

Modulation of the Host Defence System by Nematophagous Fungi and Chitosan




Abstract
1. Introduction
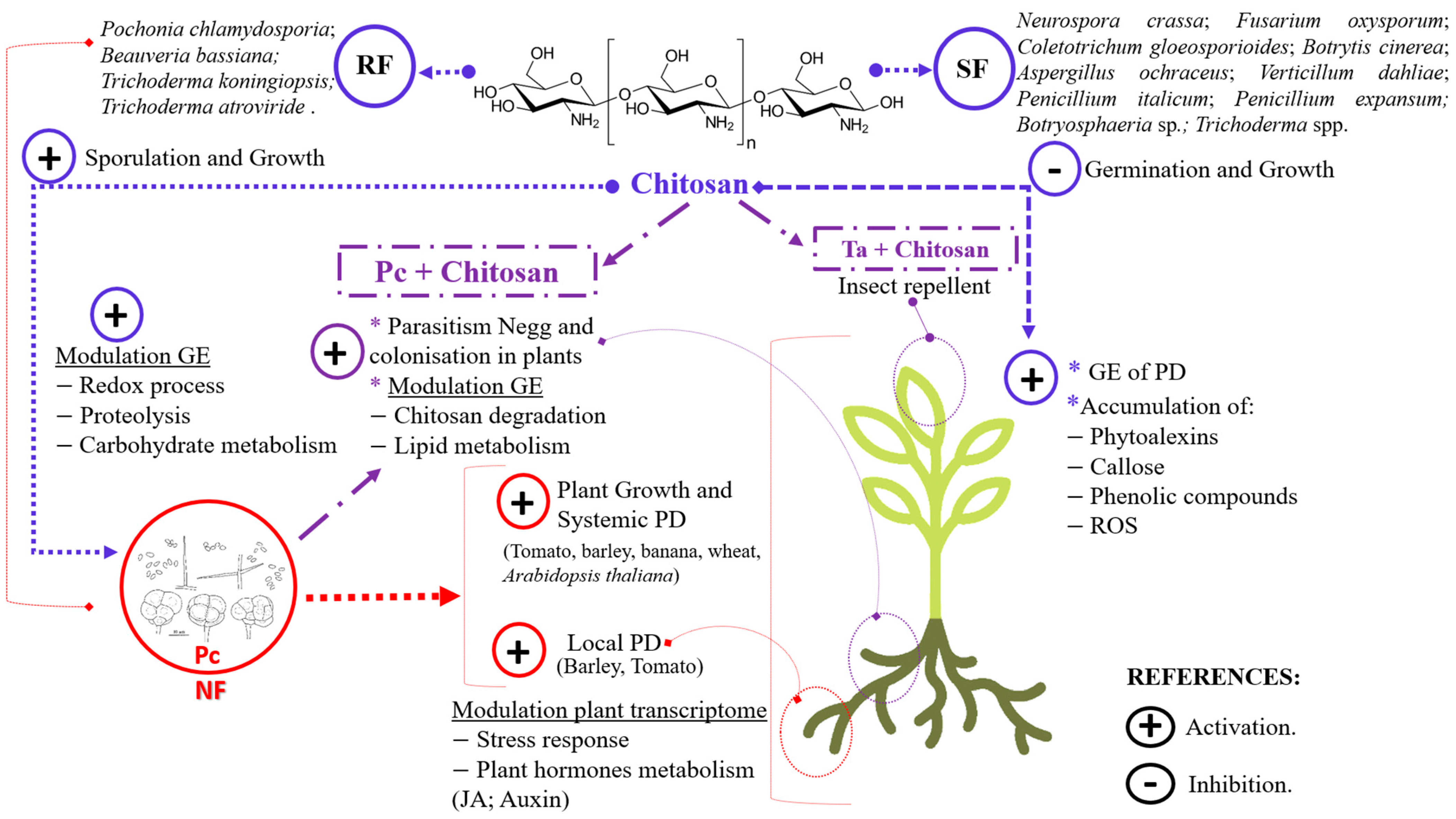
2. Nematophagous Fungi and Chitosan: Growth and Defence Modulators in Plants
3. Plant Defence Avoidance
3.1. Plant Immunity (PI)
3.2. Secreted Proteins and Effectors of Nematophagous Fungi
3.3. Enzymes Involved in the Degradation of Chitin and Chitosan
4. Host Infection and Colonisation by Nematophagous Fungi
5. Multi-Omics Tools for Understanding Integrated Pest Management
5.1. Biocontrol Mechanisms of Fungal BCAs
5.2. Omics to Elucidate BCA–Plant–Pathogen Interaction
5.3. Efficient and Stable BCA Formulates against Pests
6. Closing Remarks and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Bushley, K.E.; Raja, R.; Jaiswal, P.; Cumbie, J.S.; Nonogaki, M.; Boyd, A.E.; Owensby, C.A.; Knaus, B.J.; Elser, J.; Miller, D.; et al. The Genome of Tolypocladium inflatum: Evolution, Organization, and Expression of the Cyclosporin Biosynthetic Gene Cluster. PLoS Genet. 2013, 9, e1003496. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.-H.; Poinar, G.O.; Spatafora, J.W. The oldest fossil evidence of animal parasitism by fungi supports a Cretaceous diversification of fungal–arthropod symbioses. Mol. Phylogenet. Evol. 2008, 49, 495–502. [Google Scholar] [CrossRef]
- Bent, E.; Loffredo, A.; McKenry, M.V.; Becker, J.O.; Borneman, J. Detection and Investigation of Soil Biological Activity against Meloidogyne incognita. J. Nematol. 2008, 40, 109–118. [Google Scholar] [PubMed]
- Lamovsek, J.; Urek, G.; Trdan, S. Biological control of root-knot nematodes (Meloidogyne spp.): Microbes against the pests. Acta Agron. Slov. 2013, 101, 263–275. [Google Scholar] [CrossRef]
- Bordallo, J.J.; Lopez-Llorca, L.V.; Jansson, H.B.; Salinas, J.; Persmark, L.; Asensio, L. Colonization of plant roots by egg-parasitic and nematode-trapping fungi. New Phytol. 2002, 154, 491–499. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, A. Fungi as Biological Control Agents. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.S., Varma, A., Eds.; Springer: Cham, The Netherlands, 2019; Volume 55. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef]
- Grifoll-Romero, L.; Pascual, S.; Aragunde, H.; Biarnés, X.; Planas, A. Chitin Deacetylases: Structures, Specificities, and Biotech Applications. Polymer 2018, 10, 352. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and Chitosan Preparation from Marine Sources. Structure, Properties and Applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef]
- Escudero, N.; Ferreira, S.R.; Lopez-Moya, F.; Naranjo-Ortiz, M.A.; Marin-Ortiz, A.I.; Thornton, C.R.; Lopez-Llorca, L.V. Chitosan enhances parasitism of Meloidogyne javanica eggs by the nematophagous fungus Pochonia chlamydosporia. Fungal Biol. 2016, 120, 572–585. [Google Scholar] [CrossRef]
- Zare, R.; Gams, W.; Evans, H.C. A revision of Verticillium section Prostrata. V. The genus Pochonia, with notes on Rotiferophthora. Nova Hedwig. 2001, 73, 51–86. [Google Scholar] [CrossRef]
- Berlemont, R. Distribution and diversity of enzymes for polysaccharide degradation in fungi. Sci. Rep. 2017, 7, 222. [Google Scholar] [CrossRef] [PubMed]
- Andreou, A.; Giastas, P.; Christoforides, E.; Eliopoulos, E.E. Structural and Evolutionary Insights within the Polysaccharide Deacetylase Gene Family of Bacillus anthracis and Bacillus cereus. Genes 2018, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gao, L.; Xue, C.; Mao, X. Marine-polysaccharide degrading enzymes: Status and prospects. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2767–2796. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.G.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Chitosan, the deacetylated form of chitin, is necessary for cell wall integrity in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, I.A.; Gurr, S.J. Chitosan Mediates Germling Adhesion in Magnaporthe oryzae and Is Required for Surface Sensing and Germling Morphogenesis. PLoS Pathog. 2016, 12, e1005703. [Google Scholar] [CrossRef]
- Christodoulidou, A.; Bouriotis, V.; Thireos, G. Two Sporulation-specific Chitin Deacetylase-encoding Genes Are Required for the Ascospore Wall Rigidity of Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 31420–31425. [Google Scholar] [CrossRef]
- Sánchez-Vallet, A.; Mesters, J.R.; Thomma, B.P. The battle for chitin recognition in plant-microbe interactions. FEMS Microbiol. Rev. 2015, 39, 171–183. [Google Scholar] [CrossRef]
- White, S.; McIntyre, M.; Berry, D.R.; McNeil, B. The autolysis of industrial filamentous fungi. Crit. Rev. Biotechnol. 2002, 22, 1–14. [Google Scholar] [CrossRef]
