

Changes in Thermodynamic Parameters Induced by Pyrimidine Nucleic Bases Forming Complexes with Amino Acids and Peptides in a Buffer Solution at pH = 7.4



Abstract

1. Introduction
2. Description of Datasets
3. Results and Discussion
3.1. Characterization of Ionic State of Amino Acids and Peptides in Solution
3.2. Thermochemical Quantities of Amino Acids and Peptides in Water and Aqueous Buffers
- in the pure waterin an aqueous buffer mediumwhere ΔsolvHw is the enthalpy of the zwitterionic form solvation in water; Σ(α·ΔintH1) is the sum of the enthalpies of protonation–deprotonation of acid-active groups with the formation of other ionic forms of the amino acid/peptide, and α is the fraction of the forms, which depends on the pH of the solution; ΔintH2 is the enthalpy of amino acid/peptide–electrolyte ion interaction; and ΔsubH is the enthalpy of lattice destruction (sublimation enthalpy). If the measurements are carried out in the same buffer with a constant pH and the ionic strength of the solution, the contribution Σ(α·ΔintH1) remains constant for each amino acid/peptide.
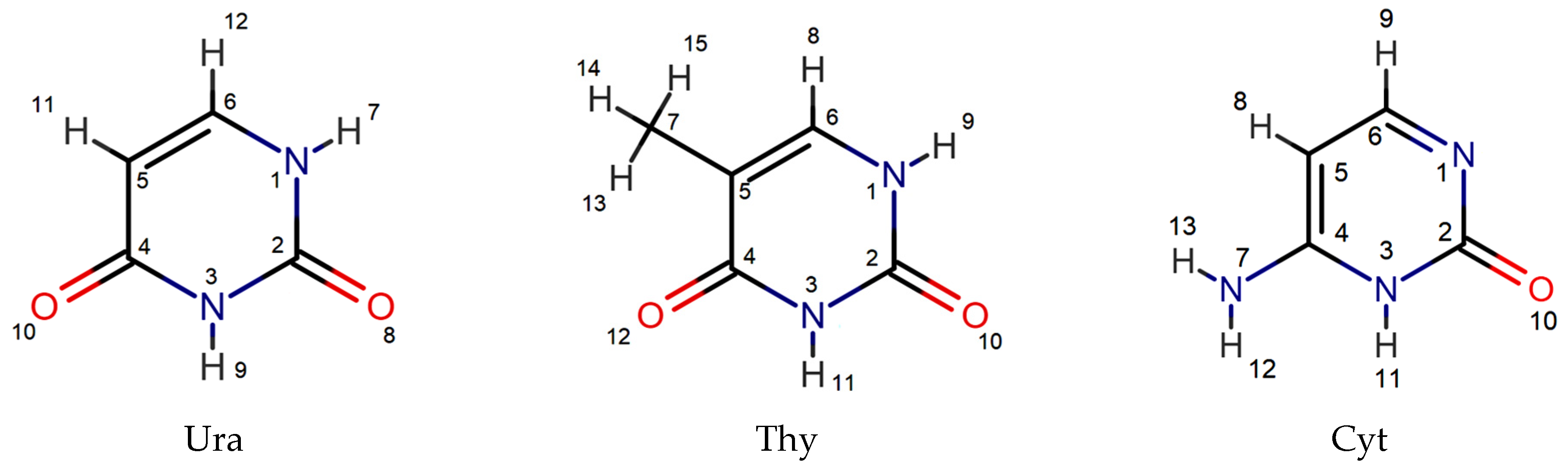
3.3. Structural Approach to Complex Formation of Amino Acids and Peptides with Nucleic Bases
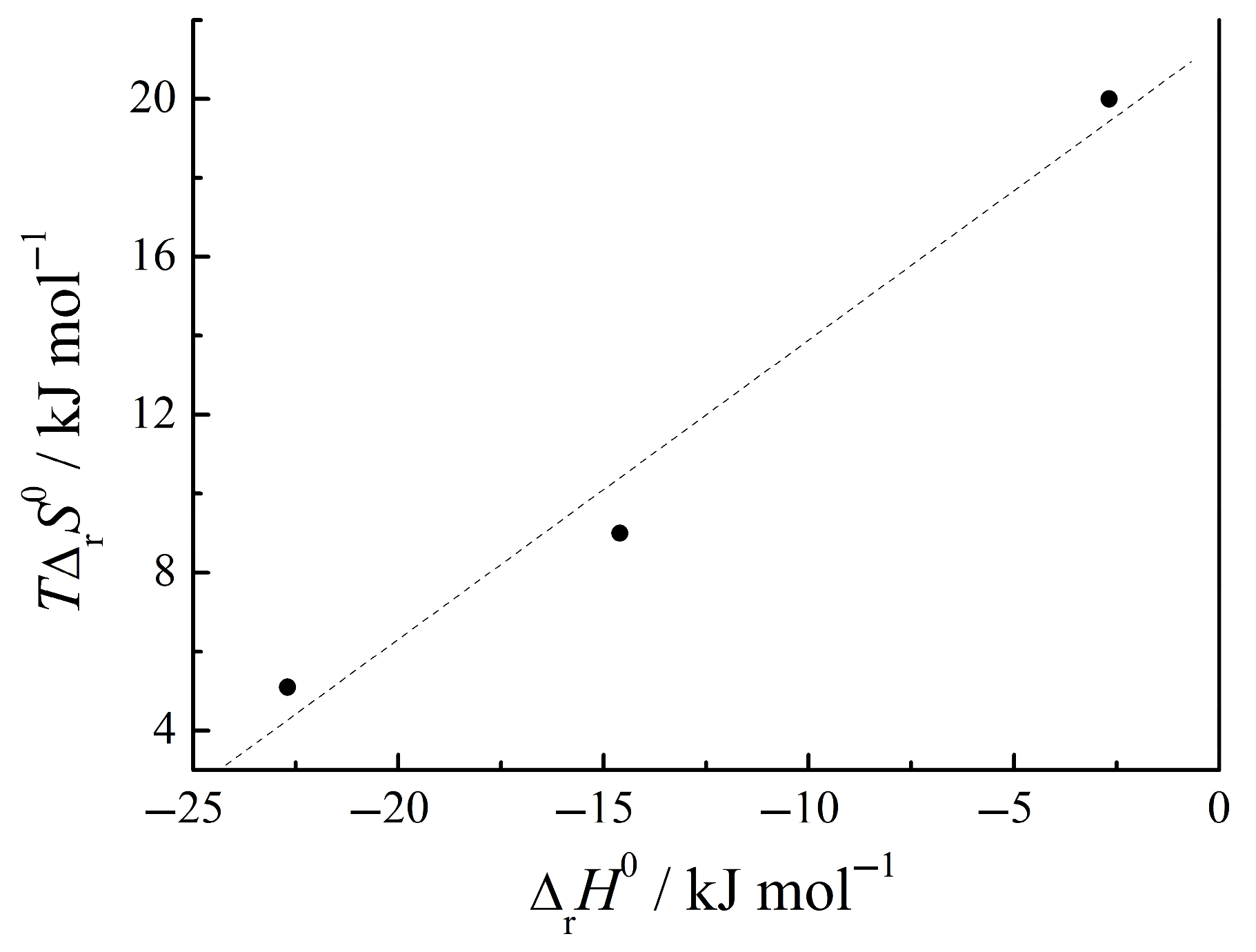
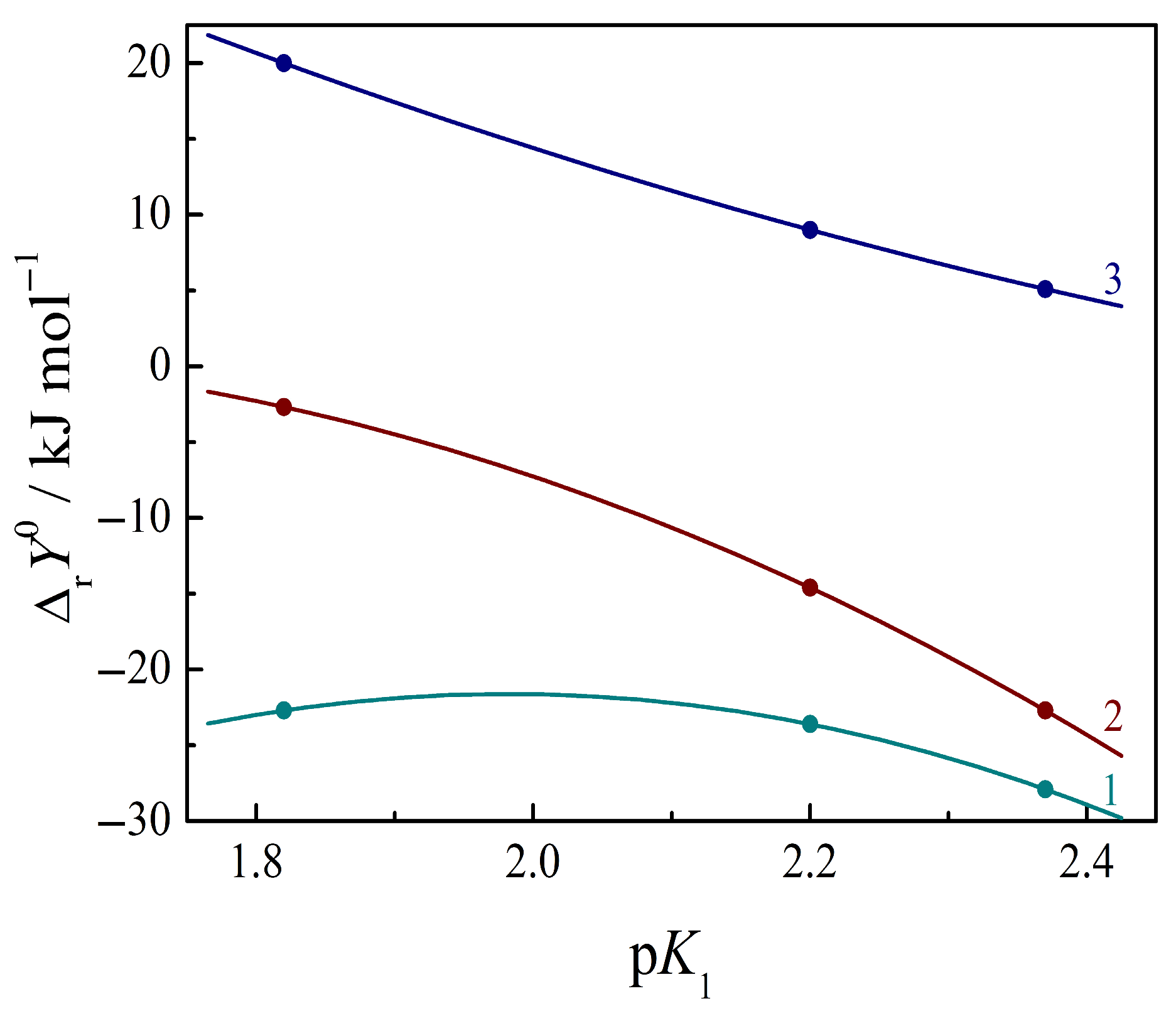
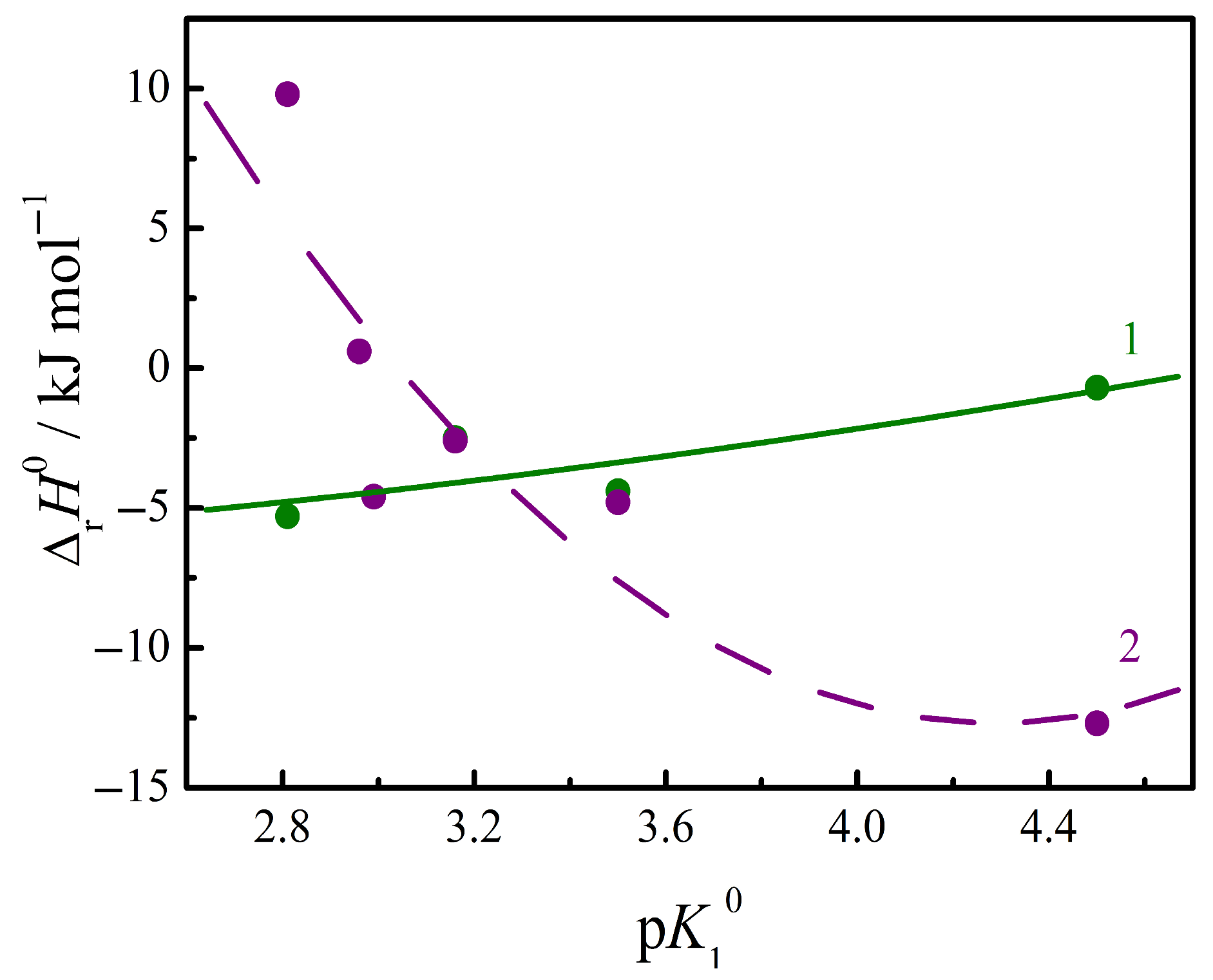
3.4. Thermodynamics of Complex Formation Between Amino Acids and Nucleic Bases in Aqueous Buffer Solutions
3.5. Thermodynamics of Complex Formation Between Peptides and Nucleic Bases in Aqueous Buffer Solutions
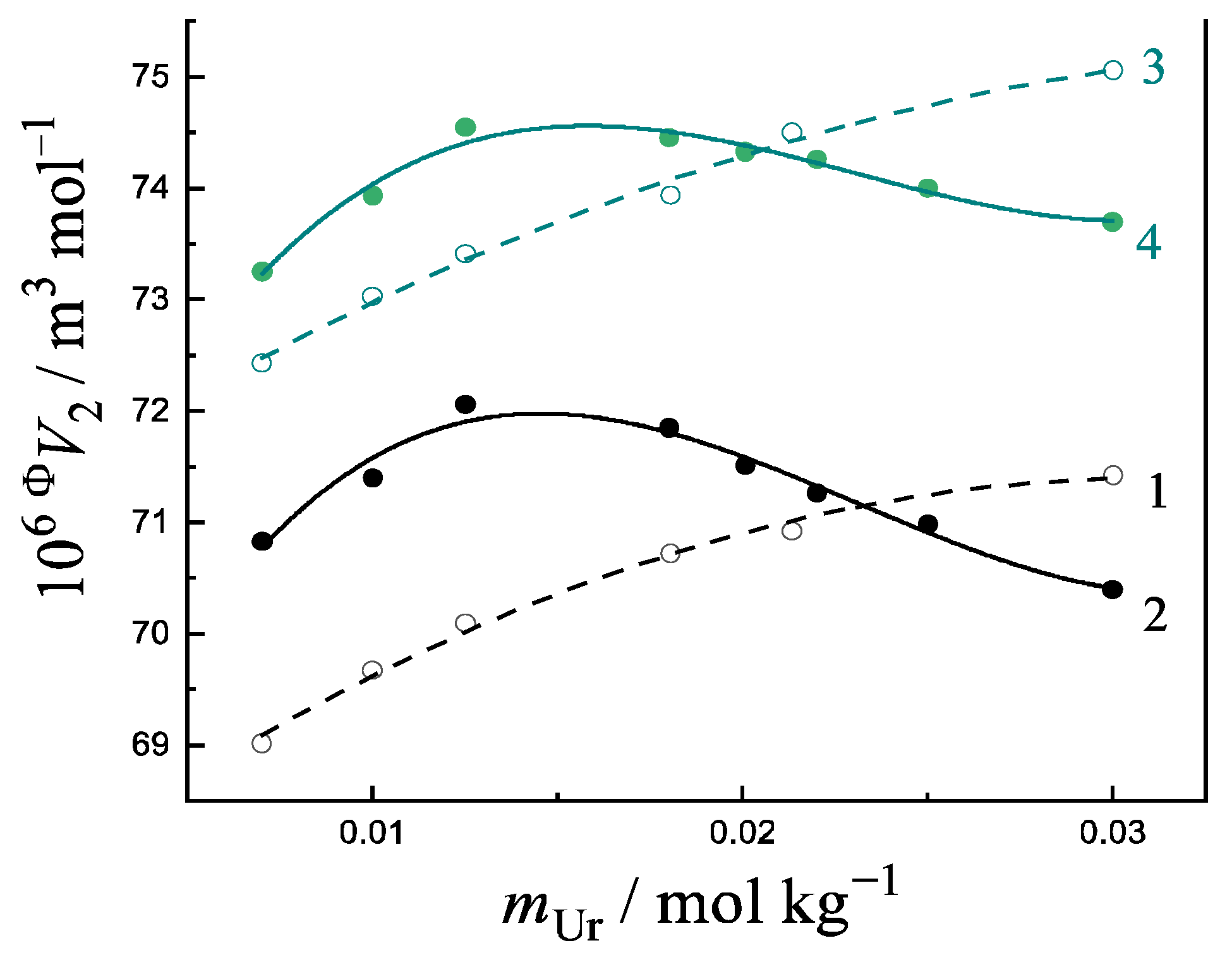
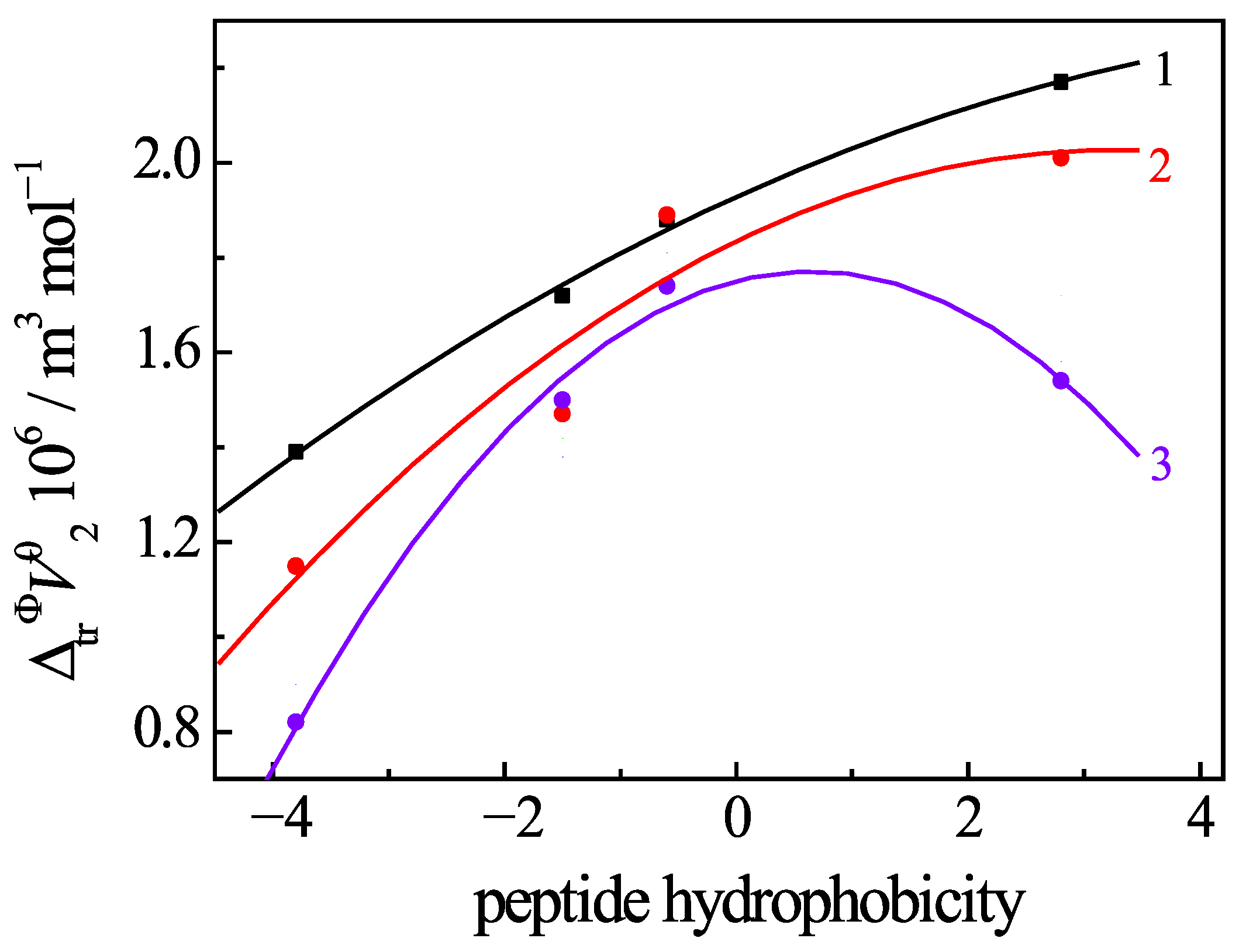
