is predominantly estuarine, a habitat that is globally noted as a hotspot for MP pollution [22]. With a broad feeding regime, from planktonic to omnivores [23], their chance of interacting with food and non-food materials, including microplastics, is high. In the present study, we exposed 240 fish larvae to four treatments of 0 (control), 1, 10 and 100 PE MPs/mL in triplicate for 65 d, followed by 60 d depuration. The small intestines were analyzed for the presence of PE MPs indicating ingestion, retention and histomorphological changes following exposure and depuration phases. The PE MPs concentrations employed were lesser than those used by some studies [10,24] and in line with marine aquatic environmental concentrations, as previously described [25].
2. Materials and Methods
2.1. Fish Husbandry
A total of 30 Oreochromis urolepis adult fish (20 females and 10 males) were obtained from Kunduchi Campus Aquaculture facilities located on the premises of the School of Aquatic Sciences and Technology, University of Dar es Salaam. The fish were introduced into concrete tanks of approximately 6.70 m3 filled with freshwater up to 80% of its volume. Upon acclimatization for 24 h, fish were fed twice a day to satiation with 4 mm pellets of commercial feed (De Heus, Lot A4, Vinh Long province, Vietnam). After two months of feeding and natural mating, the mouth broods of female fish were inspected, fertilized eggs were taken and transferred to an indoor hatchery containing sand, water filter and sterilized self-circulating water systems. Five days post hatching, larvae (fries), each with an average weight of 0.01 g and a mean length of 1.10 cm, which had exhausted their food reserve in their yolk, were randomly picked for experiment.
2.2. PE MPs Characteristics and Preparation
The Fluorescent Green Polyethylene Microspheres of 1.00 g/cc density with a product ID of UVPMS-BG in 5 g were purchased from Cospheric LLC (Santa Barbara, CA 93160, USA, order # 117025; certificate number 465). The manufacturer’s certificate showed that particle sizes of the PE MPs ranged from 22 μm to 49 μm and had a mean diameter of 42.00 ± 3.20 μm, sold as 38–45 μm diameter on the container. One gram of PE MPs contained 2.67 × 107 particles. Using fluorescent microscope and a Sedgwick rafter counter, it was confirmed that 0.0012 g in a suspension of 200 mL of distilled water with 0.1% surfactant contained approximately 145 PE MPs/mL, relatively similar to the derived value of 156 PE MPs/mL based on the manufacturer’s information. Tween 80 surfactants (product number: P8074) supplied by Sigma-Aldrich (Sigma-Aldrich, 3050 Spruce Street, Saint Louis, MO 63103, USA) were used to disperse PE MPs in suspension.
Suspensions of PE MPs were made by mixing 20 mL of distilled water, 0.1% of Tween 80 Bio-compatible surfactant with PE MPs of 0.0016 g, 0.0154 g and 0.1539 g, equivalent to 1, 10 and 100 PE MPs/mL, respectively. The suspensions were thoroughly mixed with a shaker for an hour at 150 rpm.
2.3. Fish Exposure to and Recovery from PE MPs
A total of 240 fish larvae were randomly selected from the hatchery and rinsed with distilled water. Fish larvae were then transferred into aquaria filled with filtered water containing 1, 10, 100 PE MPs/mL and control groups (no added MPs) in triplicate. Each aquarium had a size of 100 L and was filled with 40 L dechlorinated freshwater filtered through a series of 10, 5 and 1 μm filters. Fish larvae were acclimated without feed for 24 hrs. From the second day, fish larvae were fed with powdered feed at 10% of their body weight in three portions for a week, then crumbles at 8% of their body weight for three weeks. From the fifth to sixth weeks, and then the seventh week onwards, 0.5 mm and 0.8 mm juvenile pellets, respectively, were supplied, and fish were fed at 5% of their body weight twice a day. Commercial feeds (De Heus, Lot A4, Vinh Long province, Vietnam) were used throughout the experiment. Eighty percent of the water was renewed every two days, followed by the addition of PE MPs into the respective aquaria. Exposures lasted for 65 d. On day 65, two juvenile fish were picked from each aquarium for analysis (n = 6 per treatment group), while the remaining fish were rinsed with filtered freshwater and transferred into clean aquaria for 60 d depuration. During this period water renewal continued as previous, but no PE MPs were added. In both exposure and depuration phases, aeration was maintained throughout. Water temperature, pH, oxygen, and ammonia were measured regularly. Throughout the experiment, temperature ranged between 25.6 °C and 31.4 °C and pH between 7.51 and 8.61, which lies within the optimal growth range of fish. Minimum oxygen recorded was 3.32 mg/L, while maximum ammonia recorded was 0.18 mg/L.
2.4. Examination of PE MPs Ingestion and Retention
After 65 d exposure and 60 d depuration periods, two fish from each replicate were sacrificed by euthanizing with 400 mg/L tricaine (MS 222) and washed with ethanol to remove PE MPs particles from the skin. Fish were culled and half the length of the small intestine (covering the duodenum and part of the jejunum) were placed into a glass bottle and then digested with 10 mL of KOH (10% w/v) for 72 h in a water bath at 40 °C [26]. The digested small intestine was then examined under a fluorescent microscope for the presence and quantification of PE MPs.