- Zhao, Y.; Park, R.-D.; Muzzarelli, R.A. Chitin deacetylases: Properties and applications. Mar. Drugs 2010, 8, 24–46. [Google Scholar] [CrossRef]
- Cord-Landwehr, S.; Melcher, R.L.; Kolkenbrock, S.; Moerschbacher, B.M. A chitin deacetylase from the endophytic fungus Pestalotiopsis sp. efficiently inactivates the elicitor activity of chitin oligomers in rice cells. Sci. Rep. 2016, 6, 38018. [Google Scholar] [CrossRef]
- Liu, T.; Chen, Y.; Tian, S.; Li, B. Crucial Roles of Effectors in Interactions between Horticultural Crops and Pathogens. Horticulturae 2023, 9, 250. [Google Scholar] [CrossRef]
- Lopez-Llorca, L.V.; Gómez-Vidal, S.; Monfort, E.; Larriba, E.; Casado-Vela, J.; Elortza, F.; Jansson, H.B.; Salinas, J.; Martín-Nieto, J. Expression of serine proteases in egg-parasitic nematophagous fungi during barley root colonization. Fungal Genet. Biol. 2010, 47, 342–351. [Google Scholar] [CrossRef]
- Larriba, E.; Jaime, M.D.; Carbonell-Caballero, J.; Conesa, A.; Dopazo, J.; Nislow, C.; Martín-Nieto, J.; Lopez-Llorca, L.V. Sequencing and functional analysis of the genome of a nematode egg-parasitic fungus, Pochonia chlamydosporia. Fungal Genet. Biol. 2014, 65, 69–80. [Google Scholar] [CrossRef]
- Aranda-Martinez, A.; Lenfant, N.; Escudero, N.; Zavala-Gonzalez, E.A.; Henrissat, B.; Lopez-Llorca, L.V. CAZyme content of Pochonia chlamydosporia reflects that chitin and chitosan modification are involved in nematode parasitism. Environ. Microbiol. 2016, 18, 4200–4215. [Google Scholar] [CrossRef]
- Aranda-Martinez, A.; Grifoll-Romero, L.; Aragunde, H.; Sancho-Vaello, E.; Biarnés, X.; Lopez- Llorca, L.V.; Planas, A. Expression and specificity of a chitin deacetylase from the nematophagous fungus Pochonia chlamydosporia potentially involved in pathogenicity. Sci. Rep. 2018, 8, 2170. [Google Scholar] [CrossRef]
- Suarez-Fernandez, M.; Sambles, C.; Lopez-Moya, F.; Nueda, M.J.; Studholme, D.J.; Lopez-Llorca, L.V. Chitosan modulates Pochonia chlamydosporia gene expression during nematode egg parasitism. Environ. Microbiol. 2021, 23, 4980–4997. [Google Scholar] [CrossRef]
- Suarez-Fernandez, M.; Aragon-Perez, A.; Lopez-Llorca, L.V.; Lopez-Moya, F. Putative LysM Effectors Contribute to Fungal Lifestyle. Int. J. Mol. Sci. 2021, 22, 3147. [Google Scholar] [CrossRef]
- Mingot-Ureta, C.; Lopez-Moya, F.; Lopez-Llorca, L.V. Isolates of the Nematophagous Fungus Pochonia chlamydosporia Are Endophytic in Banana Roots and Promote Plant Growth. Agronomy 2020, 10, 1299. [Google Scholar] [CrossRef]
- Manzanilla-López, R.H.; Esteves, I.; Powers, S.J.; Kerry, B.R. Effects of crop plants on abundance of Pochonia chlamydosporia and other fungal parasites of root-knot and potato cyst nematodes. Ann. Appl. Biol. 2011, 159, 118–129. [Google Scholar] [CrossRef]
- Maciá-Vicente, J.G.; Rosso, L.C.; Ciancio, A.; Jansson, H.-B.; Lopez-Llorca, L.V. Colonisation of barley roots by endophytic Fusarium equiseti and Pochonia chlamydosporia: Effects on plant growth and disease. Ann. Appl. Biol 2009, 155, 391–401. [Google Scholar] [CrossRef]
- Zavala-Gonzalez, E.A.; Rodríguez-Cazorla, E.; Escudero, N.; Aranda-Martinez, A.; Martínez-Laborda, A.; Ramírez-Lepe, M.; Vera, A.; Lopez-Llorca, L.V. Arabidopsis thaliana root colonization by the nematophagous fungus Pochonia chlamydosporia is modulated by jasmonate signaling and leads to accelerated flowering and improved yield. New Phytol. 2017, 213, 351–364. [Google Scholar] [CrossRef]
- Zavala-González, E.A.; Escudero, N.; Lopez-Moya, F.; Aranda-Martinez, A.; Exposito, A.; Ricaño-Rodríguez, J.; Naranjo-Ortiz, M.A.; Ramírez-Lepe, M.; Lopez-Llorca, L.V. Some isolates of the nematophagous fungus Pochonia chlamydosporia promote root growth and reduce flowering time of tomato. Ann. Appl. Biol. 2015, 166, 472–483. [Google Scholar] [CrossRef]
- Larriba, E.; Jaime, M.D.; Nislow, C.; Martín-Nieto, J.; Lopez-Llorca, L.V. Endophytic colonization of barley (Hordeum vulgare) roots by the nematophagous fungus Pochonia chlamydosporia reveals plant growth promotion and a general defense and stress transcriptomic response. J. Plant Res. 2015, 128, 665–678. [Google Scholar] [CrossRef]