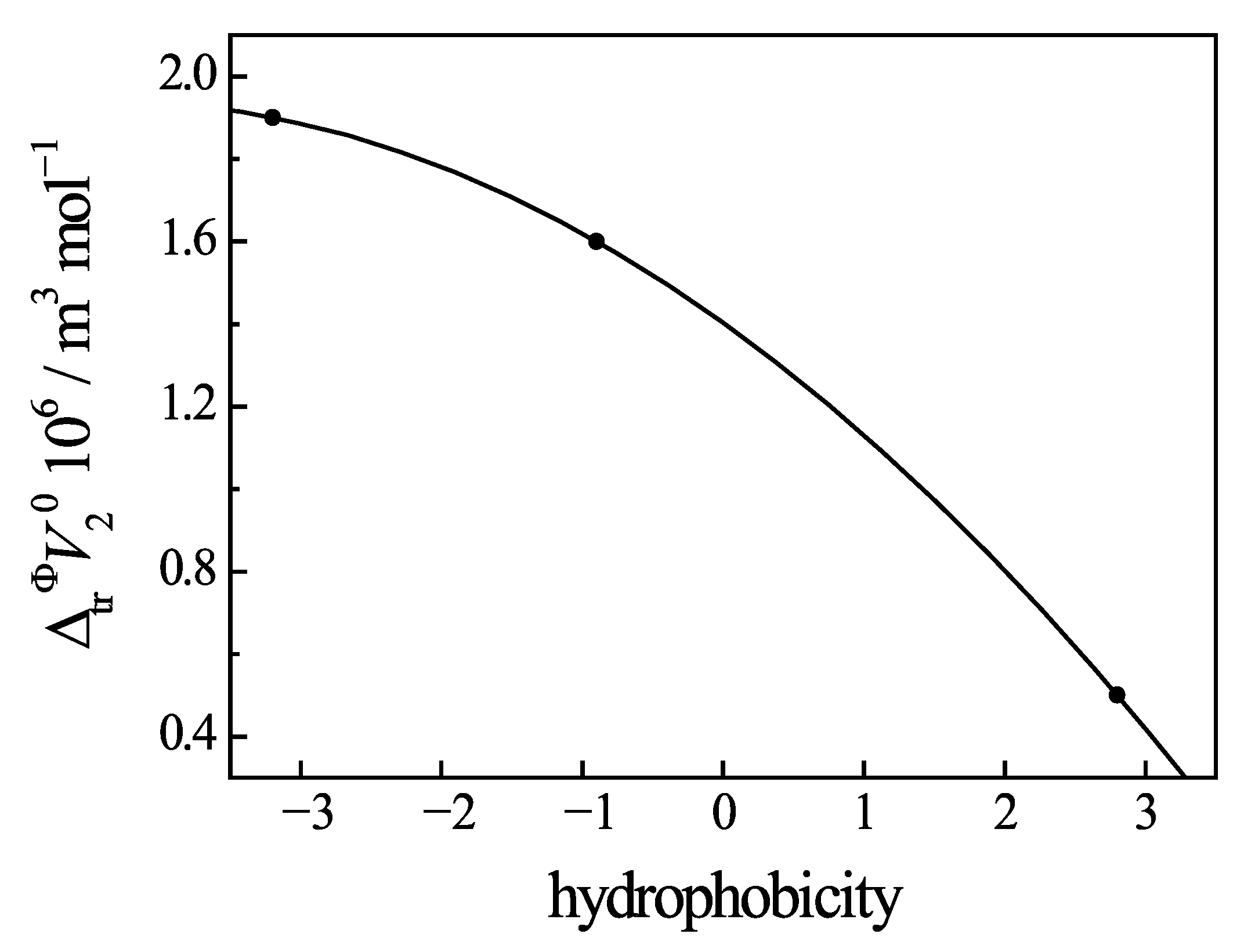
3.6. Change of Volumetric Properties of Uracil Under Addition of Amino Acids and Peptides
4. Conclusions
- -
- The manifestation of a maximum in the non-linear concentration dependences of the apparent molar volumes of Ura in a buffer saline with the additives of an amino acid/peptide shows the formation of ion–molecular complexes between a solute and co-solute (NB). The ratio of the reagent concentrations at the maximum point indicates the stoichiometry of the resulting complex.
