1. Introduction
One of V.I. Vernadsky’s—a great Russian and Soviet scientist, mineralogist, biogeochemist, and creator of the biosphere concept, main contributions to biological science, in the development of his doctrine of the biosphere, is that he actually postulated the following thesis: nothing in the science of life makes sense outside the study of this or that phenomenon within the framework of its role in the structural and functional organization of the biosphere. According to V.I. Vernadsky himself, living matter, as the totality of living organisms in the functioning biosphere, exists only in the biosphere [
1,
2].
Vernadsky considered life as a certain property of living substance: living substance (matter) means a substance possessing life that has some inherent properties, just as one speaks of a radioactive substance [
3] (p. 197). At the same time, living matter has systemic properties which influence its structure, and each element of it, each organism, has its own specific properties.
Vernadsky formulated the following three important hypotheses in the early 1920s: living matter is a cosmic phenomenon; living matter is essentially different from inert matter; and there was no beginning of life in the Earth biosphere and all empirical data do not support abiogenesis. The term “biosphere” was not central to this period of his research; Vernadsky, in fact, postulated that the emergence of life on Earth depended on the system, not on a hypothetical primary organism [
4,
5]. Life is a systemic event, not the “private” existence of paleo or more evolutionarily developed organisms.
What does this mean for biological science, in particular? First of all, living things are represented by organisms, one of the most important properties of which is self-reproduction and the transmission of genetic information; however, life in the biosphere as a planetary and cosmic phenomenon is the existence and functioning of bioinert systems. K.M. Khailov [
6] rightly and dualistically posits the following question: what is life and what is life on Earth or—more precisely—life in the biosphere? The emergence of living organisms in the biosphere, capable of endlessly maintaining the ‘circulation’ of their species, was a secondary evolutionary act based on the evolutionary development of primary protoecosystems [
7,
8,
9].
This hypothesis is quite in agreement with the position expressed by V.I. Vernadsky that the biosphere should have emerged immediately as a sufficiently complex system in which the main biospheric functions could be carried out. The maintenance of these functions in the modern biosphere is determined by its characteristic structure. The question about the structure of the biosphere is still debatable. This review, in the authors’ opinion, can make a useful contribution to discussions of the biosphere’s structure and the ways that it developed.
In this article, the main task is to show the hierarchical structure of the biosphere. The authors consider the hierarchy of biospheric subsystems and elements as a system of interconnected levels of organization, which is built on the principle of the interrelation of parts and the whole. At the same time, the authors point out the features of the functional similarity of these parts and the whole, which the authors call functional fractality.
The authors’ familiarity with the problem of biospheric structure and evolution confirms that Western scientists rarely use research from Eastern Europe; therefore, the authors intentionally predominantly used this literature in the discussions to create and maintain some useful informational bridges.
2. The Structure of the Biosphere
The structure of the biosphere is determined by its systemic properties. The authors support V.M. Shimansky’s point of view that ‘the biosphere cannot be limited to only one set of living organisms, as they cannot create a unified system outside the conditions of their existence, nor can all the most powerful organogenic formations that were once part of the biosphere of the past be attributed to the biosphere’ [
10] (p. 6). The integrity of any system is supported by the interaction of its elements and the dynamic replacement of old elements by newer ones; the dialectical connection between the whole and parts really forms the basis of the principle of system morphoprocess [
11]. The biosphere can be considered as one of the variants of the morphoprocess [
12]. On this basis, it becomes extremely important to identify the structure of the biosphere, as well as its specific interacting elements. V.I. Vernadsky was the first to approach this question. If E. Suess, introducing the term ‘biosphere’, considered its main integral feature as it representing one of the geospheres, Vernadsky singled out its main subdivisions, paying attention to the thickening of life in contact zones.
First of all, V.I. Vernadsky paid attention to the hydrosphere—the ocean—where he singled out the photic zone or photosynthesis film. In addition, the bottom film is a reducing zone, where the mineralization of organic matter and recycling of biogens for the production of new organic matter takes place. He also identified a relatively inert intermediate zone. On land, on the scale of the entire biosphere, Vernadsky distinguished only one film of life. These four large subdivisions of the biosphere are called biospheromerons [
13]. Thus, biospheromerons represent the largest structural units of the biosphere (
Figure 1). However, biospheromerons, as part of the biosphere bioinert system, do not directly consist of organisms; the latter are part of communities and are inseparably connected with inert elements of the environment, forming ecosystems.
The biosphere has a hierarchical structure, which is an important consequence of the emergent properties of its elements, which, in turn, also have a complex structure and their own structural elements. Thus, based on the systemic properties of the biosphere, it is possible to conclude that ecosystems are not directly part of the biosphere, but are elements of lower-level (relative to the biosphere) systems. The diversity of ecosystems is enormous, both in the hydrosphere and in atmobiont conditions, which is a consequence of wide ecological divergence. This is one of the areas of ecological diversogenesis, the formation of biodiversity characteristics of a particular state of the biosphere, stage of its development, and evolution. However, the analysis of the composition of the biosphere elements forces us to pay attention to the dialectical connection between broad ecological divergence and convergence. In addition to diversity and differences in structure, there is another regularity: the ‘general structure’ of ecosystems is quite typical; in similar conditions (similarity of geome (GEO)), the biotic structures at the supra-organismal level are quite convergent (similarity of biome (BIO)).
Considering the biosphere as a complex bioinert system and as a morphoprocess, it is very important to clarify its structure. This can be realized either in the form of a structural–functional fundamental model or by examining the real individual elements of the system and the connections between them. There are concepts suggesting that the biosphere represents the ‘largest ecosystem’ [
14]; that is, if consider the biosphere as a single real whole, then its fragments are also ecosystems—only smaller in size and possessing the same properties. This approach is an absolute extension of the principle of L. Evans [
15], who considered as ecosystems any bioinert systems in which there is an interaction between inert elements of the environment and living organisms. According to I. Zhirkov [
16], this approach, or the concept of the continuum, dominates current views on the structure of the biosphere. The continuum paradigm was formulated by L. Ramensky (1924) [
17] and H. Gleason (1926) [
18]. (As noted [
19], already in 1910, at the 12th Congress of Russian Naturalists and Physicians, L. Ramensky expressed the main provisions of the continualistic concept). However, it should be noted that even earlier (in 1918), the Danish hydrobiologist K. Petersen [
20] also considered marine benthos communities as continuum systems—in opposition to the concept of K. Moebius (1877) [
21].
According to the concept of continualism, species populations form associations, or ensembles, in accordance with the coincidence of individual responses of each species to a given combination of environmental conditions. These associations of populations gradually transition from one to another, creating a continuum of the Earth’s living cover. Another approach, or concept, is structuralism, which is based on the works of F. Clements [
22]. According to this concept, the populations of the planet have an endogenous structure, the elements of which are more or less clearly limited to discrete cenotic structures. Species respond to changes in environmental conditions not only individually, but also as part of integral biocenoses. Sukachev V.N. [
23] believed that the living cover of the Earth has a structure based on discrete units, or biogeocenoses.
It is obvious that strict adherence to one concept or the other leads us down a non-dialectical path. The classical ecological approach offers an alternative path. Many real ecosystems have their own rather narrow range of conditions for existence, and if they are placed in the coordinate system “Precipitation–Temperature”, a characteristic field of points is formed, resembling a triangular shape. However, this field of points can be differentiated, as E. Odum [
24] and R. Ricklefs [
25] did, into a number of areas within which ecosystems exhibit important features of similarity.
