
Generational Effects of Opioid Exposure



{kind=link}
Definition
1. Introduction
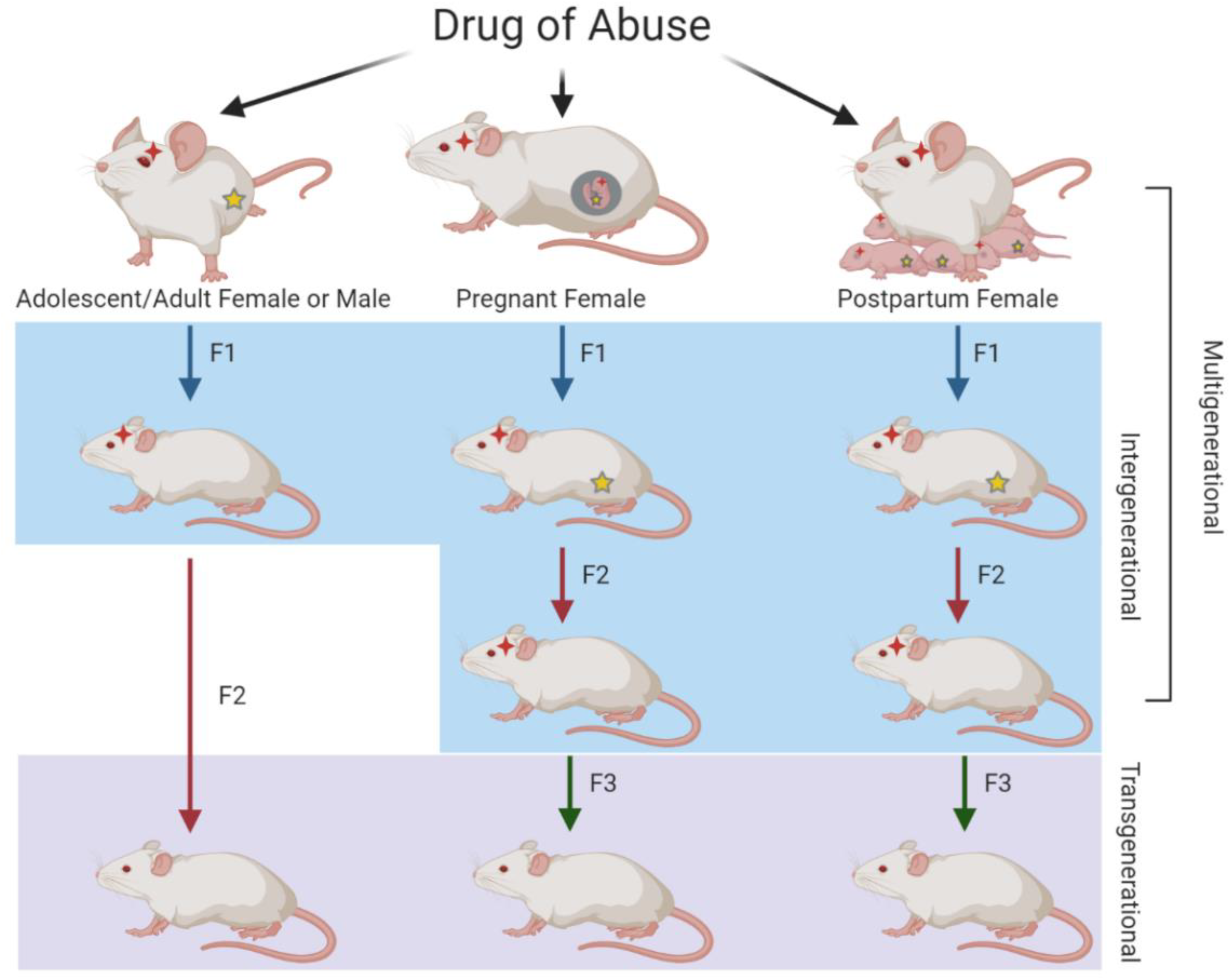
1.1. Generational Inheritance
1.2. Opioids
1.2.1. Morphine
1.2.2. Heroin
1.2.3. Oxycodone
2. Animal Studies
2.1. Morphine
2.2. Heroin
2.3. Oxycodone
3. Human Studies
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Entry Link on the Encyclopedia Platform
References
- Ostling, P.S.; Davidson, K.S.; Anyama, B.O.; Helander, E.M.; Wyche, M.Q.; Kaye, A.D. America’s Opioid Epidemic: A Comprehensive Review and Look into the Rising Crisis. Curr. Pain Headache Rep. 2018, 22, 32. [Google Scholar] [CrossRef] [PubMed]
- Badreldin, N.; Grobman, W.A.; Chang, K.T.; Yee, L.M. Opioid prescribing patterns among postpartum women. Am. J. Obstet. Gynecol. 2018, 219. [Google Scholar] [CrossRef] [PubMed]
- Bateman, B.T.; Franklin, J.M.; Bykov, K.; Avorn, J.; Shrank, W.H.; Brennan, T.A.; Landon, J.E.; Rathmell, J.P.; Huybrechts, K.F.; Fischer, M.A.; et al. Persistent opioid use following cesarean delivery: Patterns and predictors among opioid-naïve women. Am. J. Obstet. Gynecol. 2016, 215. [Google Scholar] [CrossRef] [PubMed]
- Brook, D.W.; Brook, J.S.; Zhang, C.; Cohen, P.; Whiteman, M. Drug use and the risk of major depressive disorder, alcohol dependence, and substance use disorders. Arch. Gen. Psychiatry 2002, 59, 1039–1044. [Google Scholar] [CrossRef]
- Ducci, F.; Goldman, D. The genetic basis of addictive disorders. Psychiatry Clin. N. Am. 2012, 35, 495–519. [Google Scholar] [CrossRef]
- Cloninger, C.R.; Bohman, M.; Sigvardsson, S. Inheritance of alcohol abuse. Cross-fostering analysis of adopted men. Arch. Gen. Psychiatry 1981, 38, 861–868. [Google Scholar] [CrossRef]
- Ho, M.K.; Goldman, D.; Heinz, A.; Kaprio, J.; Kreek, M.J.; Li, M.D.; Munafò, M.R.; Tyndale, R.F. Breaking barriers in the genomics and pharmacogenetics of drug addiction. Clin. Pharmacol. Ther. 2010, 88, 779–791. [Google Scholar] [CrossRef]
- Jensen, K.P. A Review of Genome-Wide Association Studies of Stimulant and Opioid Use Disorders. Mol. Neuropsychiatry 2016, 2, 37–45. [Google Scholar] [CrossRef]
- Goldberg, L.R.; Gould, T.J. Multigenerational and transgenerational effects of paternal exposure to drugs of abuse on behavioral and neural function. Eur. J. Neurosci. 2019, 50, 2453–2466. [Google Scholar] [CrossRef]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Yohn, N.L.; Bartolomei, M.S.; Blendy, J.A. Multigenerational and transgenerational inheritance of drug exposure: The effects of alcohol, opiates, cocaine, marijuana, and nicotine. Prog. Biophys. Mol. Biol. 2015, 118, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Vassoler, F.M.; Toorie, A.M.; Byrnes, E.M. Multi-, Inter-, and Transgenerational Effects of Drugs of Abuse on Behavior. Curr. Top. Behav. Neurosci. 2019, 42, 247–258. [Google Scholar] [PubMed]