- -
- The regular relationship between the volume changes, induced by the uracil–peptide interaction, and the hydrophobicity of the peptides evidences the major role the hydrophobic effect plays in the complexation process in solutions.
- -
- The effect of temperature on the volumetric properties of Ura in the solution is determined by the thermal disruption of the water structure and by the influence of hydrophilic and hydrophobic groups of the peptides that penetrate into the hydration shell of uracil. This is most clearly manifested in the values of the standard apparent molar transfer volumes and the limiting molar expansibilities.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barannikov, V.P.; Smirnov, V.I.; Mezhevoi, I.N.; Koltyshev, D.R. Patterns of interaction between peptides of various structures and pyrimidine nucleic bases in a buffered saline medium according to the analysis of thermodynamic parameters. J. Mol. Liq. 2024, 415, 126340. [Google Scholar] [CrossRef]
- Barannikov, V.P.; Smirnov, V.I.; Mezhevoi, I.N.; Koltyshev, D.R. Thermochemical study of the interaction of cytosine and uracil with peptides in a buffered saline: Complex formation with beta-endorphin 30-31 (human), L-glutathion (reduced) and α-L-alanyl-L-tyrosine. Int. J. Mol. Sci. 2023, 24, 9764. [Google Scholar] [CrossRef] [PubMed]
- Badelin, V.G.; Tyunina, E.Y.; Mezhevoi, I.N.; Tarasova, G.N. Thermodynamic characteristics of molecular interactions between L-tryptophan and nicotinic acid and uracyl in aqueous buffer solutions at 298.15 K. Russ. J. Phys. Chem. A 2015, 89, 2229–2233. [Google Scholar] [CrossRef]
- Tyunina, E.Y.; Badelin, V.G.; Mezhevoi, I.N.; Tarasova, G.N. Thermodynamics of aromatic amino acid interactions with heterocyclic ligands. J. Mol. Liq. 2015, 211, 494–497. [Google Scholar] [CrossRef]
- Tyunina, E.Y.; Badelin, V.G.; Mezhevoi, I.N. Observation of complex formation between L-histidine and heterocyclic compounds in water and aqueous buffer solution using calorimetric and spectroscopic methods. J. Mol. Liq. 2019, 278, 505–511. [Google Scholar] [CrossRef]
- Badelin, V.G.; Tarasova, G.N.; Tyunina, E.Y.; Bichkova, S.A. Investigation of interaction between L-histidine and heterocyclic substances in aqueous solutions by UV spectroscopy. Izv. Vyssh. Uchebn. Zaved. Khim. Khim. Tekhnol. 2018, 61, 11–18. [Google Scholar] [CrossRef]
- Tyunina, E.Y.; Badelin, V.G.; Mezhevoi, I.N. Volumetric and heat capacity studies of heterocyclic compounds in aqueous amino acids buffer solutions at 298.15 K. J. Chem. Thermodyn. 2019, 131, 40–48. [Google Scholar] [CrossRef]
- Tyunina, E.Y. Interaction between uracil and L-histidine in an aqueous buffer solution in the 288.15–313.15 K range of temperature. Russ. J. Phys. Chem. A 2021, 95, 2254–2262. [Google Scholar] [CrossRef]
- Barannikov, V.P.; Tyunina, E.Y.; Tarasova, G.N. Characterization of complexation of uracil with peptides through its volumetric properties in buffer saline solution at different temperatures. J. Chem. Thermodyn. 2025, 201, 107402. [Google Scholar] [CrossRef]
- Barannikov, V.P.; Tyunina, E.Y.; Tarasova, G.N. Changes in the volumetric properties of uracil in a buffered saline upon interaction with peptides: The effect of glycyl-L-tyrosine and glycyl-L-glutamic acid. J. Chem. Thermodyn. 2023, 185, 107113. [Google Scholar] [CrossRef]
- Kulikov, O.V.; Lapshev, P.V.; Parfenyuk, E.V. The role of weak and specific forces in the interaction of amino acids with cytosine, uracil and caffeine. Mendeleev Commun. 1995, 5, 72–74. [Google Scholar] [CrossRef]
- Smirnov, V.I.; Barannikov, V.P. Thermochemical characteristics of some glycylpeptides interaction with anionic micelles in a phosphate-buffered saline solution of sodium dodecyl sulfate. J. Chem. Thermodyn. 2022, 174, 106853. [Google Scholar] [CrossRef]
- Barannikov, V.P.; Kurbatova, M.S.; Mezhevoi, I.N. The influence of structure of isomolecular dipeptides of α-L-alanyl-α-Lalanine and β-alanyl-β-alanine on their behavior in aqueous micellar solution of SDS. Thermochim. Acta 2020, 689, 178647. [Google Scholar] [CrossRef]
- Barannikov, V.P.; Smirnov, V.I.; Kurbatova, M.S. The thermochemical behavior of glycyl-L-histidine and β-alanyl-L-histidine peptides in (SDS + phosphate-buffered saline) micellar solution at pH = 7.4. J. Mol. Liq. 2021, 331, 115766. [Google Scholar] [CrossRef]
- Lytkin, A.I.; Barannikov, V.P.; Badelin, V.G.; Krutova, O.N. Enthalpies of acid dissociation of L-carnosine in aqueous solution. J. Therm. Anal. Calorim. 2020, 139, 3683–3689. [Google Scholar] [CrossRef]
- Tyunina, E.Y.; Mezhevoi, I.N.; Stavnova, A.A. Molecular complexes of polar basic amino acids (L-lysine, L-histidine) with nicotinic acid in water and buffer solution: A thermodynamic aspects. J. Chem. Thermodyn. 2021, 161, 106552. [Google Scholar] [CrossRef]
- Tyunina, E.Y.; Smirnov, V.I.; Mezhevoi, I.N.; Barannikov, V.P. Thermochemical and volumetric studies of pyridine-3-carboxylic acid (vitamin B3) in a phosphate buffered saline solution with L-asparagine: Manifestation of molecular interactions. J. Mol. Liq. 2025, 423, 126902. [Google Scholar] [CrossRef]
- The Cambridge Crystallographic Data Centre, Leibniz Institute for Information Infrastructure. Available online: https://www.ccdc.cam.ac.uk/structures/Search?Ccdcid=1105666&DatabaseToSearch=Published (accessed on 1 November 2022).
