

Endemicity Is Not a Victory: The Unmitigated Downside Risks of Widespread SARS-CoV-2 Transmission

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fitness Disadvantage Incurred by Fatal Patient Outcomes
2.2. SEIRS Modeling to Predict Endemic Infection and Death Rates
2.3. Code Availability
3. Results
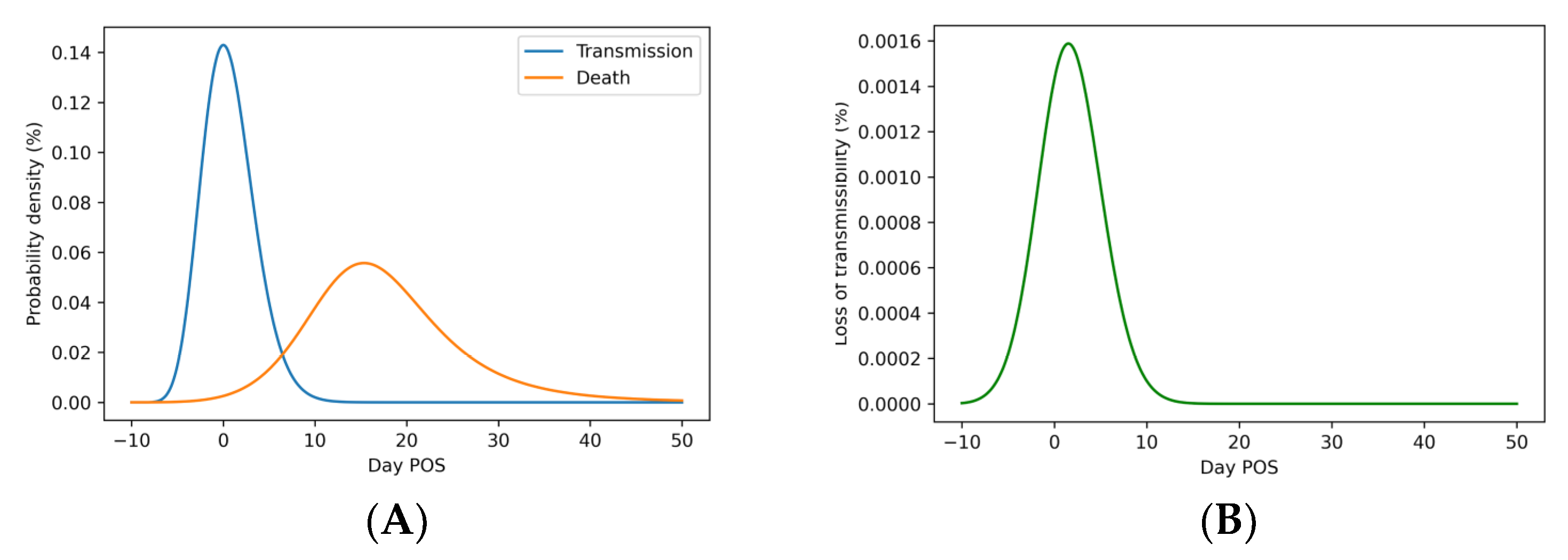
3.1. Loss of Viral Fitness Incurred by Patient Death Is Minimal
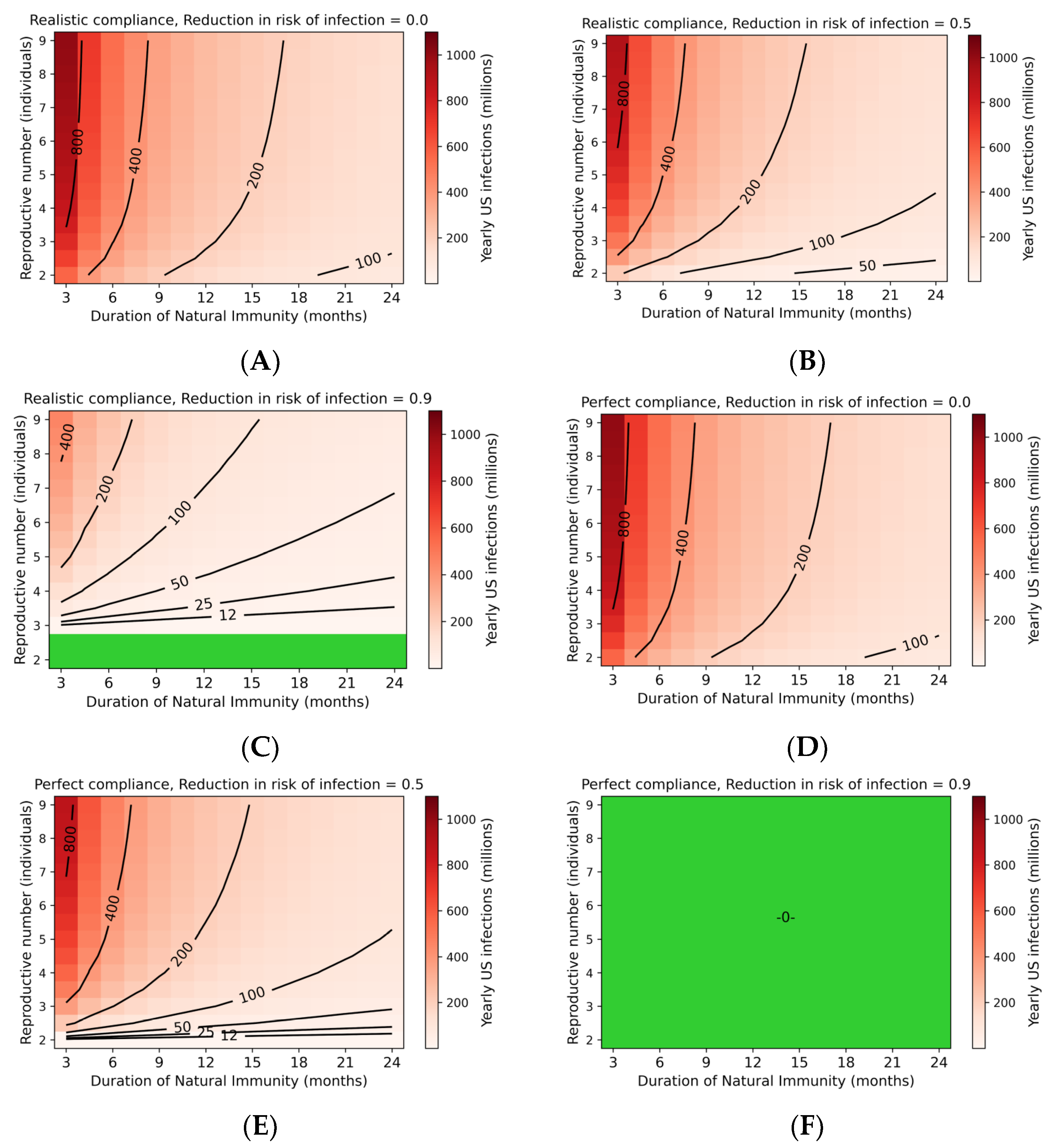
3.2. Steady-State Transmission of SARS-CoV-2 Is Extensive
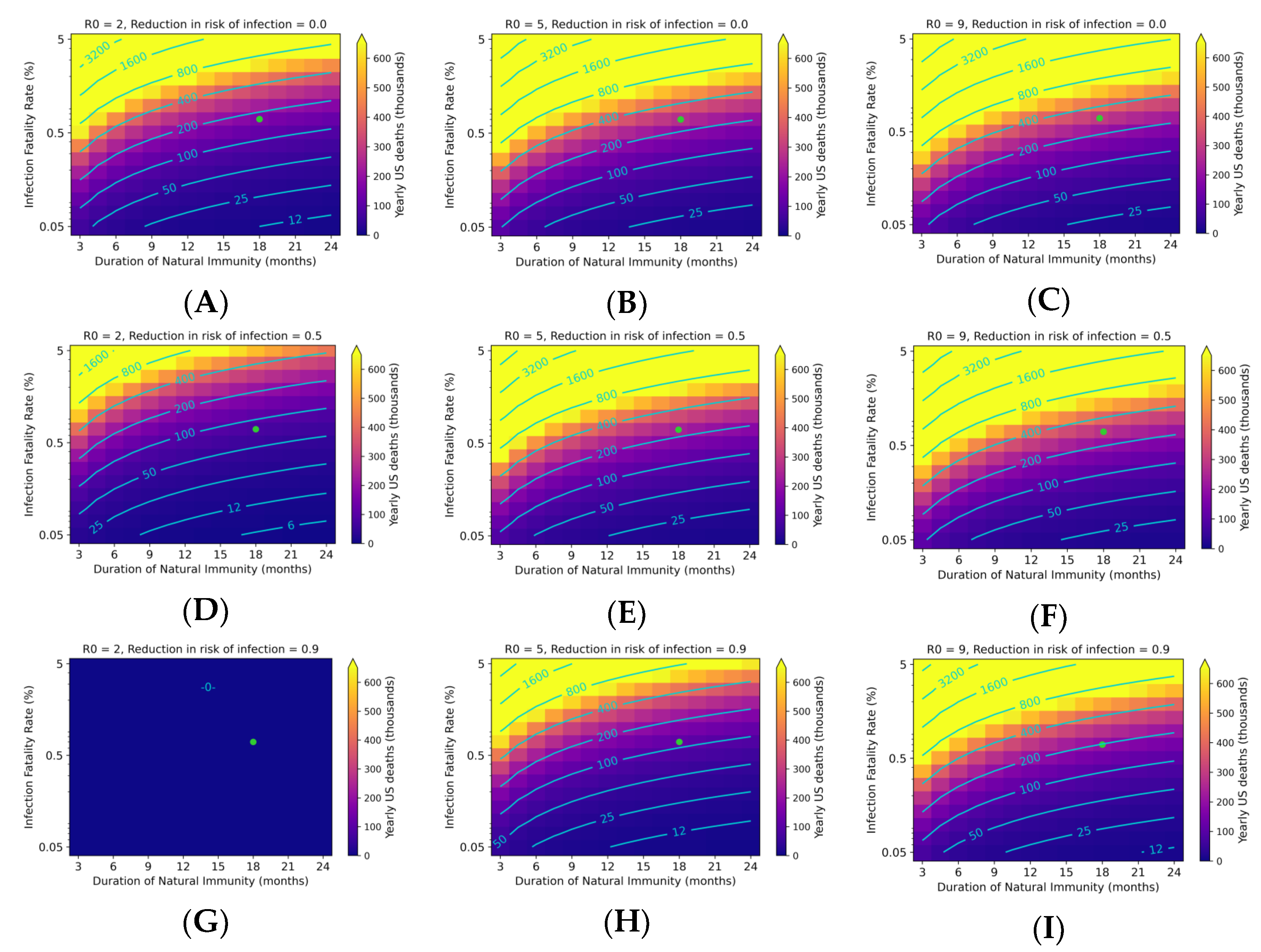
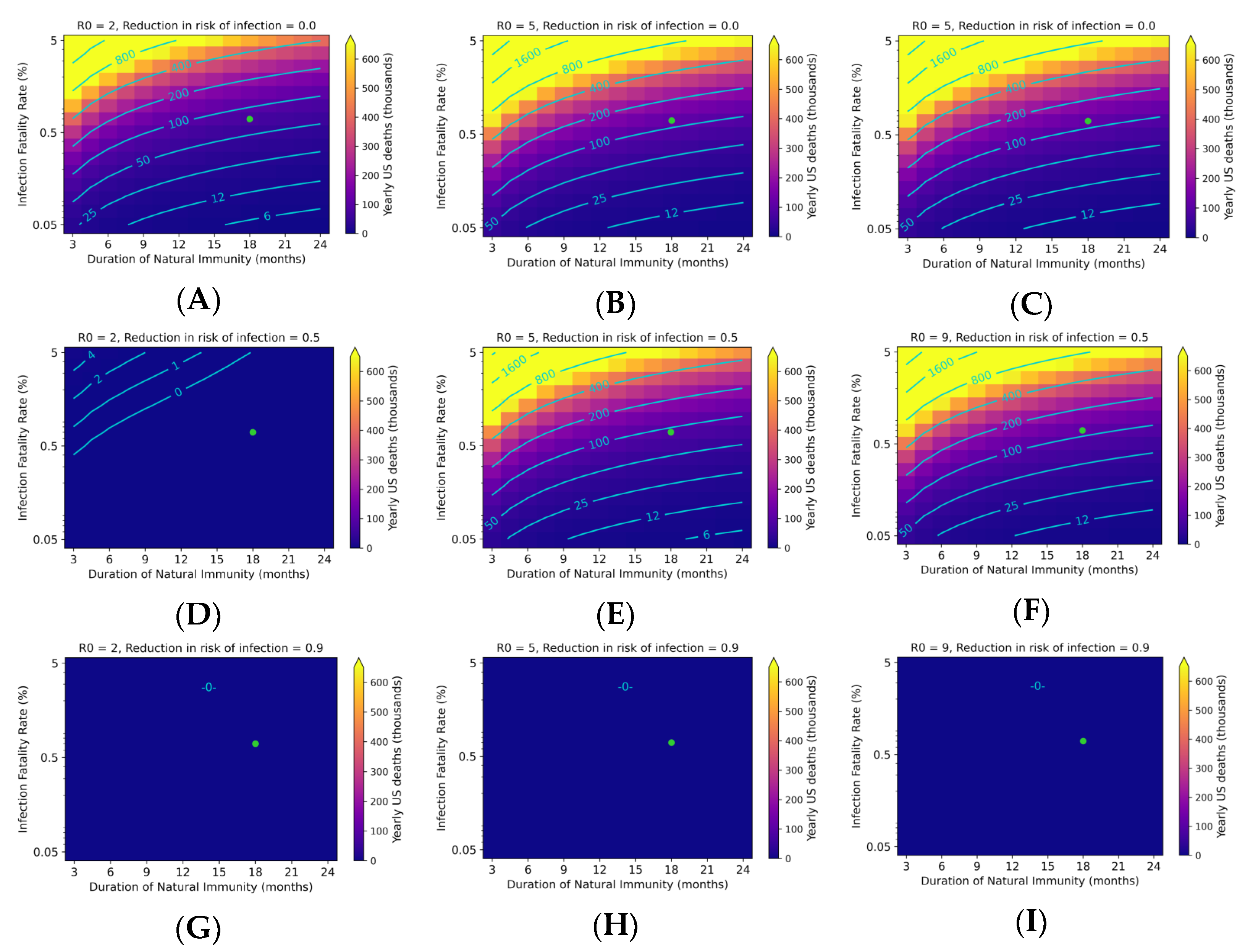
3.3. Endemic SARS-CoV-2 Death Tolls Are Highly Sensitive to Changes in IFR and Duration of Immunity
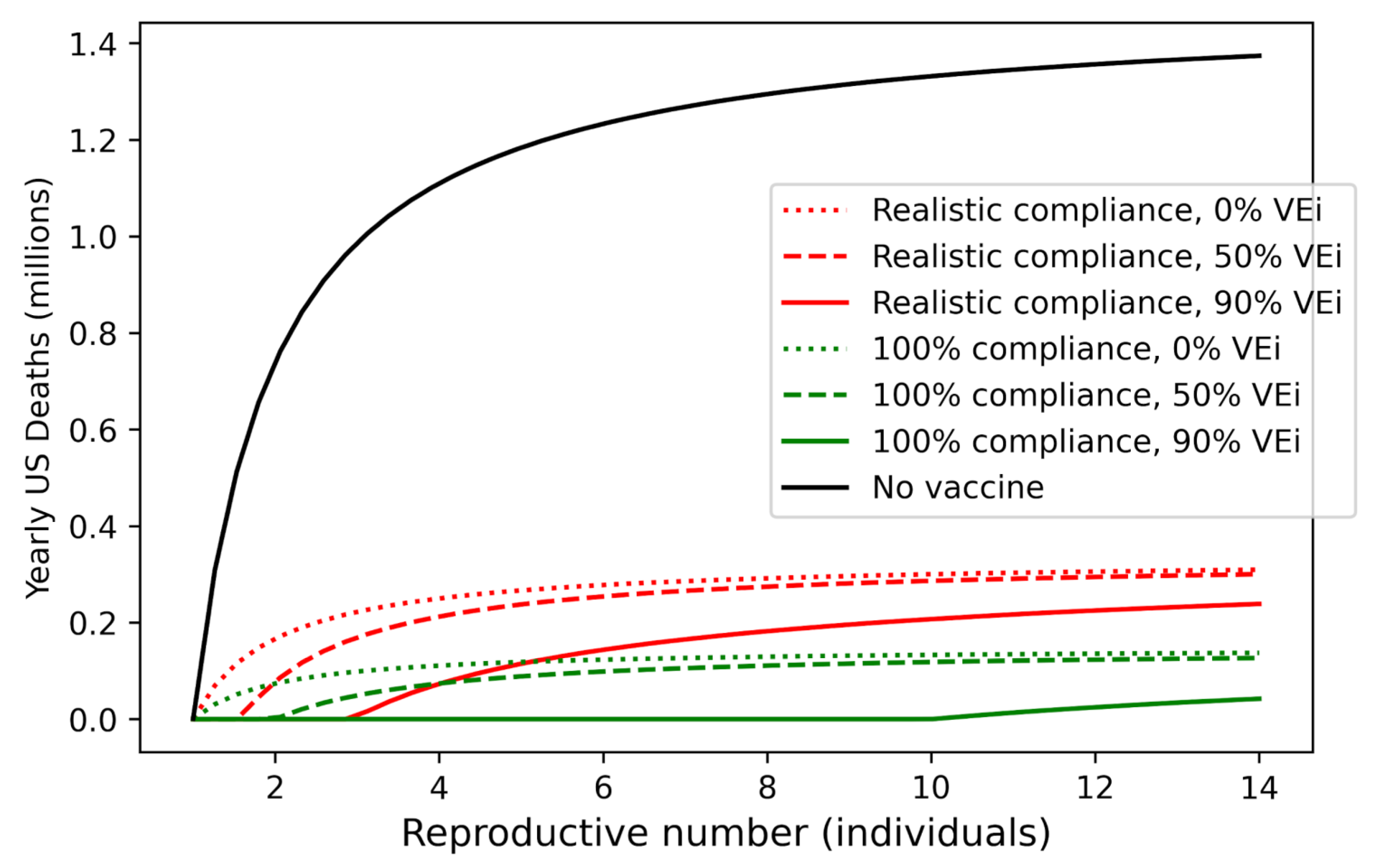
3.4. Relationship between R0 and Yearly Death Toll Is Saturating
3.5. Emergence of New Variants May Rapidly Drive Infection Levels Exceeding the Steady State
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Writer, A.P.H.S. Anthony Fauci Offers a Timeline for Ending COVID-19 Pandemic. Harvard Gazette, 10 December 2020. [Google Scholar]
- Halperin, D.T.; Hearst, N.; Hodgins, S.; Bailey, R.C.; Klausner, J.D.; Jackson, H.; Wamai, R.G.; Ladapo, J.A.; Over, M.; Baral, S.; et al. Revisiting COVID-19 Policies: 10 Evidence-Based Recommendations for Where to Go from Here. BMC Public Health 2021, 21, 2084. [Google Scholar] [CrossRef]
- Subbarao, K. The Success of SARS-CoV-2 Vaccines and Challenges Ahead. Cell Host Microbe 2021, 29, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Stieg, C. From Vaccines to Safe Socialization: Here’s What to Expect in 2021, According to Experts. CNBC, 2 January 2021. [Google Scholar]