- Skinner, M.K. Environmental epigenetic transgenerational inheritance and somatic epigenetic mitotic stability. Epigenetics 2011, 6, 838–842. [Google Scholar] [PubMed]
- Hanson, M.A.; Skinner, M.K. Developmental origins of epigenetic transgenerational inheritance. Environ. Epigenet. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Conradt, E.; Flannery, T.; Aschner, J.L.; Annett, R.D.; Croen, L.A.; Duarte, C.S.; Friedman, A.M.; Guille, C.; Hedderson, M.M.; Hofheimer, J.A.; et al. Prenatal Opioid Exposure: Neurodevelopmental Consequences and Future Research Priorities. Pediatrics 2019, 144. [Google Scholar] [CrossRef]
- Barker, D.J. Fetal origins of coronary heart disease. BMJ 1995, 311, 171–174. [Google Scholar] [CrossRef]
- Skogen, J.C.; Overland, S. The fetal origins of adult disease: A narrative review of the epidemiological literature. JRSM Short Rep. 2012, 3, 59. [Google Scholar] [CrossRef]
- Finnegan, L.P. Effects of maternal opiate abuse on the newborn. Fed. Proc. 1985, 44, 2314–2317. [Google Scholar]
- Towers, C.V.; Hyatt, B.W.; Visconti, K.C.; Chernicky, L.; Chattin, K.; Fortner, K.B. Neonatal Head Circumference in Newborns with Neonatal Abstinence Syndrome. Pediatrics 2019, 143. [Google Scholar] [CrossRef]
- Attarian, S.; Tran, L.C.; Moore, A.; Stanton, G.; Meyer, E.; Moore, R.P. The neurodevelopmental impact of neonatal morphine administration. Brain Sci. 2014, 4, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.C.; Hung, T.W.; Lee, C.W.; Yan, J.Y.; Ho, I.K. Enhancement of tolerance development to morphine in rats prenatally exposed to morphine, methadone, and buprenorphine. J. Biomed. Sci. 2010, 17, 46. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.; Berrebi, A.; Garipuy, D.; Schmitt, L.; Hammou, Y.; Chaumerliac, C.; Lapeyre-Mestre, M.; Montastruc, J.L.; Damase-Michel, C. Buprenorphine versus methadone in pregnant opioid-dependent women: A prospective multicenter study. Eur. J. Clin. Pharmacol. 2011, 67, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, K.E.; Schaal, V.L.; Clark, A.R.; Koul, S.; Gowen, A.; Sankarasubramani, J.; Xiao, P.; Guda, C.; Lisco, S.J.; Yelamanchili, S.V.; et al. Characterization of the intergenerational impact of in utero and postnatal oxycodone exposure. Transl. Psychiatry 2020, 10, 329. [Google Scholar] [CrossRef]
- Oei, J.L. Adult consequences of prenatal drug exposure. Intern. Med. J. 2018, 48, 25–31. [Google Scholar] [CrossRef]
- Nygaard, E.; Slinning, K.; Moe, V.; Walhovd, K.B. Cognitive function of youths born to mothers with opioid and poly-substance abuse problems during pregnancy. Child. Neuropsychol. 2017, 23, 159–187. [Google Scholar] [CrossRef]