2.5. Intestinal Histomorphological Change Examination
The section of the small intestine covering the jejunum and ileum was fixed with 10% buffered formaldehyde for 48 h, then processed for histomorphological analysis. Briefly, tissues were first dehydrated in increasing alcohol concentrations (70, 80, 90 and 100%), then cleared with xylene, and lastly infiltrated with paraffin wax. Intestines were sectioned from the embedded blocks at 5 μm with a microtome. Sections were transferred into a hot water bath (40 °C) and then mounted on microscopic slides. The slides were then hydrated in a series of descending alcohol concentrations (100, 90, 80 and 70%) at an interval of 2 min, and later rinsed with running water for 2 minutes. The samples were stained with hematoxylin for 15 min and washed with running water before immersing in acid ethanol for 5 seconds. The samples were again washed in a series of running water, alkaline water (lithium carbonate) and then running water. Later, the samples were stained with eosin dye for 2 min. Thereafter, samples were immersed in a series of ethanol (95, 95, 100 and 100%) at an interval of 2 min. Finally, samples were cleaned twice by xylene each for 2 min and covered by coverslip using pre-mount solution [10].
The examination of histomorphological lesions followed standard hematoxylin and eosin techniques [10,27], where reaction patterns observed were grouped into (1) circulatory disturbances, (2) inflammation and (3) regressive changes. The first two categories are the responses of blood cells (congestion of blood in blood capillaries) and leucocytes infiltration to the surrounding tissues as an inflammatory defensive mechanism. Regressive changes are the architectural and structural alterations in villi cells (crack), crypt glandular cells (damage) and goblets (reduction due to death), as well as epithelial cell degeneration. To evaluate the degree of changes and damages to the small intestine, reaction indices were computed (Table 1 and Table 2), as proposed by Bernet et al. [27]. Each alteration was assigned an importance factor depending on its histomorphological importance (Table 1). The degree and the extent of alteration was assessed using score values (Table 2). The organ and reaction pattern indices were calculated as the sum of multiplied importance factors and score values of all changes within an examined organ and reaction pattern, respectively.

Table 1.
Pathological importance of histomorphological lesion in small intestine of O. urolepis.
Table 2.
Histomorphological alteration assessment of small intestine of O. urolepis.
2.6. Statistical Analyses
Data for small intestinal ingestion and retention of PE MPs were not normally distributed, hence they were square root transformed. After conforming to normality and homoscedasticity tests, a One-Way ANOVA with post hoc Tukey HSD test was performed between treatment groups. For villi height, width and epithelial cell height, the Kruskal–Wallis test followed by pairwise comparison was employed after data deviating from normality. An evaluation of whether the frequency of occurrence of PE MPs in sections of digested small intestines and lesion indices depended on exposure concentrations was carried out using a 6 × 3 contingency table and Goodness-of-fit test, respectively, and tested with Chi-square test. The difference was considered statistically significant at p ≤ 5%. All statistical analyses were performed using the SPSS statistical software package (IBM SPSS Statistics, version 23).
3. Results
3.1. Ingestion and Retention of PE MPs after Exposure and Recovery Phase
The examination of fish juveniles indicated the presence of PE MPs in the small intestines of all PE MP exposed treatment groups (Figure 1), but not the control group. Their prevalence was significantly proportional to the PE MPs exposure concentration (χ2 = 49.54; df = 10; p < 0.05). The mean concentration of PE MPs particles retrieved were 3.6 ± 2.7, 13.5 ± 8.2 and 223.7 ± 110.2 PE MPs/mL (n = 6) corresponding significantly (to the exposure concentration of 1, 10 and 100 PE MPs/mL, respectively (one-way ANOVA, Tukey HSD, p < 0.05, Figure 2). After depuration, no PE MPs were observed in the digested small intestines of the fish of either treatment group.
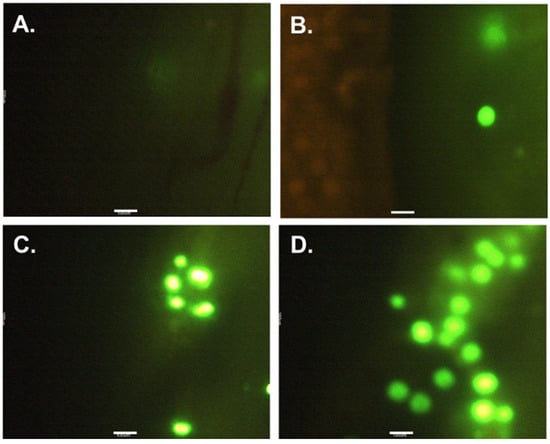
Figure 1.
Photos displaying PE MPs in longitudinal section of small intestines of O. urolepis at different treatment groups: (A) Control (no PE MPs); (B) 1 PE MPs/mL; (C) 10 PE MPs/mL; (D) 100 PE MPs/mL. Note: Scale bar = 100 μm.
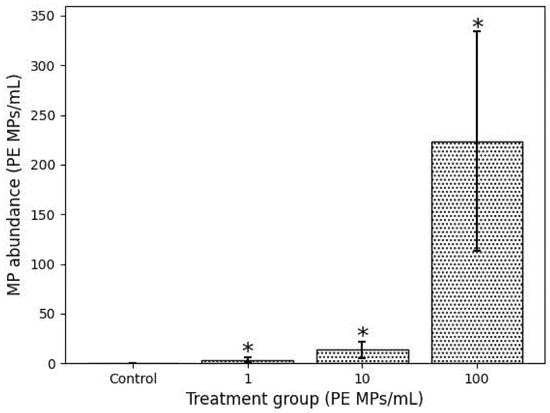
Figure 2.