Already at the beginning of the 19th century, it was generally formulated that the Earth’s surface is divided into areas with different biota. In particular, Kafanov et al. 2005, [
26] noted that it is possible to find two different areas on different continents, with almost identical conditions, ‘but almost all, even perhaps all species of organisms will be different’ (p. 199). As early as in the 18th century, J. Buffon proposed a general law for the distribution of organisms: the limitation of groups of species to certain regions with definite boundaries. According to this rule, classifying communities based on dominant low-level taxa is possible on a regional scale, but does not make sense when comparing the vegetation of different continents [
27].
Nevertheless, as noted by C. Darwin [
28], despite differences in species composition, there is a striking similarity among functionally analogous organisms in different areas of the Earth. This convergence exists due to the unity of a common “plan of structure” of communities. Thus, both the processes of divergence, which increase diversity, and convergence occur. It seems to us that the biosphere has a continuum–discrete organization. It obviously does not consist of strictly limited parts or elements that would disrupt interconnections and integrity, but it does not represent an absolute continuum either, since its elements can be distinguished as subsystems with their own components and structures. Several continuum series of ecosystems, as elementary units of the biosphere, can be identified. In one of these (the ecosystem–biogeocenotic series), it is possible to place at one pole the ecosystems “according to Evans”, which may lack natural abiogenic components, while at the other pole, biogeocenoses, as elements of natural landscapes, are located. From a chorological perspective, a different series can be presented, in which at one pole there are well-limited discrete ecosystems, and at the other pole there are ecosystems of a gradient character that gradually transition from one to another. The concept of absolute continuity of the living cover leads to a fundamental error, which, according to I. Zhirkov [
16], results in the “non-recognition of the presence of an internally endogenous spatio-temporal structure of the biosphere” (p. 199). Even V.N. Beklemishev [
29] noted that the living cover of the Earth, which he called “Geomerida”, does not consist directly of biocenoses; rather, they are part of subsystems of the biosphere at intermediate levels.
Each level of the biosphere system fulfills both functions similar to other levels (fractal similarity) and functions characteristic of itself, with processes of different scales occurring within them. Thus, the photic zone of the ocean is an aggregate of many ecosystems of various sizes [
30], differing due to variations in hydrodynamic and thermal conditions and biota composition, but they are united in the ‘surface film of the ocean’ of Vernadsky [
1], representing a single system of the biospheromeron.
3. Ecosystem
An ecosystem is associated for us with a two-component system in which a certain physical space is inhabited by such a number of living organisms as can be supported by the available amount of resources in that space, i.e., it is a system in which living and inert elements co-exist. An ecosystem can be defined as “the biodiversity of species plus the interrelationship of flows of matter, energy, and information” [
31]. The simplest definition of an ecosystem is a community of organisms living and interacting with each other in a specific environment [
32]. An ecosystem is the existence of living things in contact with the environment. This has become an ecological truism, but it is important to emphasize the following two aspects arising from these definitions: living things on our planet are represented by organisms, individuals, but Earth’s life as a whole is a more complex system in which the living is inextricably linked with inert things. Paradoxically, life does not exist without the inert matter that it is inseparably connected with it, because living organisms provide cyclic processes, energy transformation, and resource renewability. Thus, the bioinert physical system becomes a part of life on the planet and in the biosphere, not so much due to the presence of living organisms, but due to the interaction between living and inert matter.
The second aspect is that, like organisms, ecosystems possess the property of self-insufficiency, which can be defined by an analogy with the principle of the self-insufficiency of species proposed by Yu.I. Starobogatov [
33], as follows: just as no species can exist outside certain ecosystems, an ecosystem requires for its existence a specific locus within the biosphere with particular conditions, including biotic and abiotic environments. The habitats of ecosystems are other ecosystems, both neighboring and distant. This leads to the formation of systems of ecosystems. Self-insufficiency, and the consequent need for associativity, is apparently a common property of all elements of the biosphere, just as organs in an organism’s body are self-insufficient and rely on the system as a whole [
34].
However, simple definitions of an ecosystem as a unity of living and inert matter are clearly insufficient. In this regard, expanded ecosystem principles have been formulated [
35], with the main ones summarized as follows: ecosystems can store energy and matter; they store information in their structures; and ecosystems are dissipative systems open to energy, matter, and information.
Thus, an ecosystem is not merely the sum of biotic and abiotic elements, but is a morphoprocess, possessing its own emergent properties and specific characteristics of “morph” (structure), which ensure the processes of matter transformation and energy dissipation characteristic of a given ecosystem (type of ecosystem as a whole).
4. Cenotic System
Ecosystems possess a certain degree of individuality. Just as an organism cannot exist independently for a long time without interacting with other organisms, i.e., outside of populations or communities, an ecosystem also requires exchanges of matter, energy, and information with other ecosystems (except for those artificially created or with conditions maintained by humans). Under certain circumstances, ecosystems form higher-order systems known as cenotic complexes [
36]. However, the term
cenotic may be somewhat misleading, as it emphasizes the biotic component—
biocenosis—whereas the nature of a biocenosis is shaped not only by endogenous factors, but also by exogenous, environmental influences. It may be more accurate to refer to these as ecosystem systems (local or regional). Such systems can vary in type and content and may be viewed as typological or real, chorological elements in the structure of the biosphere. The biome is often regarded as one such structure.
5. Biome
It should be noted that the introduction of the term and concept of the
ecosystem by A. Tansley in 1935 [
37] was a direct critique of the concept of the
biome introduced earlier by F. Clements in 1939 [
22]. The term
biome was first used by Clements in his opening speech at the first meeting of the Ecological Society of America [
38]. Earlier, A. Schimper [
39] (cited in [
40]), though not using the term
biome, argued that the global distribution of plants is determined by environmental conditions—climate and soils shape vegetation patterns on a planetary scale. His ideas largely aligned with those of A. von Humboldt. The biome, or the complex of organisms inhabiting a given area, is convenient in some cases, Tansley wrote, but more fundamental is the concept of a holistic system that includes not only the complex of organisms, but also the complex of physical factors—habitat conditions in the broadest sense. He also emphasized that [
37] “although organisms may be our primary interest, we cannot separate them from their specific environment, with which they form a unified system”—what he called an ecosystem. From this, it clearly follows that both A. Tansley himself and the author of the term, F. Clements, understood the biome as precisely, actually, and only the complex of organisms. This understanding led Tansley to introduce a new concept, the ecosystem—which includes these organisms within a real, functioning system of BIO and GEO, of living and inert substances. Despite the differences between these elements, in the system, they acquire a new property: bioinert unity.
It is interesting to note that one of the basic concepts among the predecessors of the concept of the biome is that of biocenosis [
21,
41,
42], that is, the idea of a biome arose as the idea of a set of biocenoses, with environmental conditions considered as a “background.” In a certain sense, this concept still exists today.
One remark should be made here. The concept of the “biome” means a plant–animal formation, a basic community unit. The concept and definition of the biome itself are not clear enough: “the term biome has been applied only to those communities in which studies have established something of the processes of development and the character of the final stage or climax” [
22] (pp. v–vi). A
formation, for example, in geology, is a certain set of rocks connected by a unity of origin, formed under similar conditions, regardless of their place and time of formation. A biome is rather a concept related not to the fundamental definition of the unity of living organisms, but to their classification. It is a certain type of community with characteristic features of organization. Indeed, Clements and Shelford [
22] present steppe and other biomes as examples. This is closer to the concept of a cenotic system. The concept of the biome has become widespread as a way to denote vegetation at regional or continental levels (
Figure 2). As noted by A.I. Kafanov [
26], this is rather a landscape bionomic application of the concept.