- Vidal, S.I.; Vandeleur, C.; Rothen, S.; Gholam-Rezaee, M.; Castelao, E.; Halfon, O.; Aubry, J.M.; Ferrero, F.; Preisig, M. Risk of mental disorders in children of parents with alcohol or heroin dependence: A controlled high-risk study. Eur. Addict. Res. 2012, 18, 253–264. [Google Scholar] [CrossRef]
- Rey, R.; Wallace, L.E.; Cadden, J.A.; Cadden, S.W.; Brieger, G.H. The History of Pain; Harvard University Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Ksir, C.J.; Carl, D.; Hart, L. Drugs, Society, and Human Behavior; McGraw-Hill Education: New York, NY, USA, 2017. [Google Scholar]
- Cooper, T.E.; Chen, J.; Wiffen, P.J.; Derry, S.; Carr, D.B.; Aldington, D.; Cole, P.; Moore, R.A. Morphine for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2017, 5. [Google Scholar] [CrossRef]
- Beltrán-Campos, V.; Silva-Vera, M.; García-Campos, M.L.; Díaz-Cintra, S. Effects of morphine on brain plasticity. Neurologia 2015, 30, 176–180. [Google Scholar] [CrossRef]
- Manchikanti, L.; Helm, S.; Fellows, B.; Janata, J.W.; Pampati, V.; Grider, J.S.; Boswell, M.V. Opioid epidemic in the United States. Pain Physician 2012, 15, ES9–ES38. [Google Scholar]
- Dasgupta, N.; Beletsky, L.; Ciccarone, D. Opioid Crisis: No Easy Fix to Its Social and Economic Determinants. Am. J. Public Health 2018, 108, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Jayant, R.D. Penetration of the blood-brain barrier by peripheral neuropeptides: New approaches to enhancing transport and endogenous expression. Cell Tissue Res. 2019, 375, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Scholl, L.; Seth, P.; Kariisa, M.; Wilson, N.; Baldwin, G. Drug and Opioid-Involved Overdose Deaths-United States, 2013-2017. MMWR Morb. Mortal Wkly. Rep. 2018, 67, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.S.; Juurlink, D.N.; Perrone, J. Addressing the Opioid Epidemic. JAMA 2015, 314, 1453–1454. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, K.E.; Chand, S.; Wheeler, S.; Tiwari, S.; Flores, A.; Hernandez, J.; Savine, M.; Gowen, A.; Pendyala, G.; Yelamanchili, S.V. Role of Extracellular Vesicles in Substance Abuse and HIV-Related Neurological Pathologies. Int. J. Mol. Sci. 2020, 21, 6765. [Google Scholar] [CrossRef]
- Pacifici, G.M. Metabolism and pharmacokinetics of morphine in neonates: A review. Clinics (Sao Paulo) 2016, 71, 474–480. [Google Scholar] [CrossRef]
- Chen, M.; Zhao, Y.; Yang, H.; Luan, W.; Song, J.; Cui, D.; Dong, Y.; Lai, B.; Ma, L.; Zheng, P. Morphine disinhibits glutamatergic input to VTA dopamine neurons and promotes dopamine neuron excitation. Elife 2015, 4. [Google Scholar] [CrossRef]
- NIDA. Heroin DrugFacts; National Institute on Drug Abuse Website. Available online: https://www.drugabuse.gov/publications/drugfacts/heroin (accessed on 21 November 2019).