Mean (±SD) concentration of PE MPs recovered from digested small intestine of O. urolepis juveniles after 65 d exposure. After recovery period, no PE MPs observed in any digested intestine. Asterisks (*) denote significant difference of treatment group from other groups (One way ANOVA, p < 0.05).
3.2. Small Intestinal Histomorphological Changes after Exposure and Depuration Phase
Generally, the small intestine histomorphological lesion indices (Figure 3A) showed a significant increase in frequency of occurrence of deterioration in the small intestines of fish, corresponding to PE MPs exposure dose (χ2 = 47.37; df = 2; p < 0.05). After the recovery (depuration) phase, the frequency of occurrence of small intestine histomorphological lesion indices declined but remained significantly prevalent between treatment groups (χ2 = 12; df = 2; p < 0.05).
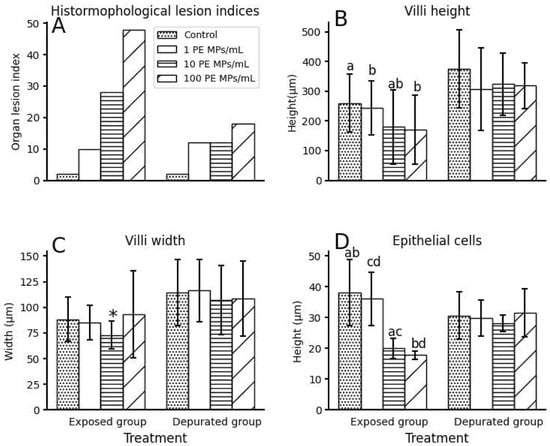
Figure 3.
Histomorphological lesions of small intestines of O. urolepis after PE MPs exposure and depuration (recovery) phases: (A) Organ indices; (B) Villi height (±SD); (C) Villi width (±SD); (D) Epithelial cells height (±SD). Similar letters on the top of error bar denote significant different across treatment groups, while asterisk denote significant difference of 10 PE MPs/mL from other treatment groups (Kruskal–Wallis post hoc pairwise comparison, p < 0.05). Note: SD = standard deviation.
The mean primary villi height of fish between treatment groups varied significantly inverse to PE MPs exposure dose (Kruskal–Wallis, p < 0.05; Figure 3B). Between treatment groups, a significant variation was found between 1 PE MPs/mL with 10 PE MPs/mL and 100 PE MPs, and the control with 10 PE MP/mL only (post hoc pairwise comparison, p < 0.05). The villi width followed a similar trend as villi height (Kruskal–Wallis, p < 0.05; Figure 3C). Post hoc pairwise comparison revealed significant variation between 10 PE MPs/mL and other treatment groups only (p < 0.05). The frequency of occurrence of villi lesion indices (combined villi height and width) increased significantly with PE MPs exposure dose (χ2 = 8.27; df = 2; p < 0.05). After the depuration phase, villi heights of all treatment groups were relatively similar, except for the control, which were the longest (Figure 3B). Villi widths of the control and 1 PE MPs/mL were slightly broader than those of 10 and 100 PE MPs/mL, which were comparable. Both villi heights and widths, however, were not statistically different between treatment groups (Kruskal–Wallis, p > 0.05). The villi recovery was very high, as shown by insignificant occurrence of villi damage indices (χ2 = 2; df = 2; p > 0.05).
Simple columnar epithelium lining in the small intestinal mucosal layer of the control group fish consisted of round to oval nuclei located towards the basement membrane, while those of 100 PE MPs/mL were lined by simple columnar to cuboidal epithelium (Figure 4). Fish exposed to 100 PE MPs/mL were most severely damaged and exhibited pale cytoplasm and pyknoic nucleus (Figure 4). In fact, epithelial cells lesion indices showed a significant increase in frequency of damage with increased PE MPs dose (χ2 = 6.67; df = 2; p < 0.05; Figure 5B). Thus, those from the 10 PE MPs exposure group were affected moderately, indicating clear cytoplasm and condensed nucleus (Figure 6), in comparison to the control group (Figure 7). The means epithelial cells height (Figure 3D) between treatment groups differed significantly (Kruskal–Wallis, p < 0.05) and were inversely related to PE MPs exposure concentration. The post hoc pairwise comparison revealed two sets; one consisted of the control and 1 PE MPs/mL groups that differed significantly from 10 and 100 PE MPs/mL groups (p < 0.05). On the contrary, depurated juvenile fish revealed high degree of epithelial cells recovery with slight variation (Figure 3D and Figure 8) in height, which resulted in insignificant change between treatment groups (Kruskal–Wallis, p > 0.05), as supported by epithelial cells lesion indices (χ2 = 2; df = 2; p > 0.05; Figure 5B).
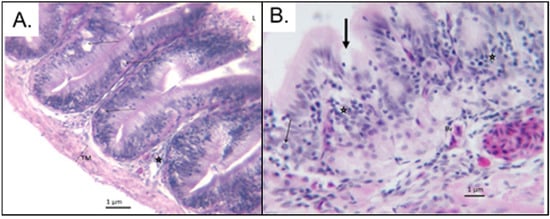
Figure 4.