- Picard, A. Opinion: How Will the Pandemic End? Not with a Moment of Triumph We’ll All Remember, but with a Slow Whimper We’ll Soon Forget. The Globe and Mail, 2 January 2020. [Google Scholar]
- Makary, M. Opinion|We’ll Have Herd Immunity by April. Wall Street Journal, 18 February 2021. [Google Scholar]
- Aridi, S. What’s the First Thing You Will Do When the Pandemic Ends? The New York Times, 2 January 2021. [Google Scholar]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing Antibody Levels Are Highly Predictive of Immune Protection from Symptomatic SARS-CoV-2 Infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Lumley, S.F.; Wei, J.; O’Donnell, D.; Stoesser, N.E.; Matthews, P.C.; Howarth, A.; Hatch, S.B.; Marsden, B.D.; Cox, S.; James, T.; et al. Duration, Dynamics, and Determinants of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Antibody Responses in Individual Healthcare Workers. Clin. Infect. Dis. 2021, 73, e699–e709. [Google Scholar] [CrossRef]
- Van Egeren, D.; Novokhodko, A.; Stoddard, M.; Tran, U.; Zetter, B.; Rogers, M.; Pentelute, B.L.; Carlson, J.M.; Hixon, M.; Joseph-McCarthy, D.; et al. Risk of Rapid Evolutionary Escape from Biomedical Interventions Targeting SARS-CoV-2 Spike Protein. PLoS ONE 2021, 16, e0250780. [Google Scholar] [CrossRef] [PubMed]
- Hirabara, S.M.; Serdan, T.D.A.; Gorjao, R.; Masi, L.N.; Pithon-Curi, T.C.; Covas, D.T.; Curi, R.; Durigon, E.L. SARS-COV-2 Variants: Differences and Potential of Immune Evasion. Front. Cell Infect. Microbiol. 2021, 11, 781429. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community Transmission and Viral Load Kinetics of the SARS-CoV-2 Delta (B.1.617.2) Variant in Vaccinated and Unvaccinated Individuals in the UK: A Prospective, Longitudinal, Cohort Study. Lancet Infect. Dis. 2022, 22, 183–195. [Google Scholar] [CrossRef]