The following idea of the essence of the biome concept has been formed: it is a biotic community finding its expression at large geographic scales, shaped by climatic factors, and perhaps better characterized by physiognomy and functional aspects, rather than by species or life form composition [
38,
41,
42].
Since biomes are considered global or subglobal entities, the concept itself is regarded as a useful methodological tool for global assessments of changes in the biosphere [
43,
44,
45].
In hydrobiology, the term “biome” has found its application as “the historically established population of a water body, the totality of all life in a water body” [
46] (p. 23). Here, it can also be seen that the concept of a biome refers exclusively to the living population, although limited to a given water body. However, conditions—as well as biocenoses—can differ significantly within a single water body, even more so than between separate ones, for example, between littoral and profundal biocenoses. The unity of conditions within a water body is undoubtedly an important prerequisite for the unity of all living things in it, but from the authors’ point of view, it would be much more important to pay attention to the convergent similarity of many littoral biocenoses, as opposed to the typical structure of biocenoses in profundal areas. V.I. Zhadin developed such a generalized model of a river system [
47]. Obviously, the concept of a biome should apply to both aquatic and terrestrial life, so the following definition appears rather strange: a biome should be interpreted as “the largest unit of terrestrial communities, e.g. tundra, temperate forest, desert, tropical rainforest, and temperate grassland” [
48] (p. 535).
Thus, A. Tansley [
37], formally and essentially, introduced the concept of an ecosystem not as the “principal construct” of a bioinert system, but rather the concept of a BioGeome—essentially adding to Clements’ “Biome” the component
Geo, meaning the abiotic environment, the totality of conditions in a given region of the planet, thereby, in some sense, overlooking the chorological approach of F. Clements. In doing so, A. Tansley laid the groundwork for a contradiction that can be labeled as the “Evans–Odum and Sukachev–Vernadsky contradiction”: between an ecosystem as a unique bioinert structure of indefinite scale and biogeocenosis as an element of the Earth’s Face.
The consideration of typical ecosystems or aggregates of similar ecosystems as “biomes” has acquired a certain traditional status, becoming a scientific cliché. A. Protasov have not escaped this tradition either: in the work “Life in the Hydrosphere” [
49], it referred to aggregates of similar aquatic ecosystems as “biomes”.
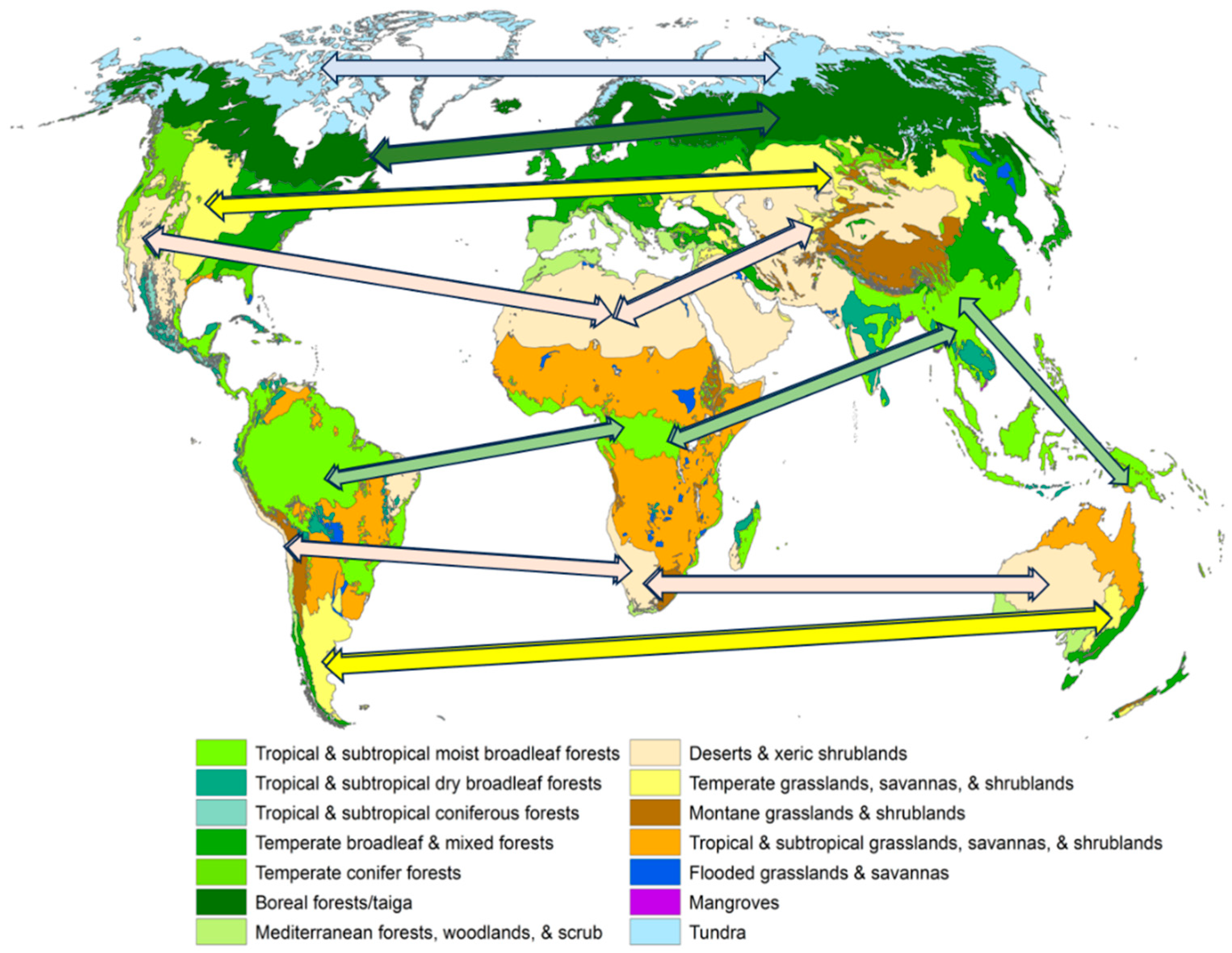
Figure 2.
Main terrestrial biomes (adapted from [
50]). The arrows connect biomes of the same name and type, which makes their mostly latitudinal location obvious.
Figure 2.
Main terrestrial biomes (adapted from [
50]). The arrows connect biomes of the same name and type, which makes their mostly latitudinal location obvious.
6. Geome and Biome as Elements of the Biogeome System
As for the term “biogeome” itself, it was previously used in stratigraphy and paleontology [
51]. The author of this work understands it as a “biogeological unity” of the biome of past epochs and its habitat, or geome. In fact, this is an attempt to reconstruct paleoecosystems. Moreover, if add the “principle of facies”—repeatability and typicality in different places and epochs—then can talk about the closeness of the concepts in the sense of the authors. It should be only added the paleobiogeome, given that a paleontologist deals with geological, not living, reality [
52]. In paleontology, the geobiome is currently understood to mean local and regional biota and embedding geosystems that evolve in tandem and exist on geological time scales [
53].
The general principle of the correspondence between biota and environmental conditions, which was laid as the foundation of ecological concepts by A. von Humboldt, Ch. Darwin, K. Roullier, E. Haeckel, and other scientists, was later reflected in various models. One of the simplest is the bell-shaped curve of population size along a gradient of a given factor, which can be found in any ecology textbook. This illustrates that only within a certain range of values for a specific factor does a population find optimal conditions and develop most intensively. In fact, the model of the multidimensional ecological niche [
54] represents a “multiplication” of such curves—an increase in the number of factors that determine the potential or actual position of a given species within a community.
