
Anti-SARS-CoV-2 Activity of Adamantanes In Vitro and in Animal Models of Infection

 , , , , , and
, , , , , and 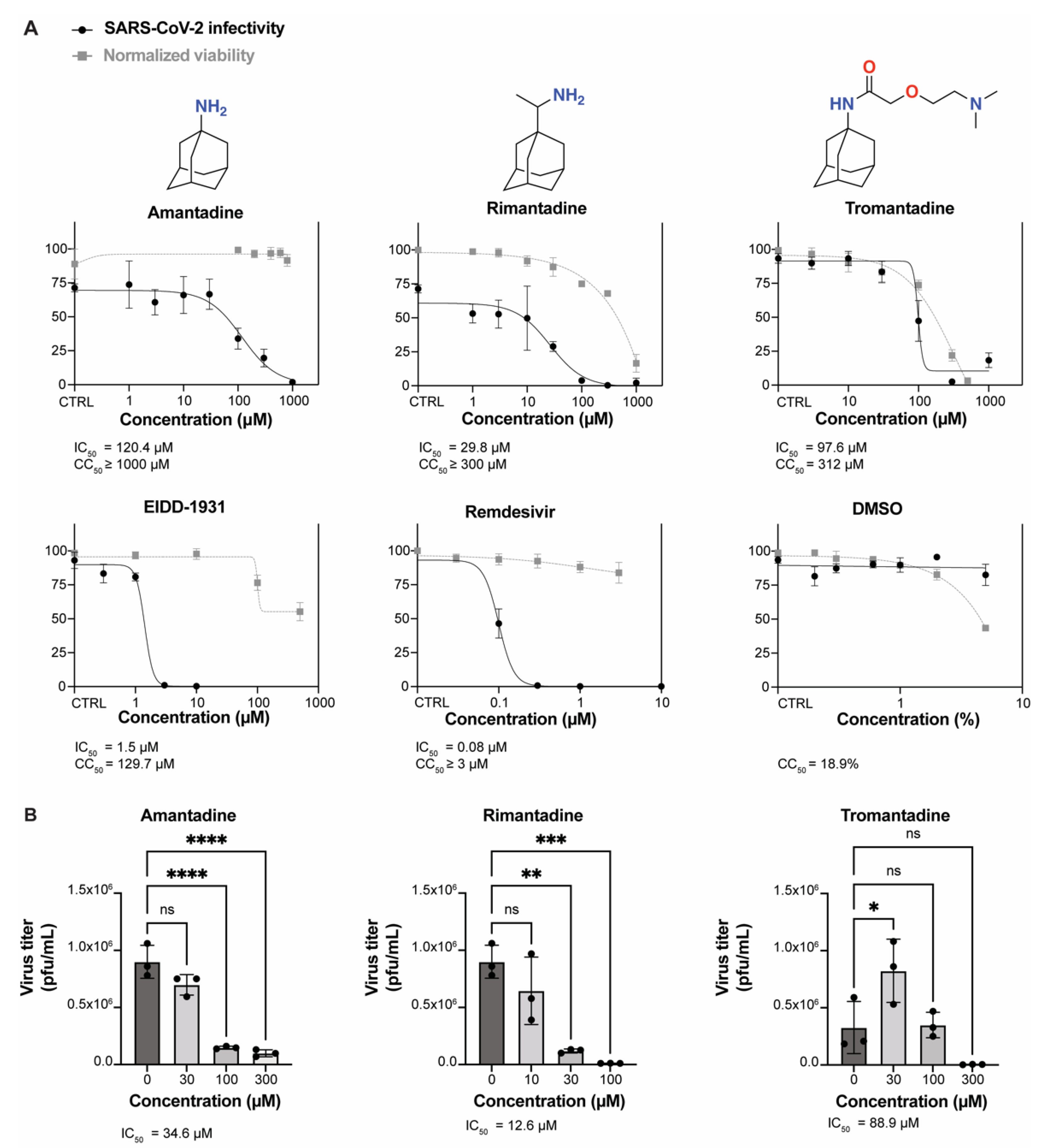{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antiviral Compounds
2.3. Antiviral Assays
2.4. Immunostaining Assay
2.5. SARS-CoV-2 Plaque Assays
2.6. Viability Assay
2.7. Animal Antiviral Studies
2.8. Statistical Analysis
3. Results
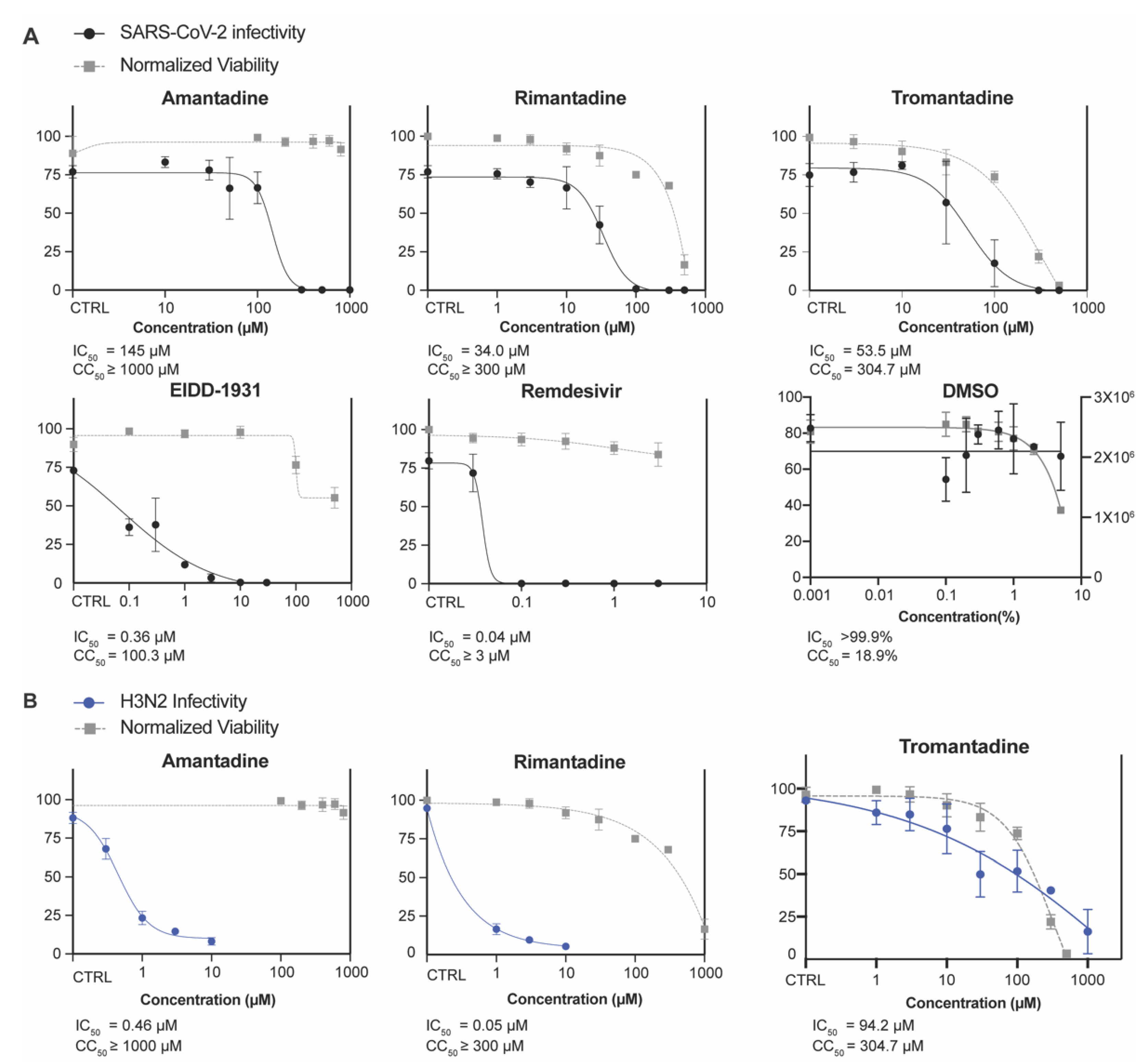
3.1. Amantadine, Rimantadine, and Tromantadine Inhibit SARS-CoV-2 In Vitro in Human Lung Epithelial Cells
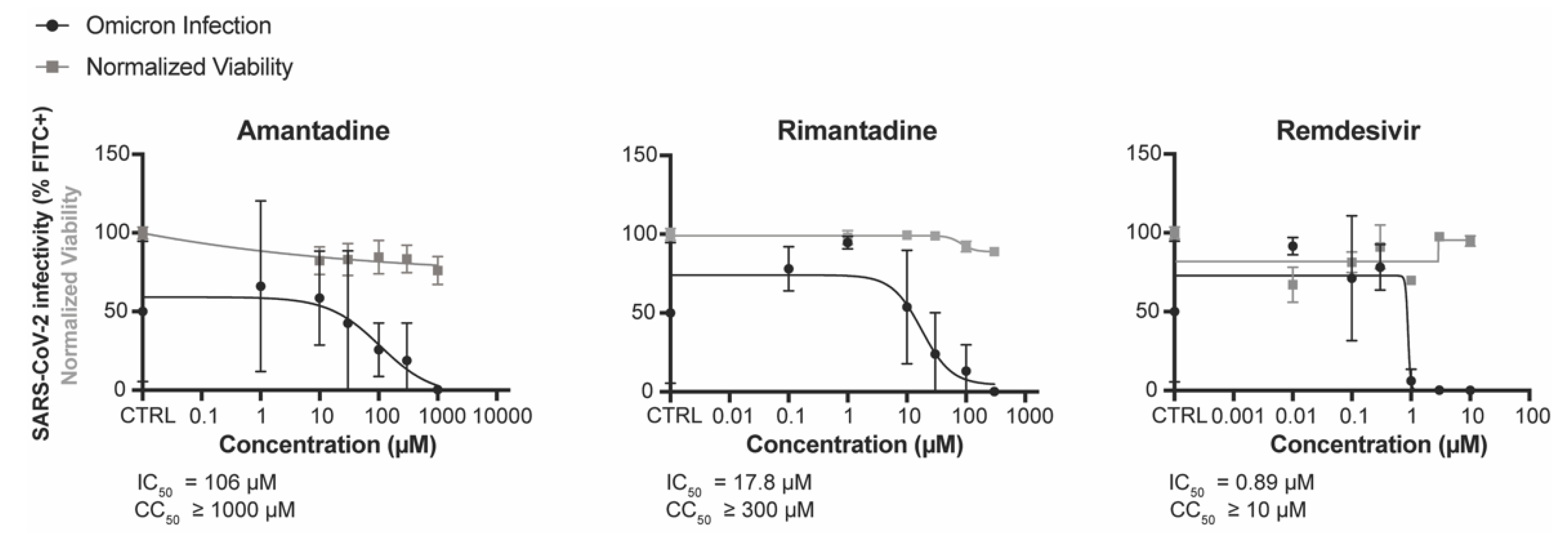
3.2. Amantadine and Rimantadine Inhibit SARS-CoV-2 Omicron Variant B.1.1.529 In Vitro in Vero E6-TMPRSS2-T2A-ACE2 (Vero E6 T/A) Cells
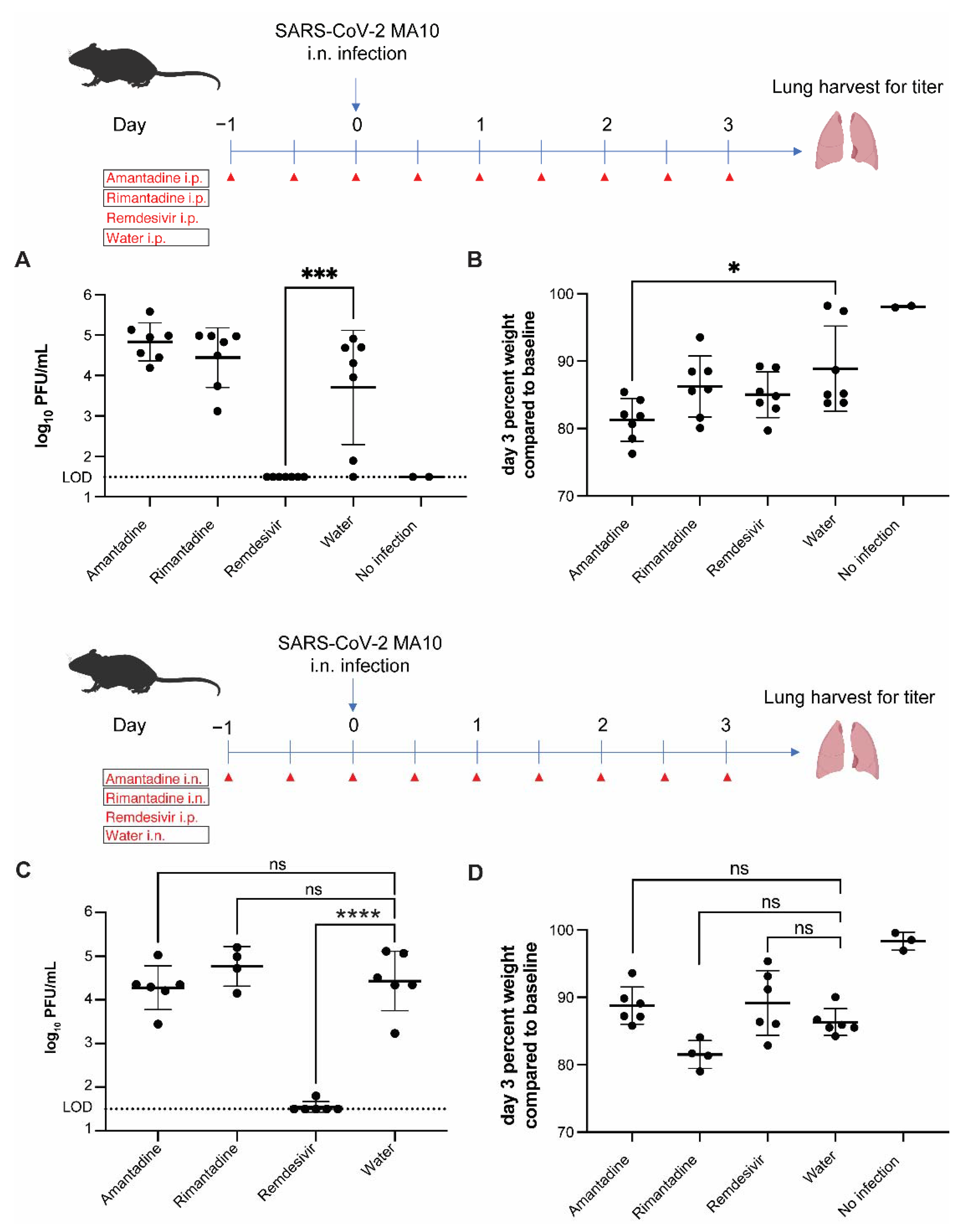
3.3. Amantadine and Rimantadine Do Not Significantly Inhibit SARS-CoV-2 Growth in Lungs of Mice When Administered by Intraperitoneal or Intranasal Route
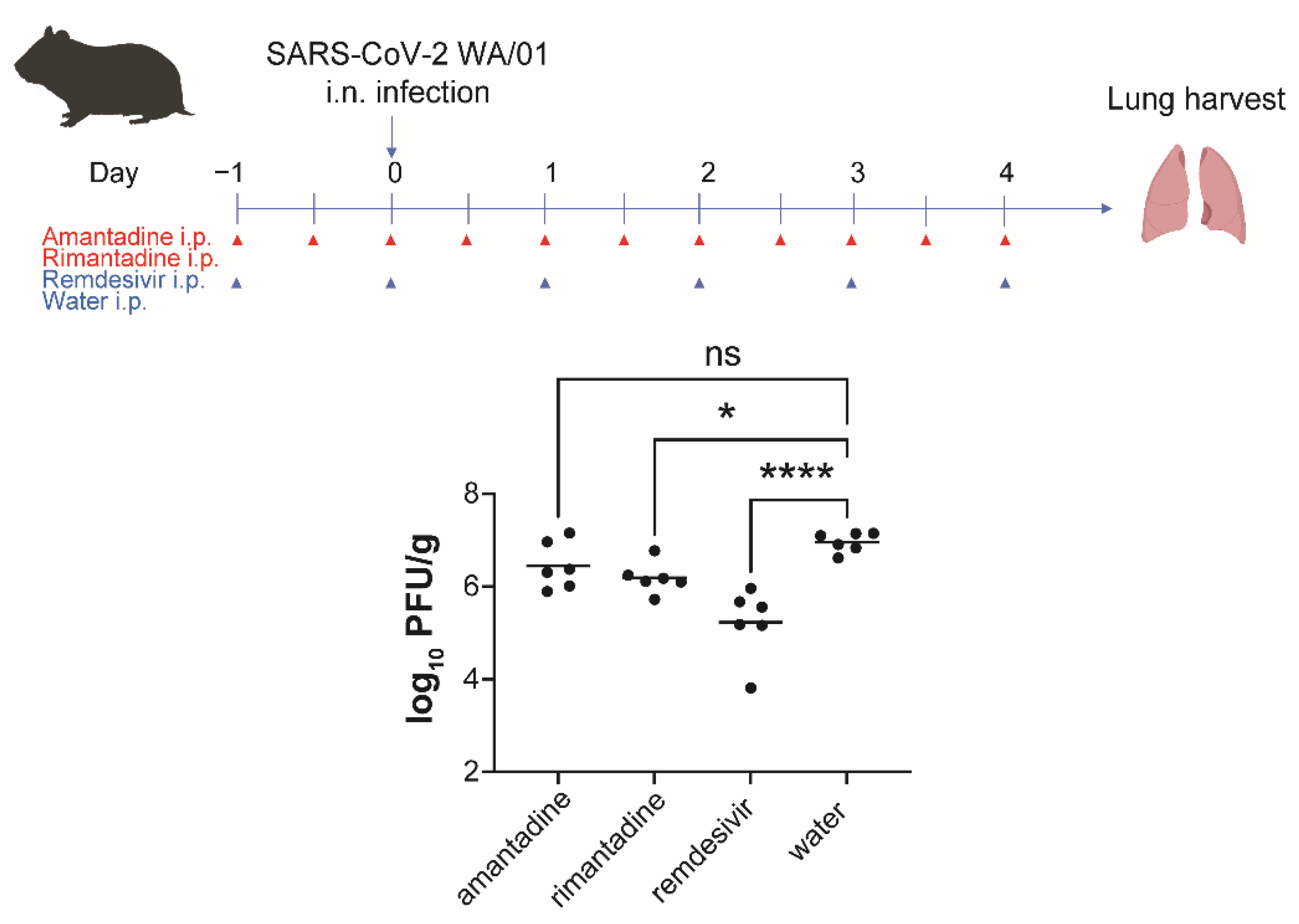
3.4. Rimantadine Inhibits SARS-CoV-2 Growth in Lungs of Hamsters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uyeki, T.M.; Bernstein, H.H.; Bradley, J.S.; Englund, J.A.; File, T.M.; Fry, A.M.; Gravenstein, S.; Hayden, F.G.; Harper, S.A.; Hirshon, J.M.; et al. Clinical Practice Guidelines by the Infectious Diseases Society of America: 