- Eyre, D.W.; Taylor, D.; Purver, M.; Chapman, D.; Fowler, T.; Pouwels, K.; Walker, A.S.; Peto, T.E. The Impact of SARS-Cov-2 Vaccination on Alpha and Delta Variant Transmission. medRxiv 2021. [Google Scholar] [CrossRef]
- Levin, A.T.; Hanage, W.P.; Owusu-Boaitey, N.; Cochran, K.B.; Walsh, S.P.; Meyerowitz-Katz, G. Assessing the Age Specificity of Infection Fatality Rates for COVID-19: Systematic Review, Meta-Analysis, and Public Policy Implications. Eur. J. Epidemiol. 2020, 35, 1123–1138. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Response Reporting Mass.Gov. Available online: https://www.mass.gov/info-details/covid-19-response-reporting (accessed on 6 March 2022).
- Brandal, L.T.; MacDonald, E.; Veneti, L.; Ravlo, T.; Lange, H.; Naseer, U.; Feruglio, S.; Bragstad, K.; Hungnes, O.; Ødeskaug, L.E.; et al. Outbreak Caused by the SARS-CoV-2 Omicron Variant in Norway, November to December 2021. Eurosurveillance 2021, 26, 2101147. [Google Scholar] [CrossRef]
- Brown, C.M. Outbreak of SARS-CoV-2 Infections, Including COVID-19 Vaccine Breakthrough Infections, Associated with Large Public Gatherings—Barnstable County, Massachusetts, July 2021. MMWR Morb. Mortal Wkly. Rep. 2021, 70, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Buttler, M. Denmark Declares Covid No Longer Poses Threat to Society. Bloomberg, 26 January 2022. [Google Scholar]
- WHO, Europe Entering “Plausible Endgame” to COVID Pandemic. ABC News, 3 February 2022.
- Press Briefing by White House COVID-19 Response Team and Public Health Officials. 2022. Available online: https://www.whitehouse.gov/briefing-room/press-briefings/2022/01/12/press-briefing-by-white-house-covid-19-response-team-and-public-health-officials-78/ (accessed on 6 March 2022).