Ecological factors are of a continuum-gradient nature, and there are limited areas of combinations, in which ecosystems are quite similar in their characteristics. Thus, in the multidimensional field of factors, there is a certain gradation of their significance for individual populations, communities, and entire ecosystems. For example, rainforest ecosystems exist at values of radiation balance ranging from 210 to 315 MJ/m
2·year and a dryness index (the total amount of energy required for evaporation of the annual sum of precipitation) close to one [
55]. Temperate forest ecosystems are formed under such a combination of conditions when the average annual temperature is about 10 °C and precipitation is about 150–200 cm/year [
56]. It follows that the characteristics of a geome can be rather precisely limited by certain values of environmental factors and in a rather narrow range of values in which a given biome exists.
Based on the analysis of the correspondence between the combination of conditions and the character of vegetation, very detailed classification systems for vegetation formations have been constructed [
57,
58]. Traditionally, in biogeography—where, in particular, studies of species ranges (as well as higher taxa) play an important role—the principles of similarity by analogy, based on the similarity of edifying vegetation species, prevail in classifications of vegetation formations. However, as recognized by biogeographers themselves [
59], under the direct and indirect influence of similar environmental conditions, deep analogies are also observed in other ecosystem characteristics, such as the level of primary and secondary production and geochemical transformations. Some adaptations can be realized in the same conditions in different ways, not to mention the fact that different fauna and flora exist in different regions, while similar functions in biocenoses and ecosystems are performed by organisms of different, often alien taxa.
The geome is the totality of factors that support life in a given place of a quite certain biome—the totality of living things. As a whole, they form a system of relations, the very bioinert unity that A. Tansley pointed out. A biogeome could be called a “large” ecosystem if its separate parts (e.g., grass biogeome or bioherm biogeome) were not located on different continents and in different oceans. The authors are dealing here, rather, with fractal similarity, the similarity of some key features of the structure and function of systems of different levels.
Considering the widest adaptations of different communities to environmental conditions, where combinations of key conditions are limited, the authors come to the term and concept of the biogeome, i.e., a set, a system of convergent ecosystems regarding their basic characteristics and properties [
60,
61].
A biogeome is not a simple sum of ecosystems in a given area of the biosphere, but one of the integral elements of the biosphere. As an integral element, it must have its own emergent, inherent properties.
First of all, it should be noted that biogeomes’ subglobal, subplanetary geographic scales [
44] are measured in thousands of kilometers. The distances between individual parts of biogeomes can also span thousands of kilometers. This scale has been emphasized repeatedly [
38,
44]. In addition, it should be noted that, unlike in ecosystems—where communities are systems of interactions between species populations—more general biotic concepts, such as ecomorphic groups and life forms, must be used to characterize a biome. These determine both physiognomic characteristics and the general direction of functioning, including energy flows and the balance of matter circulation. An ecosystem is a bio-inert system in which the biotic component is always the primary subject of research; in a biogeome, by contrast, the abiotic component (geome) can determine the features or variants of the physiognomic approach to assessment and characterization.
If the smallest unit of the biosphere structure is local, specific ecosystems, then the next level (sublevel, in the structure of the biosphere) is, obviously, regional ecosystem complexes. Their scale can vary depending on features of both the biome and geome, the individuality of the ecosystems, and the research objectives. A purely geographical approach allows us to identify, for example, regional hylea complexes in Amazonia, East Africa, and Southwest Asia. At the same time, it is useful to identify regional complexes within parts of a country, such as the northeast. Here, two ecosystem complexes are distinguished—the Amazonian and coastal Atlantic hylea [
42].
7. Diversity and Uniformity of Ecosystems
The richness of ecosystems is enormous. It is probably comparable to the species’ richness in the history of the biosphere. However, ecological convergence is also widespread. The primary ecosystems on land, in atmobiont conditions, obviously originated quite long ago, and the primary ones could consist only of elements of aquatic ecosystems [
62]. Nevertheless, the evolutionary paths of hydrosphere and terrestrial ecosystems are significantly different. They belong to different biospheromerons, and the hydrosphere is a much more complex structure as a geome than the land.
If two factors—humidity and temperature (geome)—are enough for a satisfactory description and mapping of terrestrial biomes [
55,
56], the situation is much more complicated in the hydrosphere.
The differences in geomes for atmobiont and hydrobiont organisms are great. On land, the main bioinert element of ecosystems is soil, which lies in a thin layer on the mineral base, while in the hydrosphere, the reservoir of biogenic matter is mainly water mass and bottom sediments. Water also provides biogenic element transportation. Therefore, heterotrophic communities without an autotrophic link are widespread in hydroecosystems. They exist due to the production of organic matter produced in other, rather distant ecosystems. The hydrosphere as a habitat has a high stability and uniformity of conditions over large areas [
63].
The overwhelming majority of atmobionts, dwellers of land, live in oxidizing conditions, while reducing conditions are quite widespread in the hydrosphere. The volume of the living environment for inhabitants of the hydrosphere exceeds 1 billion km3, whereas on land, this volume is no more than 200 million km3. Additionally, the environment in the hydrosphere is water with a huge heat capacity, which provides much more stable conditions.
What creates and maintains the diversity of ecosystems, and what are the prerequisites for divergent processes? There are several [
61].
The ecological individuality of species, ecomorphs, can be defined as follows:
A diversity of ways of obtaining nutrients and energy;
A variety of adaptations for reproduction, population increase, and maintaining populations amid competition and environmental influences;
A variety of ways to develop and maintain habitats.
In addition, there are a large number of biocenotic relationships that change over time.
In nature, there is a great variety of combinations of different environmental factors and great diversity of inert environments of communities of organisms. There are also many manifestations and prerequisites of convergent processes. The main ones are as follows:
Morphological, behavioral similarity of species that are part of one ecomorpha, one life form.
Similarity of trophic adaptations, limited trophic levels.
Limitations of types of life strategies.
Limitations of the main sources of energy and types of biotic transformation.
Existence of only a few basic types of habitats for organisms.
8. Ecological Convergence
A lot of data shows that ecosystems and biomes compared at fairly large distances have few common species, but ecosystems can be very similar “physiologically” in a functional sense: plant growth forms, ecomorphs, and life forms are common, while phenology and the nature of local climate manifestations can be similar [
38,
39].
The convergence of biotic systems is manifested not only in the similarity of organs and organ systems in different organisms, but also in the ecological similarity of populations that live in similar conditions and in the similarity of the structure and organization of communities under similar environmental conditions. One of the manifestations of ecological convergence is the formation of life forms, ecomorphs, which have a great similarity both in morphological and functional aspects but are formed on the basis of completely different taxa [
64]. Having paid attention to this, back in the 19th century, the great German naturalist A. von Humboldt developed the principles of plant biogeography based on life forms, identifying a relatively small number of them, about 20 [
65]. Strictly speaking, the characteristics and classification of biogeomes are determined (for a biome) primarily by nature, composition, degree of dominance, and contribution to the general habitual and functional characteristics of biocenoses of ecomorphs and life forms, and not species or other taxa [
61]. “Morphology”, if one can put it that way, and the habitual characteristics of ecosystems are closely related to the composition of ecomorphs, more than taxa.
In hydrobiology, the accumulation and analysis of materials on the structure of marine benthos biocenoses led to the creation of the concept of parallel communities [
66,
67]. At the beginning of the 20th century, the Danish hydrobiologist K. Petersen [
20] analyzed the material on the structure of many benthic marine communities and recognized no more than 14 types, which he named after the dominant organisms. Developing the idea of the convergent similarity of communities, the Danish hydrobiologist G. Thorson proposed the concept of parallel communities in the late 1950s. Its provisions were developed by A.P. Kuznetsov, who drew attention to the fact that the criterion of taxonomic similarity is insufficient and, in some cases, contradicts the concept, since closely related species may differ both in their modes of nutrition and levels of metabolism, while unrelated species perform similar functions in communities and occupy similar ecological niches. This is due to ecomorphological differences among genetically close forms. It should also be emphasized that the convergent similarity of communities is based on a rather limited combination of environmental conditions. For marine benthic ecosystems, these are the characteristics of sediment, depth, and temperature.