2018 Update on Diagnosis, Treatment, Chemoprophylaxis, and Institutional Outbreak Management of Seasonal Influenza A. Clin. Infect Dis. 2019, 68, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Abreu, G.E.; Aranda-Martinez, J.D.; Araujo, R. Use of amantadine in a patient with SARS-CoV-2. J. Med. Virol. 2020, 93, 110–111. [Google Scholar] [CrossRef]
- Cortes Borra, A. Does amantadine have a protective effect against COVID-19? Pol. J. Neurol. Neurosurg. 2020, 54, 284–285. [Google Scholar] [CrossRef]
- Rejdak, K.; Grieb, P. Adamantanes might be protective from COVID-19 in patients with neurological diseases: Multiple sclerosis, parkinsonism and cognitive impairment. Mult. Scler. Relat. Disord. 2020, 42, 102163. [Google Scholar] [CrossRef] [PubMed]
- Kamel, W.A.; Kamel, M.I.; Alhasawi, A.; Elmasry, S.; AlHamdan, F.; Al-Hashel, J.Y. Effect of Pre-exposure Use of Amantadine on COVID-19 Infection: A Hospital-Based Cohort Study in Patients with Parkinson’s Disease or Multiple Sclerosis. Front. Neurol. 2021, 12, 704186. [Google Scholar] [CrossRef]
- Mathur, A.; Beare, A.S.; Reed, S.E. In vitro antiviral activity and preliminary clinical trials of a new adamantane compound. Antimicrob. Agents Chemother. 