- Klaassen, F.; Chitwood, M.H.; Cohen, T.; Pitzer, V.E.; Russi, M.; Swartwood, N.A.; Salomon, J.A.; Menzies, N.A. Population Immunity to Pre-Omicron and Omicron SARS-CoV-2 Variants in US States and Counties through December 1, 2021. medRxiv 2022. preprint. [Google Scholar] [CrossRef]
- Lavine, J.S.; Bjornstad, O.N.; Antia, R. Immunological Characteristics Govern the Transition of COVID-19 to Endemicity. Science 2021, 371, 741–745. [Google Scholar] [CrossRef]
- Rascoe, A. Viruses Evolve and Weaken over Time—What Does That Mean for the Coronavirus? NPR, 9 January 2022. [Google Scholar]
- Ewald, P.W. Evolution of Virulence. Infect. Dis. Clin. N. Am. 2004, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Coyne, K.P.; Jones, B.R.D.; Kipar, A.; Chantrey, J.; Porter, C.J.; Barber, P.J.; Dawson, S.; Gaskell, R.M.; Radford, A.D. Lethal Outbreak of Disease Associated with Feline Calicivirus Infection in Cats. Vet. Rec. 2006, 158, 544–550. [Google Scholar] [CrossRef]
- Marshall, I.D.; Fenner, F. Studies in the Epidemiology of Infectious Myxomatosis of Rabbits. J. Hyg. 1958, 56, 288–302. [Google Scholar] [CrossRef]
- Kerr, P.J.; Cattadori, I.M.; Liu, J.; Sim, D.G.; Dodds, J.W.; Brooks, J.W.; Kennett, M.J.; Holmes, E.C.; Read, A.F. Next Step in the Ongoing Arms Race between Myxoma Virus and Wild Rabbits in Australia Is a Novel Disease Phenotype. Proc. Natl. Acad. Sci. USA 2017, 114, 9397–9402. [Google Scholar] [CrossRef]
- Best, S.M.; Kerr, P.J. Coevolution of Host and Virus: The Pathogenesis of Virulent and Attenuated Strains of Myxoma Virus in Resistant and Susceptible European Rabbits. Virology 2000, 267, 36–48. [Google Scholar] [CrossRef]
- DeJesus, E.; Costa-Hurtado, M.; Smith, D.; Lee, D.-H.; Spackman, E.; Kapczynski, D.R.; Torchetti, M.K.; Killian, M.L.; Suarez, D.L.; Swayne, D.E.; et al. Changes in Adaptation of H5N2 Highly Pathogenic Avian Influenza H5 Clade 2.3.4.4 Viruses in Chickens and Mallards. Virology 2016, 499, 52–64. [Google Scholar] [CrossRef]
- Spackman, E.; Pantin-Jackwood, M.J.; Kapczynski, D.R.; Swayne, D.E.; Suarez, D.L. H5N2 Highly Pathogenic Avian Influenza Viruses from the US 2014-2015 Outbreak Have an Unusually Long Pre-Clinical Period in Turkeys. BMC Vet. Res. 2016, 12, 260. [Google Scholar] [CrossRef]
- Herbeck, J.T.; Müller, V.; Maust, B.S.; Ledergerber, B.; Torti, C.; Di Giambenedetto, S.; Gras, L.; Günthard, H.F.; Jacobson, L.P.; Mullins, J.I.; et al. Is the Virulence of HIV Changing? A Meta-Analysis of Trends in Prognostic Markers of HIV Disease Progression and Transmission. AIDS 2012, 26, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.; Hollingsworth, T.D.; Chapman, R.; de Wolf, F.; Hanage, W.P. Variation in HIV-1 Set-Point Viral Load: Epidemiological Analysis and an Evolutionary Hypothesis. Proc. Natl. Acad. Sci. USA 2007, 104, 17441–17446. [Google Scholar] [CrossRef] [PubMed]
- Wymant, C.; Bezemer, D.; Blanquart, F.; Ferretti, L.; Gall, A.; Hall, M.; Golubchik, T.; Bakker, M.; Ong, S.H.; Zhao, L.; et al. A Highly Virulent Variant Of HIV-1 Circulating in the Netherlands. Science 2022, 375, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Department of Commerce Bureau of the Census, Mortality Statistics 1918. Available online: https://www.cdc.gov/nchs/data/vsushistorical/mortstatsh_1918.pdf (accessed on 6 March 2022).
- Tumpey, T.M.; Basler, C.F.; Aguilar, P.V.; Zeng, H.; Solórzano, A.; Swayne, D.E.; Cox, N.J.; Katz, J.M.; Taubenberger, J.K.; Palese, P.; et al. Characterization of the Reconstructed 1918 Spanish Influenza Pandemic Virus. Science 2005, 310, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kawaoka, Y. Pathogenesis of the 1918 Pandemic Influenza Virus. PLoS Pathog. 2011, 7, e1001218. [Google Scholar] [CrossRef]
- Sofonea, M.T.; Aldakak, L.; Boullosa, L.F.V.V.; Alizon, S. Can Ebola Virus Evolve to Be Less Virulent in Humans? J. Evol. Biol. 2018, 31, 382–392. [Google Scholar] [CrossRef]
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the Evolution of Host Range and Virulence. Infect. Genet. Evol. 2014, 21, 15–40. [Google Scholar] [CrossRef]
- Dumbell, K.R.; Wells, D.G. The Pathogenicity of Variola Virus. A Comparison of the Growth of Standard Strains of Variola Major and Variola Minor Viruses in Cell Cultures from Human Embryos. J. Hyg. 1982, 89, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Huq, F.; Dumbell, K.R. Virulence of Variola Viruses for Suckling Mice. Trans. R. Soc. Trop. Med. Hyg. 2003, 97, 97–99. [Google Scholar] [CrossRef]
- Fisman, D.N.; Tuite, A.R. Evaluation of the Relative Virulence of Novel SARS-CoV-2 Variants: A Retrospective Cohort Study in Ontario, Canada. CMAJ 2021, 193, E1619–E1625. [Google Scholar] [CrossRef]
- Davies, N.G.; Jarvis, C.I.; Edmunds, W.J.; Jewell, N.P.; Diaz-Ordaz, K.; Keogh, R.H. Increased Mortality in Community-Tested Cases of SARS-CoV-2 Lineage, B.1.1.7. Nature 2021, 593, 270–274. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.D.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and Epidemiology of the P.1 SARS-CoV-2 Lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Meyerowitz-Katz, G.; Merone, L. A Systematic Review and Meta-Analysis of Published Research Data on COVID-19 Infection Fatality Rates. Int. J. Infect. Dis. 2020, 101, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated Transmissibility and Impact of SARS-Cov-2 Lineage B. 1.1. 7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Abu-Raddad, L.J.; Chemaitelly, H.; Ayoub, H.H.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; Tang, P.; Hasan, M.R.; Coyle, P.; AlMukdad, S.; et al. Severity, Criticality, and Fatality of the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Beta Variant. Clin. Infect. Dis. 2021, 75, e1188–e1191. [Google Scholar] [CrossRef] [PubMed]
- Pearson, C.A.B.; Russell, T.J.; Davies, N.; Kucharski, A.J.; CMMID COVID-19 working group; Edmunds, W.J.; Eggo, R.M. Estimates of Severity and Transmissibility of Novel SARS-CoV-2 Variant 501Y.V2 in South Africa. CMMID Repository, 1 November 2021. Available online: https://cmmid.github.io/topics/covid19/reports/sa-novel-variant/2021_01_11_Transmissibility_and_severity_of_501Y_V2_in_SA.pdf (accessed on 6 March 2022).