(A) Photomicrograph of treated O. urolepis at 100 PE MPs/mL dose. Numerous degenerating epithelial cells (arrows) were observed. Note infiltrations of leukocytes (asterisk) in the lamina propria. L: lumen; TM: Tunica muscularis. (B) A higher magnification photomicrograph of treated fish at 100 PE MPs/mL dose. Thin arrow: crypt gland cell; thick arrow: degenerating epithelial cell; asterisks: leucocytic infiltration in the lamina propria; Bv: congested blood vessel.
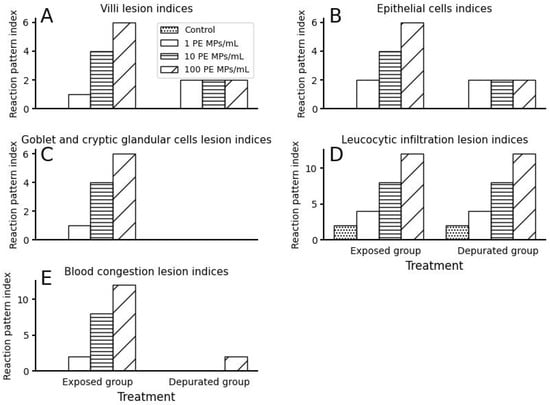
Figure 5.
Reaction pattern lesions indices of small intestines of O. urolepis after PE MPs exposure and depuration (recovery) phases: (A) Villi lesion indices; (B) Epithelial cells lesion indices; (C) Goblet and cryptic glandular cells lesion indices; (D) Leucocytic infiltration lesion indices; (E) Blood congestion lesion indices.
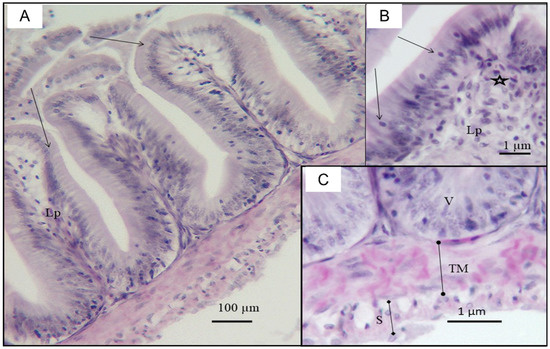
Figure 6.
Photomicrographs of the small intestine from O. urolepis exposed to 10 PE MPs/mL. (A) Note the decrease in the height of villi (arrows) and infiltration of inflammatory cells in the lamina propria (Lp); (B) Portion of villi showing degenerating epithelial cells (arrows), leucocytic infiltration (asterisk) in the lamina propria (Lp); (C) Congested capillaries in the tunica muscularis (TM); S: serosa; V: basal region villi.
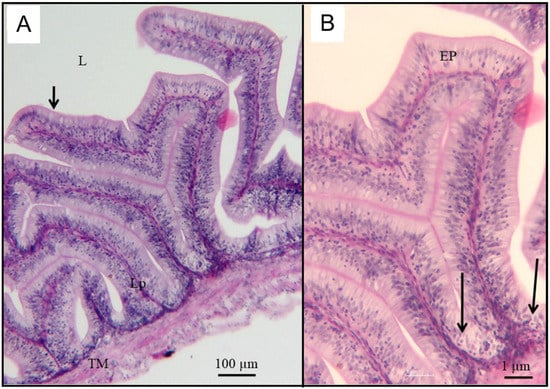
Figure 7.
Photomicrographs from a small intestine of the control O. urolepis during the exposure phase. (A) Primary villi (arrow) radiating towards the lumen (L). Lp: lamina propria; TM: tunica muscularis. (B) A higher magnification photomicrograph of a villus. Note the simple columnar epithelium (Ep) lining the villi.
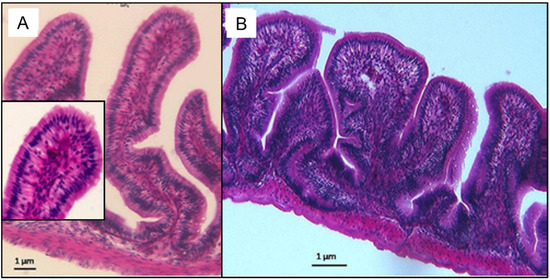
Figure 8.
Photomicrograph of the small intestines of O.urolepis following depuration period of 60 days. Note the similarity between the villi of the control group exposure (A) from the 100 PE MPs/mL exposure group (B).
Cryptic glandular cells and goblet cells were moderately affected in the 100 PE MPs/mL and 10 PE MPs/mL treatment groups compared to the control group (Figure 4 and Figure 6). Mild effects were observed in the 1 PE MPs/mL exposure group. In the control group, goblet cells appeared normal (Figure 7). Generally, the frequency of occurrence of reaction lesion indices significantly depended on PE MPs exposure dose (χ2 = 8.27; df = 2; p < 0.05; Figure 5C). Upon depuration, the goblet and cryptic glandular cells showed recovery and were indistinguishable between treatment groups (Figure 5C and Figure 8).
Leucocytic infiltration is one of the inflammatory responses that show tissue injury. Fish in the control group revealed mild scattered leucocytic cells (Figure 7A). Severe leucocytic cells infiltration was found in both 10 and 100 PE MPs/mL fish. The leucocytes infiltration lesion indices (due to leucocytes infiltration) indicated significant frequency of occurrence of inflammation in intestinal tissue fluid that diminished as the PE MPs dose decreased (χ2 = 9.08; df = 2; p < 0.05; Figure 5D). This was the same across all treatment groups, even after the recovery phase (χ2 = 9.08; df = 2; p < 0.05).