- Chahl, L.A. Opioids-mechanisms of action. Exp. Clin. Pharmacol. 1996, 19. [Google Scholar] [CrossRef]
- Freye, E. Opioids in Medicine: A Comprehensive Review on the Mode of Action and the Use of Analgesics in Different Clinical Pain States; Springer Science & Business Media: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Cicero, T.J.; Adams, M.L.; Giordano, A.; Miller, B.T.; O’Connor, L.; Nock, B. Influence of morphine exposure during adolescence on the sexual maturation of male rats and the development of their offspring. J. Pharmacol. Exp. Ther. 1991, 256, 1086–1093. [Google Scholar]
- Toorie, A.M.; Vassoler, F.M.; Qu, F.; Schonhoff, C.M.; Bradburn, S.; Murgatroyd, C.A.; Slonim, D.K.; Byrnes, E.M. A history of opioid exposure in females increases the risk of metabolic disorders in their future male offspring. Addict. Biol. 2019. [Google Scholar] [CrossRef]
- Vassoler, F.M.; Toorie, A.M.; Byrnes, E.M. Transgenerational blunting of morphine-induced corticosterone secretion is associated with dysregulated gene expression in male offspring. Brain Res. 2018, 1679, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, E.M. Chronic morphine exposure during puberty decreases postpartum prolactin secretion in adult female rats. Pharmacol. Biochem. Behav. 2005, 80, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.L.; Carini, L.; Schenk, M.E.; Stewart, M.; Byrnes, E.M. Adolescent opiate exposure in the female rat induces subtle alterations in maternal care and transgenerational effects on play behavior. Front. Psychiatry 2011, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, E.M. Transgenerational consequences of adolescent morphine exposure in female rats: Effects on anxiety-like behaviors and morphine sensitization in adult offspring. Psychopharmacology 2005, 182, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, J.J.; Babb, J.A.; Scanlan, V.F.; Byrnes, E.M. Adolescent opioid exposure in female rats: Transgenerational effects on morphine analgesia and anxiety-like behavior in adult offspring. Behav. Brain Res. 2011, 218, 200–205. [Google Scholar] [CrossRef]
- Vassoler, F.M.; Wright, S.J.; Byrnes, E.M. Exposure to opiates in female adolescents alters mu opiate receptor expression and increases the rewarding effects of morphine in future offspring. Neuropharmacology 2016, 103, 112–121. [Google Scholar] [CrossRef]
- Byrnes, J.J.; Johnson, N.L.; Carini, L.M.; Byrnes, E.M. Multigenerational effects of adolescent morphine exposure on dopamine D2 receptor function. Psychopharmacology 2013, 227, 263–272. [Google Scholar] [CrossRef]
- Pooriamehr, A.; Sabahi, P.; Miladi, G.H. Effects of environmental enrichment during abstinence in morphine dependent parents on anxiety, depressive-like behaviors and voluntary morphine consumption in rat offspring. Neurosci. Lett. 2017, 656, 37–42. [Google Scholar] [CrossRef]
- Pachenari, N.; Azizi, H.; Ghasemi, E.; Azadi, M.; Semnanian, S. Exposure to opiates in male adolescent rats alters pain perception in the male offspring. Behav. Pharmacol. 2018, 29, 255–260. [Google Scholar] [CrossRef]
- Pachenari, N.; Azizi, H.; Semnaniann, S. Adolescent Morphine Exposure in Male Rats Alters the Electrophysiological Properties of Locus Coeruleus Neurons of the Male Offspring. Neuroscience 2019, 410, 108–117. [Google Scholar] [CrossRef]