- Liu, Y.; Rocklöv, J. The Reproductive Number of the Delta Variant of SARS-CoV-2 Is Far Higher Compared to the Ancestral SARS-CoV-2 Virus. J. Travel Med. 2021, 28, taab124. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yu, Y.; Zhao, Y.; He, D. Reduction in the Infection Fatality Rate of Omicron (B.1.1.529) Variant Compared to Previous Variants in South Africa; Social Science Research Network: Rochester, NY, USA, 2022. [Google Scholar]
- Lyngse, F.P.; Mortensen, L.H.; Denwood, M.J.; Christiansen, L.E.; Møller, C.H.; Skov, R.L.; Spiess, K.; Fomsgaard, A.; Lassaunière, M.M.; Rasmussen, M.; et al. SARS-CoV-2 Omicron VOC Transmission in Danish Households. medRxiv 2021. [Google Scholar] [CrossRef]
- O’Driscoll, M.; Ribeiro Dos Santos, G.; Wang, L.; Cummings, D.A.T.; Azman, A.S.; Paireau, J.; Fontanet, A.; Cauchemez, S.; Salje, H. Age-Specific Mortality and Immunity Patterns of SARS-CoV-2. Nature 2021, 590, 140–145. [Google Scholar] [CrossRef]
- Wu, X.; Nethery, R.C.; Sabath, M.B.; Braun, D.; Dominici, F. Air Pollution and COVID-19 Mortality in the United States: Strengths And Limitations of An Ecological Regression Analysis. Sci. Adv. 2020, 6, eabd4049. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.; Kwatra, G.; Myers, J.E.; Jassat, W.; Dhar, N.; Mukendi, C.K.; Nana, A.; Blumberg, L.; Welch, R.; Ngorima-Mabhena, N.; et al. South African population immunity and severe Covid-19 with omicron variant. medRxiv 2021. [Google Scholar] [CrossRef]
- CHEST 2021: Mechanical Ventilation Associated with Higher Mortality in COVID-19 Patients. Available online: https://www.practiceupdate.com/content/chest-2021-mechanical-ventilation-associated-with-higher-mortality-in-covid-19-patients/126115 (accessed on 6 March 2022).
- Yeates, E.O.; Nahmias, J.; Chinn, J.; Sullivan, B.; Stopenski, S.; Amin, A.N.; Nguyen, N.T. Improved Outcomes over Time for Adult COVID-19 Patients with Acute Respiratory Distress Syndrome or Acute Respiratory Failure. PLoS ONE 2021, 16, e0253767. [Google Scholar] [CrossRef] [PubMed]
- Janke, A.T.; Mei, H.; Rothenberg, C.; Becher, R.D.; Lin, Z.; Venkatesh, A.K. Analysis of Hospital Resource Availability and COVID-19 Mortality Across the United States. J. Hosp. Med. 2021, 16, 211–214. [Google Scholar] [CrossRef]
- Olivas-Martínez, A.; Cárdenas-Fragoso, J.L.; Jiménez, J.V.; Lozano-Cruz, O.A.; Ortiz-Brizuela, E.; Tovar-Méndez, V.H.; Medrano-Borromeo, C.; Martínez-Valenzuela, A.; Román-Montes, C.M.; Martínez-Guerra, B.; et al. In-hospital mortality from severe COVID-19 in a tertiary care center in Mexico City; causes of death, risk factors and the impact of hospital saturation. PLoS ONE 2021, 16, e0245772. [Google Scholar] [CrossRef] [PubMed]
- SARS-CoV-2 Variants of Concern and Variants under Investigation in England. UK Health Security Agency, 31 December 2021. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1045619/Technical-Briefing-31-Dec-2021-Omicron_severity_update.pdf (accessed on 6 March 2022).
- REGEN-COV. Antibody Combination and Outcomes in Outpatients with Covid-19|NEJM. Available online: https://www.nejm.org/doi/full/10.1056/NEJMoa2108163 (accessed on 6 March 2022).
- FDA Halts Use of Regeneron and Eli Lilly Antibody Drugs That Don’t Work against Covid Omicron. Available online: https://www.cnbc.com/2022/01/24/fda-halts-use-of-antibody-drugs-that-dont-work-against-covid-omicron-.html (accessed on 6 March 2022).
- Bariola, J.R.; McCreary, E.K.; Wadas, R.J.; Kip, K.E.; Marroquin, O.C.; Minnier, T.; Koscumb, S.; Collins, K.; Schmidhofer, M.; Shovel, J.A.; et al. Impact of Bamlanivimab Monoclonal Antibody Treatment on Hospitalization and Mortality Among Nonhospitalized Adults with Severe Acute Respiratory Syndrome Coronavirus 2 Infection. Open Forum Infect. Dis. 2021, 8, ofab254. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lau, E.H.Y.; Wu, P.; Deng, X.; Wang, J.; Hao, X.; Lau, Y.C.; Wong, J.Y.; Guan, Y.; Tan, X.; et al. Temporal Dynamics in Viral Shedding and Transmissibility of COVID-19. Nat. Med. 2020, 26, 672–675. [Google Scholar] [CrossRef]
- Bai, J.; Shi, F.; Cao, J.; Wen, H.; Wang, F.; Mubarik, S.; Liu, X.; Yu, Y.; Ding, J.; Yu, C. The Epidemiological Characteristics of Deaths with COVID-19 in the Early Stage of Epidemic in Wuhan, China. Glob. Health Res. Policy 2020, 5, 54. [Google Scholar] [CrossRef]
- Abu-Raddad, L.J.; Chemaitelly, H.; Ayoub, H.H.; Tang, P.; Coyle, P.; Hasan, M.R.; Yassine, H.M.; Benslimane, F.M.; Khatib, H.A.A.; Kanaani, Z.A.; et al. Effect of Vaccination and of Prior Infection on Infectiousness of Vaccine Breakthrough Infections and Reinfections. medRxiv 2021. [Google Scholar] [CrossRef]
- Levine-Tiefenbrun, M.; Yelin, I.; Alapi, H.; Katz, R.; Herzel, E.; Kuint, J.; Chodick, G.; Gazit, S.; Patalon, T.; Kishony, R. Viral Loads of Delta-Variant SARS-CoV-2 Breakthrough Infections after Vaccination and Booster with BNT162b2. Nat. Med. 2021, 27, 2108–2110. [Google Scholar] [CrossRef] [PubMed]
- Levine-Tiefenbrun, M.; Yelin, I.; Alapi, H.; Herzel, E.; Kuint, J.; Chodick, G.; Gazit, S.; Patalon, T.; Kishony, R. Waning of SARS-CoV-2 Booster Viral-Load Reduction Effectiveness. Nat. Commun. 2022, 13, 1237. [Google Scholar] [CrossRef]
- Kuhlmann, C.; Mayer, C.K.; Claassen, M.; Maponga, T.; Burgers, W.A.; Keeton, R.; Riou, C.; Sutherland, A.D.; Suliman, T.; Shaw, M.L.; et al. Breakthrough Infections with SARS-CoV-2 Omicron despite MRNA Vaccine Booster Dose. Lancet 2022, 399, 625–626. [Google Scholar] [CrossRef]
- Stoddard, M.; Sarkar, S.; Yuan, L.; Nolan, R.P.; White, D.E.; White, L.F.; Hochberg, N.S.; Chakravarty, A. Beyond the New Normal: Assessing the Feasibility of Vaccine-Based Suppression of SARS-CoV-2. PLoS ONE 2021, 16, e0254734. [Google Scholar] [CrossRef] [PubMed]
- Lauer, S.A.; Grantz, K.H.; Bi, Q.; Jones, F.K.; Zheng, Q.; Meredith, H.R.; Azman, A.S.; Reich, N.G.; Lessler, J. The Incubation Period of Coronavirus Disease 2019 (COVID-19) From Publicly Reported Confirmed Cases: Estimation and Application. Ann. Intern. Med. 2020, 172, 577–582. [Google Scholar] [CrossRef]
- Rocklöv, J.; Sjödin, H.; Wilder-Smith, A. COVID-19 Outbreak on the Diamond Princess Cruise Ship: Estimating the Epidemic Potential and Effectiveness of Public Health Countermeasures. J. Travel Med. 2020, 27, taaa030. [Google Scholar] [CrossRef]
- Underlying Cause of Death 1999–2020. Available online: https://wonder.cdc.gov/wonder/help/ucd.html (accessed on 6 March 2022).
- CDC Surveillance System: Estimated and Projected Population in the U.S. Available online: https://nccd.cdc.gov/ckd/detail.aspx?Qnum=Q155 (accessed on 6 March 2022).
- Population of the U.S. by Sex and Age 2021. Available online: https://www.statista.com/statistics/241488/population-of-the-us-by-sex-and-age/ (accessed on 6 March 2022).