Intestinal tissue injury triggered the congestion of blood in the lamina propria, propria submucosa and the tunica muscularis of the intestinal walls proportional to PE MPs exposure dose, as indicated by the significant variation in the frequency of occurrence of blood congestion (circulatory) lesion indices (χ2 = 16.55; df = 2; p < 0.05; Figure 4E). Such effects diminished sharply in depurated juvenile fish, with only a mild occurrence in 100 PE MPs/mL treatment group individuals (χ2 = 6; df = 2; p < 0.05). No blood congestion was observed in the control juvenile fish at both experimental set ups (Figure 8A).
4. Discussion
African freshwaters are increasingly documented as being impacted by plastic and MP pollution. Population growth and poor waste management practices have contributed to this [1]. As a result, the recovery of ingested MPs from the gastrointestinal tract of wild freshwater fish is becoming common and prevalent [4,5]. Fish close to river mouths or plastic sources are more vulnerable to MP ingestion [1,6]. Relatively few studies have reported MPs retrieval, yet a knowledge gap persists on the effects of exposure and depuration. Our results showed that the long-term exposure of O. urolepis to PE MPs impaired the histomorphology of the small intestine in correspondence to the PE MPs concentration (Figure 3, Figure 4, Figure 5 and Figure 6). After prolonged depuration, most fish recovered but not fully, particularly those subjected to high PE MPs concentration. Our findings are supported by previous studies where exposure to MPs caused significant histopathological damage. The impairment of intestinal structures and functions in a dose–response manner was also reported in Oreochromis niloticus [28], Artemia pathenogenetica [29], Girella laevifrons [10] and Danio rerio [11]. The severity of the damage in depurated silver carp fish persisted after the loss of MPs [12], similar to our study. The study with silver carp, however, was an acute exposure and provided a limited snapshot of anticipated chronic effects. Gilthead seabream depurated for 30 d revealed full recovery of intestinal biomarker levels [13]. Again, the depuration time used was shorter than the current study and the study reported on physiological impacts rather than histopathological changes, which are likely to stay longer. Nevertheless, together, such studies provide a weight of evidence that MPs can cause intestinal damage after the removal of MPs from the GI tract and that the damage can be long-lasting.
The presence of MPs in small intestinal tracts of the exposed fish juveniles and their subsequent disappearance after depuration imply that fish subjected to MPs polluted habitats can ingest and egest MPs, the magnitude of which depends on the prevailing concentration in the surrounding environment. O. niloticus, a sister species of O. urolepis, exposed to 1, 10, 100 μg/L of 0.1 um polystyrene MPs, both ingested and retained amounts of MPs corresponding to the exposure concentration, although depuration was not carried out in that study [28]. Pitt et al. [30] reported a relationship between ingestion and depuration after offering zebrafish (Danio rerio) a concentration range of polystyrene nanoparticles. Both experiments were conducted with a short experimental timeframe compared to the present study. In prolonged exposure periods, as with studies with common carp [14] and gilthead seabream [13], the ingestion-depuration kinetics were similar to the short-term scenario. The implication of chronic exposure to the high quantity of MPs may have delayed depuration, extending threat to organisms and delayed recovery of injured tissues. The size and shape (i.e., fiber or bead) of MPs may further influence retention and depuration kinetics. For example, Pomacentrus amboinensis accumulated a significant quantity of MPs and showed delayed depuration when exposed to polyethylene terephthalate MPs (length 600–700 μm), compared to polypropylene (length 125–250 μm) at similar concentrations [31]. However, if the retention of larger MPs sizes, as reported in wild O. niloticus from Lake Victoria (>500 μm) [4], is coupled with increased intestinal concentrations, as described in the current study, the result may impact the feeding behavior and nutritional status of affected fish.
Against expectation, our study showed that observations of degenerated villi width were proportional to PE MPs dose concentration contrary to beheaded villi and degenerated epithelial cells height (Figure 3B–D). This could be explained by the repair of continuously worn-out villi and epithelial cells by MPs, resulting in swelling and thickening of villi width in more affected tissues. The significant decline in villi width in 10 PE MPs/mL compared to the rest of the treatments might be attributed to the energy balance to offset damaged villi and epithelial cells height. Upon depuration, a minor variation remained. The shortening and thickening in villi width was also displayed by G. laevifrons [10] and D. labrax [9] with respect to MPs exposure dose, despite lacking an explanation.
The duration and capacity of tissue or organs lesion recovery depends on the severity of injury and the type of histological alteration [27]. While recovery may occur soon for moderately affected tissues, it may persist or become irreversible due to severe damage, even post-removal of pollutants. G. laevifrons exposed to high MPs concentration revealed a severe effect on leucocytic infiltration and hyperemia compared to goblet and cryptic cell loss and villi cell loss [10], similar to this study. Ouyang et al. [14] and Qiao et al. [32] highlighted the persistence of leucocytic infiltration in lamina propria and tunica muscularis in fish small intestines following the depuration phase, consistent with this study (Figure 5D), which largely accounted for overall high small-intestine histomorphological lesion indices (Figure 3A). As a prominent line of defense to counter foreign body invasion, their magnitude mirrors scale of effect [10]. The leucocytes infiltration observed in control was temporary, likely induced by damage during peristaltic movement of chyme. Hamed et al. [33] revealed persistence in leucocytic infiltration post-depuration in O. tilapia. This situation can be further linked to microbiota dysbiosis in intestines, as described in zebrafish [32,34]. In particular, damage of the gut reduced the abundance of Actinobacteria and increased the abundance of pathogenic bacteria [14], which weakened the function of the intestinal barrier and increased sensitivity to the immune response, triggering leucocytic infiltration. Consistent to the current study, leucocytic infiltration was high after the depuration phase (Figure 5D) and accounted largely for the overall high small-intestine histomorphological lesion indices (Figure 3A). Further delay in the full recovery of villi and epithelial lesions, as indicated by their indices (Figure 5A,B), might contribute to the situation.