1973, 4, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Brison, E.; Jacomy, H.; Desforges, M.; Talbot, P.J. Novel treatment with neuroprotective and antiviral properties against a neuroinvasive human respiratory virus. J. Virol. 2014, 88, 1548–1563. [Google Scholar] [CrossRef]
- Chen, F.; Chan, K.H.; Jiang, Y.; Kao, R.Y.; Lu, H.T.; Fan, K.W.; Cheng, V.C.; Tsui, W.H.; Hung, I.F.; Lee, T.S.; et al. In vitro susceptibility of 10 clinical isolates of SARS coronavirus to selected antiviral compounds. J. Clin. Virol. 2004, 31, 69–75. [Google Scholar] [CrossRef]
- Tanner, J.A.; Zheng, B.J.; Zhou, J.; Watt, R.M.; Jiang, J.Q.; Wong, K.L.; Lin, Y.P.; Lu, L.Y.; He, M.L.; Kung, H.F.; et al. The adamantane-derived bananins are potent inhibitors of the helicase activities and replication of SARS coronavirus. Chem. Biol. 2005, 12, 303–311. [Google Scholar] [CrossRef]
- Daniloski, Z.; Jordan, T.X.; Wessels, H.H.; Hoagland, D.A.; Kasela, S.; Legut, M.; Maniatis, S.; Mimitou, E.P.; Lu, L.; Geller, E.; et al. Identification of Required Host Factors for SARS-CoV-2 Infection in Human Cells. Cell 2021, 184, 92–105.e116. [Google Scholar] [CrossRef]
- Xie, X.; Lokugamage, K.G.; Zhang, X.; Vu, M.N.; Muruato, A.E.; Menachery, V.D.; Shi, P.Y. Engineering SARS-CoV-2 using a reverse genetic system. Nat. Protoc. 2021, 16, 1761–1784. [Google Scholar] [CrossRef]