- Badelin, V.G.; Barannikov, V.P.; Tarasova, G.N.; Chernyavskaya, N.V.; Katrovtseva, A.V.; Fam, T.L. Thermodynamical characteristics of acid-base equilibria in GlycylGlycylGlycine aqueous solutions at 298 K. Russ. J. Phys. Chem. A 2012, 86, 40–44. [Google Scholar] [CrossRef]
- Gorboletova, G.G.; Gridchin, S.N.; Sazonova, E.S. Heat effects of acid-base interactions in aqueous solutions of glycylglycine. Russ. J. Phys. Chem. A 2005, 79, 1222–1226. [Google Scholar]
- Lytkin, A.I.; Chernikov, V.V.; Krutova, O.N.; Damrina, K.V.; Skvortsov, I.A. Thermochemical study of acid—Base reactions in a glycyl-tyrosine aqueous solution. Russ. J. Phys. Chem. A 2016, 90, 735–738. [Google Scholar] [CrossRef]
- Lytkin, A.I.; Krutova, O.N.; Golubev, A.A.; Badelin, V.G.; Tyunina, E.Y. Effect of the nature of background electrolyte on the thermodynamic characteristics of the acid-base reactions of L-asparagine in aqueous solutions. Russ. J. Phys. Chem. A 2019, 93, 1729–1732. [Google Scholar] [CrossRef]
- Piekarski, H.; Nowicka, B. Calorimetric studies of interactions of some peptides with electrolytes, urea and ethanol in water at 298.15 K. J. Therm. Anal. Calorim. 2010, 102, 31–36. [Google Scholar] [CrossRef]
- Vasil’ev, V.P.; Borodin, V.A.; Kozlovsky, E.V. Application of PC in Chemical Analytical Calculations; High School: Moscow, Russia, 1993. (In Russian) [Google Scholar]
- Barannikov, V.P.; Badelin, V.G.; Venediktov, E.V.; Mezhevoi, I.N.; Guseinov, S.S. Thermodynamical characteristics of the reaction of pyridoxal-5′-phosphate with L-amino acids in aqueous buffer solution. Russ. J. Phys. Chem. A 2011, 85, 16–20. [Google Scholar] [CrossRef]
- Hunter, K.C.; Millen, A.L.; Wetmore, S.D. Effects of hydrogen-bonding and stacking interactions with amino acids on the acidity of uracil. J. Phys. Chem. B 2007, 111, 1858–1871. [Google Scholar] [CrossRef] [PubMed]
- Gutowski, M.; Dąbkowska, I.; Rak, J.; Xu, S.; Nilles, J.M.; Radisic, D.; Bowen, K.H., Jr. Barrier-free intermolecular proton transfer in the uracil-glycine complex induced by excess electron attachment. Eur. Phys. J. D 2002, 20, 431. [Google Scholar] [CrossRef]
- Dąbkowska, I.; Rak, J.; Gutowski, M.; Nilles, J.M.; Stokes, S.T.; Bowen, K.H., Jr. Barrier-free intermolecular proton transfer induced by excess electron attachment to the complex of alanine with uracil. J. Chem. Phys. 2004, 120, 6064–6071. [Google Scholar] [CrossRef] [PubMed]
- Dąbkowska, I.; Rak, J.; Gutowski, M.; Nilles, J.M.; Stokes, S.T.; Radisic, D.; Bowen, K.H., Jr. Barrier-free proton transfer in anionic complex of thymine with glycine. Phys. Chem. Chem. Phys. 2004, 6, 4351–4357. [Google Scholar] [CrossRef]
- Alkorta, I.; Elguero, J. Interaction of protein backbone with nucleic acid bases. J. Phys. Chem. B 2003, 107, 5306–5310. [Google Scholar] [CrossRef]
- Kurbatova, M.S.; Barannikov, V.P. Complexes of ionized dipeptides of glycyl-L-glutamic acid and alanyl-L-tyrosine with nucleic bases of the pyrimidine series. J. Struct. Chem. 2023, 64, 2213–2225. [Google Scholar] [CrossRef]
- Kurbatova, M.S.; Barannikov, V.P. DFT-calculations and RDG analysis (topology) of complexes between glutathione and nucleic bases of pyrimidine series pyrimidine series. J. Mol. Model. 2025, 3, 98. [Google Scholar] [CrossRef]
- Rutledge, L.R.; Campbell-Verduyn, L.S.; Wetmore, S.D. Characterization of the stacking interactions between DNA or RNA nucleobases and the aromatic amino acids. Chem. Phys. Lett. 2007, 444, 167–175. [Google Scholar] [CrossRef]
- Saito, I.; Sugiyama, H.; Matsuura, T.; Fukuyama, K. Stacking interaction between tryptophan and uracil in a synthetic model compound. Tetrahedron Lett. 1985, 26, 4467–4470. [Google Scholar] [CrossRef]
- Troitiňo, D.; Bailey, L.; Peral, F. Formation of hydrogen-bonded dimers of uracil and amino acids: Cysteine, lysine and phenylalanine. A density functional theory study. J. Mol. Struct. Theochem. 2006, 767, 131–138. [Google Scholar] [CrossRef]
- Lapschov, P.V.; Kulikov, O.V. Thermodynamics and mechanism of interaction of uracil with amino acids in water. Russ. Chem. Bull. 1977, 46, 892–894. [Google Scholar] [CrossRef]
- Nagai, H.; Kuwabara, K.; Carta, G. Temperature Dependence of the Dissociation Constants of Several Amino Acids. J. Chem. Eng. Data 2008, 53, 619–627. [Google Scholar] [CrossRef]