Indiscriminate or unselective feeding is the main pathway through which fish, particularly in their early stages, ingest, microplastics [35,36]. As fish grow, their mode of feeding may change from herbivores, as exhibited by the majority of fish larvae, to either entirely carnivores or omnivores. Of the different feeding modes, omnivores have been shown to accumulate MPs more than either herbivores or carnivores [37,38]. As such, omnivorous species have been shown to suffer significant impacts from MP ingestion in terms of both histological and physiological endpoints, as demonstrated by Oreochromis niloticus [33,39] and Hypophthalmichthys molitrix [12], after exposure and depuration phases. Similar to the current study, exposure to MPs indicated a dose–effect response, of which histomorphological damage remained longer in fish in 10 and 100 PE MPs/mL treatments, even after being depurated. Arguably, other factors such as species-specificity, size and shape of fish, and size and shape of MPs should be considered, as they may affect the scale of damage and, subsequently, delay recovery [35].
Fish at their juvenile stage consume an enormous amount of food to offset catabolism (energy production) and anabolism (growth) demand. In good food supply, the efficiency of digestion and assimilation depends on health of small intestines. The acquired nutrients, together with oxygen, enhances the rapid growth of fish juveniles for quick adaptations to changing environments, reaching sexual maturity, avoiding predators and finding more food [40]. A decline in food uptake and assimilation due to MP-induced deteriorations of the small intestines may pose a great threat to juvenile fish, and, subsequently, adult fish if the damage is not reversed. The majority of studies investigating intestinal damage have used either already developed and prolonged acclimated fish [41,42] or used larvae in the short term [43]. However, in this study we elucidated the significant effects on fish larvae from the outset of feeding over a prolonged exposure period, which continued well after the removal of MPs from the environment and from the intestinal tract.
5. Conclusions
The presence of MPs in African freshwater habitats and ingestion by fish has been documented, as has the important role of the fish as a food source and economic commodity. The intestinal damage described in this study is the first for a fish species from African waters, but in line with other species world-wide. Our findings revealed that the MP-induced damage in the small intestine of O. urolepis can remain long after removal of the contaminant. The persistence of damage may impair food digestion and nutrient availability to fish, jeopardizing energy availability for growth and development to adult stages and subsequent reproduction and food security at large. Further investigation describing the interplay between damage with other physiological processes such as growth and reproduction are crucial.
Author Contributions
Conceptualization, J.J.M., D.A.S. and F.R.K.; Methodology, J.J.M. and D.A.S.; Software, J.J.M.; Validation, J.J.M., D.A.S. and D.J.; Formal analysis, J.J.M. and W.K.; Investigation, J.J.M. and D.A.S.; Resources, J.J.M., J.F.M. and D.A.S.; Data curation, J.JM. and F.R.K.; Writing—original draft preparation, J.J.M.; Writing—review and editing, F.R.K. and D.A.S.; Visualization, D.A.S. and W.K.; Supervision, D.A.S., F.R.K. and J.F.M.; Project administration, D.A.S. and D.J.; Funding acquisition, D.A.S., F.R.K. and D.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by WIOMSA under MASMA MICROMARINE project number MASMA/OP/2018/03 and URT Ministry of Education, Science and Technology.
Institutional Review Board Statement
The animal study protocol was approved by the Office of Deputy Vice Chancellor Research of UNIVERSTIY OF DAR ES SALAAM.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
My sincere gratitude to Michael and Richard Masinde for technical support in fish husbandry and lab works. I extend my thanks to University of Dar es Salaam through School of Aquatic Sciences and Fisheries Technology for hosting my study and University of Dodoma, my employer for the study leave permission.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khan, F.R.; Mayoma, B.S.; Biginagwa, F.J.; Syberg, K. Microplastics in inland African waters: Presence, sources, and fate. Freshw. Microplastics 2018, 58, 101–124. [Google Scholar]