- ter Meulen, J.; van den Brink, E.N.; Poon, L.L.; Marissen, W.E.; Leung, C.S.; Cox, F.; Cheung, C.Y.; Bakker, A.Q.; Bogaards, J.A.; van Deventer, E.; et al. Human monoclonal antibody combination against SARS coronavirus: Synergy and coverage of escape mutants. PLoS Med. 2006, 3, e237. [Google Scholar] [CrossRef]
- Rejdak, K.; Fiedor, P.; Bonek, R.; Goch, A.; Gala-Bladzinska, A.; Chelstowski, W.; Lukasiak, J.; Kiciak, S.; Dabrowski, P.; Dec, M.; et al. The use of amantadine in the prevention of progression and treatment of COVID-19 symptoms in patients infected with the SARS-CoV-2 virus (COV-PREVENT): Study rationale and design. Contemp. Clin. Trials 2022, 116, 106755. [Google Scholar] [CrossRef] [PubMed]
- Fink, K.; Nitsche, A.; Neumann, M.; Grossegesse, M.; Eisele, K.H.; Danysz, W. Amantadine Inhibits SARS-CoV-2 In Vitro. Viruses 2021, 13, 539. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gammeltoft, K.A.; Galli, A.; Offersgaard, A.; Fahnoe, U.; Ramirez, S.; Bukh, J.; Gottwein, J.M. Efficacy of Ion-Channel Inhibitors Amantadine, Memantine and Rimantadine for the Treatment of SARS-CoV-2 In Vitro. Viruses 2021, 13, 2082. [Google Scholar] [CrossRef] [PubMed]