Back in 1807, A. von Humboldt wrote the following: “As individual organic beings have their own physiognomy, so there is a physiognomy of nature that belongs exclusively to one climate” [
68] (p. 491). Thus, in similar conditions, similar communities are formed, which create the “physiognomy”, the image of nature. This regularity can be defined as the principle of the similar distribution of close life forms or the law of “correspondence of the complex of life forms to the environmental conditions” by A. von Humboldt.
In the “geographical” approach, the concept of landscape can be used as a basis for the structure of the biosphere. According to the definition of the Russian and Soviet evolutionist and geographer L.S. Berg [
69], this concept has the following content: a complex of features of relief, climate, water, soil and plant cover, animal life, and the results of human activity, which “merges into a harmonious whole, typically repeated throughout a given zone of the Earth” (p. 5). Each landscape has its own appearance; the combination of its elements is unique. This is why L.S. Berg considered landscapes as “geographical individuals” or “specimens”. This view is important because it leads us to some analogies: if individuals, living organisms possessing unconditional individuality, are united in populations—and moreover, cannot exist outside them—can be speaking about “populations” of geographical landscapes? Generally, they form “species” landscapes’ areals. Biogeography distinguishes continuous and disjunctive, i.e., broken or fragmented, areals.
But physiognomy, habitus, is only the visible part of nature’s appearance, as G.A. Zavarzin, a Russian evolutionist and microbiologist, pointed out, as follows: “...landscape can be described not only physiognomically, in the style of A. von Humboldt, but also geochemically, by the dominance of certain geochemical processes” [
70] (p. 8). This means the processes of substance transformation due to the energy entering the ecosystem. It is within the landscape ecosystem that the very exchange of atoms, about which V.I. Vernadsky wrote, takes place.
Accepting, but also clarifying V.I. Vernadsky’s synonymic approach to the definition of the Biosphere as the Face of the Earth [
1], the authors can say that the “Biosphere” has an ecosystem as the basis of its structure, and the “Face of the Earth” has landscapes. Their difference is that the concept of landscape is geocentric, while the concept of ecosystem is biocentric. Moreover, this difference is cognitive rather than natural.
9. Typization of Ecosystems by “Biotic Domination” and Formulas of Biogeomes
Thus, a quite limited combination of conditions for both hydrosphere and land inhabitants, on the one hand, and a wide convergence and similarity of adaptations, on the other hand, lead to the fact that many ecosystems in nature are quite similar. This allows us to unite them into a limited number of biogeomes (
Figure 3).
In atmobiont conditions, on land, two factors—humidity and temperature—as well as the nature of soil, act as the main geo characteristics for ecosystems. Thus, the formula of terrestrial biogeomes can be represented as (T + H + S) for the geome, where T is the thermal regime, H is the moisture regime, and S is the soil character. For the biome, this can be represented as (LF + Tr + Hr), where LF is the composition of basic life forms, Tr represents trophic features, and Hr includes chorological characteristics, as well as the nature of longitudinality, stratification, mosaicity, and boundaries.
That is, the characterization of the terrestrial biogeome (TB) can be given according to the following formula:
The relationship between GEO and BIO components in the hydrosphere is much more complex. Hydrobiogeomes are influenced by environmental characteristics that differ significantly from those on land. The formula for a biogeome in the hydrosphere may be expressed as follows:
Biome = key life forms of biota (LF, including ecomorphs and cenoecomorphs) + trophic structure (Tr, i.e., the main transformers of energy from particular sources and main trophic links) + chorology (Hr, including boundaries, tiering, stratification, and mosaicity).
Thus, the general formula for a hydrobiogeome (HB) may be written as follows:
Ecosystems can be divided, albeit conditionally, into the following three types: “biotic”, “oligobiotic”, and “nanobiotic” or “subbiotic”. This division is based on the greater or lesser importance of biotic or abiotic physiognomic/habitual and structural–functional components.
The first type includes ecosystems with a clearly expressed edifying role of biotic components. These are forest ecosystems on land and coral (bioherm) ecosystems in the ocean. Their entire appearance and spatial structure are determined by the biotic elements of the ecosystem.
Ecosystems of the “oligobiotic” type (with a low degree of biotic “creative” influence) are those in which the biocenosis does not play a defining role in the overall appearance, but may play a significant role locally. Examples include the grassland biogeome and tundra. A considerable number of ecosystems in the hydrosphere can also be attributed to the oligobiotic type, such as sea shelf ecosystems, reservoir ecosystems, and surface runoff watercourses.
Finally, the “nanobiotic” type of ecosystems is characterized by the dominance of inert elements and factors in forming their overall appearance. Examples of such ecosystems include pelagic ocean ecosystems and desert ecosystems.
10. Biogeomes of Biotic Type
A forest is a natural system in which the primary role in plant communities belongs to woody plants. Tree forms in ecosystems create a complex spatial structure of communities. This “main role” is manifested in many aspects, primarily in the fact that tree form vegetation creates a distinct biogenic biotope for other organisms. The most important factor, from the perspective of geome differentiation, is the constancy and periodicity of the environment. This significantly influences the characteristics of the biome and ecosystems in general. Taking into account the edifying role of tree ecomorphs and the periodicity or relative stability of the geome, the following two forest biogeomes can be distinguished: tropical rainforest (hylea) and seasonal forests.
Tropical rainforest ecosystems are distributed within a narrow zone of the tropical region, approximately between 10° N and 10° S. The environmental conditions in this zone are characterized by minimal fluctuations in the key factors—temperature and precipitation. Annual precipitation is high, reaching several meters in some areas [
71]. The current area occupied by these ecosystems is estimated at 17 million km
2, which is about 10% of the Earth’s land surface.
According to estimates, the area of forests worldwide (Based on The Food and Agriculture Organization (FAO) data [
72]) is 4.03 billion hectares, or 40 million km
2. This constitutes 31% of the land surface. If it accept that hylea occupies about 10% of the land area [
72], then the total area of all other forests with periodic environmental conditions is approximately 27 million km
2.
Regarding the identification of biotic-type biogeomes in the hydrosphere, the classification of biohermic ecosystems (currently represented by coral reef ecosystems) is beyond doubt. In these systems, the biotic components create an entirely new habitat for other members of the biocenosis—biogenic elements of the underwater landscape. Moreover, reef structures formed by colonial sedentary organisms not only constitute part of the biotic environment, but also shape the geome itself, effectively becoming a “geological force”, to recall the words of V.I. Vernadsky. In any case, the trophic relationship system—based on trophosymbiosis—is also unique to the key organisms of biocenoses in this biome. As with the hylea geome, the temporal periodicity of environmental conditions is weakly expressed in the bioherm biogeome.
A summary of data on more than 1000 coral reefs (McClanahan, 2002 [
73]) shows that the temperature limits for the existence of reef ecosystems range from 16.0 to 34.4 °C (with even higher temperatures recorded in the Red Sea). Water salinity varies from 23.3 to 41.8‰. The concentration of nutrients reaches up to 3.3 mmol/dm
3 for nitrates and up to 0.54 mmol/dm
3 for phosphates. The illumination zone extends from 9 to 91 m in depth. The total area of modern coral reefs is estimated at about 600,000 km
2 [
74]; however, some sources suggest a significantly higher estimate—up to 7 million km
2 [
75].