- Ghahremani, Z.; Escudero, N.; Saus, E.; Gabaldón, T.; Sorribas, F.J. Pochonia chlamydosporia induces plant-dependent systemic resistance to Meloidogyne incognita. Front. Plant Sci. 2019, 10, 945. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2019, 38, 651–664. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Lopez-Jimenez, J.A.; Pérez-Berná, A.J.; Huang, I.-C.; Jansson, H.-B.; Salinas, J.; Villalaín, J.; Read, N.D.; Lopez-Llorca, L.V. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol. Microbiol. 2010, 75, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Moya, F.; Kowbel, D.; Nueda, M.J.; Palma-Guerrero, J.; Glass, N.L.; Lopez- Llorca, L.V. Neurospora crassa transcriptomics reveals oxidative stress and plasma membrane homeostasis biology genes as key targets in response to chitosan. Mol. BioSyst. 2016, 12, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Zavala-González, E.A.; Lopez-Moya, F.; Aranda-Martinez, A.; Cruz-Valerio, M.; Lopez-Llorca, L.V.; Ramírez-Lepe, M. Tolerance to chitosan by Trichoderma species is associated with low membrane fluidity. J. Basic Microbiol. 2016, 56, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, S.; Meng, X.; Xu, Y. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest Biol. Technol. 2007, 44, 300–306. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Jansson, H.-B.; Salinas, J.; Lopez-Llorca, L.V. Effect of chitosan on hyphal growth and spore germination of plant pathogenic and biocontrol fungi. J. Appl. Microbiol. 2008, 104, 541–553. [Google Scholar] [CrossRef]
- Lee, C.G.; Koo, J.C.; Park, J.K. Antifungal Effect of Chitosan as Ca2+ Channel Blocker. Plant Pathol. J. 2016, 32, 242–250. [Google Scholar] [CrossRef]
- Wang, Y.; Li, B.; Zhang, X.; Peng, N.; Mei, Y.; Liang, Y. Low molecular weight chitosan is an effective antifungal agent against Botryosphaeria sp. and preservative agent for pear (Pyrus) fruits. Int. J. Biol. Macromol. 2017, 95, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Xoca-Orozco, L.-Á.; Cuellar-Torres, E.A.; González-Morales, S.; Gutiérrez-Martínez, P.; López-García, U.; Herrera-Estrella, L.; Vega-Arreguín, J.; Chacón-López, A. Transcriptomic Analysis of Avocado Hass (Persea americana Mill) in the Interaction System Fruit-Chitosan-Colletotrichum. Front. Plant Sci. 2017, 8, 956. [Google Scholar] [CrossRef]
- Meng, D.; Garba, B.; Ren, Y.; Yao, M.; Xia, X.; Li, M.; Wang, Y. Antifungal activity of chitosan against Aspergillus ochraceus and its possible mechanisms of action. Int. J. Biol. Macromol. 2020, 158, 1063–1070. [Google Scholar] [CrossRef]
- Riseh, R.S.; Hassanisaadi, M.; Vatankhah, M.; Babaki, S.A.; Barka, E.A. Chitosan as a potential natural compound to manage plant diseases. Int. J. Biol. Macromol. 2022, 220, 998–1009. [Google Scholar] [CrossRef]
- Orzali, L.; Corsi, B.; Forni, C.; Riccioni, L. Chitosan in agriculture: A new challenge for managing plant disease. In Biological Activities and Application of Marine Polysaccharides; Shalaby, E., Ed.; InTech: London, UK, 2017; pp. 17–36. [Google Scholar]
- Hadwiger, L.A. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant-Microbe Interact. 2011, 24, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Narasimhamurthy, K.; Udayashankar, A.C.; De Britto, S.; Lavanya, S.N.; Abdelrahman, M.; Soumya, K.; Shetty, H.S.; Srinivas, C.; Jogaiah, S. Chitosan and chitosan-derived nanoparticles modulate enhanced immune response in tomato against bacterial wilt disease. Int. J. Biol. Macromol. 2022, 220, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Escudero, N.; Lopez-Moya, F.; Ghahremani, Z.; Zavala-Gonzalez, E.A.; Alaguero-Cordovilla, A.; Ros-Ibañez, C.; Lacasa, A.; Sorribas, F.J.; Lopez-Llorca, L.V. Chitosan Increases Tomato Root Colonization by Pochonia chlamydosporia and Their Combination Reduces Root-Knot Nematode Damage. Front. Plant Sci. 2017, 8, 1415. [Google Scholar] [CrossRef]
- Kappel, L.; Kosa, N.; Gruber, S. The Multilateral Efficacy of Chitosan and Trichoderma on Sugar Beet. J. Fungi 2022, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Nürnberger, T.; Kemmerling, B. Receptor protein kinases—Pattern recognition receptors in plant immunity. Trends Plant Sci. 2006, 11, 519–522. [Google Scholar] [CrossRef]
- Lu, Y.; Tsuda, K. Intimate Association of PRR- and NLR-Mediated Signalling in Plant Immunity. Mol. Plant-Microbe Interact. 2021, 34, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-M.; Zhang, Y. Plant Immunity: Danger Perception and Signalling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, M.; Yu, L.; Zhou, Z.; Liang, X.; Liu, Z.; Cai, G.; Gao, L.; Zhang, X.; Wang, Y.; et al. The FLS2-Associated Kinase BIK1 Directly Phosphorylates the NADPH Oxidase RbohD to Control Plant Immunity. Cell Host Microbe 2014, 15, 329–338. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe Interactions: Shaping the Evolution of the Plant Immune Response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef]
- Jones, J.; Dangl, J. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Newman, M.-A.; Sundelin, T.; Nielsen, J.T.; Erbs, G. MAMP (microbe-associated molecular pattern) triggered immunity in plants. Front. Plant Sci. 2013, 4, 139. [Google Scholar] [CrossRef]