- Leist, S.R.; Dinnon, K.H., 3rd; Schafer, A.; Tse, L.V.; Okuda, K.; Hou, Y.J.; West, A.; Edwards, C.E.; Sanders, W.; Fritch, E.J.; et al. A Mouse-Adapted SARS-CoV-2 Induces Acute Lung Injury and Mortality in Standard Laboratory Mice. Cell 2020, 183, 1070–1085.e1012. [Google Scholar] [CrossRef]
- Vernier, V.G.; Harmon, J.B.; Stump, J.M.; Lynes, T.E.; Marvel, J.P.; Smith, D.H. The toxicologic and pharmacologic properties of amantadine hydrochloride. Toxicol. Appl. Pharmacol. 1969, 15, 642–665. [Google Scholar] [CrossRef]
- Chan, J.F.; Zhang, A.J.; Yuan, S.; Poon, V.K.; Chan, C.C.; Lee, A.C.; Chan, W.M.; Fan, Z.; Tsoi, H.W.; Wen, L.; et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility. Clin. Infect. Dis. 2020, 71, 2428–2446. [Google Scholar] [CrossRef]
- Rosenke, K.; Meade-White, K.; Letko, M.; Clancy, C.; Hansen, F.; Liu, Y.; Okumura, A.; Tang-Huau, T.L.; Li, R.; Saturday, G.; et al. Defining the Syrian hamster as a highly susceptible preclinical model for SARS-CoV-2 infection. Emerg. Microbes Infect. 2020, 9, 2673–2684. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- Brigham, E.F.; Johnston, T.H.; Brown, C.; Holt, J.D.S.; Fox, S.H.; Hill, M.P.; Howson, P.A.; Brotchie, J.M.; Nguyen, J.T. Pharmacokinetic/Pharmacodynamic Correlation Analysis of Amantadine for Levodopa-Induced Dyskinesia. J. Pharmacol. Exp. Ther. 2018, 367, 373–381. [Google Scholar] [CrossRef]