- Buchan, S.A.; Chung, H.; Brown, K.A. Effectiveness of COVID-19 vaccines against omicron or delta symptomatic infection and severe outcomes. medRxiv 2022. [Google Scholar] [CrossRef]
- Klompas, M. Understanding Breakthrough Infections Following MRNA SARS-CoV-2 Vaccination. JAMA 2021, 326, 2018–2020. [Google Scholar] [CrossRef]
- Iyer, A.S.; Jones, F.K.; Nodoushani, A.; Kelly, M.; Becker, M.; Slater, D.; Mills, R.; Teng, E.; Kamruzzaman, M.; Garcia-Beltran, W.F.; et al. Dynamics and Significance of the Antibody Response to SARS-CoV-2 Infection. medRxiv 2020. [Google Scholar] [CrossRef]
- Sanche, S.; Lin, Y.T.; Xu, C.; Romero-Severson, E.; Hengartner, N.; Ke, R. High Contagiousness and Rapid Spread of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. J. 2020, 26, 1470–1477. [Google Scholar] [CrossRef]
- CDC. COVID Data Tracker. Available online: https://covid.cdc.gov/covid-data-tracker (accessed on 6 March 2022).
- Coutinho, R.M.; Marquitti, F.M.D.; Ferreira, L.S.; Borges, M.E.; da Silva, R.L.P.; Canton, O.; Portella, T.P.; Poloni, S.; Franco, C.; Plucinski, M.M.; et al. Model-Based Estimation of Transmissibility and Reinfection of SARS-CoV-2 P.1 Variant. Commun. Med. 2021, 1, 1–8. [Google Scholar] [CrossRef]
- GISAID—NextStrain. Available online: https://www.gisaid.org/phylodynamics/global/nextstrain/ (accessed on 6 March 2022).
- FastStats. Available online: https://www.cdc.gov/nchs/fastats/heart-disease.htm (accessed on 6 March 2022).
- CDC Burden of Influenza. Available online: https://www.cdc.gov/flu/about/burden/index.html (accessed on 6 March 2022).
- United States COVID—Coronavirus Statistics—Worldometer. Available online: https://www.worldometers.info/coronavirus/country/us/ (accessed on 6 March 2022).
- Irons, N.J.; Raftery, A.E. Estimating SARS-CoV-2 Infections from Deaths, Confirmed Cases, Tests, and Random Surveys. Proc. Natl. Acad. Sci. USA 2021, 118, e2103272118. [Google Scholar] [CrossRef]
- Li, R.; Pei, S.; Chen, B.; Song, Y.; Zhang, T.; Yang, W.; Shaman, J. Substantial Undocumented Infection Facilitates the Rapid Dissemination of Novel Coronavirus (SARS-CoV-2). Science 2020, 368, 489–493. [Google Scholar] [CrossRef]
- Rippinger, C.; Bicher, M.; Urach, C.; Brunmeir, D.; Weibrecht, N.; Zauner, G.; Sroczynski, G.; Jahn, B.; Mühlberger, N.; Siebert, U.; et al. Evaluation of Undetected Cases during the COVID-19 Epidemic in Austria. BMC Infect. Dis. 2021, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- CDC. Cases, Data, and Surveillance. Available online: https://www.cdc.gov/coronavirus/2019-ncov/cases-updates/burden.html (accessed on 6 March 2022).
- Daily Testing Trends in the US—Johns Hopkins. Available online: https://coronavirus.jhu.edu/testing/individual-states (accessed on 6 March 2022).
- SARS-CoV-2 Viral Mutations: Impact on COVID-19 Tests. FDA, 14 September 2021. Available online: https://www.fda.gov/medical-devices/coronavirus-covid-19-and-medical-devices/sars-cov-2-viral-mutations-impact-covid-19-tests (accessed on 6 March 2022).
- Carod-Artal, F.J. Post-COVID-19 Syndrome: Epidemiology, Diagnostic Criteria and Pathogenic Mechanisms Involved. Rev. Neurol. 2021, 72, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.C.; Rosen, L.E.; Shepherd, J.G.; Spreafico, R.; da Silva Filipe, A.; Wojcechowskyj, J.A.; Davis, C.; Piccoli, L.; Pascall, D.J.; Dillen, J.; et al. Circulating SARS-CoV-2 Spike N439K Variants Maintain Fitness While Evading Antibody-Mediated Immunity. Cell 2021, 184, 1171–1187.e20. [Google Scholar] [CrossRef] [PubMed]
- Makhoul, M.; Ayoub, H.H.; Chemaitelly, H.; Seedat, S.; Mumtaz, G.R.; Al-Omari, S.; Abu-Raddad, L.J. Epidemiological Impact of SARS-CoV-2 Vaccination: Mathematical Modeling Analyses. Vaccines 2020, 8, E668. [Google Scholar] [CrossRef] [PubMed]
- Iboi, E.A.; Ngonghala, C.N.; Gumel, A.B. Will an Imperfect Vaccine Curtail the COVID-19 Pandemic in the U.S.? Infect. Dis. Model. 2020, 5, 510–524. [Google Scholar] [CrossRef]
- Escandón, K.; Rasmussen, A.L.; Bogoch, I.I.; Murray, E.J.; Escandón, K.; Popescu, S.V.; Kindrachuk, J. COVID-19 False Dichotomies and a Comprehensive Review of the Evidence Regarding Public Health, COVID-19 Symptomatology, SARS-CoV-2 Transmission, Mask Wearing, and Reinfection. BMC Infect. Dis. 2021, 21, 710. [Google Scholar] [CrossRef] [PubMed]
- FastStats. Available online: https://www.cdc.gov/nchs/fastats/leading-causes-of-death.htm (accessed on 6 March 2022).
- Woolf, S.H.; Masters, R.K.; Aron, L.Y. Effect of the Covid-19 Pandemic in 2020 on Life Expectancy across Populations in the USA and Other High Income Countries: Simulations of Provisional Mortality Data. BMJ 2021, 373, n1343. [Google Scholar] [CrossRef]
- Mazzuco, S.; Campostrini, S. Life Expectancy Drop in 2020. Estimates Based on Human Mortality Database. PLoS ONE 2022, 17, e0262846. [Google Scholar] [CrossRef] [PubMed]
- King, K.L.; Wilson, S.; Napolitano, J.M.; Sell, K.J.; Rennert, L.; Parkinson, C.L.; Dean, D. SARS-CoV-2 Variants of Concern Alpha and Delta Show Increased Viral Load in Saliva. medRxiv 2022. [Google Scholar] [CrossRef]
- Teyssou, E.; Delagrèverie, H.; Visseaux, B.; Lambert-Niclot, S.; Brichler, S.; Ferre, V.; Marot, S.; Jary, A.; Todesco, E.; Schnuriger, A.; et al. The Delta SARS-CoV-2 Variant Has a Higher Viral Load than the Beta and the Historical Variants in Nasopharyngeal Samples from Newly Diagnosed COVID-19 Patients. J. Infect. 2021, 83, e1–e3. [Google Scholar] [CrossRef]