- Yu, X.; Feng, B.; He, P.; Shan, L. From Chaos to Harmony: Responses and Signalling upon Microbial Pattern Recognition. Annu. Rev. Phytopathol. 2017, 55, 109–137. [Google Scholar] [CrossRef]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The Role of the Fungal Cell Wall in the Infection of Plants. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef]
- Felix, G.; Regenass, M.; Boller, T. Specific perception of subnanomolar concentrations of chitin fragments by tomato cells: Induction of extracellular alkalinization, changes in protein phosphorylation, and establishment of a refractory state. Plant J. 1993, 4, 307–316. [Google Scholar] [CrossRef]
- Baureithel, K.; Felix, G.; Boller, T. Specific, high affinity binding of chitin fragments to tomato cells and membranes. Competitive inhibition of binding by derivates of chitooligosaccharides and Nod factor of Rhizobium. J. Biol. Chem. 1994, 269, 17931–17938. [Google Scholar]
- Vander, P.; Vårum, K.M.; Domard, A.; Eddine El Gueddari, N.; Moerschbacher, B.M. Comparison of the ability of partially N-acetylated chitosans and chitooligosaccharides to elicit resistance reactions in wheat leaves. Plant Physiol. 1998, 118, 1353–1359. [Google Scholar] [CrossRef]
- Li, P.; Linhardt, R.J.; Cao, Z. Structural characterization of oligochitosan elicitor from Fusarium sambucinum and its elicitation of defensive responses in Zanthoxylum bungeanum. Int. J. Mol. Sci. 2016, 7, 2076. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Bravo, J.M.; Campo, S.; Murillo, I.; Coca, M.; San Segundo, B. Fungus- and wound-induced accumulation of mRNA containing a class II chitinase of the pathogenesis-related protein 4 (PR-4) family of maize. Plant Mol. Biol. 2003, 52, 745–759. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Escudero, N.; Zavala-González, E.A.; Esteve-Bruna, D.; Blázquez, M.A.; Alabadí, D.; Lopez-Llorca, L.V. Induction of auxin biosynthesis and WOX5 repression mediate changes in root development in Arabidopsis exposed to chitosan. Sci. Rep. 2017, 7, 16813. [Google Scholar] [CrossRef] [PubMed]
- Beatrice, C.; Linthorst, J.H.; Cinzia, F.; Luca, R. Enhancement of PR1 and PR5 gene expressions by chitosan treatment in kiwifruit plants inoculated with Pseudomonas syringae pv. actinidiae. Eur. J. Plant Pathol. 2017, 148, 163–179. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, B.-S.; Zhao, J.-H.; Huang, J.-F.; Jia, P.-S.; Wang, S.; Zhang, J.; Zhou, J.-M.; Guo, H.-S. Deacetylation of chitin oligomers increases virulence in soil-borne fungal pathogens. Nat. Plants 2019, 5, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-P.; Li, J.-J.; Dhar, N.; Li, J.-P.; Chen, J.-Y.; Jian, W.; Dai, X.-F.; Yang, X.-F. Lysin Motif (LysM) Proteins: Interlinking Manipulation of Plant Immunity and Fungi. Int. J. Mol. Sci. 2021, 22, 3114. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Zhang, C.; Wang, M.; Zhang, C.; Xu, X.; Huang, Y.; Chen, Y.; Lin, Y.; Lai, Z. Genome-wide identification, evolution analysis of LysM gene family members and their expression analysis in response to biotic and abiotic stresses in banana (Musa L.). Gene 2022, 845, 146849. [Google Scholar] [CrossRef]
- Bolton, M.D.; van Esse, H.P.; Vossen, J.H.; de Jonge, R.; Stergiopoulos, I.; Stulemeijer, I.J.; van Den Berg, G.C.M.; Borrás-Hidalgo, O.; Dekker, H.L.; de Koster, C.G.; et al. The novel Cladosporium fulvum lysin motif effector Ecp6 is a virulence factor with orthologues in other fungal species. Mol. Microbiol. 2008, 69, 119–136. [Google Scholar] [CrossRef]
- de Jonge, R.; van Esse, H.P.; Kombrink, A.; Shinya, T.; Desaki, Y.; Bours, R.; van Der Krol, S.; Shibuya, N.; Joosten, M.H.A.J.; Thomma, B.P. Conserved Fungal LysM Effector Ecp6 Prevents Chitin-Triggered Immunity in Plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]
- Marshall, R.; Kombrink, A.; Motteram, J.; Loza-Reyes, E.; Lucas, J.; Hammond-Kosack, K.E.; Thomma, B.P.; Rudd, J.J. Analysis of Two in Planta Expressed LysM Effector Homologs from the Fungus Mycosphaerella graminicola Reveals Novel Functional Properties and Varying Contributions to Virulence on Wheat. Plant Physiol. 2011, 156, 756–769. [Google Scholar] [CrossRef]
- Tian, H.; MacKenzie, C.I.; Rodriguez-Moreno, L.; van den Berg, G.C.; Chen, H.; Rudd, J.J.; Mesters, J.R.; Thomma, B.P. Three LysM effectors of Zymoseptoria tritici collectively disarm chitin-triggered plant immunity. Mol. Plant Pathol. 2021, 22, 683–693. [Google Scholar] [CrossRef]