- Hoffman, H.E.; Gaylord, J.C.; Blasecki, J.W.; Shalaby, L.M.; Whitney, C.C., Jr. Pharmacokinetics and metabolism of rimantadine hydrochloride in mice and dogs. Antimicrob. Agents Chemother. 1988, 32, 1699–1704. [Google Scholar] [CrossRef]
- Toft-Bertelsen, T.L.; Jeppesen, M.G.; Tzortzini, E.; Xue, K.; Giller, K.; Becker, S.; Mujezinovic, A.; Bentzen, B.H.; Andreas, L.B.; Kolocouris, A.; et al. Author Correction: Amantadine inhibits known and novel ion channels encoded by SARS-CoV-2 in vitro. Commun. Biol. 2021, 4, 1402. [Google Scholar] [CrossRef] [PubMed]
- Mandala, V.S.; McKay, M.J.; Shcherbakov, A.A.; Dregni, A.J.; Kolocouris, A.; Hong, M. Structure and drug binding of the SARS-CoV-2 envelope protein transmembrane domain in lipid bilayers. Nat. Struct. Mol. Biol. 2020, 27, 1202–1208. [Google Scholar] [CrossRef]
- Harrison, N.L.; Abbott, G.W.; Gentzsch, M.; Aleksandrov, A.; Moroni, A.; Thiel, G.; Grant, S.; Nichols, C.G.; Lester, H.A.; Hartel, A.; et al. How many SARS-CoV-2 “viroporins” are really ion channels? Commun. Biol. 2022, 5, 859. [Google Scholar] [CrossRef]
- deVries, T.; Dentiste, A.; Handiwala, L.; Jacobs, D. Bioavailability and Pharmacokinetics of Once-Daily Amantadine Extended-Release Tablets in Healthy Volunteers: Results from Three Randomized, Crossover, Open-Label, Phase 1 Studies. Neurol. Ther. 