- Shilla, D. Status updates on plastics pollution in aquatic environment of Tanzania: Data Availability, Current Challenges and Future Research Needs. Tanzan. J. Sci. 2019, 45, 101–113. [Google Scholar]
- Onyango, P. Socio-economic characteristics of the Lake Victoria Fisheries. In Lake Victoria Fisheries Resources; Springer: Berlin/Heidelberg, Germany, 2017; Volume 93, pp. 161–184. [Google Scholar]
- Biginagwa, F.J.; Mayoma, B.S.; Shashoua, Y.; Syberg, K.; Khan, F.R. First evidence of microplastics in the African Great Lakes: Recovery from Lake Victoria Nile perch and Nile tilapia. J. Great Lakes Res. 2016, 42, 146–149. [Google Scholar] [CrossRef]
- Khan, F.R.; Shashoua, Y.; Crawford, A.; Drury, A.; Sheppard, K.; Stewart, K.; Sculthorp, T. ‘The Plastic Nile’: First Evidence of Microplastic Contamination in Fish from the Nile River (Cairo, Egypt). Toxics 2020, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Merga, L.B.; Redondo-Hasselerharm, P.E.; Van den Brink, P.J.; Koelmans, A.A. Distribution of microplastic and small macroplastic particles across four fish species and sediment in an African lake. Sci. Total Environ. 2020, 741, 140527. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hoh, E.; Kurobe, T.; Teh, S.J. Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci. Rep. 2013, 3, 3263. [Google Scholar] [CrossRef]
- Kashiwada, S. Distribution of nanoparticles in the see-through medaka (Oryzias latipes). Environ. Health Perspect. 2006, 114, 1697–1702. [Google Scholar] [CrossRef] [Green Version]
- Pedà, C.; Caccamo, L.; Fossi, M.C.; Gai, F.; Andaloro, F.; Genovese, L.; Perdichizzi, A.; Romeo, T.; Maricchiolo, G. Intestinal alterations in European sea bass Dicentrarchus labrax (Linnaeus, 1758) exposed to microplastics: Preliminary results. Environ. Pollut. 2016, 212, 251–256. [Google Scholar] [CrossRef]
- Ahrendt, C.; Perez-Venegas, D.J.; Urbina, M.; Gonzalez, C.; Echeveste, P.; Aldana, M.; Pulgar, J.; Galban-Malagon, C. Microplastic ingestion cause intestinal lesions in the intertidal fish Girella laevifrons. Mar. Pollut. Bull. 2020, 151, 110795. [Google Scholar] [CrossRef]
- Lei, L.; Wu, S.; Lu, S.; Liu, M.; Song, Y.; Fu, Z.; Shi, H.; Raley-Susman, K.M.; He, D. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans. Sci. Total Environ. 2018, 619–620, 1–8. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, J.; Pan, Z.; Wang, S.; Zhang, L.; Wang, Q.; Ye, Q.; Zhou, A.; Xie, S.; Zeng, F.; et al. A dosage-effect assessment of acute toxicology tests of microplastic exposure in filter-feeding fish. Fish Shellfish Immunol. 2021, 113, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Solomando, A.; Capo, X.; Alomar, C.; Alvarez, E.; Compa, M.; Valencia, J.M.; Pinya, S.; Deudero, S.; Sureda, A. Long-term exposure to microplastics induces oxidative stress and a pro-inflammatory response in the gut of Sparus aurata Linnaeus, 1758. Environ. Pollut. 2020, 266 Pt 1, 115295. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, M.Y.; Feng, X.S.; Li, X.X.; Wen, B.; Liu, J.H.; Huang, J.N.; Gao, J.Z.; Chen, Z.Z. Microplastics intake and excretion: Resilience of the intestinal microbiota but residual growth inhibition in common carp. Chemosphere 2021, 276, 130144. [Google Scholar] [CrossRef]
- Calil, J.; Gutiérrez-Graudiņš, M.; Munguía, S.; Chin, C. Neglected: Environmental Justice Impacts of Marine Litter and Plastic Pollution; Technical Report; UN Environment (UNEP): Nairobi, Kenya, 7 April 2021. [Google Scholar]
- Alessi, E.; Di Carlo, G.; Di Carlo, E.B.; Campogianni, S. Out of the Plastic Trap: Saving the Mediterranean from Plastic Pollution; WWF Report: Rome, Italy, 2018. [Google Scholar]
- URT. Regulation on Prohibition of Plastics Carrier Bags; Government of United Republic of Tanzania Mainland: Dodoma, Tanzaniam, 2019; Volume 394. [Google Scholar]
- Directive, E. Reduction of the impact of certain plastic products on the environment. Off. J. Eur. Union 2019, L155. Available online: https://www.eumonitor.eu/9353000/1/j9vvik7m1c3gyxp/vkzb7kgzwlzz (accessed on 12 June 2019).