11. Oligobiotic Biogeomes
These ecosystems are characterized by the fact that certain abiotic factors, along with biotic ones, shape their appearance. Low winter temperatures and short summers contribute to the formation of a permafrost layer in tundra soils (a geome characteristic). This determines the prevalence of low-growing vegetation and the absence of tree forms (biome features). However, during prolonged summer insolation, a considerable amount of primary organic matter is produced, which accumulates in the form of peat and large detritus. In grassland ecosystems of the temperate zone (such as steppe or prairie), an important abiotic factor is low summer humidity and drought, which lead to a reduced rate of organic matter decomposition and its accumulation in the soil.
In the hydrosphere, a considerable number of ecosystems can be classified as oligobiotic. Shelf ecosystems are highly diverse, and the ratio of biotic to abiotic components varies significantly. In underwater “forests” of brown algae and in the upper rocky littoral, densely populated by both sedentary and mobile organisms, biotic elements clearly predominate. At the same time, life in the littoral zone is largely shaped by tidal phenomena. Overall, the fact that this part of the biosphere is where the marginal zones of three biospheremerons—surface oceanic, bottom, and intermediate (I, II, and III biospheres)—converge with the fourth, the land, compels us to consider the entire set of shelf ecosystems as a separate biogeome. This is one of the most significant contour zones in the biosphere.
It is undeniable that hydrothermal ecosystems should be recognized as a separate biogeome [
76,
77]. They appear completely isolated in contrast to the majority of ecosystems—photoautotrophic and heterotrophic. While the vast majority of Earth’s ecosystems derive their energy from the sun (even the organic matter in heterotrophic communities has a photosynthetic origin), hydrothermal communities, formed through the activity of chemosynthetic organisms, resemble islands in the ocean of “green life” (a term used by Vernadsky). Hydrothermal fluids are a geological phenomenon, and in ecosystems whose very existence depends on the intensity and duration of thermal fluid outpouring, hydrothermal activity is the determining factor. At the same time, the physiognomy of these ecosystems is shaped by clusters of unique organisms.
Ecosystems of surface runoff water bodies are closely related to terrestrial ecosystems; however, it is necessary to distinguish them as separate biogeomes, since they belong to two different environments—the hydrosphere and the atmosphere. To differentiate between the two biogeomes—the rheobiogeome (lotic ecosystems) and limnobiogeome (lentic ecosystems)—the main abiotic criterion is the intensity of water exchange. Differences in the temporal nature of processes are also associated with this factor. In reservoirs (with slow water exchange), successions are progressive. In watercourses with intensive water exchange, cyclic changes occur in both inert and biotic elements. A peculiarity of rheobiogeome ecosystems is that, spatially, their structure is linear, with the main gradients directed from the source to the mouth; temporally, these systems are characterized by pulsation processes. A watercourse can be considered both as a single system and as a series of successive local ecosystems connected along its length. However, when considering the structure of the entire rheobiogeome, its elements are integral river ecosystems.
An important step in generalizing ideas about various lotic ecosystems was the creation of the river continuum concept [
78]. This concept was an attempt to establish a foundation for describing the functioning of lotic ecosystems by generalizing knowledge about lotic systems as a whole, without emphasizing regional or geographical features. It is one of the key concepts in biogeomics in general. The concept is based on a generalized view of an entire ecosystem type, identifying typical elements and connections between them, and the idea of convergently similar communities under similar conditions. It necessitates isolating and considering the main environmental gradients and a generalized ecomorphological view of the biota. There is every reason to consider the limnetic (lake) continuum in a manner similar to the river continuum [
79].
12. Nanobiotic Biogeomes
In ecosystems of this type, inert elements determine their general physiognomy. On land, these include various deserts, whose formation and very existence are dictated by combinations of factors close to extreme conditions: high or low temperatures and extremely low water availability.
In the ocean, this type of biogeome includes the following two ecosystems where inert elements clearly predominate: the oceanic pelagic and benthic biogeomes. The primary production of the ocean pelagic zone, as a whole, does not exceed that of terrestrial deserts. However, both benthic ecosystems and parts of the pelagic ecosystems connected with the photic zone are contour ecosystems on an oceanic scale, where intense metabolic processes occur. Nevertheless, the ratio of biotic and abiotic components in these ecosystems clearly favors the latter.
In the allocation of biogeographic zones in the pelagic part of oceans, latitudinal zonation is clearly expressed [
80], however, like all biogeographic divisions, it is based on taxonomic criteria of differences and similarities [
26]. However, given the unity of the entire pelagic zone of the World Ocean as a single biogeome, the focus should be less on latitudinal zonation and more on the global hydrodynamics of the oceans associated with currents and gyres [
29]. In all three oceans (excluding the Arctic Ocean), large-scale circulations are well expressed on both sides of the equator and the intermediate complexes between them. Pelagic ecosystems are also characterized by a special type of relationship between succession in time and mobility in space [
81,
82]: when intensive zooplankton development lags behind the peak of phytoplankton development, a shift of several weeks or a month occurs, with the entire pelagic ecosystem moving in space for hundreds of kilometers. Also, all pelagic ecosystems are characterized by mass vertical migrations of hydrobionts, both daily and seasonal.
Thus, the authors distinguish 12 biogeomes that cover almost the entire biosphere of the Earth (
Table 1).
13. Extra-Biogeome Ecosystems
Obviously, the above table cannot show the many transitions between biogeomes. There are ecosystems that are difficult to categorize into any of them. Biogeomes are not absolutely linked to geographic climatic zones in their distribution; other factors, such as the nature of soils, also play a role. The great diversity of ecosystems cannot be strictly and completely reduced to a small number of biogeomes. Among the most important extra-biogeomic ecosystems are savanna, wetland ecosystems, mangrove ecosystems, and estuarine ecosystems. It should be noted that all of them have a transitional, ecotone character. High-mountain ecosystems have their own specificity.
14. Techno-Biogeome
To the diversity of combinations of natural elements, both BIO and GEO, humans add a large number of different anthropogenic-technical (TECHNO) elements. A new type of composite, anthropogenic ecosystem is definitely being formed in the biosphere. They are no longer small islands in the natural world, but are increasingly expanding their field of existence. The landscape appearances of entire countries and geographic areas have been completely transformed by human activity. Anthropogenic ecosystems are very diverse, and combining them into a conditionally single techno-biogeome is still problematic, but it is necessary to take into account their role in the biosphere.
It is important to find out what the transformation mechanism of the biosphere into the noosphere is, which V.I. Vernadsky wrote about [
2]. Evolutionarily significant restructurings in the biosphere occur through the replacement of some types of ecosystems by others, with not only the components, but also the key connection changes. In this context, does the transformation of the biosphere as a purely natural system into the noosphere occur precisely through the transformation of natural ecosystems into anthropogenic or anthropo-dependent ones?
Nothing in the biosphere remains in a static, motionless state. The flow of atoms, which V.I. Vernadsky described, exists constantly in the most diverse forms of matter transformation involving living organisms. The forms of this flow also change. These changes occur on very different time scales. Ecosystems pass through certain stages of succession, which can last years, decades, or even centuries. Changes in climatic or hydrological conditions can significantly affect the complex of ecosystems across entire regions. For example, in just a few decades, the ecosystem of the entire Aral Sea basin was completely transformed, whereas the formation of ecosystem types and biogeomes requires time measured in geological scales.