- Solomonov, B.N.; Yagofarov, M.I. The relationship between the Gibbs energies and enthalpies of hydrogen bonding and charge-transfer complex formation in non-electrolytes solutions. Is it a rule? J. Mol. Liq. 2025, 424, 127053. [Google Scholar] [CrossRef]
- Palekar, D.; Shiue, M.; Lien, E.J. Correlation of Physicochemical Parameters to the Hydrophobic Contribution Constants of Amino Acids Residues in Small Peptides. Pharm. Res. 1996, 13, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Sturtevant, J.M. Heat capacity and entropy changes in processes involving proteins. Prog. Natl. Acad. Sci. USA 1977, 74, 2236–2240. [Google Scholar] [CrossRef] [PubMed]
- Hélène, C.; Lancelot, G. Interactions between functional groups in protein-nucleic acid associations. Prog. Biophys. Mol. Biol. 1982, 39, 1–68. [Google Scholar] [CrossRef] [PubMed]
- Banipal, T.S.; Kaur, N.; Banipal, P.K. Volumetric studies on nucleic acids bases and nucleosides in aqueous guanidine hydrochloride solutions at T= (288.15 to 318.15) K and at atmospheric pressure. J. Chem. Thermodyn. 2015, 82, 12–24. [Google Scholar] [CrossRef]
- Kaur, N.; Banipal, P.K.; Banipal, T.S. Physico-chemical effects of caffeine on aqueous solutions of pyrimidine based model compounds of nucleic acids. J. Mol. Liq. 2016, 221, 721–732. [Google Scholar] [CrossRef]
- Zielenkiewicz, W.; Poznański, J.; Zielenkiewicz, A. Partial molar volumes of aqueous solutions of some halo and amino derivatives of uracil. J. Solut. Chem. 2000, 29, 757–769. [Google Scholar] [CrossRef]
- Kishore, N.; Ahluwalia, J.C. Partial molar heat capacities and volumes of transfer of nucleic acid bases, nucleosides and nucleotides from water to aqueous solutions of sodium and calcium chloride at 25 °C. J. Solut. Chem. 1990, 19, 51–64. [Google Scholar] [CrossRef]
- Patel, S.G.; Kishore, N. Thermodynamics of nucleic acid bases and nucleosides in water from 25 to 55 °C. J. Solut. Chem. 1995, 24, 25–38. [Google Scholar] [CrossRef]
- Fucaloro, A.F.; Dewey, K.; Fan, G.; Imuta, K.; Jensen, D.; Muranaka, M. Partial molar volumes of uracil, thymine, adenine in water and of adenine in aqueous solutions of uracil and thymine. J. Solut. Chem. 2008, 37, 1289–1304. [Google Scholar] [CrossRef]
- Tyunina, E.Y.; Mezhevoi, I.N.; Dunaeva, V.V. Thermodynamic properties of crystalline L-carnosine and its aqueous solutions. J. Chem. Thermodyn. 2020, 150, 106206. [Google Scholar] [CrossRef]
- Rani, R.; Rajput, S.; Sharma, K.; Baboria, V. Volumetric and viscometric properties od amino acids in aqueous solutions of various drugs at different temperatures: A review. Mol. Phys. 2022, 120, e19920259. [Google Scholar] [CrossRef]
- Barannikov, V.P.; Tyunina, E.Y. Regularities of changes in thermodynamic parameters induced by the complexes formation of uracil with some aromatic amino acids in a buffer solution at pH 7.4. Izv. Vyssh. Uchebn. Zaved. Khim. Khim. Tekhnol. ChemChemTech. 2022, 65, 38–46. [Google Scholar] [CrossRef]
- Rajput, P.; Richu; Sharma, T.; Kumar, A. Temperature dependent physicochemical investigations of some nucleic acid bases (uracil, thymine and adenine) in aqueous inositol solutions. J. Mol. Liq. 2021, 326, 115210. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, B. Analyzing solvent reorganization and hydrophobicity. Methods Enzymol. 1995, 259, 555–576. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.J.; Chaudhry, M.A. Thermodynamic study of the three pharmacologically significant drugs: Density, viscosity, and refractive index measurements at different temperatures. J. Chem. Thermodyn. 2009, 41, 221–226. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, J.; Zheng, H.; Liu, D. Volumetric properties of some α-amino acids in aqueous guanidine hydrochloride at 5, 15, 25, and 35 °C. J. Solut. Chem. 1998, 27, 473–483. [Google Scholar] [CrossRef]
- Krumgalz, B.S. Ionic limiting partial molar volumes in various solvents. J. Chem. Soc. Faraday Trans. I 1980, 76, 1887–1904. [Google Scholar] [CrossRef]
- Gurney, R.W. Ionic Processes in Solution; McGraw Hill: New York, NY, USA, 1953. [Google Scholar]
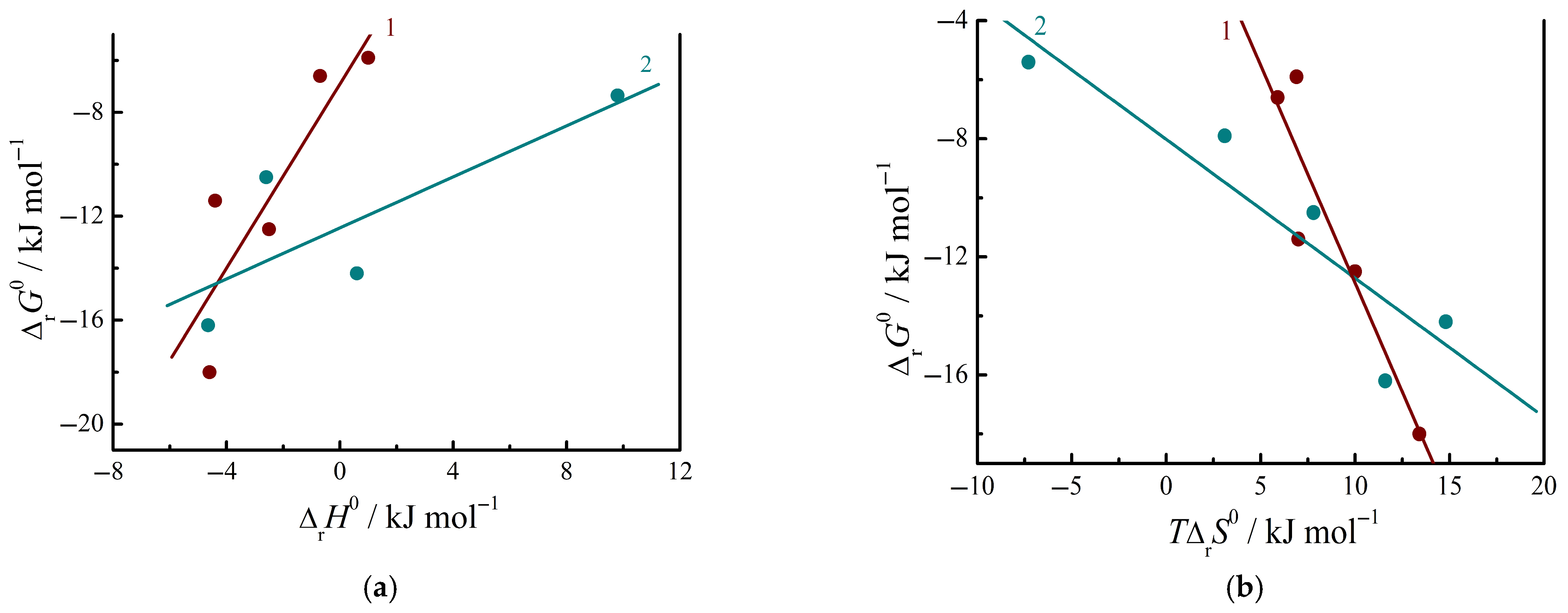




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2+H3L | +H3L | +H2L | 2+HL− | +HL− | +HL2− | L− | L2− | Ref. | |