- Kidd, M.; Richter, A.; Best, A.; Cumley, N.; Mirza, J.; Percival, B.; Mayhew, M.; Megram, O.; Ashford, F.; White, T.; et al. S-Variant SARS-CoV-2 Lineage B1.1.7 Is Associated with Significantly Higher Viral Load in Samples Tested by TaqPath Polymerase Chain Reaction. J. Infect. Dis. 2021, 223, 1666–1670. [Google Scholar] [CrossRef]
- Spiesschaert, B.; McFadden, G.; Hermans, K.; Nauwynck, H.; Van de Walle, G.R. The Current Status and Future Directions of Myxoma Virus, a Master in Immune Evasion. Vet. Res. 2011, 42, 76. [Google Scholar] [CrossRef]
- Bouayad, A. Innate Immune Evasion by SARS-CoV-2: Comparison with SARS-CoV. Rev. Med. Virol. 2020, 30, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hayn, M.; Hirschenberger, M.; Koepke, L.; Nchioua, R.; Straub, J.H.; Klute, S.; Hunszinger, V.; Zech, F.; Prelli Bozzo, C.; Aftab, W.; et al. Systematic Functional Analysis of SARS-CoV-2 Proteins Uncovers Viral Innate Immune Antagonists and Remaining Vulnerabilities. Cell Rep. 2021, 35, 109126. [Google Scholar] [CrossRef] [PubMed]
- Setaro, A.C.; Gaglia, M.M. All Hands on Deck: SARS-CoV-2 Proteins That Block Early Anti-Viral Interferon Responses. Curr. Res. Virol. Sci. 2021, 2, 100015. [Google Scholar] [CrossRef]
- Xia, H.; Cao, Z.; Xie, X.; Zhang, X.; Chen, J.Y.-C.; Wang, H.; Menachery, V.D.; Rajsbaum, R.; Shi, P.-Y. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020, 33, 108234. [Google Scholar] [CrossRef]
- Chen, D.-Y.; Khan, N.; Close, B.J.; Goel, R.K.; Blum, B.; Tavares, A.H.; Kenney, D.; Conway, H.L.; Ewoldt, J.K.; Chitalia, V.C.; et al. SARS-CoV-2 Disrupts Proximal Elements in the JAK-STAT Pathway. J. Virol. 2021, 95, e0086221. [Google Scholar] [CrossRef]
- Han, L.; Zhuang, M.-W.; Deng, J.; Zheng, Y.; Zhang, J.; Nan, M.-L.; Zhang, X.-J.; Gao, C.; Wang, P.-H. SARS-CoV-2 ORF9b Antagonizes Type I and III Interferons by Targeting Multiple Components of the RIG-I/MDA-5-MAVS, TLR3-TRIF, and CGAS-STING Signaling Pathways. J. Med. Virol. 2021, 93, 5376–5389. [Google Scholar] [CrossRef]
- Shi, C.-S.; Qi, H.-Y.; Boularan, C.; Huang, N.-N.; Abu-Asab, M.; Shelhamer, J.H.; Kehrl, J.H. SARS-Coronavirus Open Reading Frame-9b Suppresses Innate Immunity by Targeting Mitochondria and the MAVS/TRAF3/TRAF6 Signalosome. J. Immunol. 2014, 193, 3080–3089. [Google Scholar] [CrossRef]
- Dominguez Andres, A.; Feng, Y.; Campos, A.R.; Yin, J.; Yang, C.-C.; James, B.; Murad, R.; Kim, H.; Deshpande, A.J.; Gordon, D.E.; et al. SARS-CoV-2 ORF9c Is a Membrane-Associated Protein That Suppresses Antiviral Responses in Cells. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lu, F. SARS-CoV-2 ORF9c: A Mysterious Membrane-Anchored Protein That Regulates Immune Evasion? Nat. Rev. Immunol. 2020, 20, 648. [Google Scholar] [CrossRef] [PubMed]
- Thoms, M.; Buschauer, R.; Ameismeier, M.; Koepke, L.; Denk, T.; Hirschenberger, M.; Kratzat, H.; Hayn, M.; Mackens-Kiani, T.; Cheng, J.; et al. Structural Basis for Translational Shutdown and Immune Evasion by the Nsp1 Protein of SARS-CoV-2. Science 2020, 369, 1249–1255. [Google Scholar] [CrossRef]
- Kumar, A.; Ishida, R.; Strilets, T.; Cole, J.; Lopez-Orozco, J.; Fayad, N.; Felix-Lopez, A.; Elaish, M.; Evseev, D.; Magor, K.E.; et al. SARS-CoV-2 Nonstructural Protein 1 Inhibits the Interferon Response by Causing Depletion of Key Host Signaling Factors. J. Virol. 2021, 95, e0026621. [Google Scholar] [CrossRef]
- Li, J.-Y.; Liao, C.-H.; Wang, Q.; Tan, Y.-J.; Luo, R.; Qiu, Y.; Ge, X.-Y. The ORF6, ORF8 and Nucleocapsid Proteins of SARS-CoV-2 Inhibit Type I Interferon Signaling Pathway. Virus Res. 2020, 286, 198074. [Google Scholar] [CrossRef]
- Kopecky-Bromberg, S.A.; Martínez-Sobrido, L.; Frieman, M.; Baric, R.A.; Palese, P. Severe Acute Respiratory Syndrome Coronavirus Open Reading Frame (ORF) 3b, ORF 6, and Nucleocapsid Proteins Function as Interferon Antagonists. J. Virol. 2007, 81, 548–557. [Google Scholar] [CrossRef]
- Li, M.; Liu, J.; Lu, R.; Zhang, Y.; Du, M.; Xing, M.; Wu, Z.; Kong, X.; Zhu, Y.; Zhou, X.; et al. Longitudinal Immune Profiling Reveals Dominant Epitopes Mediating Long-Term Humoral Immunity in COVID-19-Convalescent Individuals. J. Allergy Clin. Immunol. 2022, 149, 1225–1241. [Google Scholar] [CrossRef]
- Premkumar, L.; Segovia-Chumbez, B.; Jadi, R.; Martinez, D.R.; Raut, R.; Markmann, A.; Cornaby, C.; Bartelt, L.; Weiss, S.; Park, Y.; et al. The Receptor Binding Domain of the Viral Spike Protein Is an Immunodominant and Highly Specific Target of Antibodies in SARS-CoV-2 Patients. Sci. Immunol. 2020, 5, eabc8413. [Google Scholar] [CrossRef]
- Thorne, L.G.; Bouhaddou, M.; Reuschl, A.-K.; Zuliani-Alvarez, L.; Polacco, B.; Pelin, A.; Batra, J.; Whelan, M.V.X.; Hosmillo, M.; Fossati, A.; et al. Evolution of Enhanced Innate Immune Evasion by SARS-CoV-2. Nature 2022, 602, 487–495. [Google Scholar] [CrossRef]
- Stoddard, M.; Yuan, L.; Sarkar, S.; Mangalaganesh, S.; Nolan, R.P.; Bottino, D.; Hather, G.; Hochberg, N.S.; White, L.F.; Chakravarty, A. Heterogeneity in vaccinal immunity to SARS-CoV-2 can be addressed by a personalized booster strategy. medRxiv 2022. [CrossRef]
- Faes, C.; Abrams, S.; Van Beckhoven, D.; Meyfroidt, G.; Vlieghe, E.; Hens, N. Time between Symptom Onset, Hospitalisation and Recovery or Death: Statistical Analysis of Belgian COVID-19 Patients. Int. J. Environ. Res. Public Health 2020, 17, 7560. [Google Scholar] [CrossRef]
- The Contribution of Nosocomial Infections to the First Wave. Public Health England and the London School of Hygiene and Tropical Medicine, 28 January 2021. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/961210/S1056_Contribution_of_nosocomial_infections_to_the_first_wave.pdf (accessed on 6 March 2022).