- Lee, W.-S.; Rudd, J.J.; Hammond-Kosack, K.E.; Kanyuka, K. Mycosphaerella graminicola LysM Effector-Mediated Stealth Pathogenesis Subverts Recognition Through Both CERK1 and CEBiP Homologues in Wheat. Mol. Plant-Microbe Interact. 2014, 27, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P.; et al. Effector-Mediated Suppression of Chitin-Triggered Immunity by Magnaporthe oryzae Is Necessary for Rice Blast Disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Takahara, H.; Hacquard, S.; KombrinK, A.; Hughes, H.B.; Halder, V.; Robin, G.P.; Hiruma, K.; Neumann, U.; Shinya, T.; Kombrink, E.; et al. Colletotrichum higginsianum extracellular LysM proteins play dual roles in appressorial function and suppression of chitin-triggered plant immunity. New Phytol. 2016, 211, 1323–1337. [Google Scholar] [CrossRef] [PubMed]
- Kombrink, A.; Rovenich, H.; Shi-Kunne, X.; Rojas-Padilla, E.; van den Berg, G.C.; Domazakis, E.; de Jonge, R.; Valkenburg, D.-J.; Sánchez-Vallet, A.; Seidl, M.F.; et al. Verticillium dahliae LysM effectors differentially contribute to virulence on plant hosts. Mol. Plant Pathol. 2017, 18, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Dölfors, F.; Holmquist, L.; Dixelius, C.; Tzelepis, G. A LysM effector protein from the basidiomycete Rhizoctonia solani contributes to virulence through suppression of chitin-triggered immunity. Mol. Genet. Genom. 2019, 294, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liao, Z.; Feng, L.; An, B.; He, C.; Wang, Q.; Luo, H. Colletotrichum gloeosporioides Cg2LysM contributed to virulence toward rubber tree through affecting invasive structure and inhibiting chitin-triggered plant immunity. Front. Microbiol. 2023, 14, 1129101. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Qin, F.; Shen, B.; Shi, Q.; Liu, C.; Zhang, X.; Jiao, Y.; Lu, J.; Gao, Y.; Suarez-Fernandez, M.; et al. Genome and secretome analysis of Pochonia chlamydosporia provide new insight into egg-parasitic mechanisms. Sci. Rep. 2018, 8, 1123. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Vidal-Diez de Ulzurrum, G.; Schwarz, E.M. Genome sequence of the oyster mushroom Pleurotua ostreatus strain PC9. G3 2021, 11, jkaa008. [Google Scholar] [CrossRef]
- Boontawon, T.; Nakazawa, T.; Inoue, C.; Osakabe, K.; Kawauchi, M.; Sakamoto, M.; Hond, Y. Efficient genome editing with CRISPR/Cas9 in Pleurotus ostreatus. AMB Expr. 2021, 11, 30. [Google Scholar] [CrossRef]
- Zhang, Y.; Qiao, M.; Xu, J.; Cao, Y.; Zhang, K.-Q.; Yu, Z.-F. Genetic diversity and recombination in natural populations of the nematode-trapping fungus Arthrobotrys oligospora from China. Ecol. Evol. 2013, 3, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Cen, K.; Li, B.; Lu, Y.; Zhang, S.; Wang, C. Divergent LysM effectors contribute to the virulence of Beauveria bassiana by evasion of insect immune defenses. PLoS Pathog. 2017, 13, e1006604. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. The insect-pathogenic fungus Metarhizium robertsii (Clavicipitaceae) is also an endophyte that stimulates plant root development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- Zeng, T.; Rodriguez-Moreno, L.; Mansurkhodzaev, A.; Wang, P.; van den Berg, W.; Gasciolli, V.; Cottaz, S.; Fort, S.; Thomma, B.P.; Bono, J.-J.; et al. A lysin motif effector subverts chitin-triggered immunity to facilitate arbuscular mycorrhizal symbiosis. New Phytol. 2020, 225, 448–460. [Google Scholar] [CrossRef]
- Romero-Contreras, Y.J.; Ramírez-Valdespino, C.A.; Guzmán-Guzmán, P.; Macías-Segoviano, J.I.; Villagómez-Castro, J.C.; Olmedo-Monfil, V. Tal6 From Trichoderma atroviride Is a LysM Effector Involved in Mycoparasitism and Plant Association. Front. Microbiol. 2019, 10, 2231. [Google Scholar] [CrossRef]
- Seidl-Seiboth, V.; Zach, S.; Frischmann, A.; Spadiut, O.; Dietzsch, C.; Herwig, C.; Ruth, C.; Rodler, A.; Jungbauer, A.; Kubicek, C.P. Spore germination of Trichoderma atroviride is inhibited by its LysM protein TAL6. FEBS J. 2013, 280, 1226–1236. [Google Scholar] [CrossRef]
- Nahar, P.; Ghormade, V.; Deshpande, M.V. The extracellular constitutive production of chitin deacetylase in Metarhizium anisopliae: Possible edge to entomopathogenic fungi in the biological control of insect pests. J. Invertebr. Pathol. 2004, 85, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Hiramatsu, S.; Songsri, P.; Fujie, M. Alternative Expression of a Chitosanase Gene Produces Two Different Proteins in Cells Infected with Chlorella Virus CVK2. Virology 1997, 230, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, B.; Li, C.; Bao, X. Knock down of chitosanase expression in phytopathogenic fungus Fusarium solani and its effect on pathogenicity. Curr. Genet. 2010, 56, 275–281. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, R.; Bolton, M.D.; Thomma, B.P. How filamentous pathogens co-opt plants: The ins and outs of fungal effectors. Curr. Opin. Plant Biol. 2011, 14, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Bharagava, R.N.; Chowdhary, P. Emerging and Eco-Friendly Approaches for Waste Management; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Gust, A.A.; Willmann, R.; Desaki, Y.; Grabherr, H.M.; Nürnberger, T. Plant LysM proteins: Modules mediating symbiosis and immunity. Trends Plant Sci. 2012, 17, 495–502. [Google Scholar] [CrossRef]