2019, 8, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Hayden, F.G.; Hoffman, H.E.; Spyker, D.A. Differences in side effects of amantadine hydrochloride and rimantadine hydrochloride relate to differences in pharmacokinetics. Antimicrob. Agents Chemother. 1983, 23, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Sperber, S.J.; Hayden, F.G. Antiviral Chemotherapyand Prophylaxis of Viral Respiratory Disease. Clin. Lab. Med. 1987, 7, 869–896. [Google Scholar] [CrossRef]
- U.S. National Library of Medicine. Available online: https://clinicaltrials.gov/ (accessed on 3 October 2022).
- Keyser, L.A.; Karl, M.; Nafziger, A.N.; Bertino, J.S., Jr. Comparison of central nervous system adverse effects of amantadine and rimantadine used as sequential prophylaxis of influenza A in elderly nursing home patients. Arch. Intern. Med. 2000, 160, 1485–1488. [Google Scholar] [CrossRef] [PubMed]
- Ilyushina, N.A.; Hoffmann, E.; Salomon, R.; Webster, R.G.; Govorkova, E.A. Amantadine-oseltamivir combination therapy for H5N1 influenza virus infection in mice. Antivir. Ther. 2007, 12, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Hurst, B.L.; Wong, M.H.; Bailey, K.W.; Morrey, J.D. Effects of double combinations of amantadine, oseltamivir, and ribavirin on influenza A (H5N1) virus infections in cell culture and in mice. Antimicrob. Agents Chemother. 