15. Evolution of Biogeomes
Any discussions about the history and evolution of biogeomes, ecosystems, and their components will always remain speculative, since the only paleontological realities are various fossils of fragments of communities that have vanished forever, elements of geomes, and petrified remains of organisms and biomes. Modern, currently existing ecosystems and scant traces of former biospheres cannot provide us with the full picture of the biosphere’s development over millions of years.
The question of the evolution of biogeomes is controversial. Do biogeomes evolve as biospheric entities, or do individual elements of the biome—species, ecomorphs, and communities—evolve independently, adapting to new states of geomes (such as changes in the ocean, soils, weather, and climate conditions) [
83]?
When considering the problem of the origin of life, the question should not be about the origin of the heredity apparatus alone, but rather about what existed first—the organism or the ecosystem? It is impossible to discuss the origin of an element separately from the origin of the entire system [
72]. This system is the biosphere, whose elementary unit has always been the ecosystem.
A difficult question deserves attention, as follows: why, within a relatively narrow range of atmospheric conditions in the modern biosphere—average temperatures ranging from –10 to 30 °C and precipitation ranging from nearly 0 to 450 cm/year—have up to ten significantly different types of atmospheric ecosystems evolved [
56], rather than just one or two with some modifications? It is highly likely that the biosphere today represents the most complex biogeome system, encompassing the largest number of biogeomes and extrabiogeomic ecosystems. “Only” 450 million years ago (compared to about 3 billion years of biosphere existence prior to that), there was no terrestrial biospheremeron and no atmospheric ecosystems or biogeomes at all. As for the hydrosphere, it should probably be assumed that all oceanic biogeomes already existed by the late Precambrian, although the composition of their ecosystems was completely different.
It is possible to draw a certain parallel between the evolution of biogeomes and ecosystems and evolution in the taxonomic trend [
9]: the evolution of ecosystems, as well as species, took place within the framework of the evolution of biogeomes as the macroevolution of “large” taxa.
The biosphere has passed through three major stages of life environment using ocean, continental waters, and land, or the transition from a purely hydrospheric and hydrobiontic biosphere to a hydrobiontic–atmobiontic one. These stages cannot be confined to precise time frames. The first stage clearly coincides with the origin of life on the planet, which most likely arose in sea basins within the ocean-continent contour zone. In any case, the earliest paleontological evidence of living organisms dates back to 3.2–3.5 billion years ago [
10,
73]. These organisms must have been part of some communities and ecosystems. Most likely, life initially existed in primary coastal ecosystems. Pelagic, coastal, deep-sea benthic, and possibly hydrothermal ecosystems already existed in the late Precambrian era.
Bioherm ecosystems, based on the vital activity of animals, could only have arisen at the beginning of the Paleozoic, since skeletal animals were entirely absent in the Precambrian. However, the bioherm biogeome, as a whole, existed much earlier, as algae in the hydrosphere formed the most ancient biogenic structures—stromatolites.
It is important to note that over a long evolutionary period, the processes of divergence—increasing the diversity of ecosystems and their composition—have constantly interacted [
84], while, simultaneously, there have always been processes of convergence, leading to the formation of certain types of ecosystems—biogeomes.
The authors adhere to the concept of the emergence of life immediately or directly in the form of ecosystem-like formations, with subsequent evolutionary steps involving the emergence of organisms and the phenomena of their self-replication and self-reproduction [
7,
8,
9]. The structure of protoecosystems is difficult to imagine, but their great diversity is unlikely. These were most likely coastal marine ecosystems functioning on the basis of formations similar to bacterial mats, in which both the production and the destruction of organic matter occurred. The diversity of the biosphere in this aspect approached zero—not only because there were few types of ecosystems, but also because one type dominated.
The complication of the biogeomic “construction” of the biosphere over evolutionary time is obvious. At present, there are many types of ecosystems (with different approaches to classification, ranging from one and a half to several dozen), but it is also important that no single biogeome currently represents an absolute dominance in the biosphere. Thus, over several billion years, the diversity of bioinert systems has grown not only in terms of increasing the number of taxa, but also in terms of increasing the number of ecosystem types and the balance of their significance within the biosphere.
In the approach to revealing the regularities of the evolution of the biosphere as a whole, it is constructive to identify elementary systems, processes, and evolutionary events both at the microlevel—within the evolution of species—and at the macrolevel, concerning the evolution of the biosphere as a whole and its major subsystems [
85]. If the elementary unit of the biosphere is the ecosystem biogeocenosis, then the elementary evolutionary process may be defined as follows: “A qualitative stage of evolution at the biosphere level—the formation of a new biogeocenosis, the metamorphosis of the metabolism of a former one” [
86] (p. 26). However, as noted by L.S. Berg [
87], such a process must be of an “epidemic character”, i.e., it must affect many ecosystems and extensive regions simultaneously.
A new ecosystem with significant differences is essentially a “chimera”, an anomaly—thus, a biogeome should have a polyphyletic basis. There are probably several mechanisms by which new biogeomes are formed: through the emergence of new ecosystems driven by evolutionary pressures and the appearance of entirely new life forms. For example, the emergence of arthropod filter feeders [
88] significantly transformed pelagic ecosystems, altering not only individual biocenoses, but the ecosystem structure—and, hence, the biogeome—as a whole.
There is reason to believe that the emergence of life on land and the formation of the atmobiont biome served as the foundation for a fundamental reorganization of the nature of surface water runoff [
62]. This, in turn, led to the formation of watercourses and rivers, which became the structural basis for the geome of rheophilic hydroecosystems and the entire rheobiogeome.
Another evolutionary pathway is associated with significant transformations of the geome. For instance, episodes of aridization—occurring at different scales across the biosphere in various geological periods—inevitably led to a radical restructuring of ecosystems, affecting both biotopes and biocenoses. This process involves not only a reduction in or disappearance of previous ecosystems, but also the formation of entirely new ones.
Moreover, as V. Krassilov [
89] pointed out, periodic geological crises also played a significant role in shaping biospheric evolution. The causes of these crises were apparently systemic and multifactorial, including both internal biospheric dynamics and external influences. According to Krassilov’s concept, during such crises, environmental destabilization occurs, leading to the elimination of dominant—typically the most specialized—species from cenotic (community) networks. This, in turn, triggers a restructuring of cenotic relationships and, on a broader scale, the reorganization of entire biogeomes.
One may ask why the biosphere did not follow the “idioadaptive” evolutionary path, using the terminology of A.N. Severtsov [
90,
91,
92], or rather, not only this path, the path of “bad” diversity [
93], the endless repeating and reproducing of “successful” ecosystem models. Instead, it followed a more complex course, in which epochs of coherent [
89], idioadaptive evolution were interrupted (or rather, separated) by periods of ecological aromorphoses. Apparently, at least three factors or mechanisms can be identified in the evolution of biogeomes.
The first is the pressure of life, in V.I. Vernadsky’s terminology. Purely biotic causes led to the emergence of new life forms, new species, which not only required new ecological niches, but also created them [
94]. However, the capacity of the ecological environment is not unlimited, and “niche conflicts” led to ecological crises.
The second path of biospheric evolution was associated with ecological aromorphoses—the emergence of significant, fundamentally new methods of adaptation [
85,
90]. Since the evolutionary ecological unit of the biosphere is the ecosystem, changes, whether more or less significant, occurred at the ecosystem level, but were fixed as biospheric evolutionary events at the level of the biogeome. Changes in the geome also led to evolutionary acts, for example, the accumulation of oxygen in the atmosphere once its geochemical consumption decreased [
85].
Finally, the influence of external, cosmic factors cannot be ruled out. For instance, V.A. Krassilov [
89] believed that the periodicity of geological crises is connected to the cyclicity of galactic phenomena.