|---|---|---|---|---|---|---|---|---|---|
| His | 0 | - | - | 0.06 | 0.94 | - | 0.02 | - | [16] |
| Trp | - | - | 0 | - | 0.99 | - | 0.01 | - | [3] |
| Phe | - | - | 0 | - | 0.98 | - | 0.02 | - | [4] |
| AlaAla | - | - | 0 | - | 0.91 | - | 0.09 | - | [13] |
| GlyGly | - | - | 0 | - | 0.89 | - | 0.11 | - | [1] |
| GlyTyr | - | - | 0 | - | 0.84 | - | 0.16 | 0 | [12] |
| AlaTyr | - | - | 0 | - | 0.77 | - | 0.23 | 0 | [2] |
| GlyPhe | - | - | 0 | - | 0.83 | - | 0.17 | - | [1] |
| AlaGln | - | - | 0 | - | 0.88 | - | 0.12 | - | [1] |
| GlyGlu | - | 0 | - | - | 0.01 | 0.86 | - | 0.13 | [2] |
| GlyHis | 0 | - | 0 | 0.15 | 0.67 | - | 0.18 | - | [14] |
| β-AlaHis | 0 | - | - | 0.19 | 0.80 | - | 0.01 | - | [15] |
| γ-GluCysGly | - | 0 | - | - | 0.95 | - | 0.05 | [2] | |
| GlyGlyGly | - | - | 0 | - | 0.81 | - | 0.19 | - | [1] |
| log Kr | ΔrG0 (kJ·mol−1) | ΔrH0 (kJ·mol−1) | TΔrS0 (kJ·mol−1) | Ref. | |
|---|---|---|---|---|---|
| His + 2Ura | 3.98 | −22.70 | −2.68 | 20.0 | [5] |
| Trp + 2Ura | 4.89 | −27.90 | −22.70 | 5.10 | [3] |
| Phe + 2Ura | 4.13 | −23.60 | −14.60 | 9.00 | [4] |
| GlyGly + Ura | 2.19 | −12.51 | −2.50 | 10.00 | [1] |
| GlyGly + Cyt | 1.84 | −10.47 | −2.63 | 7.84 | [1] |
| GlyPhe + Ura | 3.15 | −18.00 | −4.61 | 13.39 | [1] |
| GlyPhe + Cyt | 2.84 | −16.22 | −4.65 | 11.60 | [1] |
| AlaGln + Cyt | 2.50 | −14.20 | 0.62 | 14.80 | [1] |
| AlaGln + Thy | 2.27 | −13.01 | 0.79 | 13.80 | [1] |
| AlaTyr + Ura | 1.45 | −8.30 | −5.30 | 3.00 | [2] |
| AlaTyr + Cyt | 1.29 | −7.35 | 9.80 | 17.15 | [2] |
| AlaAla + Ura | 1.03 | −5.90 | 1.00 | 6.90 | [2] |
| GlyGlu + Ura | 1.16 | −6.64 | −0.70 | 5.90 | [2] |
| GlyGlu + Cyt | 0.95 | −5.40 | −12.70 | −7.30 | [2] |
| GluCysGly + Ura | 2.01 | −11.40 | −4.40 | 7.00 | [2] |
| GluCysGly + Thy | 1.24 | −7.00 | −6.70 | 0.30 | [1] |
| GluCysGly + Cyt | 1.39 | −7.89 | −4.79 | 3.10 | [2] |
| Medium | 288.15 K | 293.15 K | 298.15 K | 303.15 K | 308.15 K | 313.15 K | Ref. |
|---|---|---|---|---|---|---|---|
| ΦV°2 × 106/(m3·mol−1) | |||||||
| water | 71.86 | 74.10 | [44] | ||||
| 70.95 | 71.88 | 73.28 | 74.47 | 75.61 | [51] | ||
| 71.23 | 72.30 | 73.72 | [42] | ||||
| buffered saline | 67.60 | 68.40 | 69.30 | 70.10 | 71.10 | [10] | |
| ΔtrV°2 × 106/(m3·mol−1) | |||||||
| phosphate buffer + His | 1.7 | 1.9 | 2.4 | 4.1 | 6.2 | [8] | |
| phosphate buffer + Phe | 0.9 | 0.5 | 0.9 | 2.0 | [7] | ||
| phosphate buffer + Trp | 1.6 | [7] | |||||
| buffered saline + GlyGly | 1.9 | 1.9 | 1.6 | 1.9 | 1.7 | [9] | |
| buffered saline + GlyPhe | 2.2 | 2.0 | 1.7 | 1.6 | 1.5 | [9] | |
| buffered saline + GlyTyr | 1.7 | 1.5 | 1.4 | 1.4 | 1.5 | [10] | |
| buffered saline + GlyGlu | 1.4 | 1.2 | 0.9 | 0.9 | 0.8 | [10] | |
| ΦE° × 106/(m3·mol−1·K−1) | |||||
|---|---|---|---|---|---|
| 288.15 K | 298.15 K | 303.15 K | 308.15 K | 313.15 K | |
| Ura | 0.0581 | 0.1249 | 0.1583 | 0.1917 | 0.2251 |
| Ura + GlyPhe | 0.1410 | 0.2130 | 0.2490 | 0.2850 | 0.3210 |
| Ura + GlyGly | 0.0864 | 0.1619 | 0.1997 | 0.2375 | 0.2754 |
| Ura + GlyTyr | 0.0197 | 0.1101 | 0.1553 | 0.2005 | 0.2457 |
| Ura + GlyGlu | 0.0213 | 0.0991 | 0.1379 | 0.1768 | 0.2157 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyunina, E.Y.; Barannikov, V.P.; Mezhevoi, I.N. Changes in Thermodynamic Parameters Induced by Pyrimidine Nucleic Bases Forming Complexes with Amino Acids and Peptides in a Buffer Solution at pH = 7.4. Liquids 2025, 5, 19. https://doi.org/10.3390/liquids5030019
Tyunina EY, Barannikov VP, Mezhevoi IN. Changes in Thermodynamic Parameters Induced by Pyrimidine Nucleic Bases Forming Complexes with Amino Acids and Peptides in a Buffer Solution at pH = 7.4. Liquids. 2025; 5(3):19. https://doi.org/10.3390/liquids5030019
Chicago/Turabian StyleTyunina, Elena Yu., Vladimir P. Barannikov, and Igor N. Mezhevoi. 2025. "Changes in Thermodynamic Parameters Induced by Pyrimidine Nucleic Bases Forming Complexes with Amino Acids and Peptides in a Buffer Solution at pH = 7.4" Liquids 5, no. 3: 19. https://doi.org/10.3390/liquids5030019
APA StyleTyunina, E. Y., Barannikov, V. P., & Mezhevoi, I. N. (2025). Changes in Thermodynamic Parameters Induced by Pyrimidine Nucleic Bases Forming Complexes with Amino Acids and Peptides in a Buffer Solution at pH = 7.4. Liquids, 5(3), 19. https://doi.org/10.3390/liquids5030019