2009, 53, 2120–2128. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.-Y.; Guo, Z.; Liu, P.; McKay, L.G.A.; Storm, N.; Griffiths, A.; Qu, M.D.; Finberg, R.W.; Somasundaran, M.; Wang, J.P. Anti-SARS-CoV-2 Activity of Adamantanes In Vitro and in Animal Models of Infection. COVID 2022, 2, 1551-1563. https://doi.org/10.3390/covid2110111
Lim S-Y, Guo Z, Liu P, McKay LGA, Storm N, Griffiths A, Qu MD, Finberg RW, Somasundaran M, Wang JP. Anti-SARS-CoV-2 Activity of Adamantanes In Vitro and in Animal Models of Infection. COVID. 2022; 2(11):1551-1563. https://doi.org/10.3390/covid2110111
Chicago/Turabian StyleLim, Sun-Young, Zhiru Guo, Ping Liu, Lindsay G. A. McKay, Nadia Storm, Anthony Griffiths, Ming Da Qu, Robert W. Finberg, Mohan Somasundaran, and Jennifer P. Wang. 2022. "Anti-SARS-CoV-2 Activity of Adamantanes In Vitro and in Animal Models of Infection" COVID 2, no. 11: 1551-1563. https://doi.org/10.3390/covid2110111
APA StyleLim, S.-Y., Guo, Z., Liu, P., McKay, L. G. A., Storm, N., Griffiths, A., Qu, M. D., Finberg, R. W., Somasundaran, M., & Wang, J. P. (2022). Anti-SARS-CoV-2 Activity of Adamantanes In Vitro and in Animal Models of Infection. COVID, 2(11), 1551-1563. https://doi.org/10.3390/covid2110111