- Amri, J.; Sadegh, M.; Moulaei, N.; Palizvan, M.R. Transgenerational modification of hippocampus TNF-α and S100B levels in the offspring of rats chronically exposed to morphine during adolescence. Am. J. Drug Alcohol Abus. 2018, 44, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Azadi, M.; Azizi, H.; Haghparast, A. Paternal exposure to morphine during adolescence induces reward-resistant phenotype to morphine in male offspring. Brain Res. Bull. 2019, 147, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Vassoler, F.M.; Toorie, A.M.; Teceno, D.N.; Walia, P.; Moore, D.J.; Patton, T.D.; Byrnes, E.M. Paternal morphine exposure induces bidirectional effects on cocaine versus opioid self-administration. Neuropharmacology 2020, 162, 107852. [Google Scholar] [CrossRef] [PubMed]
- Nasiraei-Moghadam, S.; Sherafat, M.A.; Safari, M.S.; Moradi, F.; Ahmadiani, A.; Dargahi, L. Reversal of prenatal morphine exposure-induced memory deficit in male but not female rats. J. Mol. Neurosci. 2013, 50, 58–69. [Google Scholar] [CrossRef]
- Ahmadalipour, A.; Ghodrati-Jaldbakhan, S.; Samaei, S.A.; Rashidy-Pour, A. Deleterious effects of prenatal exposure to morphine on the spatial learning and hippocampal BDNF and long-term potentiation in juvenile rats: Beneficial influences of postnatal treadmill exercise and enriched environment. Neurobiol. Learn. Mem. 2018, 147, 54–64. [Google Scholar] [CrossRef]
- Yang, S.N.; Liu, C.A.; Chung, M.Y.; Huang, H.C.; Yeh, G.C.; Wong, C.S.; Lin, W.W.; Yang, C.H.; Tao, P.L. Alterations of postsynaptic density proteins in the hippocampus of rat offspring from the morphine-addicted mother: Beneficial effect of dextromethorphan. Hippocampus 2006, 16, 521–530. [Google Scholar] [CrossRef]
- Lin, C.S.; Tao, P.L.; Jong, Y.J.; Chen, W.F.; Yang, C.H.; Huang, L.T.; Chao, C.F.; Yang, S.N. Prenatal morphine alters the synaptic complex of postsynaptic density 95 with N-methyl-D-aspartate receptor subunit in hippocampal CA1 subregion of rat offspring leading to long-term cognitive deficits. Neuroscience 2009, 158, 1326–1337. [Google Scholar] [CrossRef]
- Farzin, D. Modification of naloxone-induced withdrawal signs by dextromethorphan in morphine-dependent mice. Eur. J. Pharmacol. 1999, 377, 35–42. [Google Scholar] [CrossRef]
- Mao, J.; Price, D.D.; Caruso, F.S.; Mayer, D.J. Oral administration of dextromethorphan prevents the development of morphine tolerance and dependence in rats. Pain 1996, 67, 361–368. [Google Scholar] [CrossRef]
- Sarkaki, A.; Assaei, R.; Motamedi, F.; Badavi, M.; Pajouhi, N. Effect of parental morphine addiction on hippocampal long-term potentiation in rats offspring. Behav. Brain Res. 2008, 186, 72–77. [Google Scholar] [CrossRef]
- Niu, L.; Cao, B.; Zhu, H.; Mei, B.; Wang, M.; Yang, Y.; Zhou, Y. Impaired in vivo synaptic plasticity in dentate gyrus and spatial memory in juvenile rats induced by prenatal morphine exposure. Hippocampus 2009, 19, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.W.; Duan, T.T.; Zhou, Q.X.; Ding, Z.Y.; Jing, L.; Cao, J.; Wang, L.P.; Mao, R.R.; Xu, L. Impaired contextual fear extinction and hippocampal synaptic plasticity in adult rats induced by prenatal morphine exposure. Addict. Biol. 2015, 20, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Sepehri, G.; Parsania, S.; Hajzadeh, M.A.; Haghpanah, T.; Sheibani, V.; Divsalar, K.; Shekarforoush, S.; Afarinesh, M.R. The effects of co-administration of opium and morphine with nicotine during pregnancy on spatial learning and memory of adult male offspring rats. Iran. J. Basic Med. Sci. 2014, 17, 694–701. [Google Scholar] [PubMed]