- Zhao, S.; An, B.; Guo, Y.; Hou, X.; Luo, H.; He, C.; Wang, Q. Label free proteomics and systematic analysis of secretome reveals effector candidates regulated by SGE1 and FTF1 in the plant pathogen Fusarium oxysporum f. sp. cubense tropical race 4. BMC Genom. 2020, 21, 275. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Gómez-Vidal, S.; Tikhonov, V.E.; Salinas, J.; Jansson, H.-B.; Lopez-Llorca, L.V. Comparative analysis of extracellular proteins from Pochonia chlamydosporia grown with chitosan or chitin as main carbon and nitrogen sources. Enzym. Microb. Technol. 2010, 46, 568–574. [Google Scholar] [CrossRef]
- Thadathil, N.; Velappan, S.P. Recent developments in chitosanase research and its biotechnological applications: A review. Food Chem. 2014, 150, 392–399. [Google Scholar] [CrossRef]
- Grenier, J.; Asselin, A. Some Pathogenesis-Related Proteins Are Chitosanases with Lytic Activity against Fungal Spores. Mol. Plant-Microbe Interact. 1990, 3, 401–407. [Google Scholar] [CrossRef]
- Hirano, Y.; Yamamoto, R.; Dannoura, M.; Aono, K.; Igarashi, T.; Ishii, M.; Yamase, K.; Makita, N.; Kanazawa, Y. Detection frequency of Pinus thunbergii roots by ground-penetrating radar is related to root biomass. Plant Soil 2012, 360, 363–373. [Google Scholar] [CrossRef]
- Sambles, C.; Suarez-Fernandez, M.; Lopez-Moya, F.; Lopez-Llorca, L.V.; Studholme, D.J. Chitosan induces differential transcript usage of chitosanase 3 encoding gene (csn3) in the biocontrol fungus Pochonia chlamydosporia 123. BMC Genom. 2022, 23, 101. [Google Scholar] [CrossRef]
- Larriba, E.; Martín-Nieto, J.; Lopez-Llorca, L.V. Gene cloning, molecular modeling, and phylogenetics of serine protease P32 and serine carboxypeptidase SCP1 from nematophagous fungi Pochonia rubescens and Pochonia chlamydosporia. Can. J. Microbiol. 2012, 58, 815–827. [Google Scholar] [CrossRef]
- Medie, F.M.; Davies, G.J.; Drancourt, M.; Henrissat, B. Genome analyses highlight the different biological roles of cellulases. Nat. Rev. Microbiol. 2012, 10, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Schmoll, M.; Cate, J.H.; Coradetti, S. Plant cell wall deconstruction by ascomycete fungi. Annu. Rev. Microbiol. 2013, 67, 477–498. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. NAR 2014, 42, 490–495. [Google Scholar] [CrossRef]
- Moreno-García, J.; Martín-García, F.J.; Ogawa, M.; García-Martínez, T.; Moreno, J.; Mauricio, J.C.; Bisson, L.F. FLO1, FLO5 and FLO11 flocculation gene expression impacts Saccharomyces cerevisiae attachment to Penicillium chrysogenum in a co-immobilization technique. Front. Microbiol. 2018, 9, 2586. [Google Scholar] [CrossRef] [PubMed]
- Rosso, L.C.; Finetti-Sialer, M.M.; Hirsch, P.R.; Ciancio, A.; Kerry, B.R.; Clark, I.M. Transcriptome analysis shows differential gene expression in the saprotrophic to parasitic transition of Pochonia chlamydosporia. Appl. Microbiol. Biot. 2011, 90, 1981–1994. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J. Fungal Cell Wall: Structure, Synthesis, and Assembly; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Brown, V.; Sabina, J.; Johnston, M. Specialized sugar sensing in diverse fungi. Curr. Biol. 2009, 19, 436–441. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Karaffa, L.; Fekete, E.; Gamauf, C.; Szentirmai, A.; Kubicek, C.P.; Seiboth, B. D-galactose induces cellulase gene expression in Hypocrea jecorina at low growth rates. Microbiology 2006, 152, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Nordbring-Hertz, B.; Friman, E.; Mattiasson, B. A recognition mechanism in the adhesion of nematodes to nematode-trapping fungi. In Lectins-Biology, Biochemistry and Clinical Biochemistry; Bog-Hansen, T.C., Ed.; Walter de Gruyter: Berlin, Germany, 1982; Volume 2, pp. 83–90. [Google Scholar]
- Premachandran, D.; Pramer, D. Role of N-acetylgalactosamine specific protein in trapping of nematodes by Arthrobotrys oligospora. Appl. Environ. Microbiol. 1984, 47, 1358–1359. [Google Scholar] [CrossRef]
- Sharon, E.; Spiegel, Y. Glycoprotein characterization of the gelatinous matrix in the root-knot nematode Meloidogyne javanica. J. Nematol. 1993, 25, 585–589. [Google Scholar]
- Clarke, A.J. The composition of the cyst wall of the beet cyst-nematode Heterodera schachtii. Biochem. J. 1970, 118, 315–318. [Google Scholar] [CrossRef][Green Version]