- Maione, C. Quantifying plastics waste accumulations on coastal tourism sites in Zanzibar, Tanzania. Mar. Pollut. Bull. 2021, 168, 112418. [Google Scholar] [CrossRef] [PubMed]
- Chale, F. Studies on the fisheries and biology of Oreochromis urolepis (pisces: Cichlidae) in the Mtera reservoir (Tanzania). Tanzan. J. Sci. 2004, 30, 33–39. [Google Scholar]
- Mapenzi, L.L.; Mmochi, A.J. Effect of stocking density on growth performance of hybrid of Oreochromis niloticus and Oreochromis urolepis urolepis in saline water. WIO J. Mar. Sci. 2016, 15, 67–74. [Google Scholar]
- Zhang, K.; Xiong, X.; Hu, H.; Wu, C.; Bi, Y.; Wu, Y.; Zhou, B.; Lam, P.K.; Liu, J. Occurrence and Characteristics of Microplastic Pollution in Xiangxi Bay of Three Gorges Reservoir, China. Environ. Sci. Technol. 2017, 51, 3794–3801. [Google Scholar] [CrossRef]
- Popma, T.; Masser, M. Tilapia Life History and Biology. South. Reg. Aquac. Cent. 1990, 283, 4. [Google Scholar]
- Khan, F.R.; Syberg, K.; Shashoua, Y.; Bury, N.R. Influence of polyethylene microplastic beads on the uptake and localization of silver in zebrafish (Danio rerio). Environ. Pollut. 2015, 206, 73–79. [Google Scholar] [CrossRef]
- Wang, W.; Gao, H.; Jin, S.; Li, R.; Na, G. The ecotoxicological effects of microplastics on aquatic food web, from primary producer to human: A review. Ecotoxicol. Environ. Saf. 2019, 173, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Karami, A.; Golieskardi, A.; Choo, C.K.; Romano, N.; Ho, Y.B.; Salamatinia, B. A high-performance protocol for extraction of microplastics in fish. Sci. Total Environ. 2017, 578, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Hom, P.; Wahhli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Zhang, S.; Razanajatovo, R.M.; Zou, H.; Zhu, W. Accumulation, tissue distribution, and biochemical effects of polystyrene microplastics in the freshwater fish red tilapia (Oreochromis niloticus). Environ. Pollut. 2018, 238, 1–9. [Google Scholar] [CrossRef]
- Wang, Y.; Mao, Z.; Zhang, M.; Ding, G.; Sun, J.; Du, M.; Liu, Q.; Cong, Y.; Jin, F.; Zhang, W.; et al. The uptake and elimination of polystyrene microplastics by the brine shrimp, Artemia parthenogenetica, and its impact on its feeding behavior and intestinal histology. Chemosphere 2019, 234, 123–131. [Google Scholar] [CrossRef]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, tissue distribution, and toxicity of polystyrene nanoparticles in developing zebrafish (Danio rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef]
- Santana, M.F.M.; Dawson, A.L.; Motti, C.A.; van Herwerden, L.; Lefevre, C.; Kroon, F.J. Ingestion and Depuration of Microplastics by a Planktivorous Coral Reef Fish, Pomacentrus amboinensis. Front. Environ. Sci. 2021, 9, 1–16. [Google Scholar] [CrossRef]
- Qiao, R.; Deng, Y.; Zhang, S.; Wolosker, M.B.; Zhu, Q.; Ren, H.; Zhang, Y. Accumulation of different shapes of microplastics initiates intestinal injury and gut microbiota dysbiosis in the gut of zebrafish. Chemosphere 2019, 236, 124334. [Google Scholar] [CrossRef]
- Hamed, M.; Soliman, H.A.M.; Badrey, A.E.A.; Osman, A.G.M. Microplastics induced histopathological lesions in some tissues of tilapia (Oreochromis niloticus) early juveniles. Tissue Cell 2021, 71, 101512. [Google Scholar] [CrossRef]
- Qiao, R.; Sheng, C.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics induce intestinal inflammation, oxidative stress, and disorders of metabolome and microbiome in zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Liu, Y.; Qiu, X.; Xu, X.; Takai, Y.; Ogawa, H.; Shimasaki, Y.; Oshima, Y. Uptake and depuration kinetics of microplastics with different polymer types and particle sizes in Japanese medaka (Oryzias latipes). Ecotoxicol. Environ. Saf. 2021, 212, 112007. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Sun, M.; Zhou, M.; Chang, Z.; Li, L. Polyvinyl chloride microplastics induce growth inhibition and oxidative stress in Cyprinus carpio var. larvae. Sci. Total Environ. 2020, 716, 136479. [Google Scholar] [CrossRef] [PubMed]
- Mizraji, R.; Ahrendt, C.; Perez-Venegas, D.; Vargas, J.; Pulgar, J.; Ojeda, F.P.; Duarte, C.; Galbán-Malagón, C. Is the feeding type related with the content of microplastics in intertidal fish gut? Mar. Pollut. Bull. 2017, 116, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, J.; Zhou, A.; Ye, Q.; Feng, Y.; Wang, Z.; Wang, S.; Xu, G.; Zou, J. Species-specific effect of microplastics on fish embryos and observation of toxicity kinetics in larvae. J. Hazard. Mater. 2021, 403, 123948. [Google Scholar] [CrossRef]
- Hamed, M.; Soliman, H.A.M.; Osman, A.G.M.; Sayed, A.E.H. Assessment the effect of exposure to microplastics in Nile Tilapia (Oreochromis niloticus) early juvenile: I. blood biomarkers. Chemosphere 2019, 228, 345–350. [Google Scholar] [CrossRef]
- Tran-Duy, A.; Schrama, J.W.; van Dam, A.A.; Verreth, J.A.J. Effects of oxygen concentration and body weight on maximum feed intake, growth and hematological parameters of Nile tilapia, Oreochromis niloticus. Aquaculture 2008, 275, 152–162. [Google Scholar] [CrossRef]
- Batel, A.; Baumann, L.; Carteny, C.C.; Cormier, B.; Keiter, S.H.; Braunbeck, T. Histological, enzymatic and chemical analyses of the potential effects of differently sized microplastic particles upon long-term ingestion in zebrafish (Danio rerio). Mar. Pollut. Bull. 2020, 153, 111022. [Google Scholar] [CrossRef]
- De Sales-Ribeiro, C.; Brito-Casillas, Y.; Fernandez, A.; Caballero, M.J. An end to the controversy over the microscopic detection and effects of pristine microplastics in fish organs. Sci. Rep. 2020, 10, 12434. [Google Scholar] [CrossRef]
- Huang, J.N.; Wen, B.; Meng, L.J.; Li, X.X.; Wang, M.H.; Gao, J.Z.; Chen, Z.Z. Integrated response of growth, antioxidant defense and isotopic composition to microplastics in juvenile guppy (Poecilia reticulata). J. Hazard. Mater. 2020, 399, 123044. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).