- Sabzevari, S.; Rohbani, K.; Sadat-Shirazi, M.S.; Babhadi-Ashar, N.; Shakeri, A.; Ashabi, G.; Khalifeh, S.; Ale-Ebrahim, M.; Zarrindast, M.R. Morphine exposure before conception affects anxiety-like behavior and CRF level (in the CSF and plasma) in the adult male offspring. Brain Res. Bull. 2019, 144, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Q.; Luo, Y.W.; Bi, F.F.; Cui, T.T.; Song, L.; Cao, W.Y.; Zhang, J.Y.; Li, F.; Xu, J.M.; Hao, W.; et al. Development of anxiety-like behavior via hippocampal IGF-2 signaling in the offspring of parental morphine exposure: Effect of enriched environment. Neuropsychopharmacology 2014, 39, 2777–2787. [Google Scholar] [CrossRef] [PubMed]
- Akbarabadi, A.; Niknamfar, S.; Vousooghi, N.; Sadat-Shirazi, M.S.; Toolee, H.; Zarrindast, M.R. Effect of rat parental morphine exposure on passive avoidance memory and morphine conditioned place preference in male offspring. Physiol. Behav. 2018, 184, 143–149. [Google Scholar] [CrossRef]
- Moulaei, N.; Mondanizadeh, M.; Salmani, M.E.; Palizvan, M.R.; Khansarinejad, B.; Sadegh, M. Transgenerational consequences of prepregnancy chronic morphine use on spatial learning and hippocampal Mecp2 and Hdac2 expression. Neuroreport 2018, 29, 739–744. [Google Scholar] [CrossRef]
- Ellis, A.S.; Toussaint, A.B.; Knouse, M.C.; Thomas, A.S.; Bongiovanni, A.R.; Mayberry, H.L.; Bhakta, S.; Peer, K.; Bangasser, D.A.; Wimmer, M.E. Paternal morphine self-administration produces object recognition memory deficits in female, but not male offspring. Psychopharmacology 2020, 237, 1209–1221. [Google Scholar] [CrossRef]
- Sadat-Shirazi, M.S.; Asgari, P.; Mahboubi, S.; Nouri Zadeh-Tehrani, S.; Ashabi, G.; Rohbani, K.; Sabzevari, S.; Soltani, H.; Khalifeh, S.; Zarrindast, M.R. Effect of morphine exposure on novel object memory of the offspring: The role of histone H3 and ΔFosB. Brain Res. Bull. 2020, 156, 141–149. [Google Scholar] [CrossRef]
- Ashabi, G.; Sadat-Shirazi, M.S.; Akbarabadi, A.; Vousooghi, N.; Kheiri, Z.; Toolee, H.; Khalifeh, S.; Zarrindast, M.R. Is the Nociception Mechanism Altered in Offspring of Morphine-Abstinent Rats? J. Pain 2018, 19, 529–541. [Google Scholar] [CrossRef]
- Vousooghi, N.; Sadat-Shirazi, M.S.; Safavi, P.; Zeraati, R.; Akbarabadi, A.; Makki, S.M.; Nazari, S.; Zarrindast, M.R. Adult rat morphine exposure changes morphine preference, anxiety, and the brain expression of dopamine receptors in male offspring. Int. J. Dev. Neurosci. 2018, 69, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Ashabi, G.; Matloob, M.; Monfared Neirizi, N.; Behrouzi, M.; Safarzadeh, M.; Rajabpoor Dehdashti, A.; Sadat-Shirazi, M.S.; Zarrindast, M.R. Activation of D1-like dopamine receptors is involved in the impairment of spatial memory in the offspring of morphine-abstinent rats. Neurosci. Res. 2020, 158, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Farah Naquiah, M.Z.; James, R.J.; Suratman, S.; Lee, L.S.; Mohd Hafidz, M.I.; Salleh, M.Z.; Teh, L.K. Transgenerational effects of paternal heroin addiction on anxiety and aggression behavior in male offspring. Behav. Brain Funct. 2016, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.H.; Stadlin, A. Prenatal heroin exposure. Effects on development, acoustic startle response, and locomotion in weanling rats. Neurotoxicol. Teratol. 2000, 22, 193–203. [Google Scholar] [CrossRef]