- Donnelly, C.A.; Ghani, A.C.; Leung, G.M.; Hedley, A.J.; Fraser, C.; Riley, S.; Abu-Raddad, L.J.; Ho, L.-M.; Thach, T.-Q.; Chau, P.; et al. Epidemiological Determinants of Spread of Causal Agent of Severe Acute Respiratory Syndrome in Hong Kong. Lancet 2003, 361, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Breiman, R.F.; Evans, M.R.; Preiser, W.; Maguire, J.; Schnur, A.; Bekedam, H.; MacKenzie, J.S. Role of China in the Quest to Define and Control Severe Acute Respiratory Syndrome. Emerg. Infect. Dis. 2003, 9, 1037–1041. [Google Scholar] [CrossRef]
- Al Hajjar, S.; Memish, Z.A.; McIntosh, K. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): A Perpetual Challenge. Ann. Saudi Med. 2013, 33, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.-R.; Shi, W.-Q.; Liu, K.; Li, X.-L.; Liu, M.-J.; Zhang, W.-H.; Zhao, G.-P.; Chen, J.-J.; Zhang, X.-A.; Miao, D.; et al. Epidemiology and Evolution of Middle East Respiratory Syndrome Coronavirus, 2012–2020. Infect. Dis. Poverty 2021, 10, 66. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, S.H.; Maher, A.D.; Kanagasabai, U.; Alfaraj, S.H.; Alzahrani, N.A.; Alqahtani, S.A.; Assiri, A.M.; Memish, Z.A. MERS-CoV Confirmation Among 6,873 Suspected Persons and Relevant Epidemiologic and Clinical Features, Saudi Arabia—2014 to 2019. EClinicalMedicine 2021, 41, 101191. [Google Scholar] [CrossRef]
- van der Ley, P.A.; Zariri, A.; van Riet, E.; Oosterhoff, D.; Kruiswijk, C.P. An Intranasal OMV-Based Vaccine Induces High Mucosal and Systemic Protecting Immunity Against a SARS-CoV-2 Infection. Front Immunol. 2021, 12, 781280. [Google Scholar] [CrossRef]
- Bošnjak, B.; Odak, I.; Barros-Martins, J.; Sandrock, I.; Hammerschmidt, S.I.; Permanyer, M.; Patzer, G.E.; Greorgiev, H.; Gutierrez Jauregui, R.; Tscherne, A.; et al. Intranasal Delivery of MVA Vector Vaccine Induces Effective Pulmonary Immunity Against SARS-CoV-2 in Rodents. Front Immunol. 2021, 12, 772240. [Google Scholar] [CrossRef]
- Afkhami, S.; D’Agostino, M.R.; Zhang, A.; Stacey, H.D.; Marzok, A.; Kang, A.; Singh, R.; Bavananthasivam, J.; Ye, G.; Luo, X.; et al. Respiratory Mucosal Delivery of Next-Generation COVID-19 Vaccine Provides Robust Protection against Both Ancestral and Variant Strains of SARS-CoV-2. Cell 2022, 185, 896–915.e19. [Google Scholar] [CrossRef]
- Jiang, L.; Driedonks, T.A.P.; Jong, W.S.P.; Dhakal, S.; van den Berg van Saparoea, H.B.; Sitaras, I.; Zhou, R.; Caputo, C.; Littlefield, K.; Lowman, M.; et al. A Bacterial Extracellular Vesicle-Based Intranasal Vaccine against SARS-CoV-2 Protects against Disease and Elicits Neutralizing Antibodies to Wild-Type and Delta Variants. bioRxiv 2022. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, P.; Wong, Y.-C.; Xu, H.; Lau, S.-Y.; Liu, L.; Mok, B.W.-Y.; Peng, Q.; Liu, N.; Woo, K.-F.; et al. Nasal Prevention of SARS-CoV-2 Infection by Intranasal Influenza-Based Boost Vaccination in Mouse Models. EBioMedicine 2022, 75, 103762. [Google Scholar] [CrossRef]
- Cao, H.; Mai, J.; Zhou, Z.; Li, Z.; Duan, R.; Watt, J.; Chen, Z.; Bandara, R.A.; Li, M.; Ahn, S.K.; et al. Intranasal HD-Ad Vaccine Protects the Upper and Lower Respiratory Tracts of HACE2 Mice against SARS-CoV-2. Cell Biosci. 2021, 11, 202. [Google Scholar] [CrossRef]
- Thapa, H.B.; Müller, A.M.; Camilli, A.; Schild, S. An Intranasal Vaccine Based on Outer Membrane Vesicles Against SARS-CoV-2. Front. Microbiol. 2021, 12, 752739. [Google Scholar] [CrossRef]
- Liu, X.; Luongo, C.; Matsuoka, Y.; Park, H.-S.; Santos, C.; Yang, L.; Moore, I.N.; Afroz, S.; Johnson, R.F.; Lafont, B.A.P.; et al. A Single Intranasal Dose of a Live-Attenuated Parainfluenza Virus-Vectored SARS-CoV-2 Vaccine Is Protective in Hamsters. Proc. Natl. Acad. Sci. USA 2021, 118, e2109744118. [Google Scholar] [CrossRef]
- King, R.G.; Silva-Sanchez, A.; Peel, J.N.; Botta, D.; Dickson, A.M.; Pinto, A.K.; Meza-Perez, S.; Allie, S.R.; Schultz, M.D.; Liu, M.; et al. Single-Dose Intranasal Administration of AdCOVID Elicits Systemic and Mucosal Immunity against SARS-CoV-2 and Fully Protects Mice from Lethal Challenge. Vaccines 2021, 9, 881. [Google Scholar] [CrossRef]
- Abaluck, J.; Kwong, L.H.; Styczynski, A.; Haque, A.; Kabir, M.A.; Bates-Jefferys, E.; Crawford, E.; Benjamin-Chung, J.; Raihan, S.; Rahman, S.; et al. Impact of community masking on COVID-19: A cluster-randomized trial in Bangladesh. Science 2022, 375, p.eabi9069. [Google Scholar] [CrossRef]
- Nazarenko, Y. Air Filtration and SARS-CoV-2. Epidemiol. Health 2020, 42, e2020049. [Google Scholar] [CrossRef]
- Cinaglia, P.; Cannataro, M. Forecasting COVID-19 Epidemic Trends by Combining a Neural Network with Rt Estimation. Entropy 2022, 24, 929. [Google Scholar] [CrossRef]
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Xiao, J.; Hooper, A.T.; Hamilton, J.D.; Musser, B.J.; et al. REGEN-COV antibody combination and outcomes in outpatients with Covid-19. N. Engl. J. Med. 2021, 385, e81. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed Parameter | Symbol | Value | Source |
|---|---|---|---|
| Latency period | 1/ | 3 days | [69] |
| Infectious period | 1/ | 10 days | [70] |
| US population birth rate | 0.9% annually | Fixed to death rate | |
| US population death rate | 0.9% annually | [71] | |
| US population size | N | 330 million | [72] |
| US population over 65 years | N>65 | 54 million | [73] |
| US population under 65 years | N<65 | 276 million | [73] |
| Variable Parameter | Symbol | Value | Source |
|---|---|---|---|
| Vaccine reduction in risk of infection | Vei | 0%, 50%, 90% | [74] |
| Vaccine reduction in risk of mortality | Vem | 70%, 90% | [75] |
| Duration of natural immunity | 1/ | 3 to 24 months | [76] |
| Intrinsic reproductive number | R0 | 2 to 9 individuals | [77] |
| Population IFR for unvaccinated individuals | 0.05% to 5% | See Table S3 | |
| Fraction vaccinated, over 65 years | f>65 | 90%, 100% | [78] |
| Fraction vaccinated, under 65 years | f<65 | 70%, 100% | [78] |
| Infections | IFR 0.5% | IFR 1% | IFR 2% |
|---|---|---|---|
| 50 million | 250,000 | 500,000 | 1,000,000 |
| 100 million | 500,000 | 1,000,000 | 2,000,000 |
| 200 million | 1,000,000 | 2,000,000 | 4,000,000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoddard, M.; Novokhodko, A.; Sarkar, S.; Van Egeren, D.; White, L.F.; Hochberg, N.S.; Rogers, M.S.; Zetter, B.; Joseph-McCarthy, D.; Chakravarty, A. Endemicity Is Not a Victory: The Unmitigated Downside Risks of Widespread SARS-CoV-2 Transmission. COVID 2022, 2, 1689-1709. https://doi.org/10.3390/covid2120121
Stoddard M, Novokhodko A, Sarkar S, Van Egeren D, White LF, Hochberg NS, Rogers MS, Zetter B, Joseph-McCarthy D, Chakravarty A. Endemicity Is Not a Victory: The Unmitigated Downside Risks of Widespread SARS-CoV-2 Transmission. COVID. 2022; 2(12):1689-1709. https://doi.org/10.3390/covid2120121
Chicago/Turabian StyleStoddard, Madison, Alexander Novokhodko, Sharanya Sarkar, Debra Van Egeren, Laura F. White, Natasha S. Hochberg, Michael S. Rogers, Bruce Zetter, Diane Joseph-McCarthy, and Arijit Chakravarty. 2022. "Endemicity Is Not a Victory: The Unmitigated Downside Risks of Widespread SARS-CoV-2 Transmission" COVID 2, no. 12: 1689-1709. https://doi.org/10.3390/covid2120121
APA StyleStoddard, M., Novokhodko, A., Sarkar, S., Van Egeren, D., White, L. F., Hochberg, N. S., Rogers, M. S., Zetter, B., Joseph-McCarthy, D., & Chakravarty, A. (2022). Endemicity Is Not a Victory: The Unmitigated Downside Risks of Widespread SARS-CoV-2 Transmission. COVID, 2(12), 1689-1709. https://doi.org/10.3390/covid2120121