- Forrest, J.M.S.; Robertson, W.M. Characterization and localization of saccharides on the head region of four populations of the potato cyst nematode Globodera rostochiensis and G. pallida. J. Nematol. 1986, 18, 23. [Google Scholar] [PubMed]
- Baker, K.; Cook, R.J. Biological Control of Plant Pathogens; W.H. Freeman and Company: San Francisco, CA, USA, 1974. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Motlagh, M.R.; Samimi, Z. Evaluation of Trichoderma spp., as biological agents in some of plant pathogens. Ann. Biol. Res. 2013, 4, 173–179. [Google Scholar]
- Quevedo, A.C.; Muniz, M.F.; Savian, L.G.; Sarzi, J.S.; Saldanha, M.A. Ação antagonista in vitro de Trichoderma spp. sobre Fusarium oxysporum. Ciência Florest. 2022, 32, 2288–2303. [Google Scholar] [CrossRef]
- Asad, S.A.; Ali, N.; Hameed, A.; Khan, S.A.; Ahmad, R.; Bilal, M.; Shahzad, M.; Tabassum, A. Biocontrol efficacy of different isolates of Trichoderma against soil borne pathogen Rhizoctonia solani. Pol. J. Microbiol. 2014, 63, 95–103. [Google Scholar] [CrossRef]
- Inamdar, A.A.; Morath, S.; Bennett, J.W. Fungal Volatile Organic Compounds: More Than Just a Funky Smell? Annu. Rev. Microbiol. 2020, 74, 101–116. [Google Scholar] [CrossRef]
- Lozano-Soria, A.; Picciotti, U.; Lopez-Moya, F.; Lopez-Cepero, J.; Porcelli, F.; Lopez-Llorca, L.V. Volatile Organic Compounds from Entomopathogenic and Nematophagous Fungi, Repel Banana Black Weevil (Cosmopolites sordidus). Insects 2020, 11, 509. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Harman, G.E. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; López-Bucio, J.S.; Méndez-Bravo, A.; Macías-Rodríguez, L.; Ramos-Vega, M.; Guevara-García, Á.A.; López-Bucio, J. Mitogen-Activated Protein Kinase 6 and Ethylene and Auxin Signaling Pathways Are Involved in Arabidopsis Root-System Architecture Alterations by Trichoderma atroviride. Mol. Plant-Microbe Interact. 2015, 28, 701–710. [Google Scholar] [CrossRef]
- Tjamos, E.C. Selective Elimination of Soilborne Plant Pathogens and Enhancement of Antagonists by Steaming, Sublethal Fumigation and Soil Solarization. In Biological Control of Plant Diseases; Tjamos, E.C., Papavizas, G.C., Cook, R.J., Eds.; NATO ASI Series; Springer: Boston, MA, USA, 1992; Volume 230. [Google Scholar]
- Constantin, M.E.; de Lamo, F.J.; Vlieger, B.V.; Rep, M.; Takken, F.L. Endophyte-Mediated Resistance in Tomato to Fusarium oxysporum Is Independent of ET, JA, and SA. Front. Plant Sci. 2019, 10, 979. [Google Scholar] [CrossRef]
- Afshan, N.-U.-S. Recent Advancement in Fungal Biocontrol Agents. In Plant Mycobiome; Rashad, Y.M., Baka, Z.A., Moussa, T.A., Eds.; Springer: Cham, Switzerland, 2023; pp. 203–223. [Google Scholar]
- Uchida, M.; Konishi, T.; Fujigasaki, A.; Kita, K.; Arie, T.; Teraoka, T.; Kanda, Y.; Mori, M.; Arazoe, T.; Kamakura, T. Dysfunctional Pro1 leads to female sterility in rice blast fungi. iScience 2023, 26, 107020. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Xiao, J.; Dai, L.; Huang, Y.; Mao, Z.; Lin, R.; Yao, Y.; Xie, B. Development of a high-efficiency gene knockout system for Pochonia chlamydosporia. Microbiol. Res. 2014, 170, 18–26. [Google Scholar] [CrossRef]
- Li, D.; Tang, Y.; Lin, J.; Cai, W. Methods for genetic transformation of filamentous fungi. Microb. Cell Fact. 2017, 16, 168. [Google Scholar] [CrossRef] [PubMed]
- Lichius, A.; Ruiz, D.M.; Zeilinger, S. Genetic Transformation of Filamentous Fungi: Achievements and Challenges. In Grand Challenges in Fungal Biotechnology. Grand Challenges in Biology and Biotechnology; Nevalainen, H., Ed.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Zhang, M.; Jiang, S.; Zheng, J.; Zheng, Z.; Li, X.; Pan, L.; Luo, S. Construction of an integration vector carrying hygromycin B resistance gene and its genetic transformation in Rhizopus oryzae. Chin. J. Biotechnol. 2015, 31, 1203–1218. [Google Scholar]
- Nødvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.-Q.; Liu, G.-N.; Ji, R.-Y.; Shi, K.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. CRISPR/Cas9-based genome editing of the filamentous fungi: The state of the art. Appl. Microbiol. Biotechnol. 2017, 101, 7435–7443. [Google Scholar] [CrossRef]
- Matsu-ura, T.; Baek, M.; Kwon, J.; Hong, C. Efficient gene editing in Neurospora crassa with CRISPR technology. Fungal Biol. Biotechnol. 2015, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [PubMed]
- Astudillo-Echeverría, A.; Pazmiño-Centeno, D.; Naranjo-Briceño, L. Uso de CRISPR/Cas9 como herramienta de edición de genomas en hongos filamentosos: Una revisión del estado actual y últimas tendencias. Genética Médica Y Genómica 2021, 5. [Google Scholar]