- Wang, Y.; Han, T.Z. Prenatal exposure to heroin in mice elicits memory deficits that can be attributed to neuronal apoptosis. Neuroscience 2009, 160, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Liu, X.; Long, H.; Ma, L. Effects of prenatal cocaine and heroin exposure on neuronal dendrite morphogenesis and spatial recognition memory in mice. Neurosci. Lett. 2012, 522, 128–133. [Google Scholar] [CrossRef]
- Yanai, J.; Steingart, R.A.; Snapir, N.; Gvaryahu, G.; Rozenboim, I.; Katz, A. The relationship between neural alterations and behavioral deficits after prenatal exposure to heroin. Ann. N. Y. Acad. Sci. 2000, 914, 402–411. [Google Scholar] [CrossRef]
- Shahak, H.; Slotkin, T.A.; Yanai, J. Alterations in PKCgamma in the mouse hippocampus after prenatal exposure to heroin: A link from cell signaling to behavioral outcome. Brain Res. Dev. Brain Res. 2003, 140, 117–125. [Google Scholar] [CrossRef]
- Yaniv, S.P.; Naor, Z.; Yanai, J. Prenatal heroin exposure alters cholinergic receptor stimulated activation of the PKCbetaII and PKCgamma isoforms. Brain Res. Bull. 2004, 63, 339–349. [Google Scholar] [CrossRef]
- Huleihel, R.; Yanai, J. Disruption of the development of cholinergic-induced translocation/activation of PKC isoforms after prenatal heroin exposure. Brain Res. Bull. 2006, 69, 174–181. [Google Scholar] [CrossRef]
- Devarapalli, M.; Leonard, M.; Briyal, S.; Stefanov, G.; Puppala, B.L.; Schweig, L.; Gulati, A. Prenatal Oxycodone Exposure Alters CNS Endothelin Receptor Expression in Neonatal Rats. Drug Res. 2016, 66, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Sithisarn, T.; Bada, H.S.; Charnigo, R.J.; Legan, S.J.; Randall, D.C. Effects of perinatal oxycodone exposure on the cardiovascular response to acute stress in male rats at weaning and in young adulthood. Front. Physiol. 2013, 4, 85. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.P.; Franklin, L.M.; Johnson, G.S.; Schrott, L.M. Prenatal oxycodone exposure impairs spatial learning and/or memory in rats. Behav. Brain Res. 2010, 212, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Sithisarn, T.; Legan, S.J.; Westgate, P.M.; Wilson, M.; Wellmann, K.; Bada, H.S.; Barron, S. The Effects of Perinatal Oxycodone Exposure on Behavioral Outcome in a Rodent Model. Front. Pediatr. 2017, 5, 180. [Google Scholar] [CrossRef]
- Vassoler, F.M.; Oranges, M.L.; Toorie, A.M.; Byrnes, E.M. Oxycodone self-administration during pregnancy disrupts the maternal-infant dyad and decreases midbrain OPRM1 expression during early postnatal development in rats. Pharmacol. Biochem. Behav. 2018, 173, 74–83. [Google Scholar] [CrossRef]
- Shahjin, F.; Guda, R.S.; Schaal, V.L.; Odegaard, K.; Clark, A.; Gowen, A.; Xiao, P.; Lisco, S.J.; Pendyala, G.; Yelamanchili, S.V. Brain-Derived Extracellular Vesicle microRNA Signatures Associated with In Utero and Postnatal Oxycodone Exposure. Cells 2019, 9, 21. [Google Scholar] [CrossRef]
- Mahler, S.V.; Smith, R.J.; Moorman, D.E.; Sartor, G.C.; Aston-Jones, G. Multiple roles for orexin/hypocretin in addiction. Prog. Brain Res. 2012, 198, 79–121. [Google Scholar]
- Fodor, A.; Tímár, J.; Zelena, D. Behavioral effects of perinatal opioid exposure. Life Sci. 2014, 104, 1–8. [Google Scholar] [CrossRef]
- Azuine, R.E.; Ji, Y.; Chang, H.Y.; Kim, Y.; Ji, H.; DiBari, J.; Hong, X.; Wang, G.; Singh, G.K.; Pearson, C.; et al. Prenatal Risk Factors and Perinatal and Postnatal Outcomes Associated with Maternal Opioid Exposure in an Urban, Low-Income, Multiethnic US Population. JAMA Netw. Open 2019, 2. [Google Scholar] [CrossRef]