Speaking about the emergence and evolution of biogeomes, it can turn to phenomena in biospheric evolution that are relatively well documented by paleontological research, namely, the formation of epigeic biogeomes [
62,
63]. Individual living organisms may have penetrated onto land from aquatic environments long before the Silurian, possibly forming ephemeral populations and communities. However, the true emergence of life on land and the formation of bioinert systems began not merely with the presence of life (the biome) itself, but with the establishment of a geome adequate for terrestrial conditions—primarily, the formation of soil as the main bioinert element of terrestrial biogeomes. In these processes, the biome played a key role.
Aquatic ecosystems had been “preparing” the oxygen-rich atmosphere for over a billion to a billion and a half years. Life on land could only persist once living matter—organisms through their activity—created a habitable environment for the biome, thereby forming an essential part of the new geome. Cyanobacterial mats [
70], it seems, existed only in aquatic environments; the first terrestrial ecosystems had to be built on the basis of biocenoses composed of euatmobionts.
Evolutionary processes in the biosphere have unfolded over hundreds of thousands and millions of years, but anthropogenic influence is likely to accelerate them dramatically. The fact is that natural ecosystems are being replaced by agro-, techno- and urbanized ecosystems. Considering that, for instance, most of the land in Ukraine has been transformed into agro-ecosystems [
95]—a trend observed across many regions of Europe and other continents—should be not speak of a fast-paced evolutionary process: the transformation of natural ecosystems into anthropogenic, or more precisely, anthropodependent ecosystems?
In this context, the issue of human-created agro-, techno-, and urban ecosystems becomes particularly important. Formally, these ecosystems increase the overall diversity of the biosphere, but does this not lead to a shift in ecosystem equitability and a decrease in its resilience? Unfortunately, there is reason to answer in the affirmative. However, much depends on the successional stage of the ecosystem under consideration. Moreover, humans are increasingly creating conditions that exceed the tolerance thresholds of many organisms [
96].
That is, the parameters of the environment that receives biota are limited under Earth’s conditions. For aquatic ecosystems, for example, the existence of primary producers of organic matter—photosynthetic autotrophs forming the base of the trophic pyramid—is not possible under just any environmental conditions. It is only feasible within the first quadrant of the limnosaprobity model of aquatic ecosystems [
96,
97,
98]. The remaining three quadrants of this model represent water characteristics ranging from polluted wastewater to technical fluids, where a trophic pyramid based on autotrophs cannot exist. In other words, the existence of a biogeome, like that of an ecosystem, is constrained: it is only possible under environmental conditions that allow for the succession of organism communities.
Thus, systemic properties such as discreteness, dynamism, historicity, and hierarchy are further complemented by environmental and structural thresholds that determine where an ecosystem capable of producing organic matter via photosynthesis can exist on planet Earth.
16. Conclusions
In fact, the modern concept of the biome is close to our understanding of the biogeome, since the former includes, for example, climatic factors, and if considered in the broadest sense (as a geome), it expresses the unity of ecosystems at large—planetary and subplanetary—scales and is characterized by physiognomy. A biome, indeed, is not defined by species composition. However, it is not acceptable to treat the “functional” aspect in isolation, separately from the biota itself, i.e., exclusively at the level of ecomorphs and life forms. Just as for the hylea, the key ecomorph is the woody plant form, for the bioherm biogeome, it is the hermatypic corals or other reef builders, and for the pelagic biogeome, the dominant life forms are the ecomorphs of nekton and plankton.
It is necessary to pay attention to the definition of biomes as “constructs for organizing knowledge on the structure and functioning of the world’s ecosystems, and as useful units for monitoring how the biosphere responds to anthropogenic drivers, including climate change” [
35]. That is, biomes are viewed not as elements of the biosphere’s structure, but only as cognitive constructs, in cognitive terms. This approach is obviously determined by the dominance of the concept that “the biosphere is the largest ecosystem”, which does not require consideration of the hierarchical structure of the biosphere.
The concept of the biogeome allows us to form the most general understanding of both the structure of the biosphere and its main components. The biosphere should be considered as a complex hierarchical fractal system consisting of bioinert elements and subsystems of various levels. It cannot be viewed simply as a collection of living organisms or as one of the Earth’s covers, whose conditions are favorable for life. Life on Earth exists in the form of bioinert systems at different levels, with the ecosystem as the elementary unit of biospheric life. The evolution of the biosphere proceeded such that, after periods of coherent evolution, many different ecosystems formed within specific biogeomes—ecological aromorphoses occurred. These were widespread significant changes in the structure and functioning of ecosystems, which became established in new biogeomes. The cause of this restructuring could be evolutionary changes in either the biome or the geome.
What is new in the biogeomic approach from a cognitive perspective? First, it offers the opportunity to perceive the general, typical characteristics of ecosystems with similar structures and appearances beyond the detailed study of individual ecosystems. Additionally, it provides new insights into the complex yet ordered structure and organization of the biosphere. For atmobiology, it is important that, despite the great diversity of terrestrial ecosystems, they all belong to a single biospheremeron, and in the broadest terms, their “fundamental structure” is quite similar. A relatively small number of life forms and organisms perform similar functions. This results from both ecological convergence and common origin. Moreover, the range of the main environmental factors within the geome is much narrower than in the hydrosphere.
For hydrobiology, particularly the study of aquatic ecosystems in surface runoff, such a view of generalized ecosystem models—specifically of two types, lotic and lentic—is important. Additionally, in the vast expanses of the oceans, including the ocean floor, there exist many ecosystems, yet their general structure shows certain similarities. Thousands of coral ecosystems each have their unique characteristics, but the biogeomic approach allows us to examine them from the perspective of their overall design and structure. It is quite obvious that specific studies focus on very particular water bodies, each with their own unique features. However, the “maximum goal” in studying certain aquatic ecosystems should be to gain new knowledge about the role and significance of these ecosystems within the broader functional organization of the biosphere. It should also be taken into account that anthropogenic activity significantly affects not only individual ecosystems, but also large subdivisions of the biosphere.
The activity of biotic systems—the pressure of life, as described by C. Darwin and V. Vernadsky—leads to the following two dialectically opposing processes: ecological and evolutionary divergence and convergence. Divergent processes result in an increase in the number of individuals and discrete elements; in the context under consideration, this means an increase in the diversity of ecosystems within the biosphere. The biosphere system becomes enriched with new elements, which enhances the plasticity of the entire system and promotes the expansion and continual modification of the connections between biota and habitat. Convergent processes, on the other hand, lead to a certain unification of system elements, resulting in the formation of a “block” structure within the biosphere. Global evolutionary phenomena in the biosphere manifest as the replacement of entire blocks and transformations of entire biogeomes. Overall, this contributes to an increase in the stability of the biosphere as a whole.
The identification of biogeomes as ecosystem types and as structural elements of the biosphere leads us to consider that anthropogenic activity not only directly impacts the biome of nearly all biogeomes, but also significantly alters the geome by introducing entirely new biotopic elements into the Earth’s surface. This process results in the formation of techno-, agro-, and urban ecosystems, and accordingly, a new anthropogenic biogeome. As human history shows, this biogeome, due to its considerable dependence on human energy expenditure, does not have a high stability (for example, the Mayan cities, now reclaimed by jungle, or the decaying city of Pripyat in the Chernobyl exclusion zone). However, the pathways of noospherogenesis remain unknown to us. In this context, the drastic changes occurring in seemingly stable systems such as the ocean’s pelagic biogeome are deeply concerning; the accumulation of anthropogenic debris in certain areas—driven by ocean circulation [
99]—indicates the persistence of this anomaly.






{kind=link}
{kind=link}
{kind=link}