- Wang, Q.; Coleman, J.J. Progress and Challenges: Development and Implementation of CRISPR/Cas9 Technology in Filamentous Fungi. Comput. Struct. Biotechnol. J. 2019, 17, 761–769. [Google Scholar] [CrossRef]
- Al Abdallah, Q.; Ge, W.; Fortwendel, J.R. A simple and universal system for gene manipulation in Aspergillus fumigatus: In vitro-assembled Cas9-guide RNA ribonucleoproteins coupled with microhomology repair templates. Msphere 2017, 2, e00446-17. [Google Scholar] [CrossRef]
- Ferrara, M.; Haidukowski, M.; Logrieco, A.F.; Leslie, J.F.; Mulè, G. A CRISPR-Cas9 System for Genome Editing of Fusarium proliferatum. Sci. Rep. 2019, 9, 19836. [Google Scholar] [CrossRef]


{kind=link}
| Fungus | Lifestyle | Host | Protein | Omics Strategies | References |
|---|---|---|---|---|---|
| LysM effectors | |||||
| Cladosporium fulvum | PF | tomato | Ecp6; Ecp7 | 2D-PAGE; MS; CG; AMT; Pl-C; qRT-PCR; HE-P; GW; AV; PRP | [74,75] |
| Zymoseptoria Tritici | PF | wheat | Mg1LysM; Mg2LysM; Mg3LysM; Mgx1LysM | qRT-PCR; HE-P: AMT; I-DNA; SB; IRP-K-out M; Pan; MoD | [29,76,77,78] |
| Magnaporthe oryzae | PF | rice | Slp1 | gene-GFP; G-I-Transg-cult; Pan; LM; EM; PRP | [79] |
| Colletotrichum higginsianum | PF | cruciferous crops; Arabidopsis thaliana | ChELP1; ChELP2 | Prot-S; Pan; qRT-PCR; RNA-seq; PRP; Gly-An; c-rAb; LM; ITEM; lec-C | [80] |
| Verticillum dahliae | PF | tomato; A. thaliana; Nicotiana benthamiana | Vd2LysM; Vd4LysM; Vd5LysM; Vd6LysM | qRT-PCR; Fan; HE-P; plant-PP; PRP | [81] |
| Rhizoctonia solani | PF | soybean; potato; tobacco; rice sugar beet | RsLysM | qRT-PCR; CTS; Vass; HE-P | [82] |
| Colletrotichum gloeosporioides | PF | rubber tree | Cg2LysM | qRT-PCR; G-K-out M; PE-plant | [83] |
| Pochonia chlamydosporia strain 123 | NF | A. thaliana; wheat; Meloidogyne javanica; tomato banana; barley | Pc123_Lys-1 Pc123_Lys-2 Pc123_Lys-3 Pc123_Lys-4 | GS; IPE; IPD; M-3D-SAAS; Pan; MD; qRT-PCR | [25,29] |
| Pochonia chlamydosporia strain 170 | NF | M. incognita | Pc170_1 Pc170_2 | GS; D-AS; TPAn; Pan; MD | [29,84] |
| Pleurotus ostreatus | NF | M. incognita | PlO-1 | GS; CRISPR-Cas 9; rc-PCR; PCR- IM; Pan; MD | [29,85,86] |
| Arthrobotrys oligospora | NF | Ditylenchus dipsaci; M. incognita | ArO-1 ArO-2 ArO-3 | MI-GLC; I-SNPs; PS; AV; CRAn; Pan; MD | [29,87] |
| Beauveria bassiana | EPF | insects | Blys2; Blys4; Blys5; Blys6; Blys7; Blys8 | AMT; qRT-PCR; ProtL; WB; Pan; MD | [29,88] |
| Metarhizium robertsii ARSEF | EPF | insects | Mr-1; Mr-2; Mr-3; Mr-4; Mr-5 | Pan; MD; LM; FM; CM | [29,89] |
| Rhizophagus irregularis | M | sorghum, poplar | RiSLM | ProtP; MST; qRT-PCR; GEAn; ProtOE; Pan; RNAseq | [90] |
| Trichoderma atroviride | M | tomato; A. thaliana | Tal-6 | Pl-C; ProtOE; PRP; ProtP; IFPAn; ProtopP; CWI-Ass; qRT-PCR | [91,92] |
| Chitin deacetylates (CDAs) | |||||
| Pochonia chlamydosporia strain 123 | NF | A. thaliana; wheat; M. javanica; tomato banana; barley | CDA1; CDA2 | GS; Pan; Pl-C; HECD; I-DNA; I-RNA; qRT-PCR; Prot-P-RAC; EAAss; MSA | [25,26,27] |
| Metarhizium anisopliae | EPF | Pyrilla perpusilla; Helicoverpa armigera | CDA | EAAss; UF; PAGE; DEA-NC; SDS-PAGE; Prot-S; DEA-IC | [93] |
| Pestalotiopsis sp. | PF | rice | PesCDA | I-DNA; I-RNA; Syn-cDNA; ISG Prot-SeqAn; Pl-C; BT; CRB; Prot-P-FPLC; SDS-PAGE; WB; EC; MALDI-TOF-MS; HILIC-ESI-MS; EAAss; | [22] |
| Chitosanases (CSNs) | |||||
| Pochonia chlamydosporia strain 123 | NF | A. thaliana; wheat; M. javanica; tomato banana; barley | csn1; csn3; csn4; csn5; csn6; csn7; csn8; csn9; csn10; csn11 | GS; Pan; I-DNA; I-RNA; qRT-PCR; PBSeq; GseqAssem; Ganno-ASAn; DSGI | [26,94] |
| Fusarium solani strain 0114 | PF | peas, lucerne; cucurbits | Csn1 | qRT-PCR; SB; NB; Pl-C; AMT; Eass; SBAss | [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berosich, C.M.; Lopez-Moya, F.; Lopez-Llorca, L.V. Modulation of the Host Defence System by Nematophagous Fungi and Chitosan. Encyclopedia 2024, 4, 379-394. https://doi.org/10.3390/encyclopedia4010026
Berosich CM, Lopez-Moya F, Lopez-Llorca LV. Modulation of the Host Defence System by Nematophagous Fungi and Chitosan. Encyclopedia. 2024; 4(1):379-394. https://doi.org/10.3390/encyclopedia4010026
Chicago/Turabian StyleBerosich, Carla Mariel, Federico Lopez-Moya, and Luis Vicente Lopez-Llorca. 2024. "Modulation of the Host Defence System by Nematophagous Fungi and Chitosan" Encyclopedia 4, no. 1: 379-394. https://doi.org/10.3390/encyclopedia4010026
APA StyleBerosich, C. M., Lopez-Moya, F., & Lopez-Llorca, L. V. (2024). Modulation of the Host Defence System by Nematophagous Fungi and Chitosan. Encyclopedia, 4(1), 379-394. https://doi.org/10.3390/encyclopedia4010026