- Lee, S.J.; Bora, S.; Austin, N.C.; Westerman, A.; Henderson, J.M.T. Neurodevelopmental Outcomes of Children Born to Opioid-Dependent Mothers: A Systematic Review and Meta-Analysis. Acad. Pediatr. 2020, 20, 308–318. [Google Scholar] [CrossRef]
- Geng, F.; Salmeron, B.J.; Ross, T.J.; Black, M.M.; Riggins, T. Long-term effects of prenatal drug exposure on the neural correlates of memory at encoding and retrieval. Neurotoxicol. Teratol. 2018, 65, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Walhovd, K.B.; Moe, V.; Slinning, K.; Due-Tønnessen, P.; Bjørnerud, A.; Dale, A.M.; van der Kouwe, A.; Quinn, B.T.; Kosofsky, B.; Greve, D.; et al. Volumetric cerebral characteristics of children exposed to opiates and other substances in utero. Neuroimage 2007, 36, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Griesler, P.C.; Hu, M.C.; Wall, M.M.; Kandel, D.B. Nonmedical Prescription Opioid Use by Parents and Adolescents in the US. Pediatrics 2019, 143. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.C.R.; Tiberio, S.S.; Capaldi, D.M.; Owen, L.D. Paternal and maternal prescription opioid use and misuse: General and specific risks for early adolescents’ substance use. Addict. Behav. 2020, 103. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.E.; Suess, P.E.; Newlin, D.B.; Spurgeon, L.; Porges, S.W. Vagal tone regulation during sustained attention in boys exposed to opiates in utero. Addict. Behav. 1995, 20, 43–59. [Google Scholar] [CrossRef]
- Wilson, G.S.; McCreary, R.; Kean, J.; Baxter, J.C. The development of preschool children of heroin-addicted mothers: A controlled study. Pediatrics 1979, 63, 135–141. [Google Scholar] [CrossRef]
- Minnes, S.; Lang, A.; Singer, L. Prenatal tobacco, marijuana, stimulant, and opiate exposure: Outcomes and practice implications. Addict. Sci. Clin. Pract. 2011, 6, 57–70. [Google Scholar]
- Thaithumyanon, P.; Limpongsanurak, S.; Praisuwanna, P.; Punnahitanon, S. Perinatal effects of amphetamine and heroin use during pregnancy on the mother and infant. J. Med. Assoc. Thai. 2005, 88, 1506–1513. [Google Scholar]
- Beckwith, A.M.; Burke, S.A. Identification of early developmental deficits in infants with prenatal heroin, methadone, and other opioid exposure. Clin. Pediatr. 2015, 54, 328–335. [Google Scholar] [CrossRef]
- Ornoy, A.; Michailevskaya, V.; Lukashov, I.; Bar-Hamburger, R.; Harel, S. The developmental outcome of children born to heroin-dependent mothers, raised at home or adopted. Child. Abuse Neglect. 1996, 20, 385–396. [Google Scholar] [CrossRef]
- Nygaard, E.; Slinning, K.; Moe, V.; Fjell, A.; Walhovd, K.B. Mental health in youth prenatally exposed to opioids and poly-drugs and raised in permanent foster/adoptive homes: A prospective longitudinal study. Early Hum. Dev. 2019, 140. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Odegaard, K.E.; Pendyala, G.; Yelamanchili, S.V. Generational Effects of Opioid Exposure. Encyclopedia 2021, 1, 99-114. https://doi.org/10.3390/encyclopedia1010012
Odegaard KE, Pendyala G, Yelamanchili SV. Generational Effects of Opioid Exposure. Encyclopedia. 2021; 1(1):99-114. https://doi.org/10.3390/encyclopedia1010012
Chicago/Turabian StyleOdegaard, Katherine E., Gurudutt Pendyala, and Sowmya V. Yelamanchili. 2021. "Generational Effects of Opioid Exposure" Encyclopedia 1, no. 1: 99-114. https://doi.org/10.3390/encyclopedia1010012
APA StyleOdegaard, K. E., Pendyala, G., & Yelamanchili, S. V. (2021). Generational Effects of Opioid Exposure. Encyclopedia, 1(1), 99-114. https://doi.org/10.3390/encyclopedia1010012